
Salt Stress—Regulation of Root Water Uptake in a Whole-Plant and Diurnal Context
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Plants and Salinity
1.1. Global Significance
1.2. The Major Stresses Associated with Salinity
1.2.1. Osmotic Stress
1.2.2. Ion Toxicity
1.2.3. Nutrient Imbalance
1.3. Salt Tolerance
- -
- Osmotically adjust cell water potential (ψ) so that the change in cell ψ (Δψ) equates to Δψ in the root medium; maintain cell turgor at the level observed for unstressed plants;
- -
- Tolerate xylem tensions, which increase by Δψ, without embolism;
- -
- Osmotically adjust through the use of Na+ and Cl− in the large central vacuole, and through use of alternative solutes in the cytoplasm (and nucleus), which are compatible with cell functions (‘compatible’ solutes);
- -
- Take up only as much Na+ and Cl− as required for osmotic adjustment in the vacuole;
- -
- Maintain leaf gas exchange and root water uptake at the level of non-stressed plants;
- -
- Maintain a growth rate so that growth and any seed production can be completed within the usual time frame;
- -
- Maintain the uptake of mineral nutrients, particularly K+, Ca2+, Mg2+ and NO3− at the level of non-stressed plants;
- -
- Do the above during day and night.
2. Night-Time Transpiration and Growth
2.1. Factors Which Affect Stomatal Conductance and Night-Time Transpiration
2.2. Why Do Plants Lose Water during the Night When No Carbon Can Be Gained?
2.3. Does Night-Time Transpirational Water Loss Benefit Plants under Salinity?
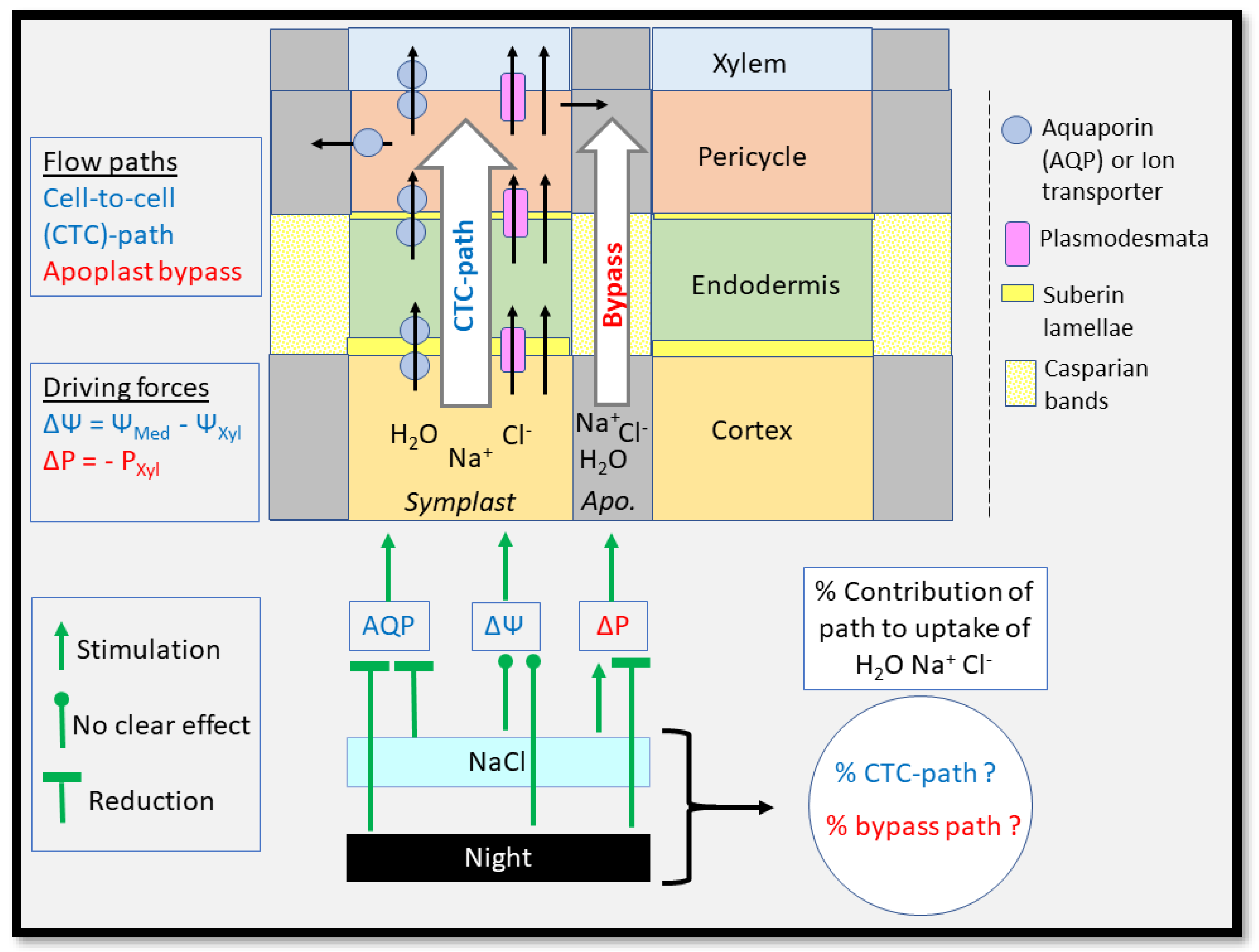
3. Root Water Uptake
3.1. Roots Are Modular, Diverse and Highly Dynamic
3.2. Root Water Uptake—Transport Paths and Driving Forces
3.3. Root Water Uptake—Hydraulic Conductance and Hydraulic Conductivity
3.4. Root Water Uptake—Regulation of the Cell-to-Cell Path Involving Aquaporins
3.5. Root Water Uptake—Water Flow through Plasmodesmata
3.6. Root Water Uptake—Water Flow along the Apoplast, and ‘Bypass’ Flow
4. Conclusions
- -
- There exists a large gap in our knowledge of the contribution of night-time-related processes, not just to the tolerance of plants to salt stress but to the functioning of plants in general.
- -
- We were surprised to see how little information there is on xylem tension during the night, for any plant species under any environmental conditions. Plants grow in size during the night and need to take up considerable amounts of water as well. They can also transpire significant quantities during the night through partly-open stomata. Surely, there must also be significant xylem tensions during the night, particularly in a low water-potential soil environment, in some species.
- -
- Maybe the largest unknown player in the regulation of the transportation of water and ions across tissues is the role of plasmodesmata. This is not so much because we do not think that they are important. It is more because we cannot study their transport properties in isolation, nor can we quantify the flow rate of water and ions through them. We tend to neglect aspects or assume that they play a minor role, as in the case of plasmodesmata, when we have no means to study them in detail. This renders the modelling of flows easier, but it does not make this modelling reflect more accurately the true situation in a plant.
- -
- It is tempting to search for and identify molecular stress tolerance mechanisms and use these for targeted breeding. However, unless these mechanisms save energy and carbon to plants, they are likely to fail. This applies more to the agricultural context, where the focus is on optimising yield; it does not have to apply to a natural, non-agronomic setting, where the emphasis is often on survival.
- -
- The endodermis is a barrier to the apoplastic (bypass) flows of water and ions across the root stele, from the cortex into the stele, and therefore, ultimately, root xylem and shoot. Is this really the entire story? Casparian bands constitute the main barrier to the radial movement of water and ions, but what about suberin lamellae? As the latter are located between the wall and plasma membrane of endodermal cells, they should also form a barrier to water and ions which exit the innermost cortex cell layer and want to enter the endodermal cell through a plasma membrane. Does that mean that the function of suberin lamellae could be more about forcing flow of water and ions between these tissue layers to occur through plasmodesmata; and to minimise the leakage of solutes out of endodermal cells through the plasma membrane into the cortex? We do not know. Could it maybe be that suberin lamellae provide an additional means to prevent the formation of air bubbles and embolism in the stelar apoplast including xylem vessels? One would not expect this based on the diameter of nanochannels in the wall and plasma membrane space, but we do not know for sure. Nor do we know how much of the tension in the xylem and stele transmits into a tension in the root cortex.
- -
- We predict here that the contribution of night-time-related processes to the tolerance of plants to salt stress and their productivity in general is far more important than we anticipate today. We also predict that this importance varies with species, soil water potential and the difference in temperature and relative humidity between day and night—sizes which are affected differentially by climate change.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Maas, E.V.; Hoffman, G.J. Crop Salt Tolerance—Current Assessment. J. Irrig. Drain. Div. 1977, 103, 115–134. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A.; Läuchli, A. Approaches to Increasing the Salt Tolerance of Wheat and Other Cereals. J. Exp. Bot. 2006, 57, 1025–1043. [Google Scholar] [CrossRef] [PubMed]
- Larcher, W. Physiological Plant Ecology: Ecophysiology and Stress Physiology of Functional Groups; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2003; ISBN 3-540-43516-6. [Google Scholar]
- Munns, R. Genes and Salt Tolerance: Bringing Them Together. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef] [PubMed]
- Carillo, P.; Grazia, M.; Pontecorvo, G.; Fuggi, A.; Woodrow, P. Salinity Stress and Salt Tolerance. In Abiotic Stress in Plants—Mechanisms and Adaptations; Shanker, A., Ed.; InTech: Rang-Du-Fliers, France, 2011; ISBN 978-953-307-394-1. [Google Scholar]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant Salinity Stress: Many Unanswered Questions Remain. Front. Plant Sci. 2019, 10, 80. [Google Scholar] [CrossRef]
- Xiao, F.; Zhou, H. Plant Salt Response: Perception, Signaling, and Tolerance. Front. Plant Sci. 2023, 13, 1053699. [Google Scholar] [CrossRef]
- Fricke, W.; Akhiyarova, G.; Wei, W.; Alexandersson, E.; Miller, A.; Kjellbom, P.O.; Richardson, A.; Wojciechowski, T.; Schreiber, L.; Veselov, D.; et al. The Short-Term Growth Response to Salt of the Developing Barley Leaf. J. Exp. Bot. 2006, 57, 1079–1095. [Google Scholar] [CrossRef]
- Rodríguez, H.G.; Roberts, M.; Jordan, W.R.; Drew, M.C. Growth, Water Relations, and Accumulation of Organic and Inorganic Solutes in Roots of Maize Seedlings during Salt Stress. Plant Physiol. 1997, 113, 881–893. [Google Scholar] [CrossRef]
- Fricke, W.; Peters, W.S. The Biophysics of Leaf Growth in Salt-Stressed Barley. A Study at the Cell Level. Plant Physiol. 2002, 129, 374–388. [Google Scholar] [CrossRef]
- Blum, A. Osmotic Adjustment Is a Prime Drought Stress Adaptive Engine in Support of Plant Production: Osmotic Adjustment and Plant Production. Plant Cell Environ. 2017, 40, 4–10. [Google Scholar] [CrossRef]
- Turner, N. Turgor Maintenance by Osmotic Adjustment—40 Years of Progress. J. Exp. Bot. 2018, 69, 3223–3233. [Google Scholar] [CrossRef] [PubMed]
- Venturas, M.D.; Sperry, J.S.; Hacke, U.G. Plant Xylem Hydraulics: What We Understand, Current Research, and Future Challenges. J. Integr. Plant Biol. 2017, 59, 356–389. [Google Scholar] [CrossRef] [PubMed]
- Flowers, T.J.; Hajibagherp, M.A.; Yeo, A.R. Ion Accumulation in the Cell Walls of Rice Plants Growing under Saline Conditions: Evidence for the Oertli Hypothesis. Plant Cell Environ. 1991, 14, 319–325. [Google Scholar] [CrossRef]
- Leigh, R.; Wyn Jones, R. Cellular Compartmentation in Plant Nutrition: The Selective Cytoplasm and the Promiscuous Vacuole. Adv. Plant Nutr. 1986, 2, 249–279. [Google Scholar]
- Leigh, R.A.; Tomos, A.D. Ion Distribution in Cereal Leaves: Pathways and Mechanisms. Phil. Trans. Royal Soc. London. Ser. B Biol. Sci. 1993, 341, 75–86. [Google Scholar]
- Brune, A.; Urbach, W.; Dietz, K.-J. Compartmentation and Transport of Zinc in Barley Primary Leaves as Basic Mechanisms Involved in Zinc Tolerance. Plant Cell Environ. 1994, 17, 153–162. [Google Scholar] [CrossRef]
- Leigh, R.A.; Storey, R. Intercellular Compartmentation of Ions in Barley Leaves in Relation to Potassium Nutrition and Salinity. J. Exp. Bot. 1993, 44, 755–762. [Google Scholar] [CrossRef]
- Dietz, K.-J.; Hollenbach, B.; Hellwege, E. The Epidermis of Barley Leaves Is a Dynamic Intermediary Storage Compartment of Carbohydrates, Amino Acids and Nitrate. Physiol. Plant. 1994, 92, 31–36. [Google Scholar] [CrossRef]
- Fricke, W.; Leigh, R.A.; Deri Tomos, A. Concentrations of Inorganic and Organic Solutes in Extracts from Individual Epidermal, Mesophyll and Bundle-Sheath Cells of Barley Leaves. Planta 1994, 192, 310–316. [Google Scholar] [CrossRef]
- Fricke, W.; Leigh, R.A.; Deri Tomos, A. Epidermal Solute Concentrations and Osmolality in Barley Leaves Studied at the Single-Cell Level: Changes along the Leaf Blade, during Leaf Ageing and NaCl Stress. Planta 1994, 192, 317–323. [Google Scholar] [CrossRef]
- Fricke, W.; Leigh, R.A.; Tomos, A.D. The Intercellular Distribution of Vacuolar Solutes in the Epidermis and Mesophyll of Barley Leaves Changes in Response to NaCI. J. Exp. Bot. 1996, 47, 1413–1426. [Google Scholar] [CrossRef]
- Conn, S.; Gilliham, M. Comparative Physiology of Elemental Distributions in Plants. Ann. Bot. 2010, 105, 1081–1102. [Google Scholar] [CrossRef] [PubMed]
- Garg, N.; Bhandari, P. Interactive Effects of Silicon and Arbuscular Mycorrhiza in Modulating Ascorbate-Glutathione Cycle and Antioxidant Scavenging Capacity in Differentially Salt-Tolerant Cicer arietinum L. Genotypes Subjected to Long-Term Salinity. Protoplasma 2016, 253, 1325–1345. [Google Scholar] [CrossRef] [PubMed]
- Ullah, S.M.; Gerzabek, M.H. Ion Uptake, Osmoregolation and Plant-Water Relations in Faba Beans (Vicia isb L.) under Salt Stress. Die Bodenkultur 1993, 44, 291–301. [Google Scholar]
- Nandwal, A.S.; Godara, M.; Kamboj, D.V.; Kundu, B.S.; Mann, A.; Kumar, B.; Sharma, S.K. Nodule Functioning in Trifoliate and Pentafoliate Mungbean Genotypes as Influenced by Salinity. Biol. Plant. 2000, 43, 459–462. [Google Scholar] [CrossRef]
- Sekeroglu, N.; Kara, M.; Dede, O.; Askin, T. Effect of Salinity on Germination, Early Seedling Growth, Na and K Constituents of Chickpea. Turk. J. Field Crops 1999, 4, e84. [Google Scholar]
- Maathuis, F. K+Nutrition and Na+Toxicity: The Basis of Cellular K+/Na+ Ratios. Ann. Bot. 1999, 84, 123–133. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity Tolerance in Halophytes*. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- Dubcovsky, J.; Maria, G.S.; Epstein, E. Mapping of the K+/Na+ Discrimination Locus Knal in Wheat. Theor. Appl. Genet. 1996, 2, 448–454. [Google Scholar] [CrossRef]
- Gorham, J.; Jones, R.G.W.; Bristol, A. Partial Characterization of the Trait for Enhanced K+−Na+ Discrimination in the D Genome of Wheat. Planta 1990, 180, 590–597. [Google Scholar] [CrossRef]
- Chen, Z.; Zhou, M.; Newman, I.A.; Mendham, N.J.; Zhang, G.; Shabala, S. Potassium and Sodium Relations in Salinised Barley Tissues as a Basis of Differential Salt Tolerance. Funct. Plant Biol. 2007, 34, 150. [Google Scholar] [CrossRef]
- Lin, H.X.; Zhu, M.Z.; Yano, M.; Gao, J.P.; Liang, Z.W.; Su, W.A.; Hu, X.H.; Ren, Z.H.; Chao, D.Y. QTLs for Na+ and K+ Uptake of the Shoots and Roots Controlling Rice Salt Tolerance. Theor. Appl. Genet. 2004, 108, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Belfield, E.J.; Cao, Y.; Smith, J.A.C.; Harberd, N.P. An Arabidopsis Soil-Salinity–Tolerance Mutation Confers Ethylene-Mediated Enhancement of Sodium/Potassium Homeostasis. Plant Cell 2013, 25, 3535–3552. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Lu, Y.; Wu, M.; Liang, E.; Li, Y.; Zhang, D.; Yin, Z.; Ren, X.; Dai, Y.; Deng, D.; et al. Ability to Remove Na+ and Retain K+ Correlates with Salt Tolerance in Two Maize Inbred Lines Seedlings. Front. Plant Sci. 2016, 7, 1716. [Google Scholar] [CrossRef] [PubMed]
- Ali, Z.; Park, H.C.; Ali, A.; Oh, D.-H.; Aman, R.; Kropornicka, A.; Hong, H.; Choi, W.; Chung, W.S.; Kim, W.-Y.; et al. TsHKT1;2, a HKT1 Homolog from the Extremophile Arabidopsis Relative Thellungiella salsuginea, Shows K+ Specificity in the Presence of NaCl. Plant Physiol. 2012, 158, 1463–1474. [Google Scholar] [CrossRef]
- Ardie, S.W.; Xie, L.; Takahashi, R.; Liu, S.; Takano, T. Cloning of a High-Affinity K+ Transporter Gene PutHKT2;1 from Puccinellia Tenuiflora and Its Functional Comparison with OsHKT2;1 from Rice in Yeast and Arabidopsis. J. Exp. Bot. 2009, 60, 3491–3502. [Google Scholar] [CrossRef]
- Rubio, F. TheHAKl Gene of Barley is a Member of a Large Gene Family and Encodes a High-Affinity Potassium Transporter. Plant Cell 1997, 9, 2281–2289. [Google Scholar]
- Bañuelos, M.A.; Garciadeblas, B.; Cubero, B.; Rodríguez-Navarro, A. Inventory and Functional Characterization of the HAK Potassium Transporters of Rice. Plant Physiol. 2002, 130, 784–795. [Google Scholar] [CrossRef]
- Frechilla, S.; Lasa, B.; Ibarretxe, L.; Lamsfus, C.; Aparicio-Tejo, P. Pea Responses to Saline Stress Is Affected by the Source of Nitrogen Nutrition (Ammonium or Nitrate). Plant Growth Reg. 2001, 35, 171–179. [Google Scholar] [CrossRef]
- Fricke, W.; Pritchard, J.; Tomos, D. Cells of the Upper and Lower Epidermis of Barley (Hordeum vulgare L.) Leaves Exhibit Distinct Patterns of Vacuolar Solutes. Planta 1994, 104, 1201–1208. [Google Scholar] [CrossRef]
- Bao, Z.; Bai, J.; Cui, H.; Gong, C. A Missing Link in Radial Ion Transport: Ion Transporters in the Endodermis. Front. Plant Sci. 2019, 10, 713. [Google Scholar] [CrossRef] [PubMed]
- Nagahashi, G.; Thomson, W.; Leonard, R. The Casparian Strip as a Barrier to the Movement of Lanthanum in Corn Roots. Science 1974, 183, 670–671. [Google Scholar] [CrossRef] [PubMed]
- Karahara, I.; Ikeda, A.; Kondo, T.; Uetake, Y. Development of the Casparian Strip in Primary Roots of Maize under Salt Stress. Planta 2004, 219, 41–47. [Google Scholar] [CrossRef]
- Shen, J.; Xu, G.; Zheng, H.Q. Apoplastic Barrier Development and Water Transport in Zea mays Seedling Roots under Salt and Osmotic Stresses. Protoplasma 2015, 252, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Hanson, A.D.; Rathinasabapathi, B.; Rivoal, J.; Burnet, M.; Dillon, M.O.; Gage, D.A. Osmoprotective Compounds in the Plumbaginaceae: A Natural Experiment in Metabolic Engineering of Stress Tolerance. Proc. Natl. Acad. Sci. USA 1994, 91, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Summers, P.S.; Nolte, K.D.; Cooper, A.J.L.; Borgeas, H.; Leustek, T.; Rhodes, D.; Hanson, A.D. Identification and Stereospecificity of the First Three Enzymes of 3-Dimethylsulfoniopropionate Biosynthesis in a Chlorophyte Alga1. Plant Physiol. 1998, 116, 369–378. [Google Scholar] [CrossRef]
- Hu, L.; Zhang, P.; Jiang, Y.; Fu, J. Metabolomic Analysis Revealed Differential Adaptation to Salinity and Alkalinity Stress in Kentucky bluegrass (Poa pratensis). Plant Mol. Biol. Rep. 2015, 33, 56–68. [Google Scholar] [CrossRef]
- Gorham, J.; Hughes, L.L.; Wyn Jones, R.G. Low-Molecular-Weight Carbohydrates in Some Salt-Stressed Plants. Physiol. Plant. 1981, 53, 27–33. [Google Scholar] [CrossRef]
- Gorham, J.; Hughes, L.L.; Wyn Jones, R.G. Chemical Composition of Salt-Marsh Plants from Ynys Môn (Anglesey): The Concept of Physiotypes. Plant Cell Environ. 1980, 3, 309–318. [Google Scholar] [CrossRef]
- Vernon, D.M.; Bohnert, H.J. Increased Expression of a Myo -Inositol Methyl Transferase in Mesembryanthemum Crystallinum Is Part of a Stress Response Distinct from Crassulacean Acid Metabolism Induction. Plant Physiol. 1992, 99, 1695–1698. [Google Scholar] [CrossRef]
- Ishitani, M.; Majumder, A.L.; Bornhouser, A.; Michalowski, C.B.; Jensen, R.G.; Bohnert, H.J. Coordinate Transcriptional Induction of Myo-Inositol Metabolism during Environmental Stress. Plant J. 1996, 9, 537–548. [Google Scholar] [CrossRef]
- Murakeözy, É.P.; Nagy, Z.; Duhazé, C.; Bouchereau, A.; Tuba, Z. Seasonal Changes in the Levels of Compatible Osmolytes in Three Halophytic Species of Inland Saline Vegetation in Hungary. J. Plant Physiol. 2003, 160, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Arndt, S.K.; Arampatsis, C.; Foetzki, A.; Li, X.; Zeng, F.; Zhang, X. Contrasting Patterns of Leaf Solute Accumulation and Salt Adaptation in Four Phreatophytic Desert Plants in a Hyperarid Desert with Saline Groundwater. J. Arid Environ. 2004, 59, 259–270. [Google Scholar] [CrossRef]
- Koyro, H.-W. Effect of Salinity on Growth, Photosynthesis, Water Relations and Solute Composition of the Potential Cash Crop Halophyte Plantago coronopus (L.). Environ. Exp. Bot. 2006, 56, 136–146. [Google Scholar] [CrossRef]
- Stoop, J.; Williamson, J.; Masonpharr, D. Mannitol Metabolism in Plants: A Method for Coping with Stress. Trends Plant Sci. 1996, 1, 139–144. [Google Scholar] [CrossRef]
- Noiraud, N.; Maurousset, L.; Lemoine, R. Transport of Polyols in Higher Plants. Plant Physiol. Biochem. 2001, 39, 717–728. [Google Scholar] [CrossRef]
- Farquharson, K.L. Targeted Overexpression of a Sodium Transporter in the Root Stele Increases Salinity Tolerance. Plant Cell 2009, 21, 1875. [Google Scholar] [CrossRef]
- James, R.A.; Davenport, R.J.; Munns, R. Physiological Characterization of Two Genes for Na+ Exclusion in Durum Wheat, Nax1 and Nax2. Plant Physiol. 2006, 142, 1537–1547. [Google Scholar] [CrossRef]
- James, R.A.; Blake, C.; Byrt, C.S.; Munns, R. Major Genes for Na+ Exclusion, Nax1 and Nax2 (Wheat HKT1;4 and HKT1;5), Decrease Na+ Accumulation in Bread Wheat Leaves under Saline and Waterlogged Conditions. J. Exp. Bot. 2011, 62, 2939–2947. [Google Scholar] [CrossRef]
- Byrt, C.S.; Platten, J.D.; Spielmeyer, W.; James, R.A.; Lagudah, E.S.; Dennis, E.S.; Tester, M.; Munns, R. HKT1;5-Like Cation Transporters Linked to Na+ Exclusion Loci in Wheat, Nax2 and Kna1. Plant Physiol. 2007, 143, 1918–1928. [Google Scholar] [CrossRef]
- Byrt, C.S.; Xu, B.; Krishnan, M.; Lightfoot, D.J.; Athman, A.; Jacobs, A.K.; Watson-Haigh, N.S.; Plett, D.; Munns, R.; Tester, M.; et al. The Na+ Transporter, TaHKT1;5-D, Limits Shoot Na+ Accumulation in Bread Wheat. Plant J. 2014, 80, 516–526. [Google Scholar] [CrossRef] [PubMed]
- Mäser, P.; Eckelman, B.; Vaidyanathan, R.; Horie, T.; Fairbairn, D.J.; Kubo, M.; Yamagami, M.; Yamaguchi, K.; Nishimura, M.; Uozumi, N.; et al. Altered Shoot/Root Na+ Distribution and Bifurcating Salt Sensitivity in Arabidopsis by Genetic Disruption of the Na+ Transporter AtHKT1. FEBS Lett. 2002, 531, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Sunarpi; Horie, T.; Motoda, J.; Kubo, M.; Yang, H.; Yoda, K.; Horie, R.; Chan, W.-Y.; Leung, H.-Y.; Hattori, K.; et al. Enhanced Salt Tolerance Mediated by AtHKT1 Transporter-Induced Na+ Unloading from Xylem Vessels to Xylem Parenchyma Cells: Na+ Unloading from Xylem by AtHKT1. Plant J. 2005, 44, 928–938. [Google Scholar] [CrossRef] [PubMed]
- Møller, I.S.; Gilliham, M.; Jha, D.; Mayo, G.M.; Roy, S.J.; Coates, J.C.; Haseloff, J.; Tester, M. Shoot Na+ Exclusion and Increased Salinity Tolerance Engineered by Cell Type–Specific Alteration of Na+ Transport in Arabidopsis. Plant Cell 2009, 21, 2163–2178. [Google Scholar] [CrossRef]
- Shi, H.; Quintero, F.J.; Pardo, J.M.; Zhu, J.-K. The Putative Plasma Membrane Na+/H+ Antiporter SOS1 Controls Long-Distance Na+ Transport in Plants. Plant Cell 2002, 14, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Parida, A.K.; Jha, B. Salt Tolerance Mechanisms in Mangroves: A Review. Trees 2010, 24, 199–217. [Google Scholar] [CrossRef]
- Thomson, W.W. The structure and function of salt glands. In Plants in Saline Environments; Poljakoff-Mayber, A., Gale, J., Eds.; Ecological Studies; Springer: Berlin/Heidelberg, Germany, 1975; Volume 15, pp. 118–146. ISBN 978-3-642-80931-6. [Google Scholar]
- Dassanayake, M.; Larkin, J.C. Making Plants Break a Sweat: The Structure, Function, and Evolution of Plant Salt Glands. Front. Plant Sci. 2017, 8, 406. [Google Scholar] [CrossRef]
- McElrone, A.J.; Pockman, W.T.; Martínez-Vilalta, J.; Jackson, R.B. Variation in Xylem Structure and Function in Stems and Roots of Trees to 20 m Depth. New Phytol. 2004, 163, 507–517. [Google Scholar] [CrossRef]
- Munns, R.; Day, D.A.; Fricke, W.; Watt, M.; Arsova, B.; Barkla, B.J.; Bose, J.; Byrt, C.S.; Chen, Z.; Foster, K.J.; et al. Energy Costs of Salt Tolerance in Crop Plants. New Phytol. 2020, 225, 1072–1090. [Google Scholar] [CrossRef]
- Tyerman, S.D.; Munns, R.; Fricke, W.; Arsova, B.; Barkla, B.J.; Bose, J.; Bramley, H.; Byrt, C.; Chen, Z.; Colmer, T.D.; et al. Energy Costs of Salinity Tolerance in Crop Plants. New Phytol. 2019, 221, 25–29. [Google Scholar] [CrossRef]
- Gambetta, G.A.; Knipfer, T.; Fricke, W.; McElrone, A.J. Aquaporins and root water uptake. In Plant Aquaporins; Chaumont, F., Tyerman, S.D., Eds.; Signaling and Communication in Plants; Springer International Publishing: Cham, Switzerland, 2017; pp. 133–153. ISBN 978-3-319-49393-0. [Google Scholar]
- Fricke, W. Night-Time Transpiration—Favouring Growth? Trends Plant Sci. 2019, 24, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Fricke, W. Energy Costs of Salinity Tolerance in Crop Plants: Night-time Transpiration and Growth. New Phytol. 2020, 225, 1152–1165. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Jeffers, R.; Raju, A.; Kenny, T.; Ratchanniyasamu, E.; Fricke, W. Does Night-time Transpiration Provide Any Benefit to Wheat (Triticum aestivum L.) Plants Which Are Exposed to Salt Stress? Physiol. Plant. 2023, 175, e13839. [Google Scholar] [CrossRef] [PubMed]
- Walter, A.; Schurr, U. Dynamics of Leaf and Root Growth: Endogenous Control versus Environmental Impact. Ann. Bot. 2005, 95, 891–900. [Google Scholar] [CrossRef] [PubMed]
- Stitt, M.; Zeeman, S.C. Starch Turnover: Pathways, Regulation and Role in Growth. Curr. Opin. Plant Biol. 2012, 15, 282–292. [Google Scholar] [CrossRef]
- Smith, A.M.; Stitt, M. Coordination of Carbon Supply and Plant Growth. Plant Cell Environ. 2007, 30, 1126–1149. [Google Scholar] [CrossRef] [PubMed]
- Sonnewald, U.; Fernie, A.R. Next-Generation Strategies for Understanding and Influencing Source–Sink Relations in Crop Plants. Curr. Opin. Plant Biol. 2018, 43, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Turgeon, R. The Sink-Source Transition in Leaves. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 119–138. [Google Scholar] [CrossRef]
- Imlau, A.; Truernit, E.; Sauer, N. Cell-to-Cell and Long-Distance Trafficking of the Green Fluorescent Protein in the Phloem and Symplastic Unloading of the Protein into Sink Tissues. Plant Cell 1999, 11, 309–322. [Google Scholar] [CrossRef]
- Graf, A.; Schlereth, A.; Stitt, M.; Smith, A.M. Circadian Control of Carbohydrate Availability for Growth in Arabidopsis Plants at Night. Proc. Natl. Acad. Sci. USA 2010, 107, 9458–9463. [Google Scholar] [CrossRef]
- Weraduwage, S.M.; Chen, J.; Anozie, F.C.; Morales, A.; Weise, S.E.; Sharkey, T.D. The Relationship between Leaf Area Growth and Biomass Accumulation in Arabidopsis thaliana. Front. Plant Sci. 2015, 6, 167. [Google Scholar] [CrossRef] [PubMed]
- Pantin, F.; Simonneau, T.; Rolland, G.; Dauzat, M.; Muller, B. Control of Leaf Expansion: A Developmental Switch from Metabolics to Hydraulics. Plant Physiol. 2011, 156, 803–815. [Google Scholar] [CrossRef] [PubMed]
- Brouquisse, R.; James, F.; Raymond, P.; Pradet, A. Study of Glucose Starvation in Excised Maize Root Tips. Plant Physiol. 1991, 96, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Caird, M.A.; Richards, J.H.; Donovan, L.A. Nighttime Stomatal Conductance and Transpiration in C3 and C4 Plants. Plant Physiol. 2007, 143, 4–10. [Google Scholar] [CrossRef]
- Dawson, T.E.; Burgess, S.S.O.; Tu, K.P.; Oliveira, R.S.; Santiago, L.S.; Fisher, J.B.; Simonin, K.A.; Ambrose, A.R. Nighttime Transpiration in Woody Plants from Contrasting Ecosystems. Tree Physiol. 2007, 27, 561–575. [Google Scholar] [CrossRef]
- Wallace, J.; McJannet, D. Climate Change Impacts on the Water Balance of Coastal and Montane Rainforests in Northern Queensland, Australia. J. Hydrol. 2012, 475, 84–96. [Google Scholar] [CrossRef]
- Rosado, B.H.P.; Oliveira, R.S.; Joly, C.A.; Aidar, M.P.M.; Burgess, S.S.O. Diversity in Nighttime Transpiration Behavior of Woody Species of the Atlantic Rain Forest, Brazil. Agric. For. Meteorol. 2012, 158–159, 13–20. [Google Scholar] [CrossRef]
- Alvarado-Barrientos, M.; Asbjornsen, H.; Holwerda, F. Is Nighttime Transpiration Enhanced after Fog Events? In Proceedings of the IX International Workshop on Sap Flow 991, Gent, Belgium, 28 May 2013; pp. 133–139. [Google Scholar]
- Zeppel, M.J.B.; Lewis, J.D.; Phillips, N.G.; Tissue, D.T. Consequences of Nocturnal Water Loss: A Synthesis of Regulating Factors and Implications for Capacitance, Embolism and Use in Models. Tree Physiol. 2014, 34, 1047–1055. [Google Scholar] [CrossRef]
- Barbeta, A.; Ogaya, R.; Peñuelas, J. Comparative Study of Diurnal and Nocturnal Sap Flow of Quercus Ilex and Phillyrea Latifolia in a Mediterranean Holm Oak Forest in Prades (Catalonia, NE Spain). Trees 2012, 26, 1651–1659. [Google Scholar] [CrossRef]
- Mitchell, P.J.; Veneklaas, E.; Lambers, H.; Burgess, S.S.O. Partitioning of Evapotranspiration in a Semi-Arid Eucalypt Woodland in South-Western Australia. Agric. For. Meteorol. 2009, 149, 25–37. [Google Scholar] [CrossRef]
- Pfautsch, S.; Keitel, C.; Turnbull, T.L.; Braimbridge, M.J.; Wright, T.E.; Simpson, R.R.; O’Brien, J.A.; Adams, M.A. Diurnal Patterns of Water Use in Eucalyptus Victrix Indicate Pronounced Desiccation-Rehydration Cycles despite Unlimited Water Supply. Tree Physiol. 2011, 31, 1041–1051. [Google Scholar] [CrossRef] [PubMed]
- Ogle, K.; Lucas, R.W.; Bentley, L.P.; Cable, J.M.; Barron-Gafford, G.A.; Griffith, A.; Ignace, D.; Jenerette, G.D.; Tyler, A.; Huxman, T.E.; et al. Differential Daytime and Night-time Stomatal Behavior in Plants from North American Deserts. New Phytol. 2012, 194, 464–476. [Google Scholar] [CrossRef] [PubMed]
- Benyon, R.G.; Marcar, N.E.; Crawford, D.F.; Nicholson, A.T. Growth and Water Use of Eucalyptus Camaldulensis and E. Occidentalis on a Saline Discharge Site near Wellington, NSW, Australia. Agric. Water Manag. 1999, 39, 229–244. [Google Scholar] [CrossRef]
- Phillips, N.G.; Lewis, J.D.; Logan, B.A.; Tissue, D.T. Inter- and Intra-Specific Variation in Nocturnal Water Transport in Eucalyptus. Tree Physiol. 2010, 30, 586–596. [Google Scholar] [CrossRef] [PubMed]
- Resco de Dios, V.; Díaz-Sierra, R.; Goulden, M.L.; Barton, C.V.M.; Boer, M.M.; Gessler, A.; Ferrio, J.P.; Pfautsch, S.; Tissue, D.T. Woody Clockworks: Circadian Regulation of Night-time Water Use in E Ucalyptus Globulus. New Phytol. 2013, 200, 743–752. [Google Scholar] [CrossRef]
- Pantin, F.; Monnet, F.; Jannaud, D.; Costa, J.M.; Renaud, J.; Muller, B.; Simonneau, T.; Genty, B. The Dual Effect of Abscisic Acid on Stomata. New Phytol. 2013, 197, 65–72. [Google Scholar] [CrossRef]
- Oren, R.; Sperry, J.S.; Katul, G.G.; Pataki, D.E.; Ewers, B.E.; Phillips, N.; Schäfer, K.V.R. Survey and Synthesis of Intra- and Interspecific Variation in Stomatal Sensitivity to Vapour Pressure Deficit: Intra- and Interspecific Variation in Stomatal Sensitivity to Vapour Pressure Deficit. Plant Cell Environ. 1999, 22, 1515–1526. [Google Scholar] [CrossRef]
- Howard, A.R.; Donovan, L.A. Soil Nitrogen Limitation Does Not Impact Nighttime Water Loss in Populus. Tree Physiol. 2010, 30, 23–31. [Google Scholar] [CrossRef]
- Furukawa, A.; Park, S.-Y.; Fujinuma, Y. Hybrid Poplar Stomata Unresponsive to Changes in Environmental Conditions. Trees 1990, 4, 191–197. [Google Scholar] [CrossRef]
- Muchow, R.; Ludlow, M.; Fisher, M.; Myers, R. Stomatal Behaviour of Kenaf and Sorghum in a Semiarid Tropical Environment. I. During the Night. Funct. Plant Biol. 1980, 7, 609–619. [Google Scholar] [CrossRef]
- Zeppel, M.J.B.; Lewis, J.D.; Chaszar, B.; Smith, R.A.; Medlyn, B.E.; Huxman, T.E.; Tissue, D.T. Nocturnal Stomatal Conductance Responses to Rising [CO2], Temperature and Drought. New Phytol. 2012, 193, 929–938. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, R. Effects of CO2 on Stomatal Conductance: Do Stomata Open at Very High CO2 Concentrations? Ann. Bot. 1999, 83, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Resco de Dios, V.; Loik, M.E.; Smith, R.; Aspinwall, M.J.; Tissue, D.T. Genetic Variation in Circadian Regulation of Nocturnal Stomatal Conductance Enhances Carbon Assimilation and Growth: On the Function of Nocturnal Stomatal Conductance. Plant Cell Environ. 2016, 39, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.M.; Monnet, F.; Jannaud, D.; Leonhardt, N.; Ksas, B.; Reiter, I.M.; Pantin, F.; Genty, B. Open all night long: The Dark Side of Stomatal Control. Plant Physiol. 2015, 167, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Rawson, H.; Clarke, J. Nocturnal Transpiration in Wheat. Funct. Plant Biol. 1988, 15, 397. [Google Scholar] [CrossRef]
- Howard, A.R.; Donovan, L.A. Helianthus Nighttime Conductance and Transpiration Respond to Soil Water but Not Nutrient Availability. Plant Physiol. 2007, 143, 145–155. [Google Scholar] [CrossRef]
- Leymarie, J.; Lasceve, G.; Vavasseur, A. Elevated CO2 Enhances Stomatal Responses to Osmotic Stress and Abscisic Acid in Arabidopsis thaliana. Plant Cell Environ. 1999, 22, 301–308. [Google Scholar] [CrossRef]
- Leymarie, J.; Lascève, G.; Vavasseur, A. Interaction of Stomatal Responses to ABA and CO2 in Arabidopsis thaliana. Funct. Plant Biol. 1998, 25, 785. [Google Scholar] [CrossRef]
- Blake, J.; Ferrell, W.K. The Association between Soil and Xylem Water Potential, Leaf Resistance, and Abscisic Acid Content in Droughted Seedlings of Douglas-Fir (Pseudotsuga menziesii). Physiol. Plant. 1977, 39, 106–109. [Google Scholar] [CrossRef]
- Ludwig, F.; Jewitt, R.A.; Donovan, L.A. Nutrient and Water Addition Effects on Day- and Night-Time Conductance and Transpiration in a C3 Desert Annual. Oecologia 2006, 148, 219–225. [Google Scholar] [CrossRef]
- Donovan, L.A.; Richards, J.H.; Linton, M.J. Magnitude and Mechanisms of Disequlibrium between Predawn Plant and Soil Water Potentials. Ecology 2003, 84, 463–470. [Google Scholar] [CrossRef]
- Scholz, F.G.; Bucci, S.J.; Goldstein, G.; Meinzer, F.C.; Franco, A.C.; Miralles-Wilhelm, F. Removal of Nutrient Limitations by Long-Term Fertilization Decreases Nocturnal Water Loss in Savanna Trees. Tree Physiol. 2007, 27, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Dodd, A.N.; Salathia, N.; Hall, A.; Kévei, E.; Tóth, R.; Nagy, F.; Hibberd, J.M.; Millar, A.J.; Webb, A.A.R. Plant Circadian Clocks Increase Photosynthesis, Growth, Survival, and Competitive Advantage. Science 2005, 309, 630–633. [Google Scholar] [CrossRef] [PubMed]
- Dodd, A.N.; Parkinson, K.; Webb, A.A.R. Independent Circadian Regulation of Assimilation and Stomatal Conductance in the Ztl-1 Mutant of Arabidopsis. New Phytol. 2004, 162, 63–70. [Google Scholar] [CrossRef]
- Lasceve, G.; Leymarie, J.; Vavasseur, A. Alterations in Light-Induced Stomatal Opening in a Starch-Deficient Mutant of Arabidopsis thaliana L. Deficient in Chloroplast Phosphoglucomutase Activity. Plant Cell Environ. 1997, 20, 350–358. [Google Scholar] [CrossRef]
- Barbour, M.M.; Cernusak, L.A.; Whitehead, D.; Griffin, K.L.; Turnbull, M.H.; Tissue, D.T.; Farquhar, G.D. Nocturnal Stomatal Conductance and Implications for Modelling Δ18O of Leaf-Respired CO2 in Temperate Tree Species. Funct. Plant Biol. 2005, 32, 1107. [Google Scholar] [CrossRef]
- Easlon, H.M.; Richards, J.H. Photosynthesis Affects Following Night Leaf Conductance in Vicia Faba. Plant Cell Environ. 2009, 32, 58–63. [Google Scholar] [CrossRef]
- Qin, H.; Arteaga, C.; Chowdhury, F.I.; Granda, E.; Yao, Y.; Han, Y.; Resco de Dios, V. Radiation and Drought Impact Residual Leaf Conductance in Two Oak Species with Implications for Water Use Models. Front. Plant Sci. 2020, 11, 603581. [Google Scholar] [CrossRef]
- Kim, H.-H. Stomatal Conductance of Lettuce Grown Under or Exposed to Different Light Qualities. Ann. Bot. 2004, 94, 691–697. [Google Scholar] [CrossRef]
- De Dios, V.R.; Turnbull, M.H.; Barbour, M.M.; Ontedhu, J.; Ghannoum, O.; Tissue, D.T. Soil Phosphorous and Endogenous Rhythms Exert a Larger Impact than CO2 or Temperature on Nocturnal Stomatal Conductance in Eucalyptus Tereticornis. Tree Physiol. 2013, 33, 1206–1215. [Google Scholar] [CrossRef]
- Running, S.W. Environmental Control of Leaf Water Conductance in Conifers. Can. J. For. Res. 1976, 6, 104–112. [Google Scholar] [CrossRef]
- Christman, M.A.; James, J.J.; Drenovsky, R.E.; Richards, J.H. Environmental Stress and Genetics Influence Night-Time Leaf Conductance in the C4 Grass Distichlis Spicata. Funct. Plant Biol. 2009, 36, 50. [Google Scholar] [CrossRef] [PubMed]
- Resco de Dios, V.; Mereed, T.E.; Ferrio, J.P.; Tissue, D.T.; Voltas, J. Intraspecific Variation in Juvenile Tree Growth under Elevated CO2 Alone and with O3: A Meta-Analysis. Tree Physiol. 2016, 36, 682–693. [Google Scholar] [CrossRef] [PubMed]
- Cernusak, L.A.; Farquhar, G.D.; Wong, S.C.; Stuart-Williams, H. Measurement and Interpretation of the Oxygen Isotope Composition of Carbon Dioxide Respired by Leaves in the Dark. Plant Physiol. 2004, 136, 3350–3363. [Google Scholar] [CrossRef]
- Daley, M.J.; Phillips, N.G. Interspecific Variation in Nighttime Transpiration and Stomatal Conductance in a Mixed New England Deciduous Forest. Tree Physiol. 2006, 26, 411–419. [Google Scholar] [CrossRef]
- Marks, C.O.; Lechowicz, M.J. The Ecological and Functional Correlates of Nocturnal Transpiration. Tree Physiol. 2007, 27, 577–584. [Google Scholar] [CrossRef]
- Snyder, K.A.; James, J.J.; Richards, J.H.; Donovan, L.A. Does Hydraulic Lift or Nighttime Transpiration Facilitate Nitrogen Acquisition? Plant Soil 2008, 306, 159–166. [Google Scholar] [CrossRef]
- Howard, A.R.; Van Iersel, M.W.; Richards, J.H.; Donovan, L.A. Night-Time Transpiration Can Decrease Hydraulic Redistribution. Plant Cell Environ. 2009, 32, 1060–1070. [Google Scholar] [CrossRef]
- Even, M.; Sabo, M.; Meng, D.; Kreszies, T.; Schreiber, L.; Fricke, W. Night-Time Transpiration in Barley (Hordeum vulgare) Facilitates Respiratory Carbon Dioxide Release and Is Regulated during Salt Stress. Ann. Bot. 2018, 122, 569–582. [Google Scholar] [CrossRef]
- Dayer, S.; Herrera, J.C.; Dai, Z.; Burlett, R.; Lamarque, L.J.; Delzon, S.; Bortolami, G.; Cochard, H.; Gambetta, G.A. Nighttime Transpiration Represents a Negligible Part of Water Loss and Does Not Increase the Risk of Water Stress in Grapevine. Plant Cell Environ. 2021, 44, 387–398. [Google Scholar] [CrossRef]
- Zeppel, M.; Tissue, D.; Taylor, D.; Macinnis-Ng, C.; Eamus, D. Rates of Nocturnal Transpiration in Two Evergreen Temperate Woodland Species with Differing Water-Use Strategies. Tree Physiol. 2010, 30, 988–1000. [Google Scholar] [CrossRef] [PubMed]
- Faiyue, B.; Al-Azzawi, M.J.; Flowers, T.J. The Role of Lateral Roots in Bypass Flow in Rice (Oryza sativa L.). Plant Cell Environ. 2010, 33, 702–716. [Google Scholar] [CrossRef]
- Steudle, E.; Peterson, C.A. How Does Water Get through Roots? J. Exp. Bot. 1998, 49, 775–788. [Google Scholar] [CrossRef]
- Yamauchi, T.; Shimamura, S.; Nakazono, M.; Mochizuki, T. Aerenchyma Formation in Crop Species: A Review. Field Crops Research 2013, 152, 8–16. [Google Scholar] [CrossRef]
- Hose, E.; Clarkson, D.T.; Steudle, E.; Schreiber, L.; Hartung, W. The Exodermis: A Variable Apoplastic Barrier. J. Exp. Bot. 2001, 52, 2245–2264. [Google Scholar] [CrossRef] [PubMed]
- Doblas, V.G.; Geldner, N.; Barberon, M. The Endodermis, a Tightly Controlled Barrier for Nutrients. Curr. Opin. Plant Biol. 2017, 39, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Mergemann, H.; Sauter, M. Ethylene Induces Epidermal Cell Death at the Site of Adventitious Root Emergence in Rice. Plant Physiol. 2000, 124, 609–614. [Google Scholar] [CrossRef]
- Schneider, H.M.; Lynch, J.P. Functional Implications of Root Cortical Senescence for Soil Resource Capture. Plant Soil 2018, 423, 13–26. [Google Scholar] [CrossRef]
- Ahmed, M.A.; Zarebanadkouki, M.; Meunier, F.; Javaux, M.; Kaestner, A.; Carminati, A. Root Type Matters: Measurement of Water Uptake by Seminal, Crown, and Lateral Roots in Maize. J. Exp. Bot. 2018, 69, 1199–1206. [Google Scholar] [CrossRef]
- De Smet, I.; White, P.J.; Bengough, A.G.; Dupuy, L.; Parizot, B.; Casimiro, I.; Heidstra, R.; Laskowski, M.; Lepetit, M.; Hochholdinger, F.; et al. Analyzing Lateral Root Development: How to Move Forward. Plant Cell 2012, 24, 15–20. [Google Scholar] [CrossRef]
- Soper, K. Root Anatomy of Grasses and Clovers: Lolium perenne L., Dactylis glomerata L., Glyceria fluitans Br., Alopecurus pratensis L., Trifolium repens L., T. pratense L., and Lotus uliginosus Schkuhr (Major Sm.). New Zealand J. Agric. Res. 1959, 2, 329–341. [Google Scholar] [CrossRef]
- Maurel, C.; Nacry, P. Root Architecture and Hydraulics Converge for Acclimation to Changing Water Availability. Nat. Plants 2020, 6, 744–749. [Google Scholar] [CrossRef] [PubMed]
- Karlova, R.; Boer, D.; Hayes, S.; Testerink, C. Root Plasticity under Abiotic Stress. Plant Physiol. 2021, 187, 1057–1070. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Eissenstat, D.M. Linking Hydraulic Conductivity to Anatomy in Plants That Vary in Specific Root Length. Am. Soc. Hort. Sci. 2000, 125, 260–264. [Google Scholar] [CrossRef]
- Steudle, E.; Oren, R.; Schulze, E.-D. Water Transport in Maize Roots: Measurement of Hydraulic Conductivity, Solute Permeability, and of Reflection Coefficients of Excised Roots Using the Root Pressure Probe. Plant Physiol. 1987, 84, 1220–1232. [Google Scholar] [CrossRef]
- Steudle, E. Water uptake by plant roots: An integration of views. In Recent Advances of Plant Root Structure and Function; Gašparíková, O., Čiamporová, M., Mistrík, I., Baluška, F., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2001; pp. 71–82. ISBN 978-90-481-5598-9. [Google Scholar]
- Schreiber, L.; Hartmann, K.; Skrabs, M. Apoplastic Barriers in Roots: Chemical Composition of Endodermal and Hypodermal Cell Walls. J. Exp. Bot. 1999, 50, 1267–1280. [Google Scholar] [CrossRef]
- Ranathunge, K.; Schreiber, L. Water and Solute Permeabilities of Arabidopsis Roots in Relation to the Amount and Composition of Aliphatic Suberin. J. Exp. Bot. 2011, 62, 1961–1974. [Google Scholar] [CrossRef]
- Lu, Y.; Fricke, W. Changes in Root Hydraulic Conductivity in Wheat (Triticum aestivum L.) in Response to Salt Stress and Day/Night Can Best Be Explained through Altered Activity of Aquaporins. Plant Cell Environ. 2023, 46, 747–763. [Google Scholar] [CrossRef]
- Frensch, J.; Steudle, E. Axial and Radial Hydraulic Resistance to Roots of Maize (Zea mays L.). Plant Physiol. 1989, 91, 719–726. [Google Scholar] [CrossRef]
- Zimmermann, U.; Schneider, H.; Wegner, L.H.; Haase, A. Water Ascent in Tall Trees: Does Evolution of Land Plants Rely on a Highly Metastable State? New Phytol. 2004, 162, 575–615. [Google Scholar] [CrossRef]
- Eissenstat, D.M. Trade-Offs in Root Form and Function. In Ecology in Agriculture; Elsevier: Amsterdam, The Netherlands, 1997; pp. 173–199. ISBN 978-0-12-378260-1. [Google Scholar]
- Bramley, H.; Turner, N.C.; Turner, D.W.; Tyerman, S.D. Roles of Morphology, Anatomy, and Aquaporins in Determining Contrasting Hydraulic Behavior of Roots. Plant Physiol. 2009, 150, 348–364. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.X.; Ranathunge, K.; Lee, S.; Lee, Y.; Lee, D.; Sung, J. Composite Transport Model and Water and Solute Transport across Plant Roots: An Update. Front. Plant Sci. 2018, 9, 193. [Google Scholar] [CrossRef] [PubMed]
- Knipfer, T.; Besse, M.; Verdeil, J.-L.; Fricke, W. Aquaporin-facilitated Water Uptake in Barley (Hordeum vulgare L.) Roots. J. Exp. Bot 2011, 62, 4115–4126. [Google Scholar] [CrossRef] [PubMed]
- Hachez, C.; Zelazny, E.; Chaumont, F. Modulating the Expression of Aquaporin Genes in Planta: A Key to Understand Their Physiological Functions? Biochim. Biophys. Acta—Biomembr. 2006, 1758, 1142–1156. [Google Scholar] [CrossRef]
- Hachez, C.; Veljanovski, V.; Reinhardt, H.; Guillaumot, D.; Vanhee, C.; Chaumont, F.; Batoko, H. The Arabidopsis Abiotic Stress-Induced TSPO-Related Protein Reduces Cell-Surface Expression of the Aquaporin PIP2;7 through Protein-Protein Interactions and Autophagic Degradation. Plant Cell 2014, 26, 4974–4990. [Google Scholar] [CrossRef]
- Boursiac, Y.; Boudet, J.; Postaire, O.; Luu, D.-T.; Tournaire-Roux, C.; Maurel, C. Stimulus-Induced Downregulation of Root Water Transport Involves Reactive Oxygen Species-Activated Cell Signalling and Plasma Membrane Intrinsic Protein Internalization. Plant J. 2008, 56, 207–218. [Google Scholar] [CrossRef]
- Boursiac, Y.; Chen, S.; Luu, D.-T.; Sorieul, M.; van den Dries, N.; Maurel, C. Early Effects of Salinity on Water Transport in Arabidopsis Roots. Molecular and Cellular Features of Aquaporin Expression. Plant Physiol. 2005, 139, 790–805. [Google Scholar] [CrossRef]
- Meng, D.; Walsh, M.; Fricke, W. Rapid Changes in Root Hydraulic Conductivity and Aquaporin Expression in Rice (Oryza sativa L.) in Response to Shoot Removal—Xylem Tension as a Possible Signal. Ann. Bot. 2016, 118, 809–819. [Google Scholar] [CrossRef]
- Vandeleur, R.K.; Mayo, G.; Shelden, M.C.; Gilliham, M.; Kaiser, B.N.; Tyerman, S.D. The Role of Plasma Membrane Intrinsic Protein Aquaporins in Water Transport through Roots: Diurnal and Drought Stress Responses Reveal Different Strategies between Isohydric and Anisohydric Cultivars of Grapevine. Plant Physiol. 2009, 149, 445–460. [Google Scholar] [CrossRef]
- Sakurai-Ishikawa, J.; Murai-Hatano, M.; Hayashi, H.; Ahamed, A.; Fukushi, K.; Matsumoto, T.; Kitagawa, Y. Transpiration from Shoots Triggers Diurnal Changes in Root Aquaporin Expression: Transpiration Triggers Root Aquaporin Expression. Plant Cell Environ. 2011, 34, 1150–1163. [Google Scholar] [CrossRef]
- Calvo-Polanco, M.; Sánchez-Romera, B.; Aroca, R. Mild Salt Stress Conditions Induce Different Responses in Root Hydraulic Conductivity of Phaseolus Vulgaris Over-Time. PLoS ONE 2014, 9, e90631. [Google Scholar] [CrossRef] [PubMed]
- Knipfer, T.; Danjou, M.; Vionne, C.; Fricke, W. Salt Stress Reduces Root Water Uptake in Barley (Hordeum vulgare L.) through Modification of the Transcellular Transport Path. Plant Cell Environ. 2021, 44, 458–475. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.; Fricke, W. Changes in Root Hydraulic Conductivity Facilitate the Overall Hydraulic Response of Rice (Oryza sativa L.) Cultivars to Salt and Osmotic Stress. Plant Physiol. Biochem. 2017, 113, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Knipfer, T.; Fricke, W. Root aquaporins. In Root Engineering: Basic and Applied Concepts; Morte, A., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 269–296. ISBN 978-3-642-54276-3. [Google Scholar]
- Rutschow, H.L.; Baskin, T.I.; Kramer, E.M. Regulation of Solute Flux through Plasmodesmata in the Root Meristem. Plant Physiol. 2011, 155, 1817–1826. [Google Scholar] [CrossRef] [PubMed]
- Cheval, C.; Samwald, S.; Johnston, M.G.; de Keijzer, J.; Breakspear, A.; Liu, X.; Bellandi, A.; Kadota, Y.; Zipfel, C.; Faulkner, C. Chitin Perception in Plasmodesmata Characterizes Submembrane Immune-Signaling Specificity in Plants. Proc. Natl. Acad. Sci. USA 2020, 117, 9621–9629. [Google Scholar] [CrossRef]
- Maule, A.J.; Benitez-Alfonso, Y.; Faulkner, C. Plasmodesmata—Membrane Tunnels with Attitude. Curr. Opin. Plant Biol. 2011, 14, 683–690. [Google Scholar] [CrossRef]
- Peters, W.S.; Jensen, K.H.; Stone, H.A.; Knoblauch, M. Plasmodesmata and the Problems with Size: Interpreting the Confusion. J. Plant Physiol. 2021, 257, 153341. [Google Scholar] [CrossRef]
- Robards, A.W.; Clarkson, D.T. The role of plasmodesmata in the transport of water and nutrients across roots. In Intercellular Communication in Plants: Studies on Plasmodesmata; Gunning, B.E.S., Robards, A.W., Eds.; Springer: Berlin/Heidelberg, Germany, 1976; pp. 181–201. ISBN 978-3-642-66296-6. [Google Scholar]
- Lee, D.-K.; Sieburth, L.E. Plasmodesmata Formation: Poking Holes in Walls with Ise. Curre. Biol. 2010, 20, R488–R490. [Google Scholar] [CrossRef]
- Itaya, A.; Woo, Y.-M.; Masuta, C.; Bao, Y.; Nelson, R.S.; Ding, B. Developmental Regulation of Intercellular Protein Trafficking through Plasmodesmata in Tobacco Leaf Epidermis. Plant Physiol. 1998, 118, 373–385. [Google Scholar] [CrossRef]
- Burch-Smith, T.M.; Stonebloom, S.; Xu, M.; Zambryski, P.C. Plasmodesmata during Development: Re-Examination of the Importance of Primary, Secondary, and Branched Plasmodesmata Structure versus Function. Protoplasma 2011, 248, 61–74. [Google Scholar] [CrossRef]
- Faulkner, C. Plasmodesmata and the Symplast. Curr. Biol. 2018, 28, R1374–R1378. [Google Scholar] [CrossRef] [PubMed]
- Duckett, C.M.; Oparka, K.J.; Prior, D.A.M.; Dolan, L.; Roberts, K. Dye-Coupling in the Root Epidermis of Arabidopsis Is Progressively Reduced during Development. Development 1994, 120, 3247–3255. [Google Scholar] [CrossRef]
- Ma, F.; Peterson, C.A. Frequencies of Plasmodesmata in Allium cepa L. Roots: Implications for Solute Transport Pathways. J. Exp. Bot. 2001, 52, 1051–1061. [Google Scholar] [CrossRef] [PubMed]
- Sevilem, I.; Miyashima, S.; Helariutta, Y. Cell-to-Cell Communication via Plasmodesmata in Vascular Plants. Cell Adhes. Migr. 2013, 7, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Sager, R.E.; Lee, J.-Y. Plasmodesmata at a Glance. J. Cell Sci. 2018, 131, jcs209346. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sager, R.; Cui, W.; Zhang, C.; Lu, H.; Lee, J.-Y. Salicylic Acid Regulates Plasmodesmata Closure during Innate Immune Responses in Arabidopsis. Plant Cell 2013, 25, 2315–2329. [Google Scholar] [CrossRef] [PubMed]
- Knox, J.P.; Benitez-Alfonso, Y. Roles and Regulation of Plant Cell Walls Surrounding Plasmodesmata. Curr. Opin. Plant Biol. 2014, 22, 93–100. [Google Scholar] [CrossRef]
- Schulz, A. Plasmodesmal Widening Accompanies the Short-Term Increase in Symplasmic Phloem Unloading in Pea Root Tips under Osmotic Stress. Protoplasma 1995, 188, 22–37. [Google Scholar] [CrossRef]
- Hukin, D.; Doering-Saad, C.; Thomas, C.R.; Pritchard, J. Sensitivity of Cell Hydraulic Conductivity to Mercury Is Coincident with Symplasmic Isolation and Expression of Plasmalemma Aquaporin Genes in Growing Maize Roots. Planta 2002, 215, 1047–1056. [Google Scholar] [CrossRef]
- Barberon, M. The Endodermis as a Checkpoint for Nutrients. New Phytol. 2017, 213, 1604–1610. [Google Scholar] [CrossRef]
- Taleisnik, E. Water Retention Capacity in Root Segments Differing in the Degree of Exodermis Development. Ann. Bot. 1999, 83, 19–27. [Google Scholar] [CrossRef]
- Rincon, C.A.; Raper, C.D., Jr.; Patterson, R.P. Genotypic Differences in Root Anatomy Affecting Water Movement through Roots of Soybean. Int. J. Plant Sci. 2003, 164, 543–551. [Google Scholar] [CrossRef]
- Sutka, M.; Li, G.; Boudet, J.; Boursiac, Y.; Doumas, P.; Maurel, C. Natural Variation of Root Hydraulics in Arabidopsis Grown in Normal and Salt-Stressed Conditions. Plant Physiol. 2011, 155, 1264–1276. [Google Scholar] [CrossRef] [PubMed]
- van Zelm, E.; Zhang, Y.; Testerink, C. Salt Tolerance Mechanisms of Plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [PubMed]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity Induced Physiological and Biochemical Changes in Plants: An Omic Approach towards Salt Stress Tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef]
- Yıldırım, K.; Yağcı, A.; Sucu, S.; Tunç, S. Responses of Grapevine Rootstocks to Drought through Altered Root System Architecture and Root Transcriptomic Regulations. Plant Physiol. Biochem. 2018, 127, 256–268. [Google Scholar] [CrossRef]
- Kreszies, T.; Shellakkutti, N.; Osthoff, A.; Yu, P.; Baldauf, J.A.; Zeisler-Diehl, V.V.; Ranathunge, K.; Hochholdinger, F.; Schreiber, L. Osmotic Stress Enhances Suberization of Apoplastic Barriers in Barley Seminal Roots: Analysis of Chemical, Transcriptomic and Physiological Responses. New Phytol. 2019, 221, 180–194. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, Y.; Zhong, J.; Zhang, T.; Li, D.; Ba, T.; Xu, T.; Chang, L.; Zhang, Q.; Sun, M. Root Physiological Traits and Transcriptome Analyses Reveal That Root Zone Water Retention Confers Drought Tolerance to Opisthopappus Taihangensis. Sci Rep. 2020, 10, 2627. [Google Scholar] [CrossRef]
- Henry, A.; Cal, A.J.; Batoto, T.C.; Torres, R.O.; Serraj, R. Root Attributes Affecting Water Uptake of Rice (Oryza sativa) under Drought. J. Exp. Bot. 2012, 63, 4751–4763. [Google Scholar] [CrossRef]
- Barberon, M.; Vermeer, J.E.M.; De Bellis, D.; Wang, P.; Naseer, S.; Andersen, T.G.; Humbel, B.M.; Nawrath, C.; Takano, J.; Salt, D.E.; et al. Adaptation of Root Function by Nutrient-Induced Plasticity of Endodermal Differentiation. Cell 2016, 164, 447–459. [Google Scholar] [CrossRef]
- Xu, C.; Fu, X.; Liu, R.; Guo, L.; Ran, L.; Li, C.; Tian, Q.; Jiao, B.; Wang, B.; Luo, K. PtoMYB170 Positively Regulates Lignin Deposition during Wood Formation in Poplar and Confers Drought Tolerance in Transgenic Arabidopsis. Tree Physiol. 2017, 37, 1713–1726. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Luo, L.; Zheng, L. Lignins: Biosynthesis and Biological Functions in Plants. IJMS 2018, 19, 335. [Google Scholar] [CrossRef] [PubMed]
- Kosma, D.K.; Murmu, J.; Razeq, F.M.; Santos, P.; Bourgault, R.; Molina, I.; Rowland, O. At MYB 41 Activates Ectopic Suberin Synthesis and Assembly in Multiple Plant Species and Cell Types. Plant J. 2014, 80, 216–229. [Google Scholar] [CrossRef]
- Yamauchi, T.; Noshita, K.; Tsutsumi, N. Climate-Smart Crops: Key Root Anatomical Traits That Confer Flooding Tolerance. Breed. Sci. 2021, 71, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Faiyue, B.; Al-Azzawi, M.J.; Flowers, T.J. A New Screening Technique for Salinity Resistance in Rice (Oryza sativa L.) Seedlings Using Bypass Flow: Bypass Flow and Screening for Salt Tolerance. Plant Cell Environ. 2012, 35, 1099–1108. [Google Scholar] [CrossRef]
- Flam-Shepherd, R.; Huynh, W.Q.; Coskun, D.; Hamam, A.M.; Britto, D.T.; Kronzucker, H.J. Membrane Fluxes, Bypass Flows, and Sodium Stress in Rice: The Influence of Silicon. J. Exp. Bot. 2018, 69, 1679–1692. [Google Scholar] [CrossRef]
- Emonet, A.; Hay, A. Development and Diversity of Lignin Patterns. Plant Physiol. 2022, 190, 31–43. [Google Scholar] [CrossRef]
- Kumar, P.; Choudhary, M.; Halder, T.; Prakash, N.R.; Singh, V.; Vineeth, T.V.; Sheoran, S.; Ravikiran, K.T.; Longmei, N.; Rakshit, S.; et al. Salinity Stress Tolerance and Omics Approaches: Revisiting the Progress and Achievements in Major Cereal Crops. Heredity 2022, 128, 497–518. [Google Scholar] [CrossRef]
- Knipfer, T.; Fricke, W. Water Uptake by Seminal and Adventitious Roots in Relation to Whole-Plant Water Flow in Barley (Hordeum vulgare L.). J. Exp. Bot. 2011, 62, 717–733. [Google Scholar] [CrossRef]
- Kreszies, T.; Schreiber, L.; Ranathunge, K. Suberized Transport Barriers in Arabidopsis, Barley and Rice Roots: From the Model Plant to Crop Species. J. Plant Physiol. 2018, 227, 75–83. [Google Scholar] [CrossRef]
- Garcia, A.; Rizzo, C.A.; Ud-Din, J.; Bartos, S.L.; Senadhira, D.; Flowers, T.J.; Yeo, A.R. Sodium and Potassium Transport to the Xylem Are Inherited Independently in Rice, and the Mechanism of Sodium: Potassium Selectivity Differs between Rice and Wheat. Plant Cell Environ. 1997, 20, 1167–1174. [Google Scholar] [CrossRef]
- Faiyue, B.; Vijayalakshmi, C.; Nawaz, S.; Nagato, Y.; Taketa, S.; Ichii, M.; Al-Azzawi, M.J.; Flowers, T.J. Studies on Sodium Bypass Flow in Lateral Rootless Mutants Lrt1 and Lrt2, and Crown Rootless Mutant Crl1 of Rice (Oryza sativa L.). Plant Cell Environ. 2010, 33, 687–701. [Google Scholar] [CrossRef] [PubMed]
- Chaumont, F.; Tyerman, S.D. Aquaporins: Highly Regulated Channels Controlling Plant Water Relations. Plant Physiol. 2014, 164, 1600–1618. [Google Scholar] [CrossRef] [PubMed]
- Tyerman, S.D.; Skerrett, M.; Garrill, A.; Findlay, G.P.; Leigh, R.A. Pathways for the Permeation of Na + and CI- into Protoplasts Derived from the Cortex of Wheat Roots. J. Exp. Bot. 1997, 48, 459–480. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Y.; Fricke, W. Salt Stress—Regulation of Root Water Uptake in a Whole-Plant and Diurnal Context. Int. J. Mol. Sci. 2023, 24, 8070. https://doi.org/10.3390/ijms24098070
Lu Y, Fricke W. Salt Stress—Regulation of Root Water Uptake in a Whole-Plant and Diurnal Context. International Journal of Molecular Sciences. 2023; 24(9):8070. https://doi.org/10.3390/ijms24098070
Chicago/Turabian StyleLu, Yingying, and Wieland Fricke. 2023. "Salt Stress—Regulation of Root Water Uptake in a Whole-Plant and Diurnal Context" International Journal of Molecular Sciences 24, no. 9: 8070. https://doi.org/10.3390/ijms24098070