
Holoprosencephaly with a Special Form of Anophthalmia Result from Experimental Induction of bmp4, Oversaturating BMP Antagonists in Zebrafish
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
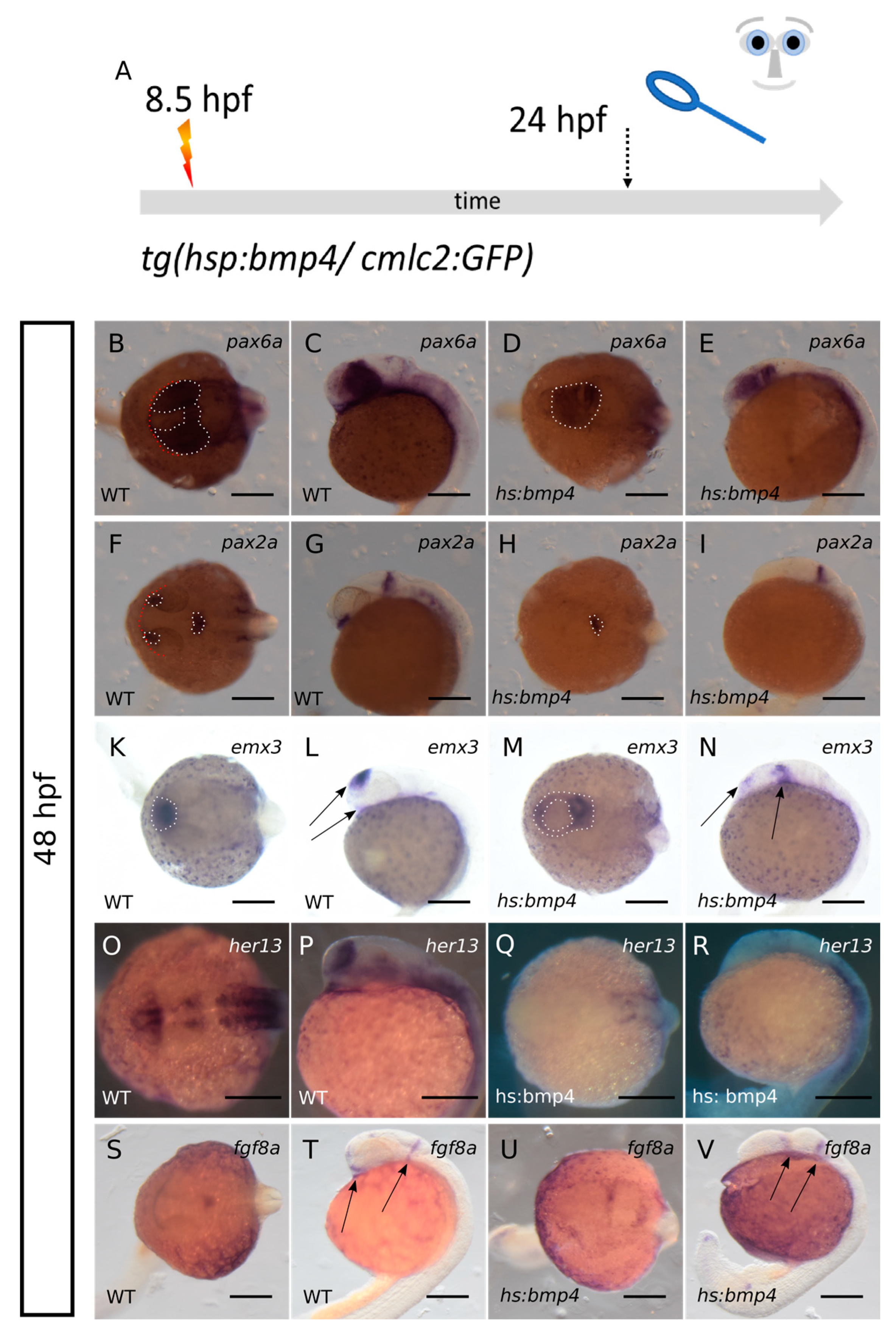
2.1. Bmp4 Induction at 8.5 hpf Results in HPE and a “Crypt-Oculoid”
2.2. ANP Development Is Severely Hampered after bmp4 Induction at 8.5 hpf
2.3. Bmp4 Induction Results in Loss of rx3 Expression but Sustained Expression of rx2
2.4. Bmp4 Induction Alters Expression of zic2a, shha/b, alcamb and cxcr4a
2.5. Bmp4 Induction Hampers Hypothalamic Subduction during Neurulation and Neural Keel Formation
3. Discussion
4. Materials and Methods
4.1. Zebrafish Care
4.1.1. Heat Shock Procedures
4.1.2. Laser Scanning Confocal Microscopy
4.1.3. Image Processing
4.1.4. In Situ hybridization
- Emx3 F
- 5′-GAAGTGCTTCACGATTGAATC-3′; R 5′-TGAAATGACGTCAATGTCCTC-3′
- Fgf8a F:
- 5′-GACTCATACCTTCACGGTTGAG-3′; R: 5′-TGCGTTTAGTCCGTCTGTTG-3′
- Foxg1a F:
- 5′-ATGTTGGATATGGGAGAAAG-3′; R: 5′-AAGAAATAACTGGTCTGACC-3′
- Nog2: For:
- 5′-ATGGGCAGCATCACCCG-3′ Rev: 5′- TCAGCACGAGCACTTGCA-3′
- Chrd: For:
- 5′-TTGTATGGCAGCAGGCGTAT-3′ Rev: 5′-TTGTATGGCAGCAGGCGTAT-3′
- Her13 F:
- 5′-CCACGCTGCTGAACTTAGAAA-3′; R: 5′-TCATCCAGGTCAGAGCAGAGA-3′
- Pax6a F:
- 5′-AGATGGTTGCCAACAGTCAG-3′; R: 5′-GGGACATGTCTGGTTCACTG-3′
- Rx2 F:
- 5′-GCCTCTCCACAGAAAGCTAC-3′; R: 5′-CGATACTAGAACTGCGGTCG-3′
- Rx3 F:
- 5′-ATGAGGCTTGTTGGATCTCAG-3′; R: 5′-ATGAGGCTTGTTGGATCTCAG-3′
- Six3b F:
- 5′-TTTGGTCGTTGCCCGTAGCACC-3′; R: 5′-CATCGAAATCAGAGTCACTGTC-3′
- Zic2a F:
- 5′-ATTAAGCAAGAGCTCATCTG-3′; R: 5′-AACTGTGGACCGCTGAGGAAG-3′
4.1.5. CRISPR/Cas9 F0 Analysis (Crispants)
- rx3 T1:
- CCCGGCGTTTCCATATGGAT
- rx3 T2:
- TGAACGTGGTTCGGTTCCGC
- rx3 T3:
- CTTCGAGAAGTCGCACTATC
- rx3 T4:
- GAGATGGGGCCGGTCAACCA
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rembold, M.; Loosli, F.; Adams, R.J.; Wittbrodt, J. Individual Cell Migration Serves as the Driving Force for Optic Vesicle Evagination. Science 2006, 313, 1130–1134. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.E.; Keller, P.J.; Ramialison, M.; Rembold, M.; Stelzer, E.H.K.; Loosli, F.; Wittbrodt, J. Nlcam Modulates Midline Convergence during Anterior Neural Plate Morphogenesis. Dev. Biol. 2010, 339, 14–25. [Google Scholar] [CrossRef]
- Bielen, H.; Houart, C. BMP Signaling Protects Telencephalic Fate by Repressing Eye Identity and Its Cxcr4-Dependent Morphogenesis. Dev. Cell 2012, 23, 812–822. [Google Scholar] [CrossRef] [PubMed]
- England, S.J.; Blanchard, G.B.; Mahadevan, L.; Adams, R.J. A Dynamic Fate Map of the Forebrain Shows How Vertebrate Eyes Form and Explains Two Causes of Cyclopia. Development 2006, 133, 4613–4617. [Google Scholar] [CrossRef]
- Fallet-Bianco, C. Neuropathology of Holoprosencephaly. Am. J. Med. Genet. C Semin. Med. Genet. 2018, 178, 214–228. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.; Litingtung, Y.; Lee, E.; Young, K.E.; Corden, J.L.; Westphal, H.; Beachy, P.A. Cyclopia and Defective Axial Patterning in Mice Lacking Sonic Hedgehog Gene Function. Nature 1996, 383, 407–413. [Google Scholar] [CrossRef]
- Roessler, E.; Belloni, E.; Gaudenz, K.; Jay, P.; Berta, P.; Scherer, S.W.; Tsui, L.C.; Muenke, M. Mutations in the Human Sonic Hedgehog Gene Cause Holoprosencephaly. Nat. Genet. 1996, 14, 357–360. [Google Scholar] [CrossRef]
- Houart, C.; Caneparo, L.; Heisenberg, C.; Barth, K.; Take-Uchi, M.; Wilson, S. Establishment of the Telencephalon during Gastrulation by Local Antagonism of Wnt Signaling. Neuron 2002, 35, 255–265. [Google Scholar] [CrossRef]
- Cavodeassi, F. Integration of Anterior Neural Plate Patterning and Morphogenesis by the Wnt Signaling Pathway. Dev. Neurobiol. 2014, 74, 759–771. [Google Scholar] [CrossRef]
- Bachiller, D.; Klingensmith, J.; Kemp, C.; Belo, J.A.; Anderson, R.M.; May, S.R.; McMahon, J.A.; McMahon, A.P.; Harland, R.M.; Rossant, J.; et al. The Organizer Factors Chordin and Noggin Are Required for Mouse Forebrain Development. Nature 2000, 403, 658–661. [Google Scholar] [CrossRef] [PubMed]
- Khokha, M.K.; Yeh, J.; Grammer, T.C.; Harland, R.M. Depletion of Three BMP Antagonists from Spemann’s Organizer Leads to a Catastrophic Loss of Dorsal Structures. Dev. Cell 2005, 8, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Eckert, P.; Knickmeyer, M.D.; Schütz, L.; Wittbrodt, J.; Heermann, S. Morphogenesis and Axis Specification Occur in Parallel during Optic Cup and Optic Fissure Formation, Differentially Modulated by BMP and Wnt. Open. Biol. 2019, 9, 180179. [Google Scholar] [CrossRef] [PubMed]
- Heermann, S.; Schütz, L.; Lemke, S.; Krieglstein, K.; Wittbrodt, J. Eye Morphogenesis Driven by Epithelial Flow into the Optic Cup Facilitated by Modulation of Bone Morphogenetic Protein. eLife 2015, 4, e05216. [Google Scholar] [CrossRef]
- Knickmeyer, M.D.; Mateo, J.L.; Eckert, P.; Roussa, E.; Rahhal, B.; Zuniga, A.; Krieglstein, K.; Wittbrodt, J.; Heermann, S. TGFβ-Facilitated Optic Fissure Fusion and the Role of Bone Morphogenetic Protein Antagonism. Open Biol. 2018, 8, 170134. [Google Scholar] [CrossRef]
- Eckert, P.; Knickmeyer, M.D.; Heermann, S. In Vivo Analysis of Optic Fissure Fusion in Zebrafish: Pioneer Cells, Basal Lamina, Hyaloid Vessels, and How Fissure Fusion Is Affected by BMP. Int. J. Mol. Sci. 2020, 21, 2760. [Google Scholar] [CrossRef]
- Hettige, N.C.; Ernst, C. FOXG1 Dose in Brain Development. Front. Pediatr. 2019, 7, 482. [Google Scholar] [CrossRef] [PubMed]
- Duggan, C.D.; DeMaria, S.; Baudhuin, A.; Stafford, D.; Ngai, J. Foxg1 Is Required for Development of the Vertebrate Olfactory System. J. Neurosci. 2008, 28, 5229–5239. [Google Scholar] [CrossRef] [PubMed]
- Loosli, F.; Staub, W.; Finger-Baier, K.C.; Ober, E.A.; Verkade, H.; Wittbrodt, J.; Baier, H. Loss of eyes in zebrafish caused by mutation of chokh/rx3. EMBO Rep. 2003, 4, 894–899. [Google Scholar] [CrossRef]
- Loosli, F.; Winkler, S.; Burgtorf, C.; Wurmbach, E.; Ansorge, W.; Henrich, T.; Grabher, C.; Arendt, D.; Carl, M.; Krone, A.; et al. Medaka Eyeless Is the Key Factor Linking Retinal Determination and Eye Growth. Development 2001, 128, 4035–4044. [Google Scholar] [CrossRef]
- Wu, R.S.; Lam, I.I.; Clay, H.; Duong, D.N.; Deo, R.C.; Coughlin, S.R. A Rapid Method for Directed Gene Knockout for Screening in G0 Zebrafish. Dev. Cell. 2018, 46, 112–125.e4. [Google Scholar] [CrossRef] [PubMed]
- Warr, N.; Powles-Glover, N.; Chappell, A.; Robson, J.; Norris, D.; Arkell, R.M. Zic2 -Associated Holoprosencephaly Is Caused by a Transient Defect in the Organizer Region during Gastrulation. Hum. Mol. Genet. 2008, 17, 2986–2996. [Google Scholar] [CrossRef] [PubMed]
- Sanek, N.A.; Taylor, A.A.; Nyholm, M.K.; Grinblat, Y. Zebrafish Zic2a Patterns the Forebrain through Modulation of Hedgehog-Activated Gene Expression. Development 2009, 136, 3791–3800. [Google Scholar] [CrossRef]
- Li, H.; Tierney, C.; Wen, L.; Wu, J.Y.; Rao, Y. A Single Morphogenetic Field Gives Rise to Two Retina Primordia under the Influence of the Prechordal Plate. Development 1997, 124, 603–615. [Google Scholar] [CrossRef]
- Sagai, T.; Amano, T.; Maeno, A.; Ajima, R.; Shiroishi, T. SHH Signaling Mediated by a Prechordal and Brain Enhancer Controls Forebrain Organization. Proc. Natl. Acad. Sci. USA 2019, 116, 23636–23642. [Google Scholar] [CrossRef]
- Chow, R.L.; Lang, R.A. Early Eye Development in Vertebrates. Annu. Rev. Cell Dev. Biol. 2001, 17, 255–296. [Google Scholar] [CrossRef]
- Jung, J.; Choi, I.; Ro, H.; Huh, T.-L.; Choe, J.; Rhee, M. march5 Governs the Convergence and Extension Movement for Organization of the Telencephalon and Diencephalon in Zebrafish Embryos. Mol. Cells 2020, 43, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Paridaen, J.T.M.L.; Danesin, C.; Elas, A.T.; van de Water, S.; Houart, C.; Zivkovic, D. Apc1 Is Required for Maintenance of Local Brain Organizers and Dorsal Midbrain Survival. Dev. Biol. 2009, 331, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, E.; Shiota, K. Holoprosencephaly in Human Embryos: Epidemiologic Studies of 150 Cases. Teratology 1977, 16, 261–272. [Google Scholar] [CrossRef]
- Pineda-Alvarez, D.E.; Solomon, B.D.; Roessler, E.; Balog, J.Z.; Hadley, D.W.; Zein, W.M.; Hadsall, C.K.; Brooks, B.P.; Muenke, M. A Broad Range of Ophthalmologic Anomalies Is Part of the Holoprosencephaly Spectrum. Am. J. Med. Genet. A 2011, 155A, 2713–2720. [Google Scholar] [CrossRef]
- Roessler, E.; Hu, P.; Muenke, M. Holoprosencephaly in the Genomics Era. Am. J. Med. Genet. C Semin. Med. Genet. 2018, 178, 165–174. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of Embryonic Development of the Zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Collery, R.F.; Link, B.A. Dynamic Smad-Mediated BMP Signaling Revealed through Transgenic Zebrafish. Dev. Dyn. 2011, 240, 712–722. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Quiring, R.; Wittbrodt, B.; Henrich, T.; Ramialison, M.; Burgtorf, C.; Lehrach, H.; Wittbrodt, J. Large-Scale Expression Screening by Automated Whole-Mount in Situ Hybridization. Mech. Dev. 2004, 121, 971–976. [Google Scholar] [CrossRef] [PubMed]
- Hua, R.; Yu, S.; Liu, M.; Li, H. A PCR-Based Method for RNA Probes and Applications in Neuroscience. Front. Neurosci. 2018, 12, 266. [Google Scholar] [CrossRef]
- Stemmer, M.; Thumberger, T.; del Sol Keyer, M.; Wittbrodt, J.; Mateo, J.L. CCTop: An Intuitive, Flexible and Reliable CRISPR/Cas9 Target Prediction Tool. PLoS ONE 2015, 10, e0124633. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bulk, J.; Kyrychenko, V.; Rensinghoff, P.M.; Ghaderi Ardekani, Z.; Heermann, S. Holoprosencephaly with a Special Form of Anophthalmia Result from Experimental Induction of bmp4, Oversaturating BMP Antagonists in Zebrafish. Int. J. Mol. Sci. 2023, 24, 8052. https://doi.org/10.3390/ijms24098052
Bulk J, Kyrychenko V, Rensinghoff PM, Ghaderi Ardekani Z, Heermann S. Holoprosencephaly with a Special Form of Anophthalmia Result from Experimental Induction of bmp4, Oversaturating BMP Antagonists in Zebrafish. International Journal of Molecular Sciences. 2023; 24(9):8052. https://doi.org/10.3390/ijms24098052
Chicago/Turabian StyleBulk, Johannes, Valentyn Kyrychenko, Philipp M. Rensinghoff, Zahra Ghaderi Ardekani, and Stephan Heermann. 2023. "Holoprosencephaly with a Special Form of Anophthalmia Result from Experimental Induction of bmp4, Oversaturating BMP Antagonists in Zebrafish" International Journal of Molecular Sciences 24, no. 9: 8052. https://doi.org/10.3390/ijms24098052