
Predictive Value of MUC5AC Signature in Pancreatic Ductal Adenocarcinoma: A Hypothesis Based on Preclinical Evidence

Abstract
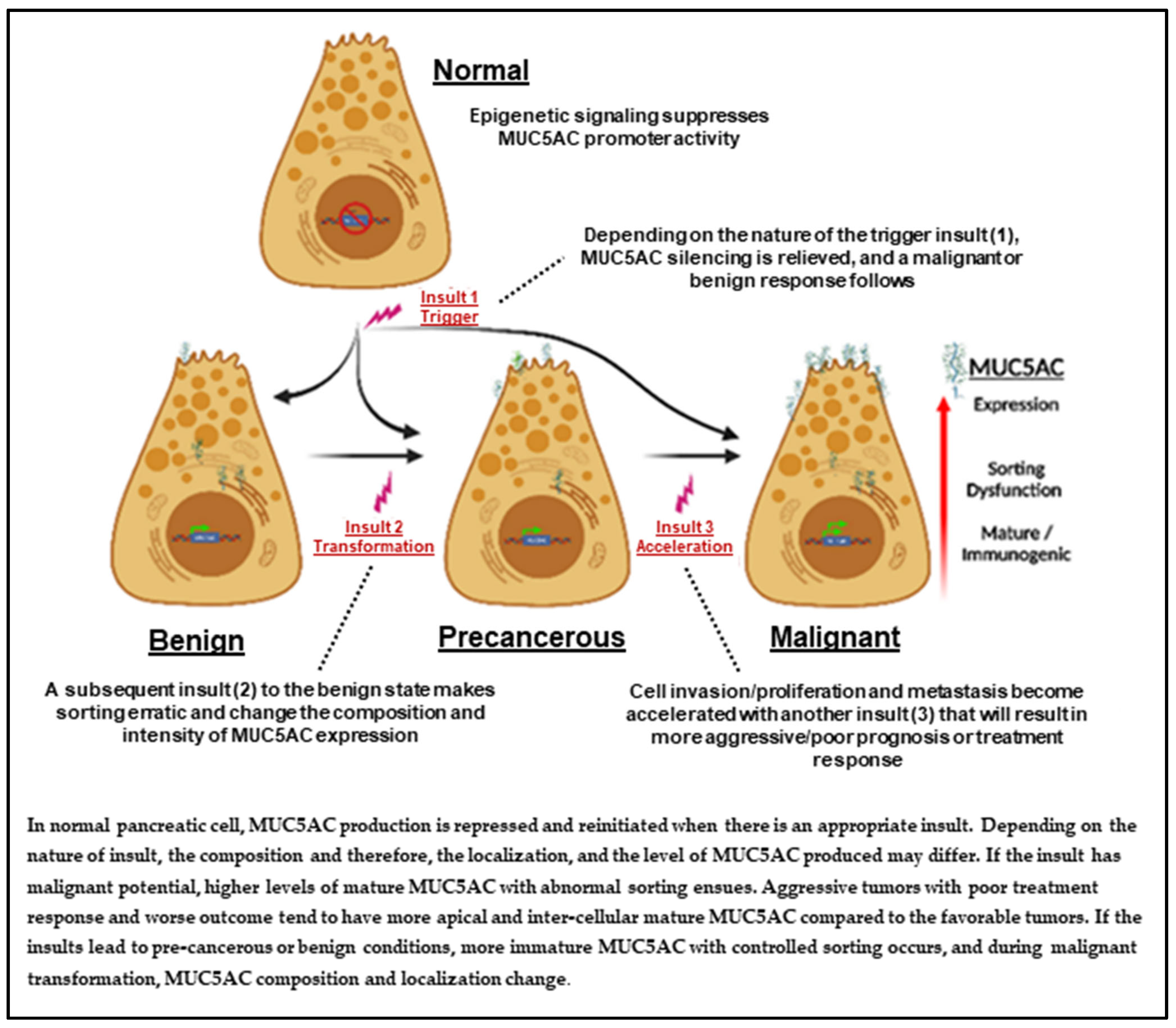
:1. Introduction
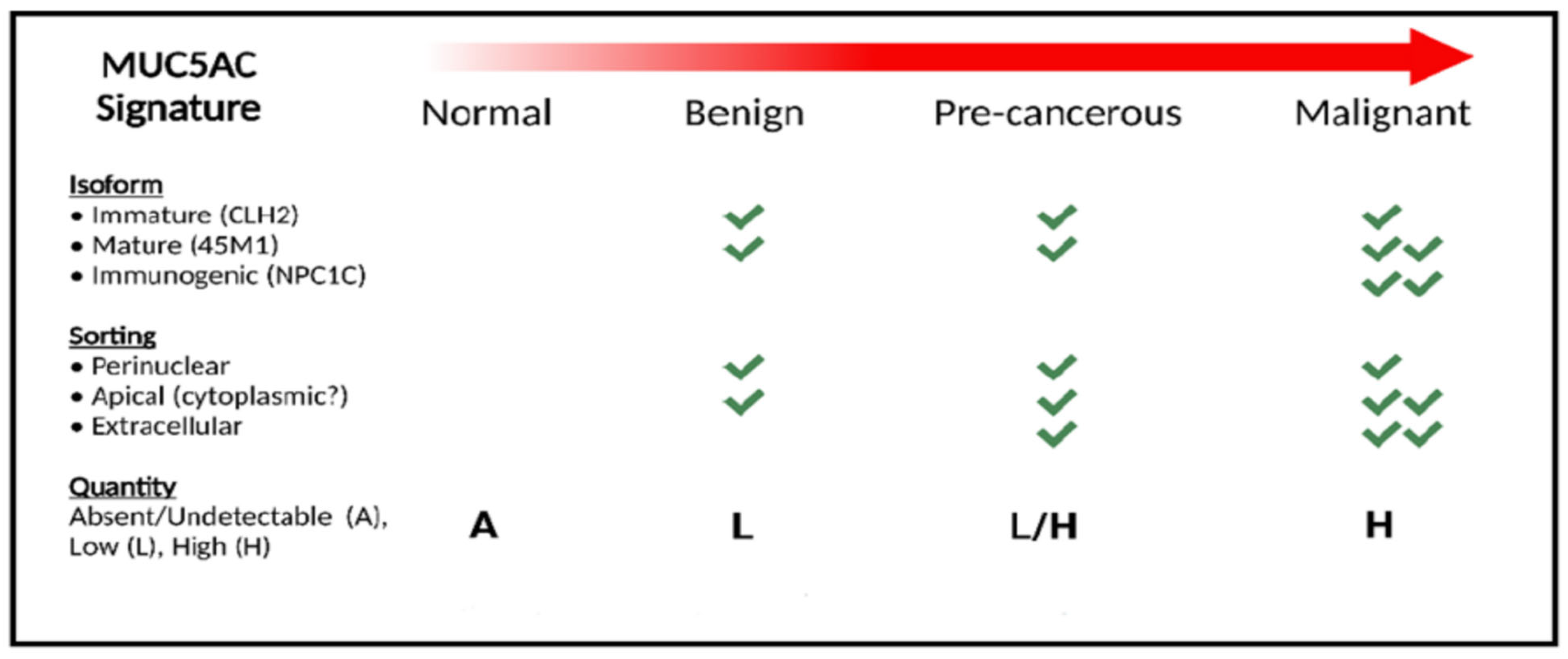
2. MUC5AC Signature
Strengthening Our Hypothesis Using MUC5AC Signature
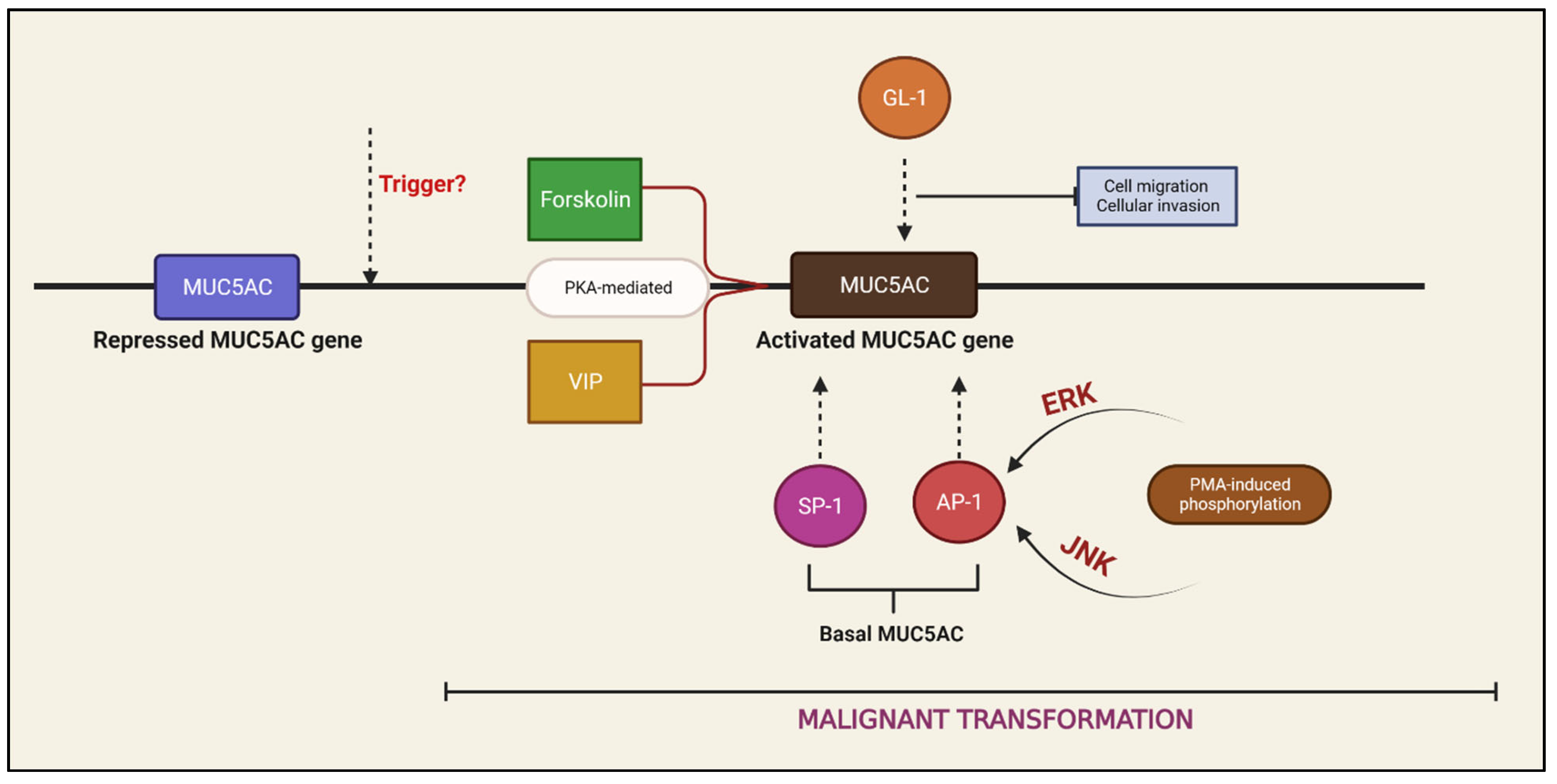
3. Regulation of MUC5AC Expression
4. Predictive Value of MUC5AC
4.1. Comparing PCLs Based on MUC5AC Expression
4.2. Comparing Basic Pathological Characteristics
4.3. Comparing Malignant Potential
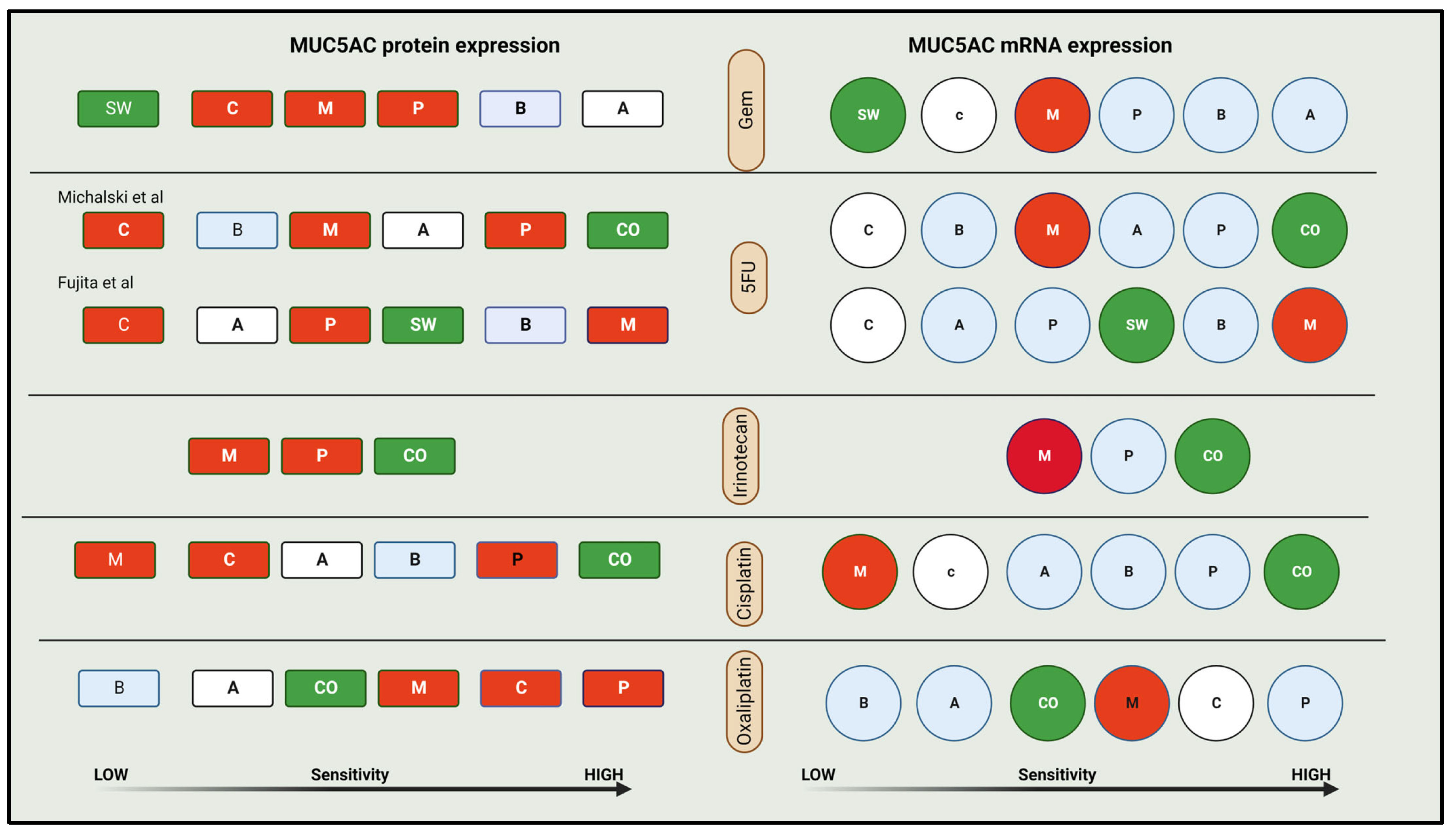
4.4. Comparing Chemosensitivity
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Surveillance, Epidemiology, and End Results (SEER) Program. SEER*Stat Database: Incidence—SEER Research Data, 9 Registries, Nov 2019 Sub (1975–2017). Available online: www.seer.cancer.gov (accessed on 17 April 2023).
- Wainberg, Z.A.; Melisi, D.; Macarulla, T.; Pazo-Cid, R.; Chandana, S.R.; De La Fouchardiere, C.; Dean, A.P.; Kiss, I.; Lee, W.; Goetze, T.O.; et al. NAPOLI-3: A randomized, open-label phase 3 study of liposomal irinotecan + 5-fluorouracil/leucovorin + oxaliplatin (NALIRIFOX) versus nab-paclitaxel + gemcitabine in treatment-naïve patients with metastatic pancreatic ductal adenocarcinoma (mPDAC). J. Clin. Oncol. 2023, 41, LBA661. [Google Scholar] [CrossRef]
- Sheel, A.; Addison, S.; Nuguru, S.P.; Manne, A. Is Cell-Free DNA Testing in Pancreatic Ductal Adenocarcinoma Ready for Prime Time? Cancers 2022, 14, 3453. [Google Scholar] [CrossRef] [PubMed]
- Bansil, R.; Turner, B.S. Mucin structure, aggregation, physiological functions and biomedical applications. Curr. Opin. Colloid Interface Sci. 2006, 11, 164–170. [Google Scholar] [CrossRef]
- Kebouchi, M.; Hafeez, Z.; Le Roux, Y.; Dary-Mourot, A.; Genay, M. Importance of digestive mucus and mucins for designing new functional food ingredients. Food Res. Int. 2020, 131, 108906. [Google Scholar] [CrossRef] [PubMed]
- Manne, A.; Esnakula, A.; Abushahin, L.; Tsung, A. Understanding the Clinical Impact of MUC5AC Expression on Pancreatic Ductal Adenocarcinoma. Cancers 2021, 13, 3059. [Google Scholar] [CrossRef]
- Pan, S.; Chen, R.; Tamura, Y.; Crispin, D.A.; Lai, L.A.; May, D.; McIntosh, M.W.; Goodlett, D.R.; Brentnall, T.A. Quantitative Glycoproteomics Analysis Reveals Changes in N-Glycosylation Level Associated with Pancreatic Ductal Adenocarcinoma. J. Proteome Res. 2014, 13, 1293–1306. [Google Scholar] [CrossRef]
- Sheehan, J.K.; Kirkham, S.; Howard, M.; Woodman, P.; Kutay, S.; Brazeau, C.; Buckley, J.; Thornton, D.J. Identification of Molecular Intermediates in the Assembly Pathway of the MUC5AC Mucin. J. Biol. Chem. 2004, 279, 15698–15705. [Google Scholar] [CrossRef]
- Luka, J.; Arlen, P.M.; Bristol, A. Development of a serum biomarker assay that differentiates tumor-associated MUC5AC (NPC-1C ANTIGEN) from normal MUC5AC. J. Biomed. Biotechnol. 2011, 2011, 934757. [Google Scholar] [CrossRef]
- Patel, S.P.; Bristol, A.; Saric, O.; Wang, X.-P.; Dubeykovskiy, A.; Arlen, P.M.; Morse, M.A. Anti-tumor activity of a novel monoclonal antibody, NPC-1C, optimized for recognition of tumor antigen MUC5AC variant in preclinical models. Cancer Immunol. Immunother. 2013, 62, 1011–1019. [Google Scholar] [CrossRef]
- Gold, D.V.; Hollingsworth, P.; Kremer, T.; Nelson, D. Identification of a human pancreatic duct tissue-specific antigen. Cancer Res. 1983, 43, 235–238. [Google Scholar]
- Gold, D.V.; Lew, K.; Maliniak, R.; Hernandez, M.; Cardillo, T. Characterization of monoclonal antibody PAM4 reactive with a pancreatic cancer mucin. Int. J. Cancer 1994, 57, 204–210. [Google Scholar] [CrossRef]
- Reis, C.A.; David, L.; Carvalho, F.; Mandel, U.; de Bolós, C.; Mirgorodskaya, E.; Clausen, H.; Sobrinho-Simões, M. Immunohistochemical Study of the Expression of MUC6 Mucin and Co-expression of Other Secreted Mucins (MUC5AC and MUC2) in Human Gastric Carcinomas. J. Histochem. Cytochem. 2000, 48, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Nollet, S.; Forgue-Lafitte, M.E.; Kirkham, P.; Bara, J. Mapping of two new epitopes on the apomucin encoded by MUC5AC gene: Expression in normal GI tract and colon tumors. Int. J. Cancer 2002, 99, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Chang, C.-H.; Gold, D.V.; Goldenberg, D.M. Identification of PAM4 (clivatuzumab)-reactive epitope on MUC5AC: A promising biomarker and therapeutic target for pancreatic cancer. Oncotarget 2015, 6, 4274–4285. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.E.; Bae, H.; Park, H.; Kuan, S.; Crawley, S.C.; Ho, J.J.; Kim, Y.S. Aberrant expression of MUC5AC and MUC6 gastric mucins and sialyl Tn antigen in intraepithelial neoplasms of the pancreas. Gastroenterology 2002, 123, 1052–1060. [Google Scholar] [CrossRef]
- Ho, J.J.; Crawley, S.; Pan, P.L.; Farrelly, E.R.; Kim, Y.S. Secretion of MUC5AC mucin from pancreatic cancer cells in response to forskolin and VIP. Biochem. Biophys. Res. Commun. 2002, 294, 680–686. [Google Scholar] [CrossRef]
- Yamazoe, S.; Tanaka, H.; Iwauchi, T.; Yoshii, M.; Ito, G.; Amano, R.; Yamada, N.; Sawada, T.; Ohira, M.; Hirakawa, K. Identification of HLA-A*0201- and A*2402-Restricted Epitopes of Mucin 5AC Expressed in Advanced Pancreatic Cancer. Pancreas 2011, 40, 896–904. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.J.; Bi, N.; Yan, P.S.; Yuan, M.; Norton, K.A.; Kim, Y.S. Characterization of new pancreatic cancer-reactive monoclonal antibodies directed against purified mucin. Cancer Res. 1991, 51, 372–380. [Google Scholar]
- Lidell, M.E.; Bara, J.; Hansson, G.C. Mapping of the 45M1 epitope to the C-terminal cysteine-rich part of the human MUC5AC mucin. FEBS J. 2008, 275, 481–489. [Google Scholar] [CrossRef]
- Bara, J.; Chastre, E.; Mahiou, J.; Singh, R.L.; Forgue-Lafitte, M.E.; Hollande, E.; Godeau, F. Gastric M1 mucin, an early oncofetal marker of colon carcinogenesis, is encoded by the MUC5AC gene. Int. J. Cancer 1998, 75, 767–773. [Google Scholar] [CrossRef]
- Inaguma, S.; Kasai, K.; Ikeda, H. GLI1 facilitates the migration and invasion of pancreatic cancer cells through MUC5AC-mediated attenuation of E-cadherin. Oncogene 2011, 30, 714–723. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, M.; Kondo, F.; Ishihara, T.; Yamaguchi, T.; Ito, R.; Tsuyuguchi, T.; Tawada, K.; Yokosuka, O. Evaluation of pancreatic intraepithelial neoplasia and mucin expression in normal pancreata. J. Hepato-Biliary-Pancreatic Sci. 2011, 19, 242–248. [Google Scholar] [CrossRef]
- Ohuchida, K.; Mizumoto, K.; Yamada, D.; Fujii, K.; Ishikawa, N.; Konomi, H.; Nagai, E.; Yamaguchi, K.; Tsuneyoshi, M.; Tanaka, M. Quantitative analysis of MUC1 and MUC5AC mRNA in pancreatic juice for preoperative diagnosis of pancreatic cancer. Int. J. Cancer 2006, 118, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Giorgadze, T.A.; Peterman, H.; Baloch, Z.W.; Furth, E.E.; Pasha, T.; Shiina, N.; Zhang, P.J.; Gupta, P.K. Diagnostic utility of mucin profile in fine-needle aspiration specimens of the pancreas. Cancer 2006, 108, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Higashi, M.; Yokoyama, S.; Yamamoto, T.; Goto, Y.; Kitazono, I.; Hiraki, T.; Taguchi, H.; Hashimoto, S.; Fukukura, Y.; Koriyama, C.; et al. Mucin Expression in Endoscopic Ultrasound-Guided Fine-Needle Aspiration Specimens Is a Useful Prognostic Factor in Pancreatic Ductal Adenocarcinoma. Pancreas 2015, 44, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Jinfeng, M.; Kimura, W.; Hirai, I.; Sakurai, F.; Moriya, T.; Mizutani, M. Expression of MUC5AC and MUC6 in invasive ductal carcinoma of the pancreas and relationship with prognosis. Int. J. Gastrointest. Cancer 2003, 34, 9–18. [Google Scholar] [CrossRef]
- Aloysius, M.M.; Zaitoun, A.M.; Awad, S.; Ilyas, M.; Rowlands, B.J.; Lobo, D.N. Mucins and CD56 as markers of tumour invasion and prognosis in periampullary cancer. Br. J. Surg. 2010, 97, 1269–1278. [Google Scholar] [CrossRef]
- Takano, Y.; Ohike, N.; Tajiri, T.; Asonuma, K.; Harada, K.; Takahashi, H.; Morohoshi, T. Gastric- and intestinal-type marker expression in invasive ductal adenocarcinoma of the pancreas. Hepatobiliary Pancreat. Dis. Int. 2012, 11, 424–428. [Google Scholar] [CrossRef]
- Yamazoe, S.; Tanaka, H.; Sawada, T.; Amano, R.; Yamada, N.; Ohira, M.; Hirakawa, K. RNA interference suppression of mucin 5AC (MUC5AC) reduces the adhesive and invasive capacity of human pancreatic cancer cells. J. Exp. Clin. Cancer Res. 2010, 29, 53. [Google Scholar] [CrossRef]
- Hoshi, H.; Sawada, T.; Uchida, M.; Iijima, H.; Kimura, K.; Hirakawa, K.; Wanibuchi, H. MUC5AC protects pancreatic cancer cells from TRAIL-induced death pathways. Int. J. Oncol. 2013, 42, 887–893. [Google Scholar] [CrossRef]
- Hoshi, H.; Sawada, T.; Uchida, M.; Saito, H.; Iijima, H.; Toda-Agetsuma, M.; Wada, T.; Yamazoe, S.; Tanaka, H.; Kimura, K.; et al. Tumor-associated MUC5AC stimulates in vivo tumorigenicity of human pancreatic cancer. Int. J. Oncol 2011, 38, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, K.; Krishn, S.R.; Jahan, R.; Atri, P.; Rachagani, S.; Rauth, S.; Xi, H.; Lu, Y.; Batra, S.; Kaur, S. Abstract 65: Gel-forming mucin MUC5AC as the nexus for cell-adhesion molecules governing pancreatic cancer aggressiveness and chemoresistance. Cancer Res. 2019, 79, 65. [Google Scholar] [CrossRef]
- Krishn, S.R.; Ganguly, K.; Kaur, S.; Batra, S.K. Ramifications of secreted mucin MUC5AC in malignant journey: A holistic view. Carcinogenesis 2018, 39, 633–651. [Google Scholar] [CrossRef]
- Vincent, A.; Perrais, M.; Desseyn, J.-L.; Aubert, J.-P.; Pigny, P.; Van Seuningen, I. Epigenetic regulation (DNA methylation, histone modifications) of the 11p15 mucin genes (MUC2, MUC5AC, MUC5B, MUC6) in epithelial cancer cells. Oncogene 2007, 26, 6566–6576. [Google Scholar] [CrossRef] [PubMed]
- Yamada, N.; Nishida, Y.; Yokoyama, S.; Tsutsumida, H.; Houjou, I.; Kitamoto, S.; Goto, M.; Higashi, M.; Yonezawa, S. Expression of MUC5AC, an early marker of pancreatobiliary cancer, is regulated by DNA methylation in the distal promoter region in cancer cells. J. Hepato-Biliary-Pancreatic Sci. 2010, 17, 844–854. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.J.; Han, S.W.; Pan, P.L.; Deng, G.; Kuan, S.-F.; Kim, Y.S. Methylation status of promoters and expression of MUC2 and MUC5AC mucins in pancreatic cancer cells. Int. J. Oncol. 2003, 22, 273–279. [Google Scholar] [CrossRef]
- Kato, S.; Hokari, R.; Crawley, S.; Gum, J.; Ahn, D.-H.; Kim, J.-W.; Kwon, S.-W.; Miura, S.; Basbaum, C.B.; Kim, Y.S. MUC5AC mucin gene regulation in pancreatic cancer cells. Int. J. Oncol. 2006, 29, 33–40. [Google Scholar] [CrossRef]
- di Magliano, M.P.; Sekine, S.; Ermilov, A.; Ferris, J.; Dlugosz, A.A.; Hebrok, M. Hedgehog/Ras interactions regulate early stages of pancreatic cancer. Genes Dev. 2006, 20, 3161–3173. [Google Scholar] [CrossRef]
- Mills, L.D.; Zhang, Y.; Marler, R.J.; Herreros-Villanueva, M.; Zhang, L.; Almada, L.L.; Couch, F.; Wetmore, C.; di Magliano, M.P.; Fernandez-Zapico, M.E. Loss of the Transcription Factor GLI1 Identifies a Signaling Network in the Tumor Microenvironment Mediating KRAS Oncogene-induced Transformation. J. Biol. Chem. 2013, 288, 11786–11794. [Google Scholar] [CrossRef]
- Kasai, K.; Inaguma, S.; Yoneyama, A.; Yoshikawa, K.; Ikeda, H. SCL/TAL1 Interrupting Locus Derepresses GLI1 from the Negative Control of Suppressor-of-Fused in Pancreatic Cancer Cell. Cancer Res. 2008, 68, 7723–7729. [Google Scholar] [CrossRef]
- Ho, J.J.; Farrelly, E.R.; Hong, J.; Pan, P.; Kim, Y.S. Forskolin increases the expression of the pancreatic tumor antigen, Nd2, and uptake of Nd2 antibody. Int. J. Oncol. 2000, 17, 881–888. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.J.L.; Farrelly, E.R.; Kim, Y.S. EGF enhances the effect of vasoactive intestinal hormone in elevating the level of the pancreatic tumor antigen ND2. Gastroenterology 1998, 114, A469. [Google Scholar] [CrossRef]
- Jiang, S.; Kopras, E.; McMichael, M.; Bell, R.H., Jr.; Ulrich, C.D., 2nd. Vasoactive intestinal peptide (VIP) stimulates in vitro growth of VIP-1 receptor-bearing human pancreatic adenocarcinoma-derived cells. Cancer Res. 1997, 57, 1475–1480. [Google Scholar] [PubMed]
- Zimmerman, N.P.; Roy, I.; Hauser, A.D.; Wilson, J.M.; Williams, C.L.; Dwinell, M.B. Cyclic AMP regulates the migration and invasion potential of human pancreatic cancer cells. Mol. Carcinog. 2013, 54, 203–215. [Google Scholar] [CrossRef]
- Tortora, G.; Ciardiello, F. Targeting of epidermal growth factor receptor and protein kinase A: Molecular basis and therapeutic applications. Ann. Oncol. 2000, 11, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Korc, M. Role of Growth Factors in Pancreatic Cancer. Surg. Oncol. Clin. N. Am. 1998, 7, 25–41. [Google Scholar] [CrossRef]
- Komatsu, H.; Tanji, E.; Sakata, N.; Aoki, T.; Motoi, F.; Naitoh, T.; Katayose, Y.; Egawa, S.; Unno, M.; Furukawa, T. A GNAS Mutation Found in Pancreatic Intraductal Papillary Mucinous Neoplasms Induces Drastic Alterations of Gene Expression Profiles with Upregulation of Mucin Genes. PLoS ONE 2014, 9, e87875. [Google Scholar] [CrossRef]
- Perrais, M.; Pigny, P.; Copin, M.-C.; Aubert, J.-P.; Van Seuningen, I. Induction of MUC2 and MUC5AC Mucins by Factors of the Epidermal Growth Factor (EGF) Family Is Mediated by EGF Receptor/Ras/Raf/Extracellular Signal-regulated Kinase Cascade and Sp1*. J. Biol. Chem. 2002, 277, 32258–32267. [Google Scholar] [CrossRef]
- Takeyama, K.; Jung, B.; Shim, J.J.; Burgel, P.-R.; Dao-Pick, T.; Ueki, I.F.; Protin, U.; Kroschel, P.; Nadel, J.A. Activation of epidermal growth factor receptors is responsible for mucin synthesis induced by cigarette smoke. Am. J. Physiol. Lung Cell. Mol. Physiol. 2001, 280, L165–L172. [Google Scholar] [CrossRef]
- Young, H.W.J.; Williams, O.W.; Chandra, D.; Bellinghausen, L.K.; Pérez, G.; Suárez, A.; Tuvim, M.J.; Roy, M.G.; Alexander, S.N.; Moghaddam, S.J.; et al. Central Role of Muc5ac Expression in Mucous Metaplasia and Its Regulation by Conserved 5′ Elements. Am. J. Respir. Cell Mol. Biol. 2007, 37, 273–290. [Google Scholar] [CrossRef]
- Zhong, T.; Perelman, J.M.; Kolosov, V.P.; Zhou, X.-D. MiR-146a negatively regulates neutrophil elastase-induced MUC5AC secretion from 16HBE human bronchial epithelial cells. Mol. Cell. Biochem. 2011, 358, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.; Zhang, R.; Liu, C.; Zhou, L.; Wang, H.; Zhuang, W.; Huang, Y.; Hong, Z. miR-143 inhibits interleukin-13-induced inflammatory cytokine and mucus production in nasal epithelial cells from allergic rhinitis patients by targeting IL13Rα1. Biochem. Biophys. Res. Commun. 2015, 457, 58–64. [Google Scholar] [CrossRef]
- Qin, Y.; Jiang, Y.; Sheikh, A.S.; Shen, S.; Liu, J.; Jiang, D. Interleukin-13 stimulates MUC5AC expression via a STAT6-TMEM16A-ERK1/2 pathway in human airway epithelial cells. Int. Immunopharmacol. 2016, 40, 106–114. [Google Scholar] [CrossRef]
- Thai, P.; Loukoianov, A.; Wachi, S.; Wu, R. Regulation of Airway Mucin Gene Expression. Annu. Rev. Physiol. 2008, 70, 405–429. [Google Scholar] [CrossRef]
- Lindén, S.; Mahdavi, J.; Hedenbro, J.; Borén, T.; Carlstedt, I. Effects of pH on Helicobacter pylori binding to human gastric mucins: Identification of binding to non-MUC5AC mucins. Biochem. J. 2004, 384, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Morinaga, Y.; Yanagihara, K.; Kaku, N.; Harada, Y.; Uno, N.; Nakamura, S.; Imamura, Y.; Hasegawa, H.; Miyazaki, T.; et al. Azithromycin inhibits MUC5AC induction via multidrug-resistant Acinetobacter baumannii in human airway epithelial cells. Pulm. Pharmacol. Ther. 2014, 28, 165–170. [Google Scholar] [CrossRef]
- Krishn, S.R. Secretory Mucin MUC5AC in Gastrointestinal Malignancies. Ph.D. Thesis, University of Nebraska Medical Center, Omaha, NE, USA, 2016. Available online: https://digitalcommons.unmc.edu/etd/110 (accessed on 17 April 2023).
- Kolb, A.; Kleeff, J.; Guweidhi, A.; Esposito, I.; Giese, N.; Adwan, H.; Giese, T.; Büchler, M.W.; Berger, M.; Friess, H. Osteopontin influences the invasiveness of pancreatic cancer cells and is increased in neoplastic and inflammatory conditions. Cancer Biol. Ther. 2005, 4, 740–746. [Google Scholar] [CrossRef]
- Kleeff, J.; Maruyama, H.; Ishiwata, T.; Sawhney, H.; Friess, H.; Büchler, M.W.; Korc, M. Bone morphogenetic protein 2 exerts diverse effects on cell growth in vitro and is expressed in human pancreatic cancer in vivo. Gastroenterology 1999, 116, 1202–1216. [Google Scholar] [CrossRef]
- Grzesiak, J.J.; Smith, K.C.; Burton, D.W.; Deftos, L.J.; Bouvet, M. Integrin-mediated laminin-1 adhesion upregulates CXCR4 and IL-8 expression in pancreatic cancer cells. Surgery 2007, 141, 804–814. [Google Scholar] [CrossRef] [PubMed]
- Grzesiak, J.J.; Bouvet, M. Determination of the Ligand-Binding Specificities of the α2β1 and α1β1 Integrins in a Novel 3-Dimensional In Vitro Model of Pancreatic Cancer. Pancreas 2007, 34, 220–228. [Google Scholar] [CrossRef]
- Michalski, C.W.; Erkan, M.; Sauliunaite, D.; Giese, T.; Stratmann, R.; Sartori, C.; Giese, N.A.; Friess, H.; Kleeff, J. Ex vivo chemosensitivity testing and gene expression profiling predict response towards adjuvant gemcitabine treatment in pancreatic cancer. Br. J. Cancer 2008, 99, 760–767. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, C.; Buttenschoen, K.; Straeter, J.; Henne-Bruns, D.; Kornmann, M. Pre-clinical evaluation of the activity of irinotecan as a basis for regional chemotherapy. Anticancer. Res. 2005, 25, 795–804. [Google Scholar] [PubMed]
- Deer, E.L.; González-Hernández, J.; Coursen, J.D.; Shea, J.E.; Ngatia, J.; Scaife, C.L.; Firpo, M.A.; Mulvihill, S.J. Phenotype and Genotype of Pancreatic Cancer Cell Lines. Pancreas 2010, 39, 425–435, Erratum in Pancreas 2018, 47, e37. [Google Scholar] [CrossRef] [PubMed]
- Sawai, H.; Yamamoto, M.; Okada, Y.; Sato, M.; Akamo, Y.; Takeyama, H.; Manabe, T. Alteration of Integrins by Interleukin-1α in Human Pancreatic Cancer Cells. Pancreas 2001, 23, 399–405. [Google Scholar] [CrossRef]
- Greco, E.; Basso, D.; Fogar, P.; Mazza, S.; Navaglia, F.; Zambon, C.-F.; Falda, A.; Pedrazzoli, S.; Ancona, E.; Plebani, M. Pancreatic Cancer Cells Invasiveness is Mainly Affected by Interleukin-1β not by Transforming Growth Factor-β1. Int. J. Biol. Markers 2005, 20, 235–241. [Google Scholar] [CrossRef]
- Funahashi, H.; Takeyama, H.; Sawai, H.; Furuta, A.; Sato, M.; Okada, Y.; Hayakawa, T.; Tanaka, M.; Manabe, T. Alteration of Integrin Expression by Glial Cell Line-Derived Neurotrophic Factor (GDNF) in Human Pancreatic Cancer Cells. Pancreas 2003, 27, 190–196. [Google Scholar] [CrossRef]
- Takada, M.; Nakamura, Y.; Koizumi, T.; Toyama, H.; Kamigaki, T.; Suzuki, Y.; Takeyama, Y.; Kuroda, Y. Suppression of Human Pancreatic Carcinoma Cell Growth and Invasion by Epigallocatechin-3-Gallate. Pancreas 2002, 25, 45–48. [Google Scholar] [CrossRef]
- Raut, C.P.; Nawrocki, S.; Lashinger, L.M.; Davis, D.W.; Khanbolooki, S.; Xiong, H.; Ellis, L.M.; McConkey, D.J. Celecoxib inhibits angiogenesis by inducing endothelial cell apoptosis in human pancreatic tumor xenografts. Cancer Biol. Ther. 2004, 3, 1217–1224. [Google Scholar] [CrossRef]
- Luo, J.; Guo, P.; Matsuda, K.; Truong, N.; Lee, A.; Chun, C.; Cheng, S.-Y.; Korc, M. Pancreatic cancer cell-derived vascular endothelial growth factor is biologically activein vitro and enhances tumorigenicityin vivo. Int. J. Cancer 2001, 92, 361–369. [Google Scholar] [CrossRef]
- Matsuo, Y.; Sawai, H.; Funahashi, H.; Takahashi, H.; Sakamoto, M.; Yamamoto, M.; Okada, Y.; Hayakawa, T.; Manabe, T. Enhanced Angiogenesis Due to Inflammatory Cytokines from Pancreatic Cancer Cell Lines and Relation to Metastatic Potential. Pancreas 2004, 28, 344–352. [Google Scholar] [CrossRef]
- Miknyoczki, S.J.; Chang, H.; Klein-Szanto, A.; Dionne, C.A.; Ruggeri, B.A. The Trk tyrosine kinase inhibitor CEP-701 (KT-5555) exhibits significant antitumor efficacy in preclinical xenograft models of human pancreatic ductal adenocarcinoma. Clin. Cancer Res. 1999, 5, 2205–2212. [Google Scholar]
- Sipos, B.; Möser, S.; Kalthoff, H.; Török, V.; Löhr, M.; Klöppel, G. A comprehensive characterization of pancreatic ductal carcinoma cell lines: Towards the establishment of an in vitro research platform. Virchows Archiv 2003, 442, 444–452. [Google Scholar] [CrossRef]
- Fujita, H.; Kurata, N.; Ohuchida, K.; Mizumoto, K.; Mahawithitwong, P.; Sakai, H.; Onimaru, M.; Manabe, T.; Ohtsuka, T.; Tanaka, M. Predicting the chemosensitivity of pancreatic cancer cells by quantifying the expression levels of genes associated with the metabolism of gemcitabine and 5-fluorouracil. Int. J. Oncol. 2011, 39, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Takagi, K.; Imura, J.; Shimomura, A.; Noguchi, A.; Minamisaka, T.; Tanaka, S.; Nishida, T.; Hatta, H.; Nakajima, T. Establishment of highly invasive pancreatic cancer cell lines and the expression of IL-32. Oncol. Lett. 2020, 20, 2888–2896. [Google Scholar] [CrossRef] [PubMed]
- Moore, S. A Comparison of Immunohistochemistry and Western Blot. News-Medical. 2020. Available online: https://www.news-medical.net/life-sciences/A-Comparison-of-Immunohistochemistry-and-Western-Blot.aspx (accessed on 17 April 2023).
- Kaur, S.; Smith, L.M.; Patel, A.; Menning, M.; Watley, D.C.; Malik, S.S.; Krishn, S.R.; Mallya, K.; Aithal, A.; Sasson, A.R.; et al. A Combination of MUC5AC and CA19-9 Improves the Diagnosis of Pancreatic Cancer: A Multicenter Study. Am. J. Gastroenterol. 2017, 112, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Huffman, B.M.; Mallick, A.B.; Horick, N.K.; Wang-Gillam, A.; Hosein, P.J.; Morse, M.A.; Beg, M.S.; Murphy, J.E.; Mavroukakis, S.; Zaki, A.; et al. Effect of a MUC5AC Antibody (NPC-1C) Administered with Second-Line Gemcitabine and Nab-Paclitaxel on the Survival of Patients With Advanced Pancreatic Ductal Adenocarcinoma. JAMA Netw. Open 2023, 6, e2249720. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Results |
|---|---|
| MUC5AC expression by IHC intumor-tissues | 80% of pancreatic cancer surgical specimens (positive for histology score > 0.01) Not detected in the normal tissue Localization was not reported |
| In pancreatic cell lines (PCL) | |
| MUC5AC-mRNA expression(by quantitative-polymerase chain reaction) | Higher in PCLs compared to normal pancreatic cell lines No expression in some PCLs (MIA PaCa, PANC10.05, QGP1) |
| MUC5AC protein expression(by western blot) | Positive in some PCL (ASPC1, BXPC3, COLO357, SW1990, and T3M4) Negative in some PCL (CD18/HPAF, CAPAN1, MIA PaCa, Panc-1). |
| Localization by confocalmicroscopy) | Stain the cytoplasm and intercellular junction (typical for mature MUC5AC) |
| In MUC5AC-knockdown PCL | |
| Decreased | Pancreatic cancer cell viability, anchorage-independent growth, cell motility, adhesion to the extracellular matrix, and angiogenesis Sensitivity to Gemcitabine (β-catenin mediated resistance) |
| Increased | Apoptosis |
| Nude mice with orthotopically transplanted MUC5AC knockdown PCL in pancreas * | Lesser pancreatic tumor weight Fewer metastatic sites |
| KrasG12D; Pdx1-Cre; Muc5ac-/- mouse models ** | Delay in onset and progression of pancreatic cancer cells |
| High | Medium | Low | No | |
|---|---|---|---|---|
| mRNA | COLO357 SW 1990 SU 8686 T3M4 | BXPC3 PANC1 AsPc3 | CAPAN | MIA PaCa |
| Protein | COLO357 SW 1990 | BXPC3 | AsPc3 T3M4 | PANC-1 CAPAN MIA PaCa |
| Features in CO Compared to M | Associated Features | |
|---|---|---|
| Prognostic factors [59,60,61,62] | Higher osteopontin | Invasiveness |
| Higher BMP2 | Poor survival | |
| Higher CXCR 4 (receptor and protein) | Cell survival, proliferation, migration, invasion, and metastasis. | |
| Predictive factors [63,64] | More sensitive to 5FU, Irinotecan, Cisplatin | |
| Less sensitive to Gem and Oxaliplatin |
| Cell Lines Compared | MUC5AC Expression | Results |
|---|---|---|
| M vs. BxPC3 | No vs. M (mRNA & protein) | BxPC3—More invasion, angiogenic potential, and tumorigenicity. More sensitive to Gem and 5FU |
| ASPC-1 vs. BxPC3 | L vs. M (mRNA & protein) | BxPC3 has -More invasion, angiogenic potential, and tumorigenicity. More resistant to 5FU and less-similar resistance to Gemcitabine |
| ASPC-1 vs. CO | L vs. H (mRNA & protein) | CO—less sensitive to 5FU and more sensitive to Gem |
| SU86.86 vs. Panc-1 | H vs. L (mRNA) | SU86.86 is less adhesive, more invasive and angiogenic potential |
| M vs. SW 1990 | L vs. H (mRNA & protein) | SW1990 is more resistant to Gem and more sensitive to 5FU (IC 50 of 9 vs. 5.68 μM) |
| CO > SU86686 > BxPC3 > M | H vs. H vs. M vs. No (P) H vs. M vs. M vs. No (mRNA) | Osteopontin influenced invasiveness |
| BxPC-3 > AsPC-1 or M | M vs. L vs. No (P and mRNA) | Tendency to invade—MPC has a minimum tendency to invade |
| Drug Tested | Sensitivity to the Drugs | |
|---|---|---|
| Protein Expression | mRNA Expression | |
| Gemcitabine | H < No < L-M | NDP |
| 5FU | NDP | NDP |
| Irinotecan | No < H | No < L-M < H |
| Oxaliplatin | L-M < H < No | H < No, NDP for L-M |
| Paclitaxel * | NDP | NDP |
| Cisplatin | No < H, no NDP for L-M | No < L-M < H |
| Nab-paclitaxel | NT | NT |
| Nanoliposomal irinotecan | NT | NT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manne, A.; Kasi, A.; Esnakula, A.K.; Paluri, R.K. Predictive Value of MUC5AC Signature in Pancreatic Ductal Adenocarcinoma: A Hypothesis Based on Preclinical Evidence. Int. J. Mol. Sci. 2023, 24, 8087. https://doi.org/10.3390/ijms24098087
Manne A, Kasi A, Esnakula AK, Paluri RK. Predictive Value of MUC5AC Signature in Pancreatic Ductal Adenocarcinoma: A Hypothesis Based on Preclinical Evidence. International Journal of Molecular Sciences. 2023; 24(9):8087. https://doi.org/10.3390/ijms24098087
Chicago/Turabian StyleManne, Ashish, Anup Kasi, Ashwini Kumar Esnakula, and Ravi Kumar Paluri. 2023. "Predictive Value of MUC5AC Signature in Pancreatic Ductal Adenocarcinoma: A Hypothesis Based on Preclinical Evidence" International Journal of Molecular Sciences 24, no. 9: 8087. https://doi.org/10.3390/ijms24098087