

Serelaxin Alleviates Fibrosis in Thyroid-Associated Ophthalmopathy via the Notch Pathway
 ,
,
Abstract
:1. Introduction
2. Results
2.1. mRNA Expression Profile and Functional Enrichment Analysis of DEGs
2.2. Validation of DEGs in the Relaxin Signaling Pathway
2.3. Relaxin Signaling Pathway Expression Level in Human Orbital Connective Tissue
2.4. RLN2 Expression in Serum and Supernatants
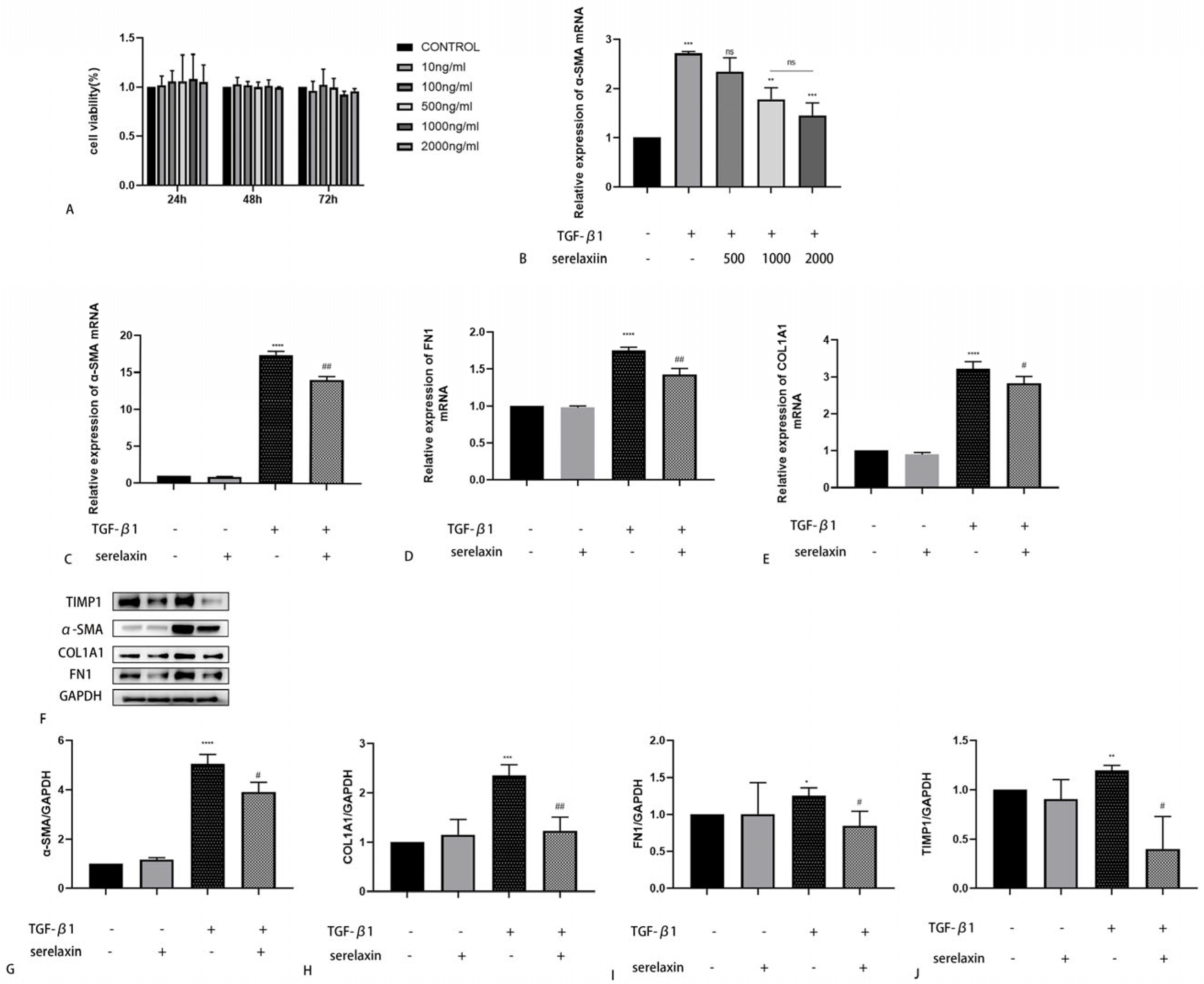
2.5. Serelaxin Alleviates Fibrosis in OFs of TAO
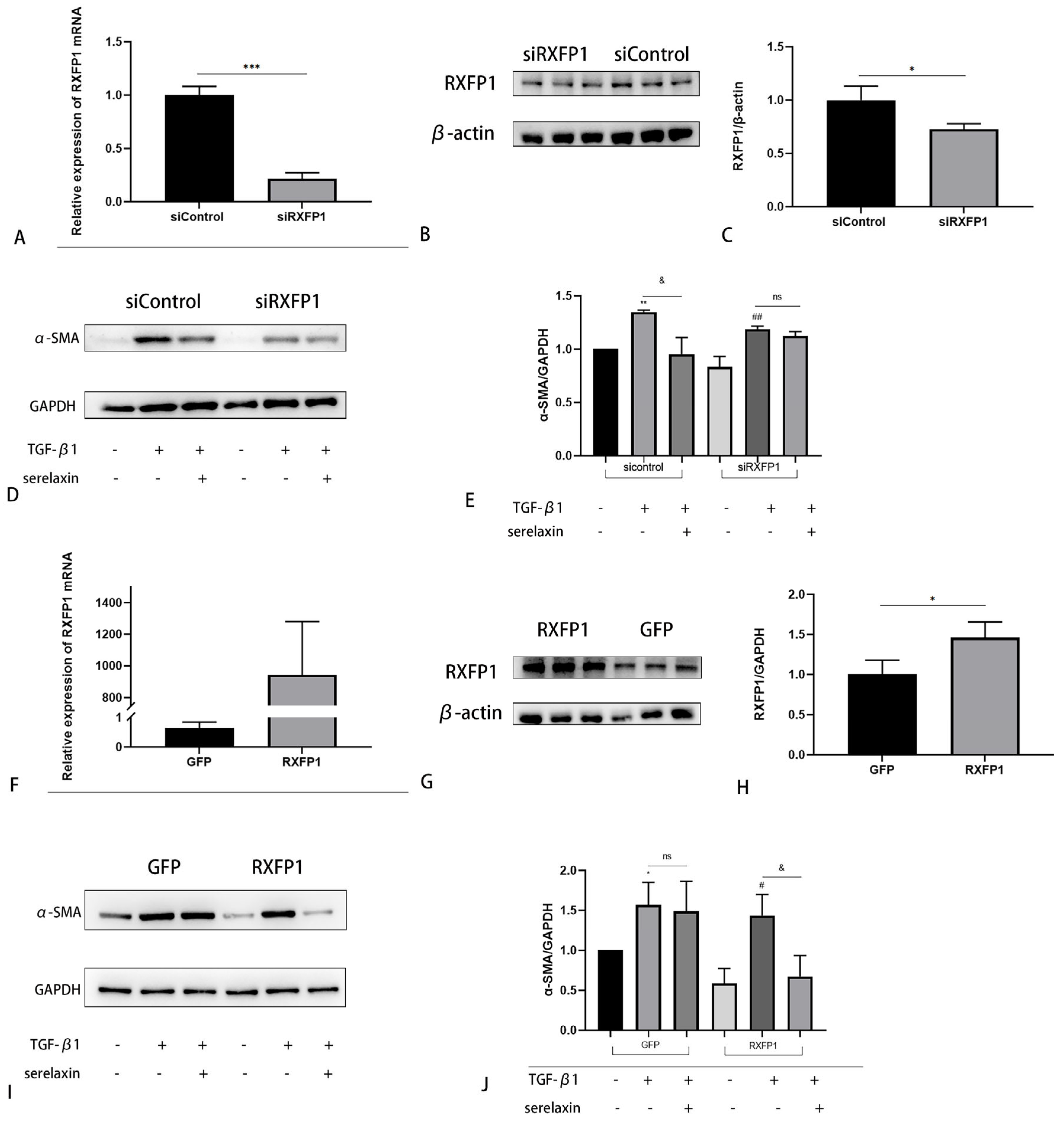
2.6. The Antifibrotic Effect of Serelaxin Is Dependent on RXFP1
2.7. TGF-β1 Regulates RXFP1 Expression in OFs
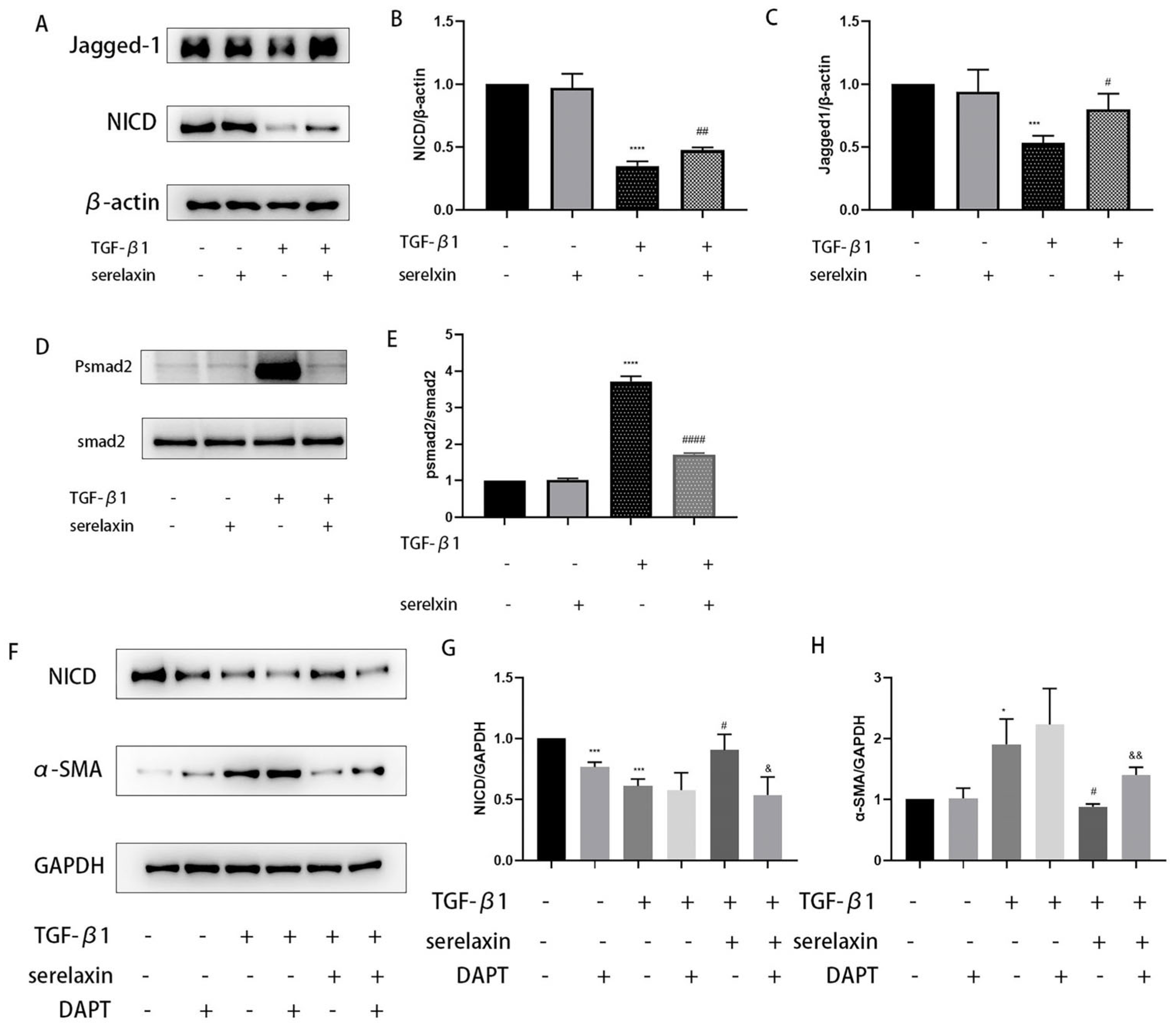
2.8. Serelaxin Alleviates Inflammation in the OFs of TAO
3. Discussion
4. Materials and Methods
4.1. Patient Selection and Tissue Samples
4.2. RNA Sequencing
4.3. Functional Enrichment Analysis
4.4. Construction of a PPI Network and Identification of Hub Genes
4.5. Quantitative Real-Time Polymerase Chain Reaction (qRT–PCR)
4.6. Histologic Staining
4.7. Immunofluorescence (IF)
4.8. Cell Culture
4.9. Cell Proliferation/Toxicity Assay
4.10. Western Blotting
4.11. Enzyme-Linked Immunosorbent Assay (ELISA)
4.12. Overexpression or Silencing of RXFP1
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bahn, R.S. Graves’ ophthalmopathy. N. Engl. J. Med. 2010, 362, 726–738. [Google Scholar] [CrossRef]
- Kendall-Taylor, P. The pathogenesis of graves’ ophthalmopathy. Clin. Endocrinol. Metab. 1985, 14, 331–349. [Google Scholar] [CrossRef]
- Place, R.F.; Krieger, C.C.; Neumann, S.; Gershengorn, M.C. Inhibiting thyrotropin/insulin-like growth factor 1 receptor crosstalk to treat Graves’ ophthalmopathy: Studies in orbital fibroblasts in vitro. Br. J. Pharm. 2017, 174, 328–340. [Google Scholar] [CrossRef]
- Aniszewski, J.P.; Valyasevi, R.W.; Bahn, R.S. Relationship between Disease Duration and Predominant Orbital T Cell Subset in Graves’ Ophthalmopathy. J. Clin. Endocrinol. Metab. 2000, 85, 776–780. [Google Scholar] [CrossRef] [PubMed]
- GWick, G.; Grundtman, C.; Mayerl, C.; Wimpissinger, T.F.; Feichtinger, J.; Zelger, B.; Sgonc, R.; Wolfram, D. The Immunology of Fibrosis. Annu. Rev. Immunol. 2013, 31, 107–135. [Google Scholar]
- Kahaly, G.J.; Riedl, M.; König, J.; Pitz, S.; Ponto, K.; Diana, T.; Kampmann, E.; Kolbe, E.; Eckstein, A.; Moeller, L.; et al. Mycophenolate plus methylprednisolone versus methylprednisolone alone in active, moderate-to-severe Graves’ orbitopathy (MINGO): A randomised, observer-masked, multicentre trial. Lancet Diabetes Endocrinol. 2018, 6, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Bartalena, L.; Kahaly, G.J.; Baldeschi, L.; Dayan, C.M.; Eckstein, A.; Marcocci, C.; Marinò, M.; Vaidya, B.; Wiersinga, W.M.; Ayvaz, G.; et al. The 2021 European Group on Graves’ orbitopathy (EUGOGO) clinical practice guidelines for the medical management of Graves’ orbitopathy. Eur. J. Endocrinol. 2021, 185, G43–G67. [Google Scholar] [CrossRef]
- Sherwood, O.D. Relaxin’s Physiological Roles and Other Diverse Actions. Endocr. Rev. 2004, 25, 205–234. [Google Scholar] [CrossRef]
- Ivell, R.; Einspanier, A. Relaxin peptides are new global players. Trends Endocrinol. Metab. 2002, 13, 343–348. [Google Scholar] [CrossRef]
- Dschietzig, T.; Bartsch, C.; Baumann, G.; Stangl, K. Relaxin—A pleiotropic hormone and its emerging role for experimental and clinical therapeutics. Pharmacol. Ther. 2006, 112, 38–56. [Google Scholar] [CrossRef]
- Samuel, C.; Hewitson, T. Relaxin in cardiovascular and renal disease. Kidney Int. 2006, 69, 1498–1502. [Google Scholar] [CrossRef] [PubMed]
- Samuel, C.S.; Summers, R.J.; Hewitson, T.D. Antifibrotic Actions of Serelaxin–New Roles for an Old Player. Trends Pharmacol. Sci. 2016, 37, 485–497. [Google Scholar] [CrossRef]
- Metra, M.; Teerlink, J.R.; Cotter, G.; Davison, B.A.; Felker, G.M.; Filippatos, G.; Greenberg, B.H.; Pang, P.S.; Ponikowski, P.; Voors, A.A.; et al. Effects of Serelaxin in Patients with Acute Heart Failure. N. Engl. J. Med. 2019, 381, 716–726. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Sun, A.; Xiao, W.; Zhang, T.; Xu, Z.; Shi, L.; Sha, X.; Yang, H. Differential Circular RNA Expression Profiling of Orbital Connective Tissue From Patients with Type I and Type II Thyroid-Associated Ophthalmopathy. Investig. Opthalmology Vis. Sci. 2022, 63, 27. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Bennett, R.G. Relaxin signaling activates peroxisome proliferator-activated receptor gamma. Mol. Cell. Endocrinol. 2010, 315, 239–245. [Google Scholar] [CrossRef]
- Hombach-Klonisch, S.; Bialek, J.; Trojanowicz, B.; Weber, E.; Holzhausen, H.-J.; Silvertown, J.D.; Summerlee, A.J.; Dralle, H.; Hoang-Vu, C.; Klonisch, T. Relaxin Enhances the Oncogenic Potential of Human Thyroid Carcinoma Cells. Am. J. Pathol. 2006, 169, 617–632. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.J.; Hegedüs, L. Graves’ Disease. N. Engl. J. Med. 2016, 375, 1552–1565. [Google Scholar] [CrossRef]
- Ponto, K.A.; Hommel, G.; Pitz, S.; Elflein, H.; Pfeiffer, N.; Kahaly, G.J. Quality of Life in a German Graves Orbitopathy Population. Am. J. Ophthalmol. 2011, 152, 483–490.e1. [Google Scholar] [CrossRef]
- Chen, C.Z.; Raghunath, M. Focus on collagen: In vitro systems to study fibrogenesis and antifibrosis_state of the art. Fibrogenesis Tissue Repair 2009, 2, 7. [Google Scholar] [CrossRef]
- Menconi, F.; Profilo, M.A.; Leo, M.; Sisti, E.; Altea, M.A.; Rocchi, R.; Marinò, M. Spontaneous improvement of untreated mild Graves’ ophthalmopathy: Rundle’s curve revisited. Thyroid 2014, 24, 60–66. [Google Scholar] [CrossRef]
- Zhao, P.; Deng, Y.; Gu, P.; Wang, Y.; Zhou, H.; Hu, Y.; Chen, P.; Fan, X. Insulin-like growth factor 1 promotes the proliferation and adipogenesis of orbital adipose-derived stromal cells in thyroid-associated ophthalmopathy. Exp. Eye Res. 2013, 107, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhou, R.; Diao, J.; Chen, X.; Huang, J.; Xu, K.; Ling, L.; Xia, W.; Liang, Y.; Liu, G.; et al. Differentially expressed circular RNAs in orbital adipose/connective tissue from patients with thyroid-associated ophthalmopathy. Exp. Eye Res. 2020, 196, 108036. [Google Scholar] [CrossRef]
- Ng, H.H.; Shen, M.; Samuel, C.S.; Schlossmann, J.; Bennett, R.G. Relaxin and extracellular matrix remodeling: Mechanisms and signaling pathways. Mol. Cell. Endocrinol. 2019, 487, 59–65. [Google Scholar] [CrossRef]
- Teerlink, J.R.; Cotter, G.; Davison, B.A.; Felker, G.M.; Filippatos, G.; Greenberg, B.H.; Ponikowski, P.; Unemori, E.; Voors, A.A.; Adams, K.F.; et al. Serelaxin, recombinant human relaxin-2, for treatment of acute heart failure (RELAX-AHF): A randomised, placebo-controlled trial. Lancet 2013, 381, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Prabhakar, B.S.; Bahn, R.S.; Smith, T. Current Perspective on the Pathogenesis of Graves’ Disease and Ophthalmopathy. Endocr. Rev. 2003, 24, 802–835. [Google Scholar] [CrossRef]
- Hemmann, S.; Graf, J.; Roderfeld, M.; Roeb, E. Expression of MMPs and TIMPs in liver fibrosis-a systematic review with special emphasis on antifibrotic strategies. J. Hepatol. 2007, 46, 955–975. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Huang, Y.; Wang, S.; Zhang, Y.; Luo, X.; Liu, L.; Zhong, S.; Liu, X.; Li, D.; Liang, R.; et al. IL-17A Exacerbates Fibrosis by Promoting the Proinflammatory and Profibrotic Function of Orbital Fibroblasts in TAO. J. Clin. Endocrinol. Metab. 2016, 101, 2955–2965. [Google Scholar] [CrossRef]
- Zenz, R.; Eferl, R.; Scheinecker, C.; Redlich, K.; Smolen, J.; Schonthaler, H.B.; Kenner, L.; Tschachler, E.; Wagner, E.F. Activator protein 1 (Fos/Jun) functions in inflammatory bone and skin disease. Thromb. Haemost. 2008, 10, 201. [Google Scholar] [CrossRef] [PubMed]
- Romero-Kusabara, I.L.; Filho, J.V.; Scalissi, N.M.; Melo, K.C.; Demartino, G.; Longui, C.A.; Melo, M.R.; Cury, A.N. Distinct inflammatory gene expression in extraocular muscle and fat from patients with Graves’ orbitopathy. Eur. J. Endocrinol. 2017, 176, 481–488. [Google Scholar] [CrossRef]
- Su, H.; Xie, J.; Wen, L.; Wang, S.; Chen, S.; Li, J.; Qi, C.; Zhang, Q.; He, X.; Zheng, L.; et al. LncRNA Gas5 regulates Fn1 deposition via Creb5 in renal fibrosis. Epigenomics 2021, 13, 699–713. [Google Scholar] [CrossRef]
- Bouter, Y.; Kacprowski, T.; Weissmann, R.; Dietrich, K.; Borgers, H.; Brauß, A.; Bayer, T.A. Deciphering the molecular profile of plaques, memory decline and neuron loss in two mouse models for Alzheimer’s disease by deep sequencing. Front. Aging Nerosci. 2014, 6, 75. [Google Scholar]
- Zhu, D.; Gu, X.; Lin, Z.; Yu, D.; Wang, J. High expression of PSMC2 promotes gallbladder cancer through regulation of GNG4 and predicts poor prognosis. Oncogenesis 2021, 10, 43. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.Y.T. New insights into the evolution of the relaxin–LGR signaling system. Trends Endocrinol. Metab. 2003, 14, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Sheau, Y.H.; Nakabayashi, K.; Nishi, S.; Kumagai, J.; Kudo, M.; Sherwood, O.D.; Hsueh, A.J.W. Activation of orphan receptors by the hormone relaxin. Science 2002, 295, 671–674. [Google Scholar] [CrossRef]
- Unemori, E.N.; Pickford, L.B.; Salles, A.L.; Piercy, C.E.; Grove, B.H.; Erikson, M.E.; Amento, E.P. Relaxin induces an extracellular matrix-degrading phenotype in human lung fibroblasts in vitro and inhibits lung fibrosis in a murine model in vivo. J. Clin. Investig. 1996, 98, 2739–2745. [Google Scholar] [CrossRef] [PubMed]
- Dik, W.A.; Virakul, S.; van Steensel, L. Current perspectives on the role of orbital fibroblasts in the pathogenesis of Graves’ ophthalmopathy. Exp. Eye Res. 2016, 142, 83–91. [Google Scholar] [CrossRef]
- Kisseleva, T.; Brenner, D. Molecular and cellular mechanisms of liver fibrosis and its regression. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 151–166. [Google Scholar] [CrossRef]
- Rybinski, B.; Franco-Barraza, J.; Cukierman, E. The wound healing, chronic fibrosis, and cancer progression triad. Physiol. Genom. 2014, 46, 223–244. [Google Scholar] [CrossRef]
- Visse, R.; Nagase, H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: Structure, function, and biocheistry. Circ. Res. 2003, 92, 827–839. [Google Scholar] [CrossRef]
- Artavanis-Tsakonas, S.; Rand, M.D.; Lake, R.J. Notch Signaling: Cell Fate Control and Signal Integration in Development. Science 1999, 284, 770–776. [Google Scholar] [CrossRef]
- Bray, S.J. Notch signalling: A simple pathway becomes complex. Nat. Rev. Mol. Cell Biol. 2006, 7, 678–689. [Google Scholar] [CrossRef]
- Aoyagi-Ikeda, K.; Maeno, T.; Matsui, H.; Ueno, M.; Hara, K.; Aoki, Y.; Kurabayashi, M. Notch induces myofibroblast differentiation of alveolar epithelial cells via transforming growth fac-tor-{beta}-Smad3 pathway. Am. J. Respir. Cell. Mol. Biol. 2011, 45, 136–144. [Google Scholar] [PubMed]
- Sweetwyne, M.T.; Tao, J.; Susztak, K. Kick it up a notch: Notch signaling and kidney fibrosis. Kidney Int. Suppl. 2014, 4, 91–96. [Google Scholar] [CrossRef]
- Chen, S.; Xu, L.; Lin, N.; Pan, W.; Hu, K.; Xu, R. Activation of Notch1 signaling by marrow-derived mesenchymal stem cells through cell–cell contact inhibits proliferation of hepatic stellate cells. Life Sci. 2011, 89, 975–981. [Google Scholar] [CrossRef]
- Nemir, M.; Metrich, M.; Plaisance, I.; Lepore, M.; Cruchet, S.; Berthonneche, C.; Sarre, A.; Radtke, F.; Pedrazzini, T. The Notch pathway controls fibrotic and regenerative repair in the adult heart. Eur. Heart J. 2014, 35, 2174–2185. [Google Scholar] [CrossRef] [PubMed]
- Dees, C.; Tomcik, M.; Zerr, P.; Akhmetshina, A.; Horn, A.; Palumbo, K.; Distler, J.H. Notch signalling regulates fibroblast activation and collagen release in systemic sclerosis. Ann. Rheum. Dis. 2011, 70, 1304–1310. [Google Scholar] [CrossRef]
- Hu, B.; Phan, S.H. Notch in fibrosis and as a target of antifibrotic therapy. Pharmacol. Res. 2016, 108, 57–64. [Google Scholar] [CrossRef]
- Zhou, X.; Chen, X.; Cai, J.J.; Chen, L.Z.; Gong, Y.S.; Wang, L.X.; Gao, Z.; Zhang, H.Q.; Huang, W.J.; Zhou, H. Relaxin inhibits cardiac fibrosis and endothelial–mesenchymal transition via the Notch pathway. Drug Des. Dev. Ther. 2015, 9, 4599–4611. [Google Scholar] [CrossRef] [PubMed]
- Sassoli, C.; Chellini, F.; Pini, A.; Tani, A.; Nistri, S.; Nosi, D.; Formigli, L. Relaxin prevents cardiac fibroblast-myofibroblast transition via notch-1-mediated inhibition of TGF-beta/Smad3 signaling. PLoS ONE 2013, 8, e63896. [Google Scholar] [CrossRef]
- Wilhelmi, T.; Xu, X.; Tan, X.; Hulshoff, M.S.; Maamari, S.; Sossalla, S.; Zeisberg, M.; Zeisberg, E.M. Serelaxin alleviates cardiac fibrosis through inhibiting endothelial-to-mesenchymal transition via RXFP1. Theranostics 2020, 10, 3905–3924. [Google Scholar] [CrossRef]
- Mazella, J.; Tang, M.; Tseng, L. Disparate effects of relaxin and TGFbeta1: Relaxin increases, but TGFbeta1 inhibits, the relaxin receptor and the production of IGFBP-1 in human endometrial stromal/decidual cells. Hum. Reprod. 2004, 19, 1513–1518. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Wang, Y.; Liu, Z.; Yu, Z.; Guan, K.; Liu, M.; Wang, M.; Tan, J.; Huang, L. Hepatic macrophages act as a central hub for relaxin-mediated alleviation of liver fibrosis. Nat. Nanotechnol. 2021, 16, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Diao, J.; Chen, X.; Mou, P.; Ma, X.; Wei, R. Potential Therapeutic Activity of Berberine in Thyroid-Associated Ophthalmopathy: Inhibitory Effects on Tissue Remodeling in Orbital Fibroblasts. Investig. Opthalmol. Vis. Sci. 2022, 63, 6. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.S.; Ghosh, S. Shared principles in NF-kappaB signaling. Cell 2008, 132, 344–362. [Google Scholar] [CrossRef]
- Méndez-Samperio, P. Expression and regulation of chemokines in mycobacterial infection. J. Infect. 2008, 57, 374–384. [Google Scholar] [CrossRef]
- Cassetta, L.; Fragkogianni, S.; Sims, A.H.; Swierczak, A.; Forrester, L.M.; Zhang, H.; Soong, D.Y.H.; Cotechini, T.; Anur, P.; Lin, E.Y.; et al. Human Tumor-Associated Macrophage and Monocyte Transcriptional Landscapes Reveal Cancer-Specific Reprogramming, Biomarkers, and Therapeutic Targets. Cancer Cell 2019, 35, 588–602.e10. [Google Scholar] [CrossRef] [PubMed]
- Pawlowski, P.; Reszec, J.; Eckstein, A.; Johnson, K.; Grzybowski, A.; Chyczewski, L.; Mysliwiec, J. Markers of inflammation and fibrosis in the orbital fat/connective tissue of patients with Graves’ orbitopathy: Clinical implications. Mediat. Inflamm. 2014, 2014, 412158. [Google Scholar] [CrossRef]
- Bartley, G.B.; Gorman, C.A. Diagnostic Criteria for Graves’ Ophthalmopathy. Am. J. Ophthalmol. 1995, 119, 792–795. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Characteristics | RNA-Sequencing Stage | Experiments Stage | ||||
|---|---|---|---|---|---|---|
| TAO (n = 12) | Control (n = 6) | p | TAO (n = 6) | Control (n = 6) | p | |
| Age, y | 41.5 ± 13.5 | 44.5 ± 12.8 | 0.657 | 47.2 ± 13.0 | 46.7 ± 13.5 | 0.949 |
| Sex, M/F | 4/8 | 3/3 | 0.494 | 4/2 | 3/3 | 0.558 |
| CAS | 1 (range: 0–2) | - | NA | 1 (range: 0–2) | - | NA |
| NOSPECS score | 4 (range: 3–6) | - | NA | 4 (range: 3–6) | - | NA |
| TAO duration, mo | 25 (IQR: 18–29.8; range: 12–36) | - | NA | 27.5 (IQR: 22–34.3; range: 19–35) | - | NA |
| TAO (n = 22) | Control (n = 10) | p | |
|---|---|---|---|
| Sex (M/F) | 13/9 | 6/4 | 0.961 |
| Age (years) | 47.68 ± 10.13 | 41.6 ± 12.23 | 0.1389 |
| CAS | 2 (range: 0–2) | - | NA |
| Duration of TAO (mo) | 17.5 (IQR: 12–24; range: 12–38) | - | NA |
| Therapy History | Glucocorticoids: 7/22 Thyroid surgery: 3/22 | - | NA |
| Thyroid Function | Euthyroid: 20/22 Hypothyroid: 2/22 Hyperthyroid: 0/22 | - | NA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, A.; Ye, H.; Xu, Z.; Chen, J.; Xiao, W.; Zhang, T.; Sha, X.; Bi, S.; Zhou, T.; Yang, H. Serelaxin Alleviates Fibrosis in Thyroid-Associated Ophthalmopathy via the Notch Pathway. Int. J. Mol. Sci. 2023, 24, 8356. https://doi.org/10.3390/ijms24098356
Sun A, Ye H, Xu Z, Chen J, Xiao W, Zhang T, Sha X, Bi S, Zhou T, Yang H. Serelaxin Alleviates Fibrosis in Thyroid-Associated Ophthalmopathy via the Notch Pathway. International Journal of Molecular Sciences. 2023; 24(9):8356. https://doi.org/10.3390/ijms24098356
Chicago/Turabian StyleSun, Anqi, Huijing Ye, Zhihui Xu, Jingqiao Chen, Wei Xiao, Te Zhang, Xiaotong Sha, Shaowei Bi, Tianyi Zhou, and Huasheng Yang. 2023. "Serelaxin Alleviates Fibrosis in Thyroid-Associated Ophthalmopathy via the Notch Pathway" International Journal of Molecular Sciences 24, no. 9: 8356. https://doi.org/10.3390/ijms24098356