
Inflammasome Coordinates Senescent Chronic Wound Induced by Thalassophryne nattereri Venom
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
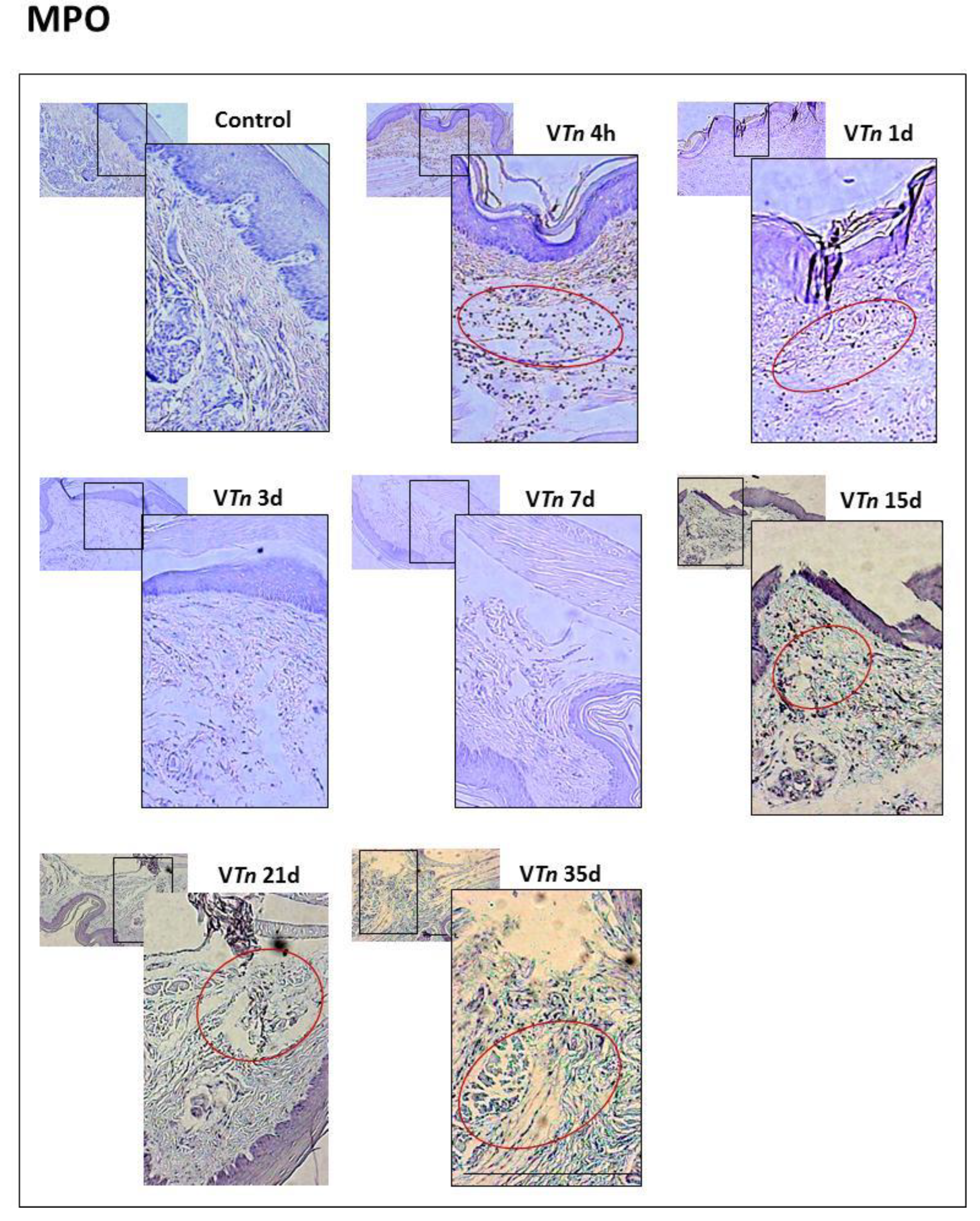
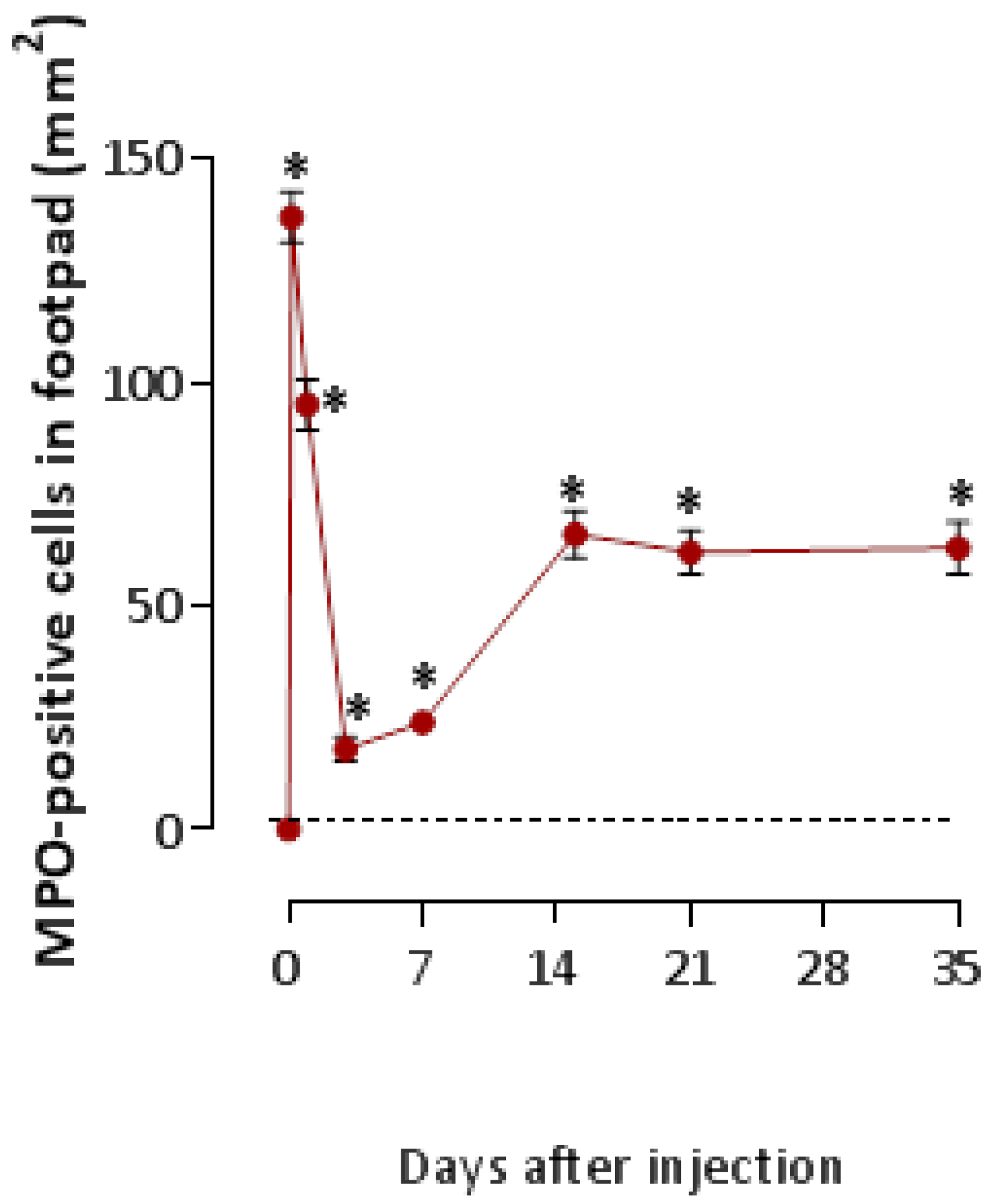
2.1. T. nattereri Venom Induces Long-Term Necrosis
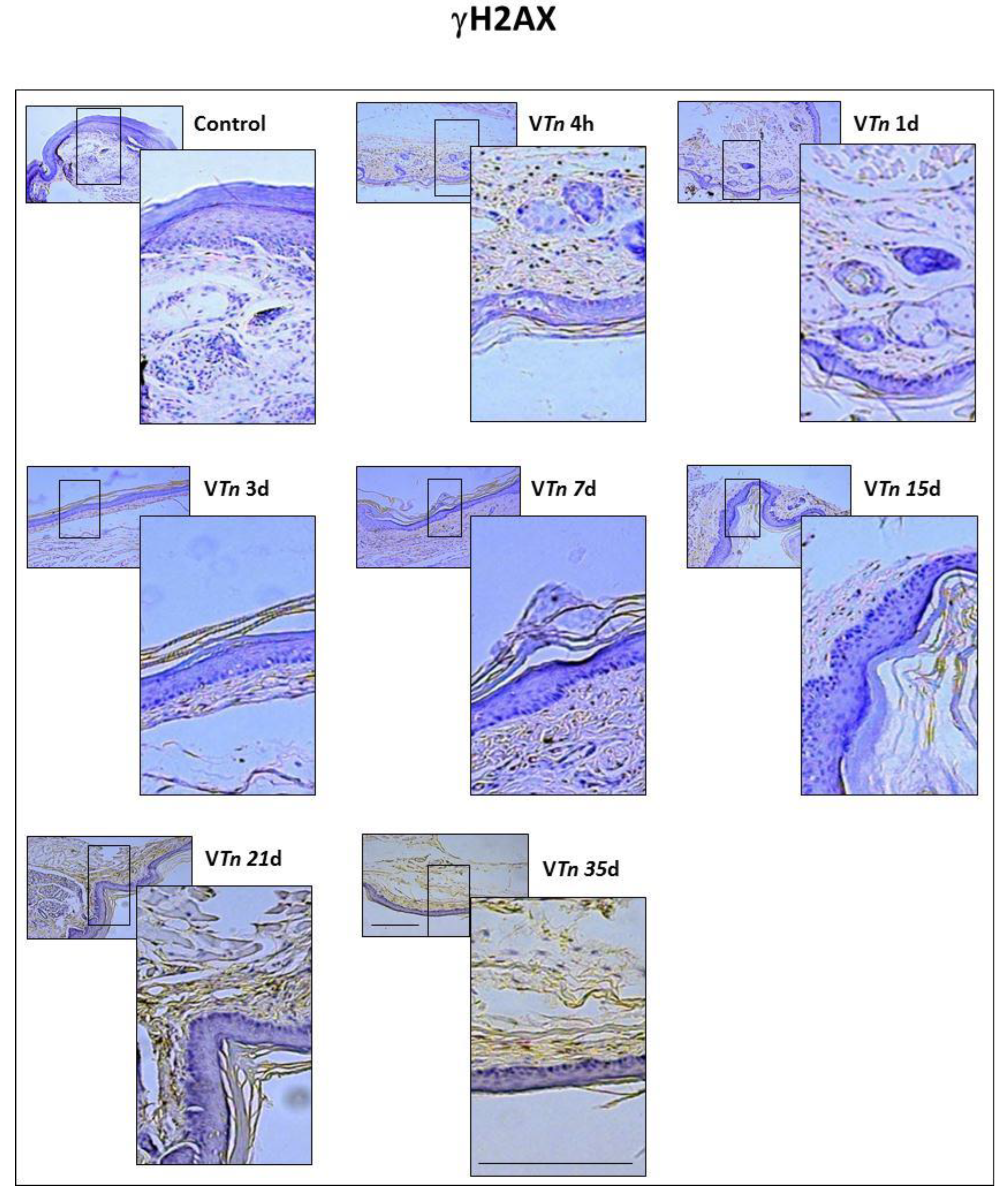
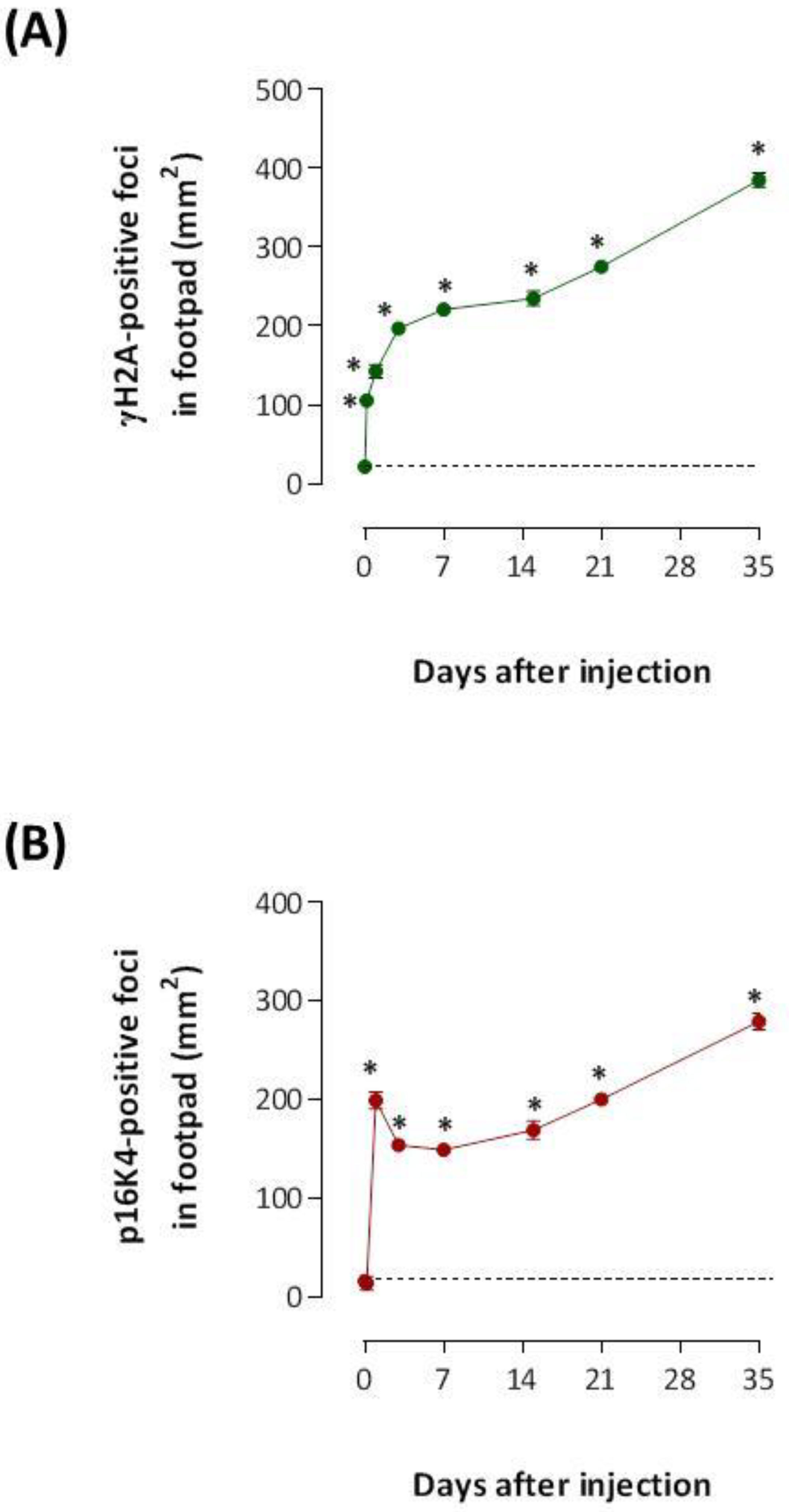
2.2. The Venom of T. nattereri Induces Senescent Wound
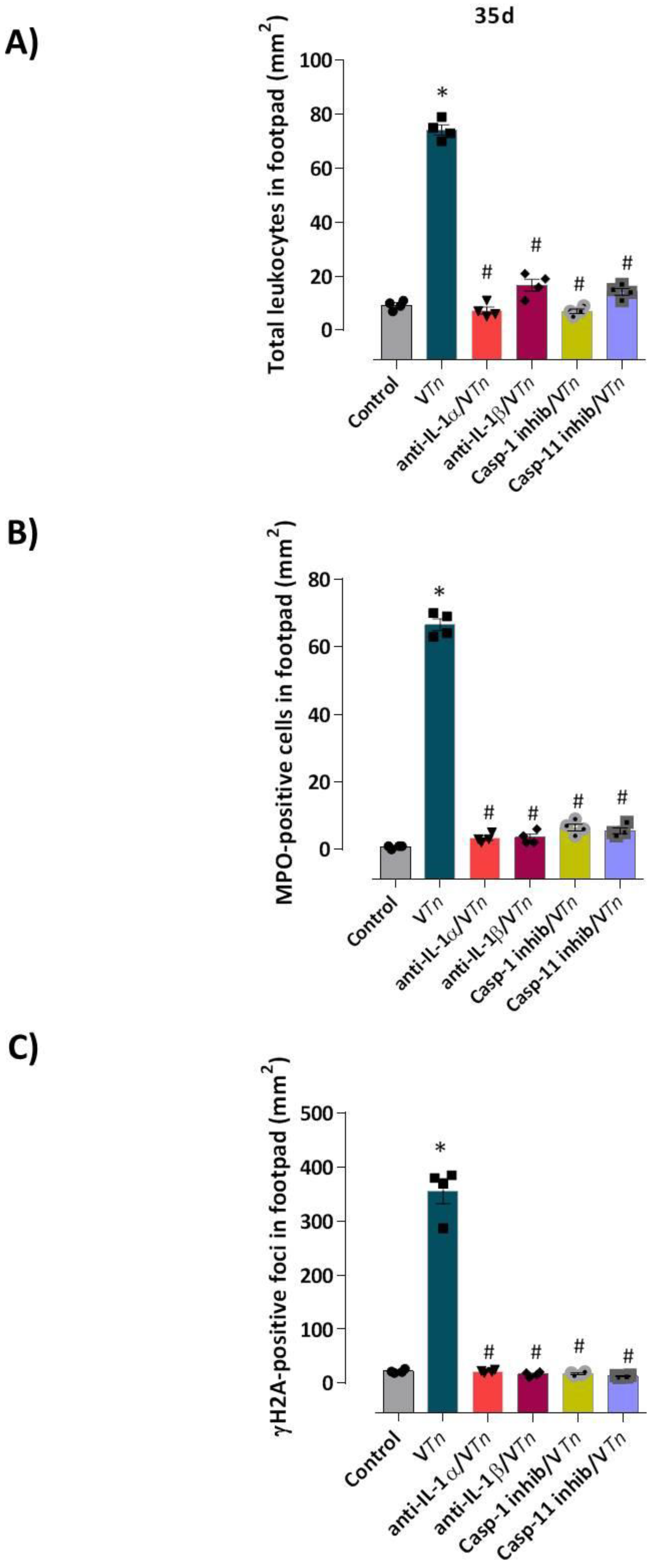
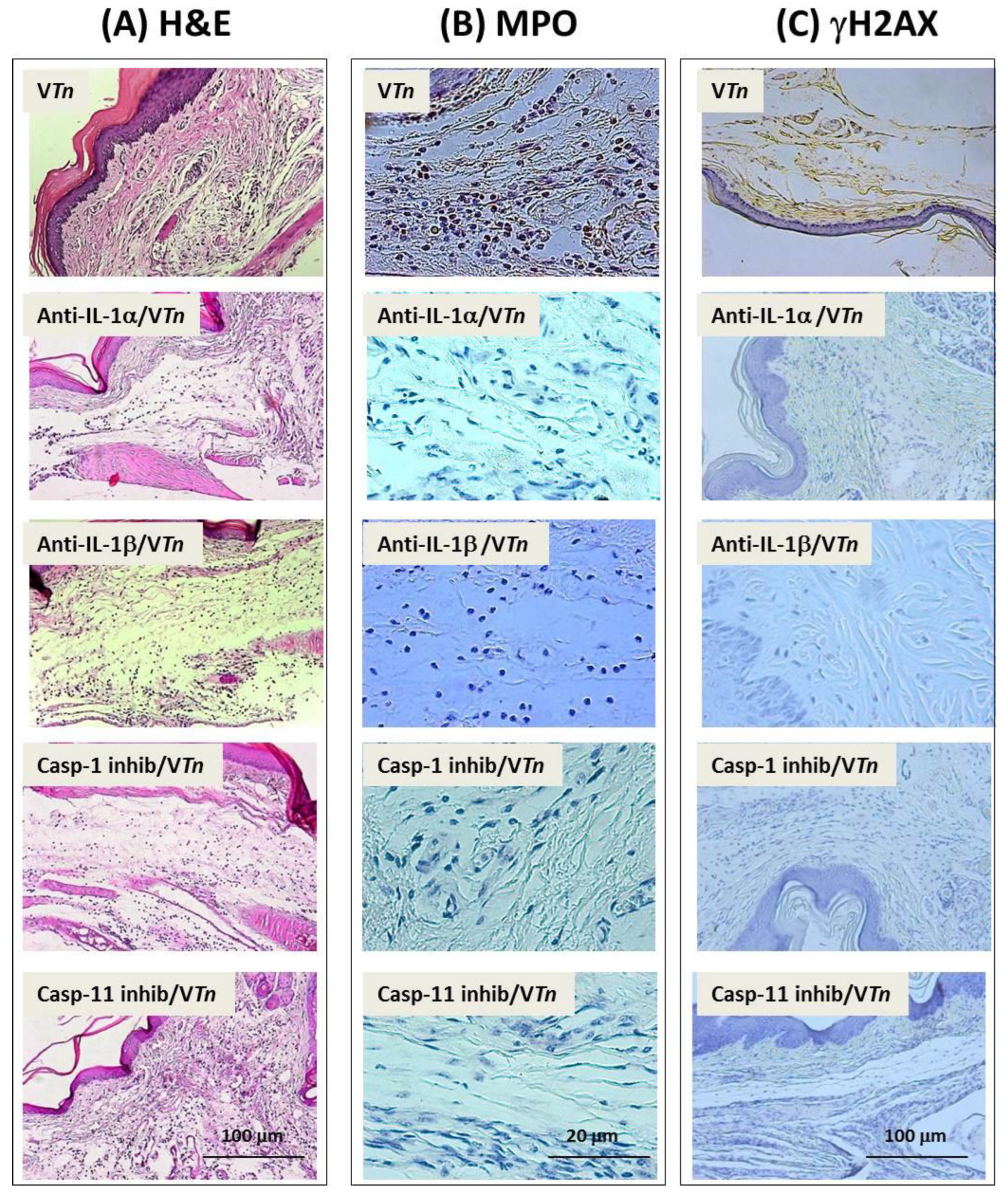
2.3. VTn-Induced Wound Is Associated with IL-1α and IL-1β via Activation of Inflammatory Caspases
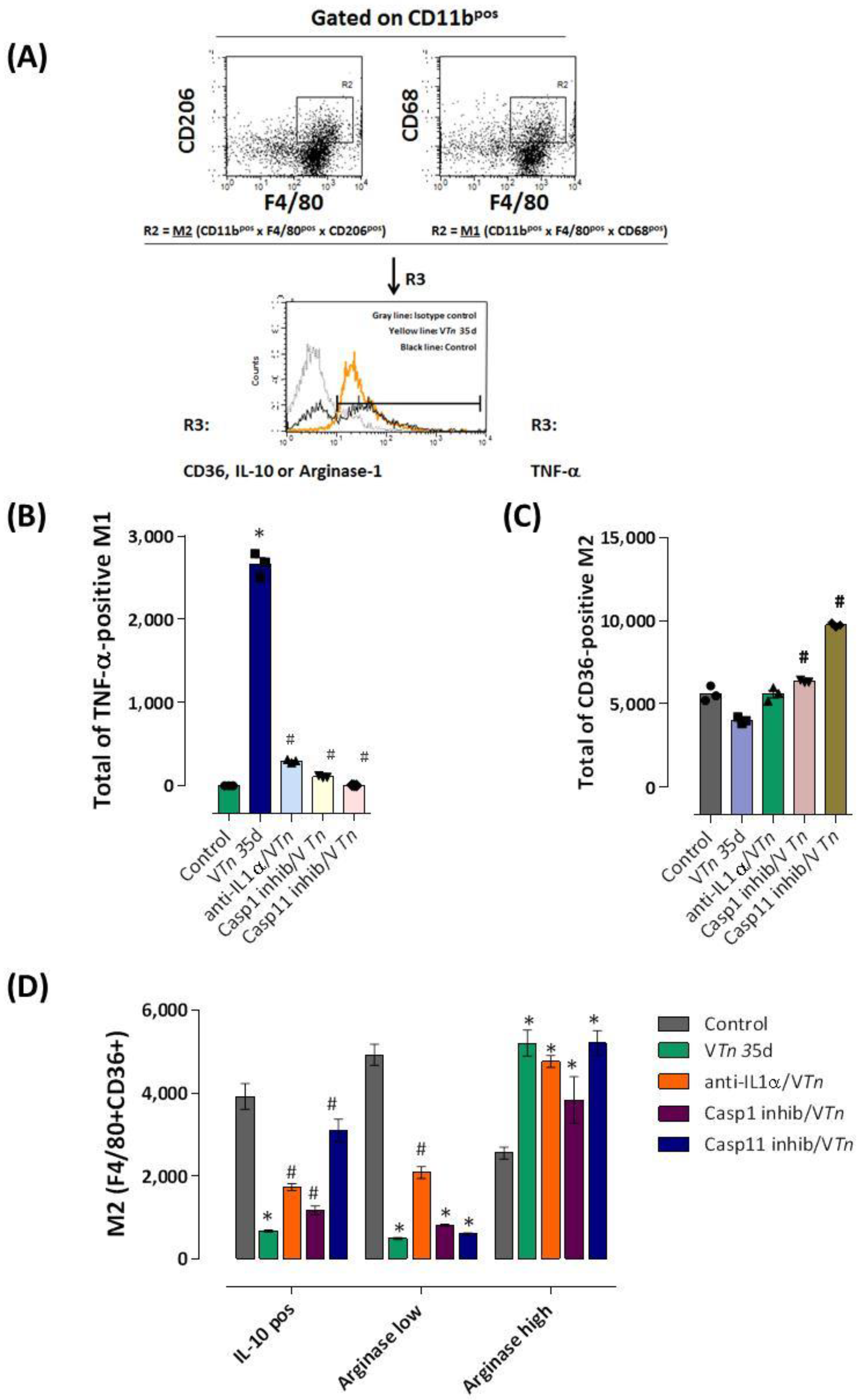
2.4. M1/M2 Macrophage Imbalance in VTn-Induced Chronic Injury Is Dependent on IL-1α and Caspase-1 and Caspase-11
3. Materials and Methods
3.1. Mice
3.2. Thalassophryne Nattereri Venom
3.3. Induction of Paw Injury by VTn and Treatments
3.4. Cell Suspension Collection and Phenotypic Analysis by Flow Cytometry
3.5. Hematoxylin/Eosin Staining
3.6. Immunohistochemistry
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodrigues, M.; Kosaric, N.; Bonham, C.A.; Gurtner, G.C. Wound Healing: A Cellular Perspective. Physiol. Rev. 2019, 99, 665–706. [Google Scholar] [CrossRef]
- Raziyeva, K.; Kim, Y.; Zharkinbekov, Z.; Kassymbek, K.; Jimi, S.; Saparov, A. Immunology of Acute and Chronic Wound Healing. Biomolecules 2021, 11, 700. [Google Scholar] [CrossRef]
- Li, M.; Hou, Q.; Zhong, L.; Zhao, Y.; Fu, X. Macrophage Related Chronic Inflammation in Non-Healing Wounds. Front. Immunol. 2021, 12, 681710. [Google Scholar] [CrossRef] [PubMed]
- Haddad, V.; Pardal, P.P.O.; Cardoso, J.L.C.; Martins, I.A. The Venomous Toadfish Thalassophryne Nattereri (Niquim or Miquim): Report of 43 Injuries Provoked in Fishermen of Salinopolis (Para State) and Aracaju (Sergipe State), Brazil. Rev. Inst. Med. Trop. Sao Paulo 2003, 45, 221–223. [Google Scholar] [CrossRef] [PubMed]
- Facó, P.E.; Bezerra, G.P.; Barbosa, P.S.F.; Martins, A.M.C.; Guimarães, J.A.; Ferreira, M.L.; Monteiro, H.S.A. Epidemiologia Dos Acidentes Por Thalassophryne Nattereri (Niquim) No Estado Do Ceará (1992–2002). Rev. Soc. Bras. Med. Trop. 2005, 38, 479–482. [Google Scholar] [CrossRef]
- Fonseca, L.A.; Lopes-Ferreira, M. Clinical and Experimental Studies Regarding Poisoning Caused by a Fish Thalassophryne Nattereri (Niquim). An. Bras. Dermat. 2000, 75, 435–443. [Google Scholar]
- Lopes-Ferreira, M.; Barbaro, K.C.; Cardoso, D.F.; Moura-Da-Silva, A.M.; Mota, I. Thalassophryne Nattereri Fish Venom: Biological and Biochemical Characterixation and Serum Neutralization of Its Toxic Activities. Toxicon 1998, 36, 405–410. [Google Scholar] [CrossRef]
- Lopes-Ferreira, M.; Emim, J.A.D.S.; Oliveira, V.; Puzer, L.; Cezari, M.H.; Araújo, M.D.S.; Juliano, L.; Lapa, A.J.; Souccar, C.; Moura-Da-Silva, A.M. Kininogenase Activity of Thalassophryne Nattereri Fish Venom. Biochem. Pharmacol. 2004, 68, 2151–2157. [Google Scholar] [CrossRef]
- Lima, C.; Clissa, P.B.; Piran-Soares, A.A.; Tanjoni, I.; Moura-da-Silva, A.M.; Lopes-Ferreira, M. Characterisation of Local Inflammatory Response Induced by Thalassophryne Nattereri Fish Venom in a Mouse Model of Tissue Injury. Toxicon 2003, 42, 499–507. [Google Scholar] [CrossRef]
- Lopes-Ferreira, M.; Núñez, J.; Rucavado, A.; Farsky, S.H.P.; Lomonte, B.; Angulo, Y.; Moura Da Silva, A.M.; Gutiérrez, J.M. Skeletal Muscle Necrosis and Regeneration after Injection of Thalassophryne Nattereri (Niquim) Fish Venom in Mice. Int. J. Exp. Pathol. 2001, 82, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Pareja-Santos, A.; Saraiva, T.C.; Costa, E.P.; Santos, M.F.; Zorn, T.T.; Souza, V.M.O.; Lopes-Ferreira, M.; Lima, C. Delayed Local Inflammatory Response Induced by Thalassophryne Nattereri Venom Is Related to Extracellular Matrix Degradation. Int. J. Exp. Pathol. 2009, 90, 34–43. [Google Scholar] [CrossRef]
- Lopes-Ferreira, M.; Moura-Da-Silva, A.M.; Piran-Soares, A.A.; Angulo, Y.; Lomonte, B.; Gutiérrez, J.M.; Farsky, S.H.P. Hemostatic Effects Induced by Thalassophryne Nattereri Fish Venom: A Model of Endothelium-Mediated Blood Flow Impairment. Toxicon 2002, 40, 1141–1147. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Li, M.; Zheng, Z.; Ma, J.; Gao, Y.; Chen, L.; Peng, Y.; Yu, S.; Yang, L. Senescence in Chronic Wounds and Potential Targeted Therapies. Burn. Trauma 2022, 10, tkab045. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Bano, D.; Ehninger, D. Cellular Senescence in Vivo: From Cells to Tissues to Pathologies. Mech. Ageing Dev. 2020, 190, 111308. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, S.; Mancini, E.; Xu, L.; Moore, A.; Jahanbani, F.; Hebestreit, K.; Srinivasan, R.; Li, X.; Devarajan, K.; Prélot, L.; et al. Heterogeneity in old fibroblasts is linked to variability in reprogramming and wound healing. Nature 2019, 574, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Dipietro, L.A. Factors affecting wound healing. J. Dent. Res. 2010, 89, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Chelvarajan, R.L.; Liu, Y.; Popa, D.; Getchell, M.L.; Getchell, T.V.; Stromberg, A.J.; Bondada, S. Molecular basis of age-associated cytokine dysregulation in LPS-stimulated macrophages. J. Leukoc. Biol. 2006, 79, 1314–1327. [Google Scholar] [CrossRef]
- Gosain, A.; DiPietro, L.A. Aging and wound healing. World J. Surg. 2004, 28, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Dellago, H.; Preschitz-Kammerhofer, B.; Terlecki-Zaniewicz, L.; Schreiner, C.; Fortschegger, K.; Chang, M.W.; Hackl, M.; Monteforte, R.; Kühnel, H.; Schosserer, M.; et al. High levels of oncomiR-21 contribute to the senescence-induced growth arrest in normal human cells and its knock-down increases the replicative lifespan. Aging Cell 2013, 12, 446–458. [Google Scholar] [CrossRef]
- Wiggins, K.A.; Clarke, M.C.H. Senescence Utilises Inflammatory Caspases to Drive the SASP. Aging 2019, 11, 3891–3892. [Google Scholar] [CrossRef]
- Fernández-Duran, I.; Quintanilla, A.; Tarrats, N.; Birch, J.; Hari, P.; Millar, F.R.; Lagnado, A.B.; Smer-Barreto, V.; Muir, M.; Brunton, V.G.; et al. Cytoplasmic Innate Immune Sensing by the Caspase-4 Non-Canonical Inflammasome Promotes Cellular Senescence. Cell Death Differ. 2022, 29, 1267–1282. [Google Scholar] [CrossRef] [PubMed]
- Phillipson, M.; Kubes, P. The Healing Power of Neutrophils. Trends Immunol. 2019, 40, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Krzyszczyk, P.; Schloss, R.; Palmer, A.; Berthiaume, F. The Role of Macrophages in Acute and Chronic Wound Healing and Interventions to Promote Pro-Wound Healing Phenotypes. Front. Physiol. 2018, 9, 419. [Google Scholar] [CrossRef] [PubMed]
- Slater, T.W.; Finkielsztein, A.; Mascarenhas, L.A.; Mehl, L.C.; Butin-Israeli, V.; Sumagin, R. Neutrophil Microparticles Deliver Active Myeloperoxidase to Injured Mucosa To Inhibit Epithelial Wound Healing. J. Immunol. 2017, 198, 2886–2897. [Google Scholar] [CrossRef]
- Chen, W.Y.J.; Rogers, A.A. Recent Insights into the Causes of Chronic Leg Ulceration in Venous Diseases and Implications on Other Types of Chronic Wounds. Wound Repair Regen. 2007, 15, 434–449. [Google Scholar] [CrossRef]
- Wilkinson, H.N.; Hardman, M.J. Senescence in Wound Repair: Emerging Strategies to Target Chronic Healing Wounds. Front. Cell Dev. Biol. 2020, 8, 773. [Google Scholar] [CrossRef]
- Wang, Z.; Shi, C. Cellular Senescence Is a Promising Target for Chronic Wounds: A Comprehensive Review. Burn. Trauma 2020, 8, tkaa021. [Google Scholar] [CrossRef]
- Krishnamurthy, J.; Torrice, C.; Ramsey, M.R.; Kovalev, G.I.; Al-Regaiey, K.; Su, L.; Sharpless, N.E. Ink4a/Arf Expression Is a Biomarker of Aging. J. Clin. Investig. 2004, 114, 1299–1307. [Google Scholar] [CrossRef] [PubMed]
- Smirnov, A.; Panatta, E.; Lena, A.M.; Castiglia, D.; Daniele, N.D.; Melino, G.; Candi, E. FOXM1 Regulates Proliferation, Senescence and Oxidative Stress in Keratinocytes and Cancer Cells. Aging 2016, 8, 1384–1397. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.; Wu, S.; Zhang, L.; Chen, G.; Lai, W. Retarding the Senescence of Human Vascular Endothelial Cells Induced by Hydrogen Peroxide: Effects of 17beta-Estradiol (E2) Mediated Mitochondria Protection. Biogerontology 2014, 15, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Coppé, J.P.; Rodier, F.; Patil, C.K.; Freund, A.; Desprez, P.Y.; Campisi, J. Tumor Suppressor and Aging Biomarker P16 INK4a Induces Cellular Senescence without the Associated Inflammatory Secretory Phenotype. J. Biol. Chem. 2011, 286, 36396–36403. [Google Scholar] [CrossRef]
- Freund, A.; Patil, C.K.; Campisi, J. P38MAPK Is a Novel DNA Damage Response-Independent Regulator of the Senescence-Associated Secretory Phenotype. EMBO J. 2011, 30, 1536–1548. [Google Scholar] [CrossRef] [PubMed]
- Viganò, E.; Diamond, C.E.; Spreafico, R.; Balachander, A.; Sobota, R.M.; Mortellaro, A. Human Caspase-4 and Caspase-5 Regulate the One-Step Non-Canonical Inflammasome Activation in Monocytes. Nat. Commun. 2015, 6, 8761. [Google Scholar] [CrossRef]
- Daniels, M.J.D.; Brough, D. Unconventional Pathways of Secretion Contribute to Inflammation. Int. J. Mol. Sci. 2017, 18, 102. [Google Scholar] [CrossRef]
- Ratitong, B.; Marshall, M.; Pearlman, E. β-Glucan-Stimulated Neutrophil Secretion of IL-1α Is Independent of GSDMD and Mediated through Extracellular Vesicles. Cell Rep. 2021, 35, 109139. [Google Scholar] [CrossRef] [PubMed]
- Kuilman, T.; Michaloglou, C.; Vredeveld, L.C.W.; Douma, S.; van Doorn, R.; Desmet, C.J.; Aarden, L.A.; Mooi, W.J.; Peeper, D.S. Oncogene-Induced Senescence Relayed by an Interleukin-Dependent Inflammatory Network. Cell 2008, 133, 1019–1031. [Google Scholar] [CrossRef]
- Orjalo, A.V.; Bhaumik, D.; Gengler, B.K.; Scott, G.K.; Campisi, J. Cell Surface-Bound IL-1α Is an Upstream Regulator of the Senescence-Associated IL-6/IL-8 Cytokine Network. Proc. Natl. Acad. Sci. USA 2009, 106, 17031–17036. [Google Scholar] [CrossRef] [PubMed]
- Gardner, S.E.; Humphry, M.; Bennett, M.R.; Clarke, M.C.H. Senescent Vascular Smooth Muscle Cells Drive Inflammation through an Interleukin-1α-Dependent Senescence-Associated Secretory Phenotype. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 1963–1974. [Google Scholar] [CrossRef]
- Acosta, J.C.; Banito, A.; Wuestefeld, T.; Georgilis, A.; Janich, P.; Morton, J.P.; Athineos, D.; Kang, T.W.; Lasitschka, F.; Andrulis, M.; et al. A complex secretory program orchestrated by the inflammasome controls paracrine senescence. Nat. Cell Biol. 2013, 15, 978–990. [Google Scholar] [CrossRef]
- Wiggins, K.A.; Parry, A.J.; Cassidy, L.D.; Humphry, M.; Webster, S.J.; Goodall, J.C.; Narita, M.; Clarke, M.C.H. IL-1α Cleavage by Inflammatory Caspases of the Noncanonical Inflammasome Controls the Senescence-Associated Secretory Phenotype. Aging Cell 2019, 18, e12946. [Google Scholar] [CrossRef]
- Prattichizzo, F.; De Nigris, V.; Mancuso, E.; Spiga, R.; Giuliani, A.; Matacchione, G.; Lazzarini, R.; Marcheselli, F.; Recchioni, R.; Testa, R.; et al. Short-Term Sustained Hyperglycaemia Fosters an Archetypal Senescence-Associated Secretory Phenotype in Endothelial Cells and Macrophages. Redox Biol. 2018, 15, 170–181. [Google Scholar] [CrossRef]
- Covarrubias, A.J.; Kale, A.; Perrone, R.; Dominguez, J.A.L.-; Pisco, A.O.; Kasler, H.G.; Schmidt, M.S.; Heckenbach, I.; Kwok, R.; Wiley, C.D.; et al. Senescent Cells Promote Tissue NAD+ Decline during Ageing via the Activation of CD38+ Macrophages Anthony. Nat. Metab. 2021, 2, 1265–1283. [Google Scholar] [CrossRef] [PubMed]
- Hall, B.M.; Balan, V.; Gleiberman, A.S.; Strom, E.; Krasnov, P.; Virtuoso, L.P.; Rydkina, E.; Vujcic, S.; Balan, K.; Gitlin, I.; et al. Aging of Mice Is Associated with P16(Ink4a)- and β-Galactosidasepositive Macrophage Accumulation That Can Be Induced in Young Mice by Senescent Cells. Aging 2016, 8, 1294–1315. [Google Scholar] [CrossRef] [PubMed]
- Lefèvre, L.; Iacovoni, J.S.; Martini, H.; Bellière, J.; Maggiorani, D.; Dutaur, M.; Marsal, D.J.; Decaunes, P.; Pizzinat, N.; Mialet-Perez, J.; et al. Kidney Inflammaging Is Promoted by CCR2+ Macrophages and Tissue-Derived Micro-Environmental Factors. Cell. Mol. Life Sci. 2021, 78, 3485–3501. [Google Scholar] [CrossRef]
- Littlewood-Evans, A.; Sarret, S.; Apfel, V.; Loesle, P.; Dawson, J.; Zhang, J.; Muller, A.; Tigani, B.; Kneuer, R.; Patel, S.; et al. GPR91 Senses Extracellular Succinate Released from Inflammatory Macrophages and Exacerbates Rheumatoid Arthritis. J. Exp. Med. 2016, 213, 1655–1662. [Google Scholar] [CrossRef]
- Fremder, M.; Kim, S.W.; Khamaysi, A.; Shimshilashvili, L.; Eini-Rider, H.; Park, I.S.; Hadad, U.; Cheon, J.H.; Ohana, E. A Transepithelial Pathway Delivers Succinate to Macrophages, Thus Perpetuating Their pro-Inflammatory Metabolic State. Cell Rep. 2021, 36, 109521. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, E.M.; Menga, A.; Martín-Pérez, R.; Quinto, A.; Riera-Domingo, C.; De Tullio, G.; Hooper, D.C.; Lamers, W.H.; Ghesquière, B.; McVicar, D.W.; et al. Pharmacologic or Genetic Targeting of Glutamine Synthetase Skews Macrophages toward an M1-like Phenotype and Inhibits Tumor Metastasis. Cell Rep. 2017, 20, 1654–1666. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Roos, A.; Schlagwein, N.; Woltman, A.M.; Daha, M.R.; Van Kooten, C. IL-10-Producing Macrophages Preferentially Clear Early Apoptotic Cells. Blood 2006, 107, 4930–4937. [Google Scholar] [CrossRef] [PubMed]
- Shouval, D.S.; Biswas, A.; Goettel, J.A.; McCann, K.; Conaway, E.; Redhu, N.S.; Mascanfroni, I.D.; Adham, Z.A.; Lavoie, S.; Ibourk, M.; et al. Interleukin-10 Receptor Signaling in Innate Immune Cells Regulates Mucosal Immune Tolerance and Anti-Inflammatory Macrophage Function. Immunity 2014, 30, 1289–1303. [Google Scholar] [CrossRef] [PubMed]
- Ip, W.K.E.; Hoshi, N.; Shouval, D.S.; Snapper, S.; Medzhitov, R. Anti-Inflammatory Effect of IL-10 Mediated by Metabolic Reprogramming of Macrophages. Science (80-) 2017, 176, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Yue, Z.; Xu, M.; Zhang, M.; Shen, X.; Ma, Z.; Li, J.; Xie, X. Macrophages Play a Key Role in Tissue Repair and Regeneration. PeerJ 2022, 10, e14053. [Google Scholar] [CrossRef] [PubMed]
- Szondi, D.C.; Wong, J.K.; Vardy, L.A.; Cruickshank, S.M. Arginase Signalling as a Key Player in Chronic Wound Pathophysiology and Healing. Front. Mol. Biosci. 2021, 8, 773866. [Google Scholar] [CrossRef] [PubMed]
- Pennathur, S.; Pasichnyk, K.; Bahrami, N.M.; Zeng, L.; Febbraio, M.; Yamaguchi, I.; Okamura, D.M. The Macrophage Phagocytic Receptor CD36 Promotes Fibrogenic Pathways on Removal of Apoptotic Cells during Chronic Kidney Injury. Am. J. Pathol. 2015, 185, 2232–2245. [Google Scholar] [CrossRef] [PubMed]
- Sindrilaru, A.; Peters, T.; Wieschalka, S.; Baican, C.; Baican, A.; Peter, H.; Hainzl, A.; Schatz, S.; Qi, Y.; Schlecht, A.; et al. An unrestrained proinflammatory M1 macrophage population induced by iron impairs wound healing in humans and mice. J. Clin. Investig. 2011, 121, 985–997. [Google Scholar] [CrossRef] [PubMed]
- Shimada, N.; Sakata, A.; Igarashi, T.; Takeuchi, M.; Nishimura, S. M1 Macrophage Infiltration Exacerbate Muscle/Bone Atrophy after Peripheral Nerve Injury. BMC Musculoskelet. Disord. 2020, 21, 44. [Google Scholar] [CrossRef]
- Li, M.; Sun, X.; Zhao, J.; Xia, L.; Li, J.; Xu, M.; Wang, B.; Guo, H.; Yu, C.; Gao, Y.; et al. CCL5 Deficiency Promotes Liver Repair by Improving Inflammation Resolution and Liver Regeneration through M2 Macrophage Polarization. Cell. Mol. Immunol. 2020, 17, 753–764. [Google Scholar] [CrossRef]
- Zenaro, E.; Pietronigro, E.; Bianca, V.D.; Piacentino, G.; Marongiu, L.; Budui, S.; Turano, E.; Rossi, B.; Angiari, S.; Dusi, S.; et al. Neutrophils Promote Alzheimer’s Disease-like Pathology and Cognitive Decline via LFA-1 Integrin. Nat. Med. 2015, 21, 880–886. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima, C.; Andrade-Barros, A.I.; Carvalho, F.F.; Falcão, M.A.P.; Lopes-Ferreira, M. Inflammasome Coordinates Senescent Chronic Wound Induced by Thalassophryne nattereri Venom. Int. J. Mol. Sci. 2023, 24, 8453. https://doi.org/10.3390/ijms24098453
Lima C, Andrade-Barros AI, Carvalho FF, Falcão MAP, Lopes-Ferreira M. Inflammasome Coordinates Senescent Chronic Wound Induced by Thalassophryne nattereri Venom. International Journal of Molecular Sciences. 2023; 24(9):8453. https://doi.org/10.3390/ijms24098453
Chicago/Turabian StyleLima, Carla, Aline Ingrid Andrade-Barros, Fabiana Franco Carvalho, Maria Alice Pimentel Falcão, and Monica Lopes-Ferreira. 2023. "Inflammasome Coordinates Senescent Chronic Wound Induced by Thalassophryne nattereri Venom" International Journal of Molecular Sciences 24, no. 9: 8453. https://doi.org/10.3390/ijms24098453