
Preliminary Study on the Formation Mechanism of Malformed Sweet Cherry (Prunus avium L.) Fruits in Southern China Using Transcriptome and Metabolome Data

Abstract
:1. Introduce
2. Results
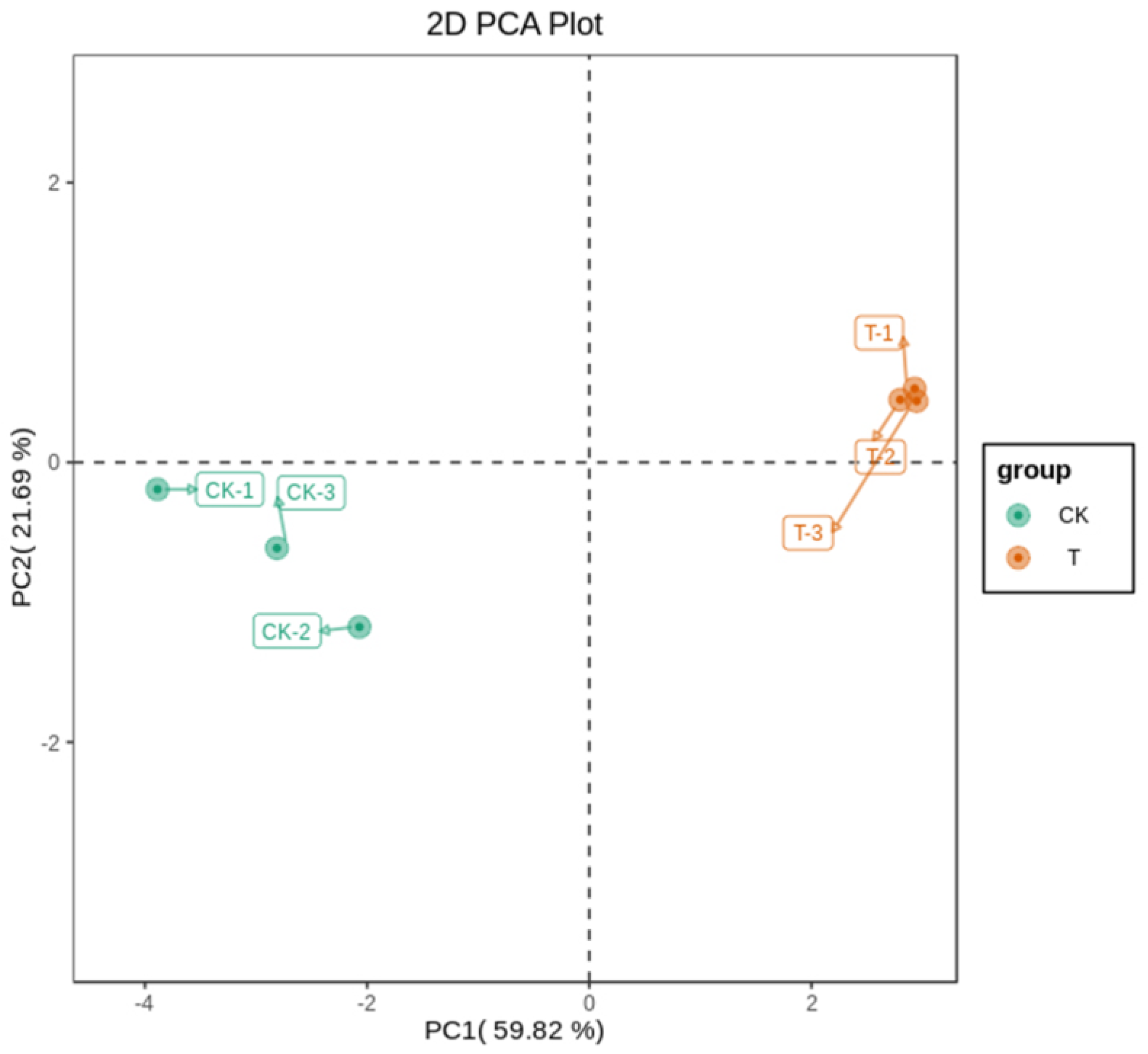
2.1. Metabolomics Analysis of Sweet Cherry
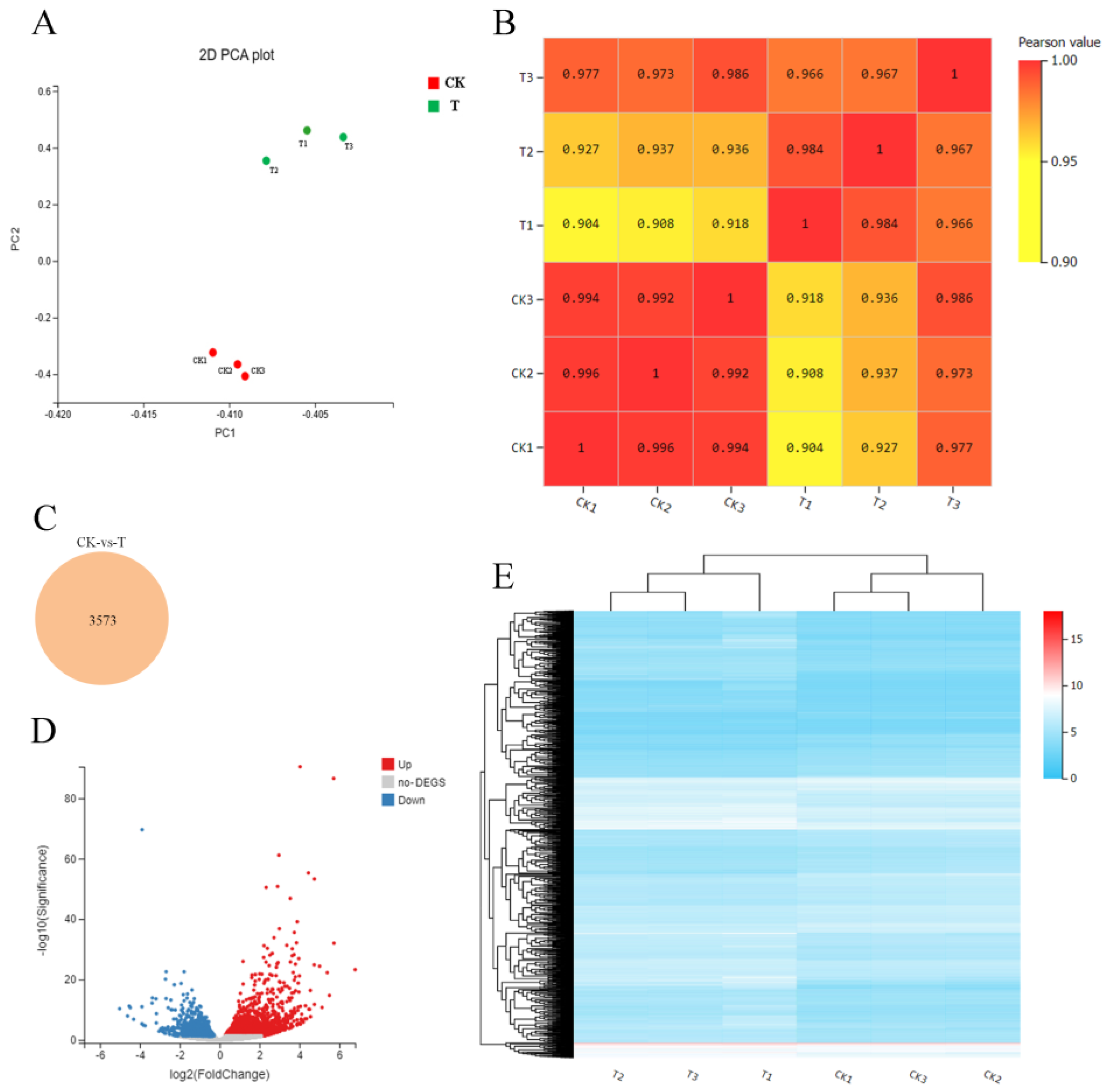
2.2. Transcriptome Analysis of Sweet Cherry Fruit
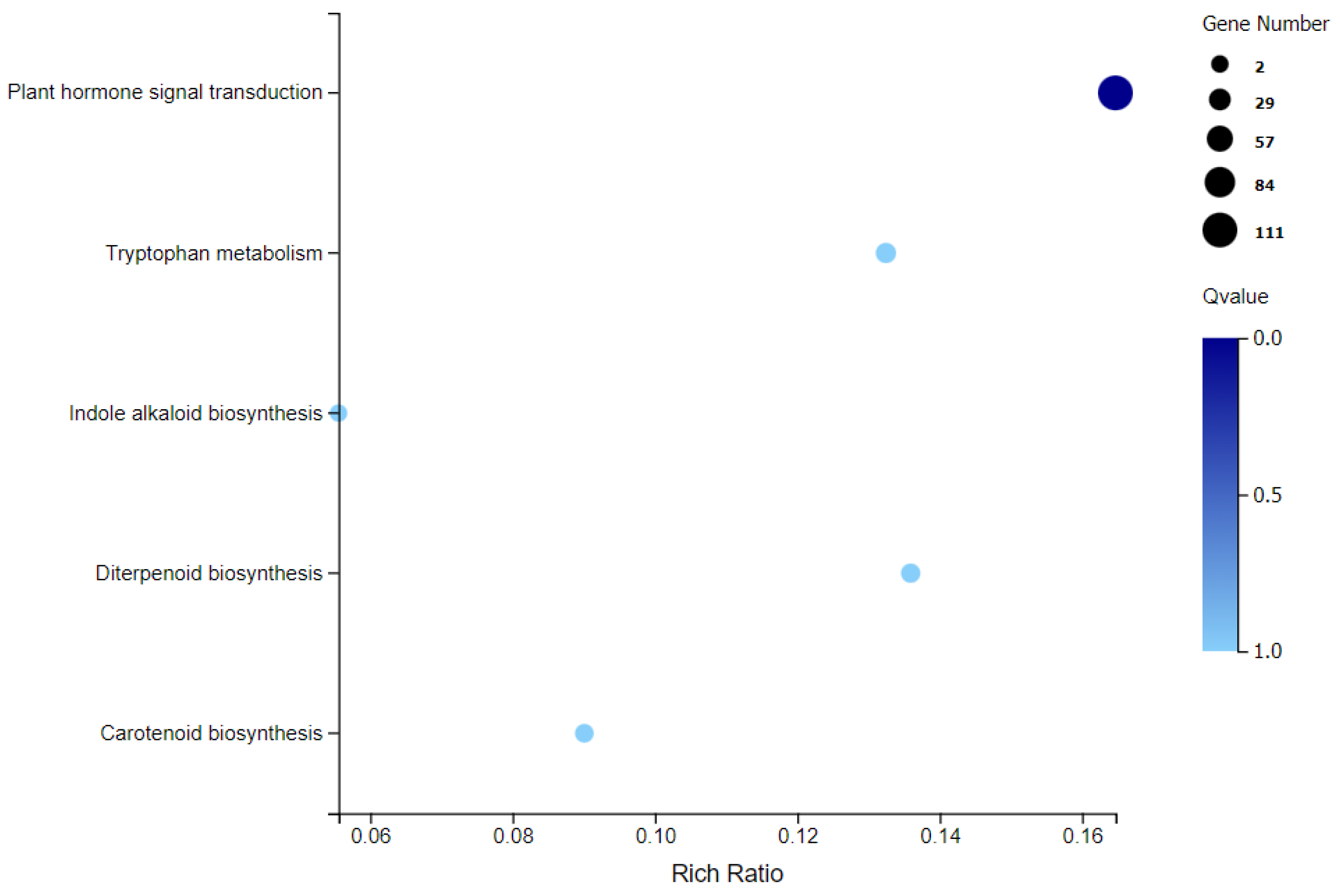
2.3. Combined Metabolome and Transcriptome Analysis
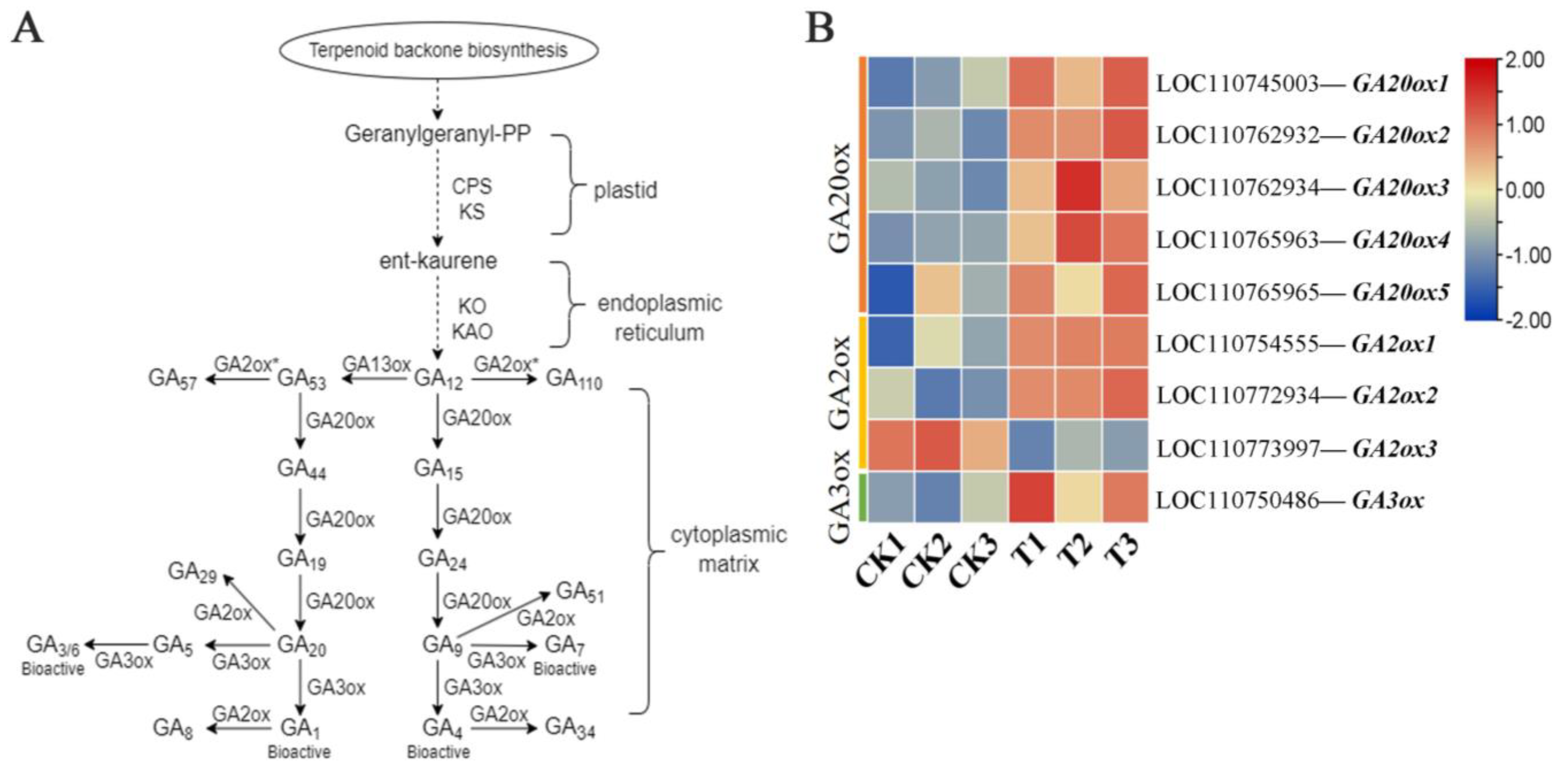
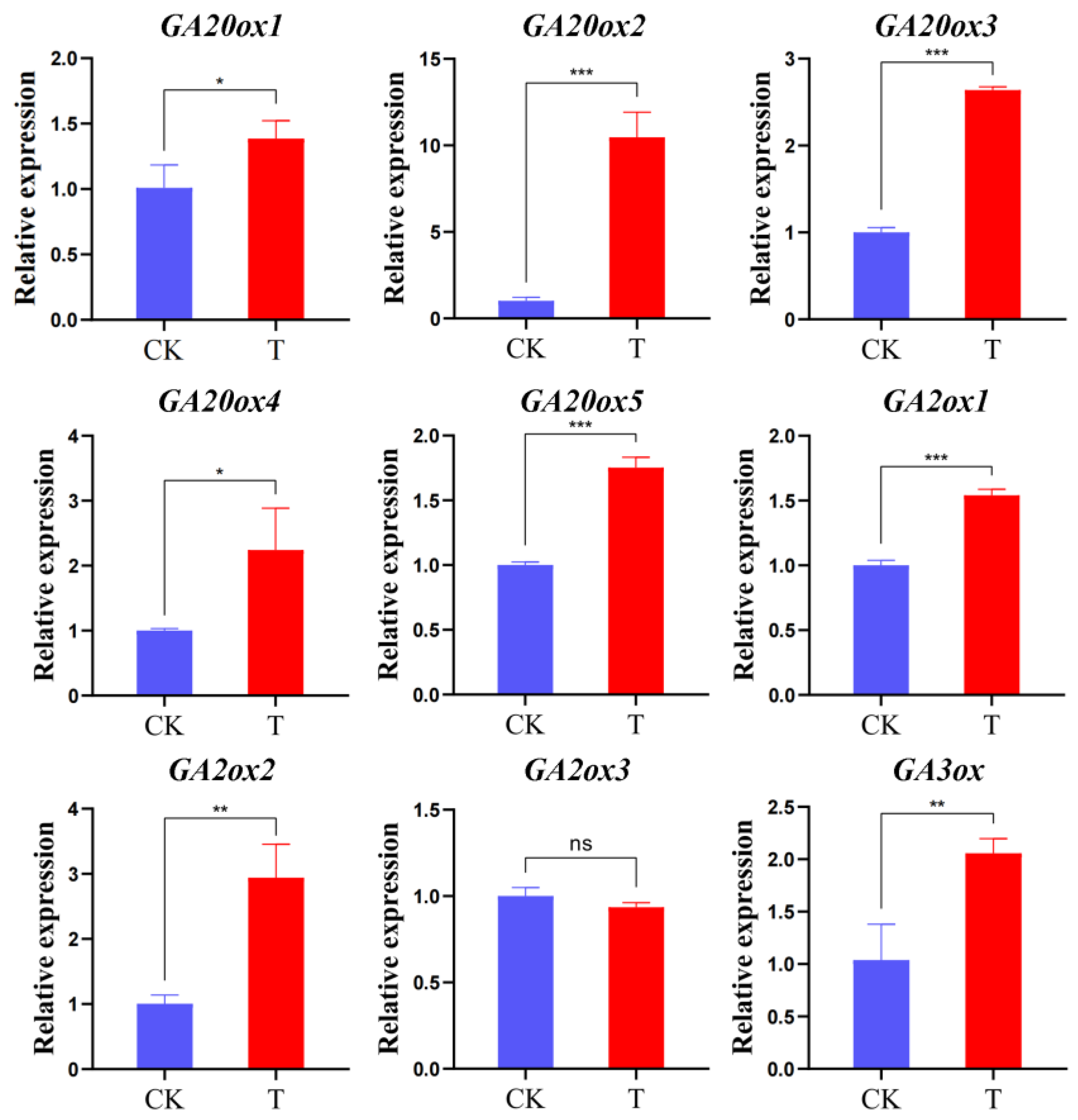
2.4. Expression of Gibberellin-Related Synthase Gene
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Metabolite Extraction and Profiling
4.3. RNA Extraction and RNA-Sequencing
4.4. Combined Metabolome and Transcriptome Analysis
4.5. Different Expression Gene (DEG), KEGG, and Gene Ontology (GO) Enrichment Analysis
4.6. Quantitative Real-Time Reverse Transcription PCR (qRT-PCR) Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Papapetros, S.; Louppis, A.; Kosma, I.; Kontakos, S.; Badeka, A.; Kontominas, M.G. Characterization and differentiation of botanical and geographical origin of selected popular sweet cherry cultivars grown in Greece. J. Food Compos. Anal. 2018, 72, 48–56. [Google Scholar] [CrossRef]
- Faienza, M.F.; Corbo, F.; Carocci, A.; Catalano, A.; Clodoveo, M.L.; Grano, M.; Wang, D.Q.; Amato, G.; Muraglia, M.; Franchini, C.; et al. Novel insights in health-promoting properties of sweet cherries. J. Funct. Foods 2020, 69, 103945. [Google Scholar] [CrossRef] [PubMed]
- Askarieh, A.; Suleiman, S.; Tawakalna, M. Sweet cherry (Prunus avium L.) fruit drop reduction by plant growth regulators (Naphthalene acetic acid NAA and gibberellic acid GA3). Am. J. Plant Sci. 2021, 12, 1338–1346. [Google Scholar] [CrossRef]
- Chen, C.Q.; Chen, H.X.; Chen, Y.F.; Yang, W.L.; Li, M.Y.; Sun, B.; Song, H.Y.; Tang, W.J.; Zhang, Y.; Gong, R.G. Joint metabolome and transcriptome analysis of the effects of exogenous GA3 on endogenous hormones in sweet cherry and mining of potential regulatory genes. Front. Plant Sci. 2022, 13, 1041068. [Google Scholar] [CrossRef] [PubMed]
- Mesejo, C.; Yuste, R.; Reig, C.; Martínez-Fuentes, A.; Iglesias, D.J.; Muñoz-Fambuena, N.; Bermejo, A.; Germanà, M.A.; Primo-Millo, E.; Agustí, M. Gibberellin reactivates and maintains ovary-wall cell division causing fruit set in parthenocarpic Citrus species. Plant Sci. 2016, 247, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Liang, J.J.; Chang, X.; Yang, H.T.; Li, T.Z.; Hu, J.F. Enhanced vacuolar invertase activity and capability for carbohydrate import in GA-treated inflorescence correlate with increased fruit set in grapevine. Tree Genet. Genomes 2017, 13, 21–33. [Google Scholar] [CrossRef]
- Jiang, S.; Luo, J.; Xu, F.J.; Zhang, X.Y. Transcriptome Analysis Reveals Candidate Genes Involved in Gibberellin-Induced Fruit Setting in Triploid Loquat (Eriobotrya japonica). Front. Plant Sci. 2016, 7, 1924. [Google Scholar] [CrossRef]
- Wen, B.B.; Song, W.L.; Sun, M.Y.; Chen, M.; Mu, Q.; Zhang, X.H.; Wu, Q.J.; Chen, X.D.; Gao, D.S.; Wu, H.Y. Identification and characterization of cherry (Cerasus pseudocerasus G. Don) genes responding to parthenocarpy induced by GA3 through transcriptome analysis. BMC Genet. 2019, 20, 65. [Google Scholar] [CrossRef]
- Watanabe, M.; Segawa, H.; Murakami, M.; Sagawa, S.; Komori, S. Effects of plant growth regulators on fruit set and fruit shape of parthenocarpic apple fruits. J. Jpn. Soc. Hortic. Sci. 2008, 77, 350–357. [Google Scholar] [CrossRef]
- Liu, C.S.; Xiao, P.S.; Jiang, F.; Wang, S.Y.; Liu, Z.; Song, G.Z.; Li, W.; Lv, T.X.; Li, J.; Wang, D.M.; et al. Exogenous gibberellin treatment improves fruit quality in self-pollinated apple. Plant Physiol. Biochem. 2022, 174, 11–21. [Google Scholar] [CrossRef]
- Du, Y.X.; Li, J.X.; Dong, J.M.; Hu, C.X.; Li, D.P.; Tan, Q.L.; Zhang, J.Z.; Li, J.; Zhou, X.Y.; Zhu, C.H.; et al. Effects of Different Regulating Measures on the Floral and Nutritional Physiology of Lemon. Agronomy 2022, 12, 2381. [Google Scholar] [CrossRef]
- Elfving, D.C.; Whiting, M.D.; Lenahan, O.M. Gibberellic acid inhibits floral bud induction and improves ‘Bing’ sweet cherry fruit quality. HortScience 2006, 41, 654–659. [Google Scholar]
- Einhorn, T.C.; Wang, Y.; Turner, J. Sweet Cherry Fruit Firmness and Postharvest Quality of Late-maturing Cultivars Are Improved with Low-rate, Single Applications of Gibberellic Acid. HortScience 2013, 48, 1010–1017. [Google Scholar] [CrossRef]
- Kuhn, N.; Ponce, C.; Arellano, M.; Time, A.; Sagredo, B.; Donoso, J.M.; Meisel, L.A. Gibberellic acid modifies the transcript abundance of ABA pathway orthologs and modulates sweet cherry (Prunus avium) fruit ripening in early- and mid-season varieties. Plants 2020, 9, 1796. [Google Scholar] [CrossRef] [PubMed]
- Ozturk, B.; Aglar, E.; Saracoglu, O.; Karakaya, O.; Gun, S. Effects of GA3, CACl2 and Modified Atmosphere Packaging (MAP) Applications on Fruit Quality of Sweet Cherry at Cold Storage. Int. J. Fruit Sci. 2022, 22, 696–710. [Google Scholar] [CrossRef]
- Correia, S.; Aires, A.; Queirós, F.; Carvalho, R.; Schouten, R.; Silva, A.P.; Gonçalves, B. Climate conditions and spray treatments induce shifts in health promoting compounds in cherry (Prunus avium L.) fruits. Sci. Hortic. 2020, 263, 109147. [Google Scholar] [CrossRef]
- Chen, J.; Pan, B.Q.; Li, Z.X.; Xu, Y.; Cao, X.M.; Jia, J.J.; Shen, H.L.; Sun, L. Fruit shape loci sun, ovate, fs8.1 and their interactions affect seed size and shape in tomato. Front. Plant Sci. 2022, 13, 1091639. [Google Scholar] [CrossRef]
- Li, J.Y.; Tao, X.Y.; Li, L.; Mao, L.C.; Luo, Z.S.; Khan, Z.U.; Ying, T.J. Comprehensive RNA-Seq Analysis on the Regulation of Tomato Ripening by Exogenous Auxin. PLoS ONE 2016, 11, e0156453. [Google Scholar] [CrossRef]
- Li, J.Y.; Khan, Z.U.; Tao, X.Y.; Mao, L.C.; Luo, Z.S.; Ying, T.J. Effects of exogenous auxin on pigments and primary metabolite profile of postharvest tomato fruit during ripening. Sci. Hortic. 2017, 219, 90–97. [Google Scholar] [CrossRef]
- Ren, Z.; Liu, R.; Gu, W.; Dong, X. The Solanum lycopersicum auxin response factor SlARF2 participates in regulating lateral root formation and flower organ senescence. Plant Sci. 2017, 256, 103–111. [Google Scholar] [CrossRef]
- Liu, J.; Hua, W.; Hu, Z.Y.; Yang, H.L.; Zhang, L.; Li, R.J.; Deng, L.B.; Sun, X.C.; Wang, X.F.; Wang, H.Z. Natural variation in ARF18 gene simultaneously affects seed weight and silique length in polyploid rapeseed. Proc. Natl. Acad. Sci. USA 2015, 112, E5123–E5132. [Google Scholar] [CrossRef] [PubMed]
- Devoghalaere, F.; Doucen, T.; Guitton, B.; Keeling, J.; Payne, W.; Ling, T.J.; Ross, J.J.; Hallett, I.C.; Gunaseelan, K.; Dayatilake, G.A.; et al. A genomics approach to understanding the role of auxin in apple (Malus × domestica) fruit size contro. BMC Plant Biol. 2012, 12, 7. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Li, H.; Ecker, J.R. Ethylene Biosynthesis and Signaling Networks. Plant Cell 2002, 1, S131–S151. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Xu, Y.X.; Zhang, L.C.; Ji, Y.L.; Tan, D.M.; Yuan, H.; Wang, A.D. The Jasmonate-Activated Transcription Factor MdMYC2 Regulates ETHYLENE RESPONSE FACTOR and Ethylene Biosynthetic Genes to Promote Ethylene Biosynthesis during Apple Fruit Ripening. Plant Cell 2017, 29, 1316–1334. [Google Scholar] [CrossRef]
- Muday, G.K.; Rahman, A.; Binder, B.M. Auxin and ethylene: Collaborators or competitors? Trends Plant Sci. 2012, 17, 181–195. [Google Scholar] [CrossRef]
- Cheng, C.X.; Xu, X.Z.; Singer, S.D.; Li, J.; Zhang, H.J.; Gao, M.; Wang, L.; Song, J.Y.; Wang, X.P. Effect of GA3 Treatment on Seed Development and Seed-Related Gene Expression in Grape. PLoS ONE 2013, 8, e80044. [Google Scholar] [CrossRef]
- Bouré, N.; Arnaud, N. Molecular GA pathways as conserved integrators for adaptive responses. Plant Biol. 2023, 25, 647–660. [Google Scholar] [CrossRef]
- Tang, N.; Deng, W.; Hu, G.J.; Hu, N.; Li, Z.G. Transcriptome Profiling Reveals the Regulatory Mechanism Underlying Pollination Dependent and Parthenocarpic Fruit Set Mainly Mediated by Auxin and Gibberellin. PLoS ONE 2015, 10, e0125355. [Google Scholar] [CrossRef]
- Mignolli, F.; Vidoz, M.L.; Picciarelli, P.; Mariotti, L. Gibberellins modulate auxin responses during tomato (Solanum lycopersicum L.) fruit development. Physiol. Plant. 2019, 165, 768–779. [Google Scholar] [CrossRef]
- Blanco-Ulate, B.; Vincenti, E.; Powell, A.L.T.; Cantu, D. Tomato transcriptome and mutant analyses suggest a role for plant stress hormones in the interaction between fruit and Botrytis cinerea . Front. Plant Sci. 2013, 4, 142–158. [Google Scholar] [CrossRef]
- Etesami, H.; Alikhani, H.A.; Akbari, A.A. Evaluation of Plant Growth Hormones Production (IAA) Ability by Iranian Soils Rhizobial Strains and Effects of Superior Strains Application on Wheat Growth Indexes. World Appl. Sci. J. 2009, 6, 1576–1584. [Google Scholar]
- Zahir, Z.A.; Asghar, H.N.; Akhtar, M.J.; Arshad, M. Precursor (L-tryptophan)-Inoculum (Azotobacter) Interaction for Improving Yields and Nitrogen Uptake of Maize. J. Plant Nutr. 2005, 28, 805–817. [Google Scholar] [CrossRef]
- Naveed, M.; Qureshi, M.A.; Zahir, Z.A.; Hussain, M.B.; Sessitsch, A.; Mitter, B. L-Tryptophan-dependent biosynthesis of indole-3-acetic acid (IAA) improves plant growth and colonization of maize by Burkholderia phytofirmans PsJN. Ann. Microbiol. 2015, 65, 1381–1389. [Google Scholar] [CrossRef]
- Sun, T.P.; Kamiya, Y.J. The Arabidopsis GA1 locus encodes the cyclase ent-kaurene synthetase A of gibberellin biosynthesis. Plant Cell 1994, 6, 1509–1518. [Google Scholar] [PubMed]
- Hedden, P.; Thomas, S.G. Gibberellin biosynthesis and its regulation. Biochem. J. 2012, 444, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Wuddineh, W.A.; Mazarei, M.; Zhang, J.Y.; Poovaiah, C.R.; Mann, D.G.J.; Ziebell, A.; Sykes, R.W.; Davis, M.F.; Udvardi, M.K.; Stewart, C.N. Identification and overexpression of gibberellin 2-oxidase (GA2ox) in switchgrass (Panicum virgatum L.) for improved plant architecture and reduced biomass recalcitrance. Plant Biotechnol. J. 2014, 13, 636–647. [Google Scholar] [CrossRef]
- Lee, D.H.; Lee, I.C.; Kim, K.J.; Kim, D.S.; Na, H.J.; Lee, I.J.; Kang, S.M.; Jeon, H.W.; Le, P.Y.; Ko, J.H. Expression of gibberellin 2-oxidase 4 from Arabidopsis under the control of a senescence-associated promoter results in a dominant semi-dwarf plant with normal flowering. J. Plant Biol. 2014, 57, 106–116. [Google Scholar] [CrossRef]
- Fukazawa, J.; Mori, M.; Watanabe, S.; Miyamoto, C.; Ito, T.; Takahashi, Y. DELLA-GAF1 Complex Is a Main Component in Gibberellin Feedback Regulation of GA20 Oxidase 2. Plant Physiol. 2017, 175, 1395–1406. [Google Scholar] [CrossRef]
- Shang, M.Y.; Wang, X.T.; Zhang, J.; Qi, X.H.; Ping, A.; Hou, L.P.; Xing, G.M.; Li, G.Z.; Li, M.L. Genetic Regulation of GA Metabolism during Vernalization, Floral Bud Initiation and Development in Pak Choi (Brassica rapa ssp. chinensis Makino). Front. Plant Sci. 2017, 8, 1533. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Li, A.; Liu, X.Q.; Sun, J.X.; Guo, W.J.; Zhang, J.W.; Lyu, Y.M. Changes in the morphology of the bud meristem and the levels of endogenous hormones after low temperature treatment of different Phalaenopsis cultivars. S. Afr. J. Bot. 2019, 125, 499–504. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, C.X.; Yan, X.J.; Zhang, J.R.; Xu, J.L. Simultaneous analysis of ten phytohormones in Sargassum horneri by high-performance liquid chromatography with electrospray ionization tandem mass spectrometry. J. Sep. Sci. 2016, 39, 1804–1813. [Google Scholar] [CrossRef] [PubMed]
- Floková, K.; Tarkowská, D.; Miersch, O.; Strnad, M.; Wasternack, C.; Novák, O. UHPLC-MS/MS based target profiling of stress-induced phytohormones. Phytochemistry 2014, 105, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.D.; Zhu, J.X.; Gao, Q.; Luo, D.; Yuan, B.F.; Feng, Y.Q. Rapid and high-throughput determination of endogenous cytokinins in Oryza sativa by bare Fe3O4 nanoparticles-based magnetic solid-phase extraction. J. Chromatogr. A 2014, 1340, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Niu, Q.F.; Zong, Y.; Qian, M.J.; Yang, F.X.; Teng, Y.W. Simultaneous quantitative determination of major plant hormones in pear flowers and fruit by UPLC/ESI-MS/MS. Anal. Methods 2014, 6, 1766–1773. [Google Scholar] [CrossRef]
- Xiao, H.M.; Cai, W.J.; Ye, T.T.; Ding, J.; Feng, Y.Q. Spatio-temporal profiling of abscisic acid, indoleacetic acid and jasmonic acid in single rice seed during seed germination. Anal. Chim. Acta 2018, 1031, 119–127. [Google Scholar] [CrossRef]
- Pan, X.; Welti, R.; Wang, X. Quantitative analysis of major plant hormones in crude plant extracts by high-performance liquid chromatography-mass spectrometry. Nat. Protoc. 2010, 5, 986–992. [Google Scholar] [CrossRef] [PubMed]
- Simura, J.; Antoniadi, I.; Siroká, J.; Tarkowská, D.; Strnad, M.; Ljung, K.; Novák, O. Plant Hormonomics: Multiple Phytohormone Profiling by Targeted Metabolomics. Plant Physiol. 2018, 177, 476–489. [Google Scholar] [CrossRef] [PubMed]
- Cui, K.Y.; Lin, Y.Y.; Zhou, X.; Li, S.C.; Liu, H.; Zeng, F.; Zhu, F.; Ouyang, G.F.; Zeng, Z.X. Comparison of sample pretreatment methods for the determination of multiple phytohormones in plant samples by liquid chromatography-electrospray ionization-tandem mass spectrometry. Microchem. J. 2015, 121, 25–31. [Google Scholar] [CrossRef]
- Tang, S.; Chen, N.M.; Song, B.; He, J.Q.; Zhou, Y.J.; Jenks, M.A.; Xu, X.J. Compositional and transcriptomic analysis associated with cuticle lipid production on rosette and inflorescence stem leaves in the extremophyte Thellungiella salsuginea . Physiol. Plant. 2019, 165, 584–603. [Google Scholar] [CrossRef]
- Min, T.; Bao, Y.Q.; Zhou, B.X.; Yi, Y.; Wang, L.M.; Hou, W.F.; Ai, Y.W.; Wang, H.X. Transcription Profiles Reveal the Regulatory Synthesis of Phenols during the Development of Lotus Rhizome (Nelumbo nucifera Gaertn). Int. J. Mol. Sci. 2019, 20, 2735. [Google Scholar] [CrossRef]
- Li, R.; Li, Y.; Kristiansen, K.; Wang, J. SOAP: Short oligonucleotide alignment program. Bioinformatics 2008, 24, 713–714. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Landmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef]
- Audic, S.; Claverie, J.M. The significance of digital gene expression profile. Genome Res. 1997, 7, 986–995. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Index | Compounds | CK (ng/g) | T (ng/g) |
|---|---|---|---|---|
| ABA | ABA | Abscisic acid | 12.97 ± 2.15 | 12.23 ± 0.19 |
| ABA | ABA-GE | ABA-glucosyl ester | 42.20 ± 6.31 | 49.68 ± 1.23 |
| Auxin | IAA-Glu | Indole-3-acetyl glutamic acid | UD | 0.30 ± 0.04 |
| Auxin | IAA-Asp | Indole-3-acetyl-L-aspartic acid | UD | 2.88 ± 0.52 |
| Auxin | TRA | Tryptamine | 0.09 ± 0.04 | 1.81 ± 1.65 |
| Auxin | IPA | 3-Indolepropionic acid | 0.09 ± 0.01 | 0.15 ± 0.02 |
| Auxin | ICA | Indole-3-carboxylic acid | 8.13 ± 2.68 | 10.01 ± 1.65 |
| Auxin | TRP | L-tryptophan | 3297.76 ± 81.85 | 3552.37 ± 97.42 |
| Auxin | MEIAA | Methyl indole-3-acetate | 0.27 ± 0.05 | 4.37 ± 0.34 |
| Auxin | ICAld | Indole-3-carboxaldehyde | 12.94 ± 2.25 | 12.73 ± 0.34 |
| Auxin | IAN | 3-Indoleacetonitrile | 0.19 ± 0.02 | 0.27 ± 0.02 |
| Auxin | IAA-Val-Me | Indole-3-acetyl-L-valine methyl ester | 0.01 ± 0.00 | 0.01 ± 0.00 |
| Auxin | IAA | Indole-3-acetic acid | 1.84 ± 0.24 | 9.25 ± 7.15 |
| ETH | ACC | 1-Aminocyclopropanecarboxylic acid | 177.57 ± 11.87 | 210.49 ± 3.47 |
| GA | GA20 | Gibberellin A20 | 0.57 ± 0.06 | 1.75 ± 0.54 |
| GA | GA4 | Gibberellin A4 | 0.18 ± 0.02 | 0.98 ± 0.03 |
| GA | GA7 | Gibberellin A7 | 0.04 ± 0.01 | 0.24 ± 0.01 |
| GA | GA3 | Gibberellin A3 | 55.49 ± 2.57 | 378.21 ± 32.27 |
| GA | GA1 | Gibberellin A1 | 34.91 ± 4.54 | 35.02 ± 4.25 |
| GA | GA19 | Gibberellin A19 | 0.69 ± 0.01 | 1.71 ± 0.12 |
| Sample | Total Raw Reads (M) | Total Clean Reads (M) | Total Clean Bases (Gb) | Clean Reads Q20 (%) | Clean Reads Q30 (%) | Clean Reads Ratio (%) |
|---|---|---|---|---|---|---|
| CK1 | 43.82 | 42.45 | 6.37 | 97.84 | 93.62 | 96.87 |
| CK2 | 43.82 | 42.43 | 6.36 | 97.78 | 93.39 | 96.82 |
| CK3 | 45.44 | 42.11 | 6.32 | 96.45 | 89.48 | 92.67 |
| T1 | 45.44 | 42.49 | 6.37 | 96.62 | 89.86 | 93.50 |
| T2 | 45.44 | 42.45 | 6.37 | 96.34 | 89.04 | 93.43 |
| T3 | 44.23 | 40.86 | 6.13 | 96.45 | 89.51 | 92.38 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Xu, Y.; Jing, L.; Jiang, B.; Wang, Q.; Wang, Y. Preliminary Study on the Formation Mechanism of Malformed Sweet Cherry (Prunus avium L.) Fruits in Southern China Using Transcriptome and Metabolome Data. Int. J. Mol. Sci. 2024, 25, 153. https://doi.org/10.3390/ijms25010153
Zhang W, Xu Y, Jing L, Jiang B, Wang Q, Wang Y. Preliminary Study on the Formation Mechanism of Malformed Sweet Cherry (Prunus avium L.) Fruits in Southern China Using Transcriptome and Metabolome Data. International Journal of Molecular Sciences. 2024; 25(1):153. https://doi.org/10.3390/ijms25010153
Chicago/Turabian StyleZhang, Wangshu, Yue Xu, Luyang Jing, Baoxin Jiang, Qinghao Wang, and Yuxi Wang. 2024. "Preliminary Study on the Formation Mechanism of Malformed Sweet Cherry (Prunus avium L.) Fruits in Southern China Using Transcriptome and Metabolome Data" International Journal of Molecular Sciences 25, no. 1: 153. https://doi.org/10.3390/ijms25010153