
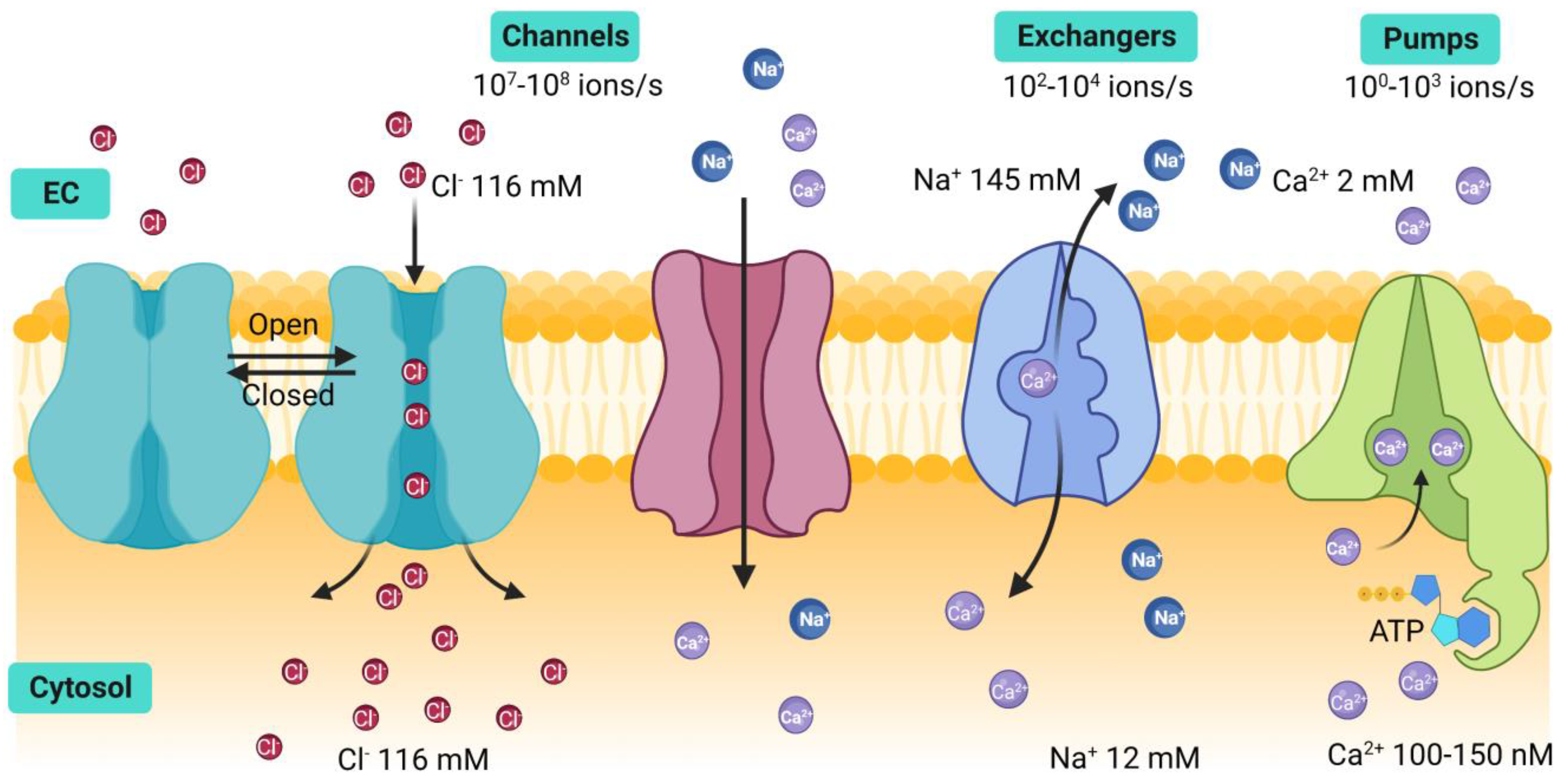
Estrogenic Modulation of Ionic Channels, Pumps and Exchangers in Airway Smooth Muscle
 , , , , and
, , , , and
Abstract
:1. Introduction
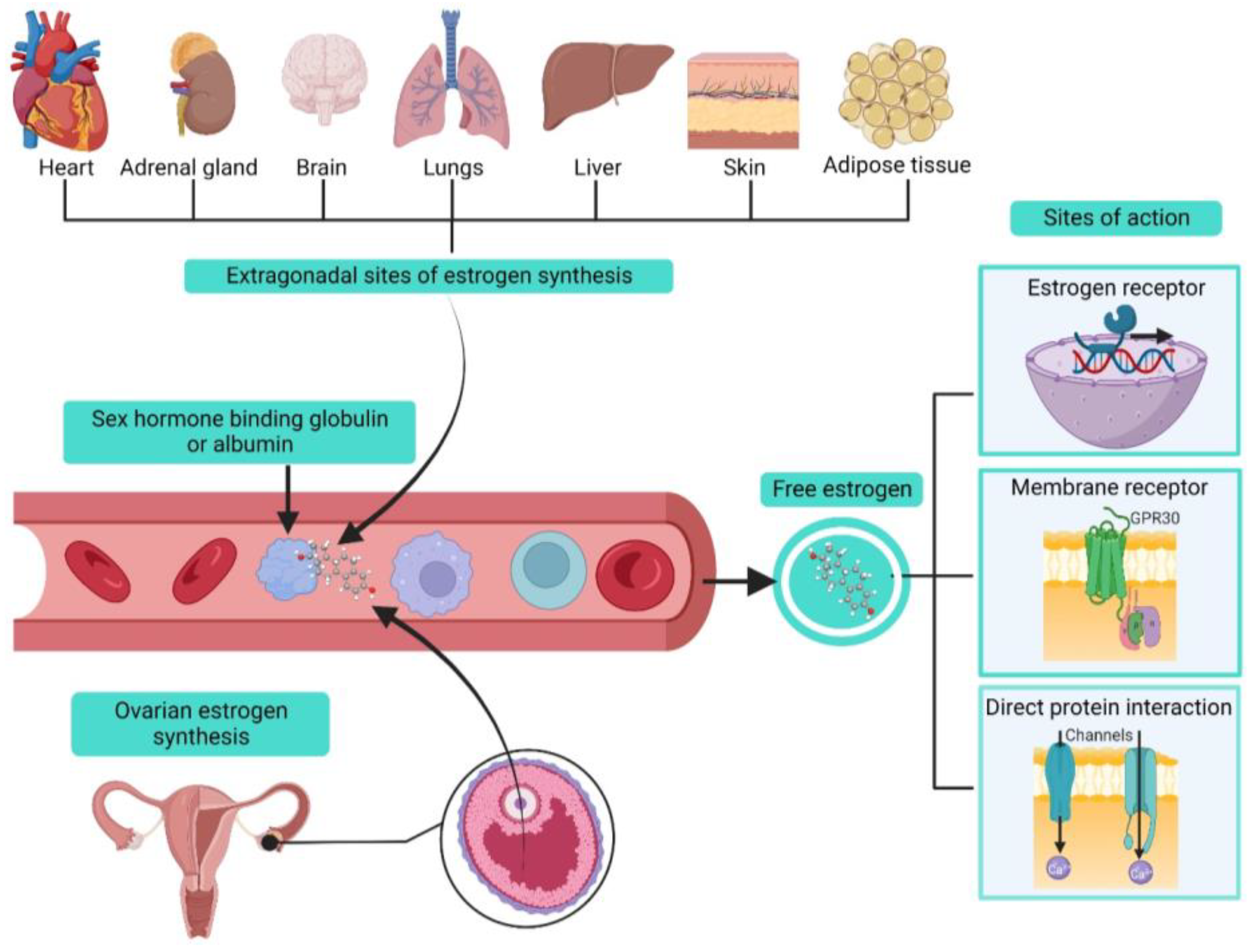
2. Estrogens Biosynthesis and Modes of Action
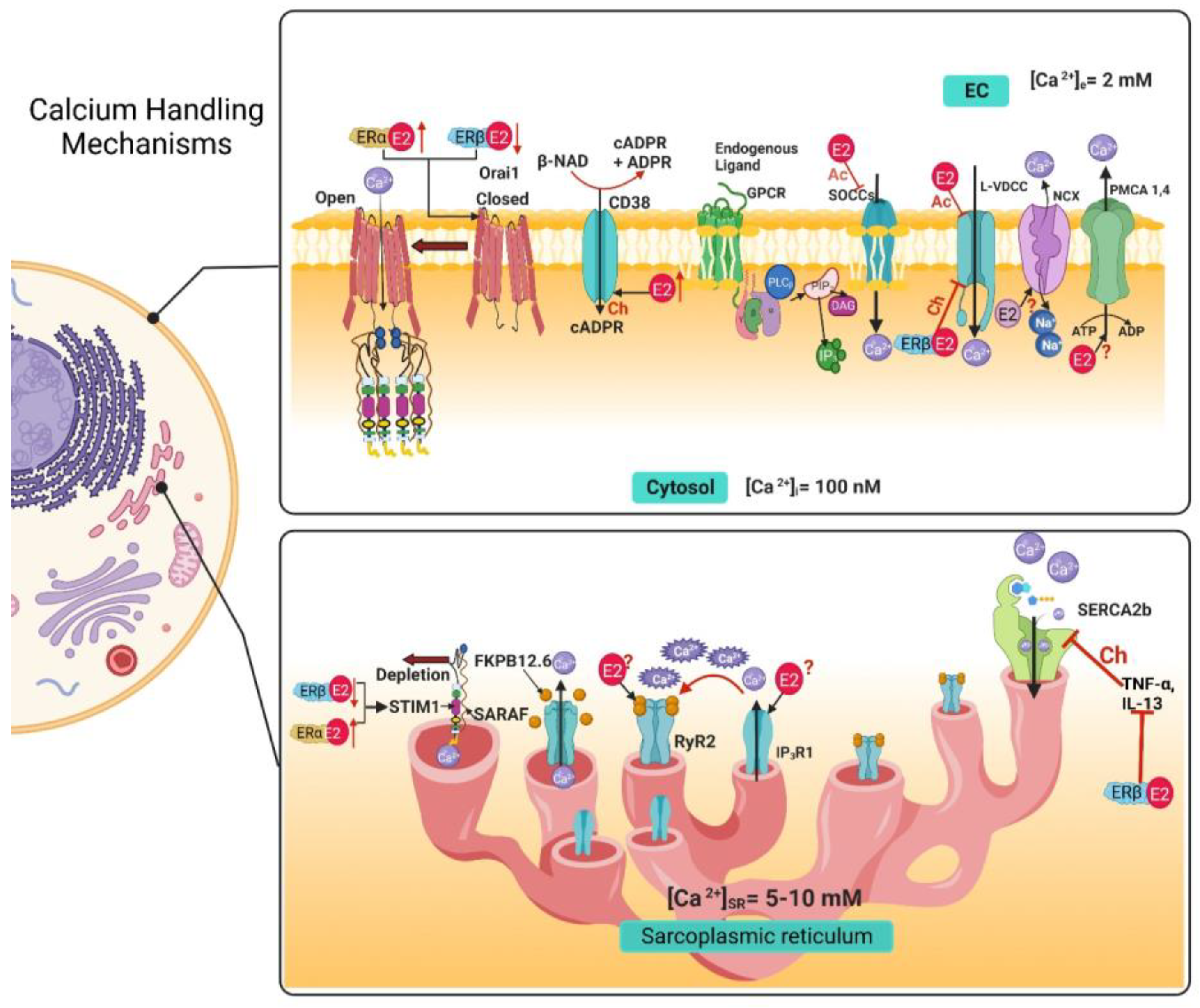
3. Airway Smooth Muscle Calcium Handling Mechanisms and Estrogens
3.1. Voltage-Dependent Ca2+ Channels
3.2. Store-Operated Calcium Channels
3.3. Ryanodine Receptor
3.4. IP3 Receptor
3.5. Na+/Ca2+ Exchanger
3.6. Plasma Membrane Ca2+ ATPase
3.7. Sarcoplasmic Reticulum Ca2+ ATPase

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Acute | Chronic | |||
|---|---|---|---|---|
| Calcium Handling Mechanisms | Pathway | Effect | Pathway | Effect |
| Voltage-dependent Ca2+ channels (VDCCs) | ERα | Inhibition [20,34] | ERβ | Inhibition [35] |
| Store-Operated Calcium Channels (SOCCs) | ERα | Inhibition via STIM1 phosphorylation [20,50] | ERβ | Downregulated STIM1 and Orai1 expression [49] |
| ERα | Upregulated STIM1 and Orai1 expression [49] | |||
| Ryanodine Receptor (RyR) | Unknown | Unknown | ERs | Upregulates CD38 expression [64] |
| IP3 Receptor (IP3R) | Unknown | Unknown | Unknown | Unknown |
| Na+/Ca2+ Exchanger (NCX) | No effect | No effect [20] | Unknown | Unknown |
| Plasma Membrane Ca2+ ATPase (PMCA) | Unknown | Unknown | Unknown | Unknown |
| Sarcoplasmic Reticulum Ca2+ ATPase (SERCA) | No effect | No effect | ERβ | Upregulates SERCA2 expression [35] |
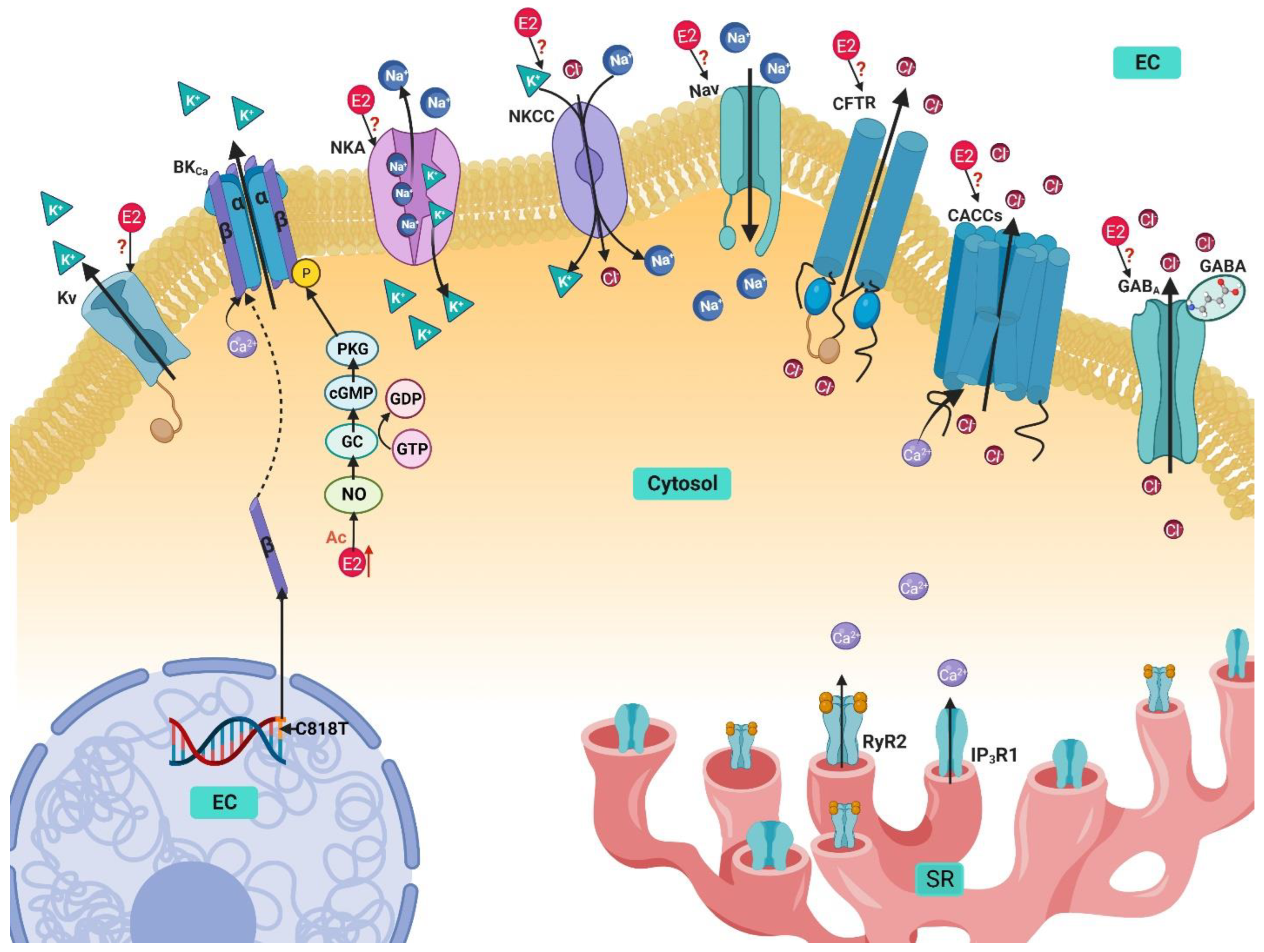
4. Potassium Handling Mechanisms in Airway Smooth Muscle and Their Modulation by Estrogens
4.1. Ca2+-Activated K+ Channels
4.2. Voltage-Activated K+ Channels
4.3. Na+/K+ ATPase
4.4. Na+/K+/Cl− Cotransporter
5. Sodium Handling in Airway Smooth Muscle and Its Modulation by Estrogen
Voltage-Gated Na+ Channels
6. Chlorine Handling Mechanisms in Airway Smooth Muscle and Their Modulation by Estrogen
6.1. Ca2+ activated Cl− Channels
6.2. Cystic Fibrosis Transmembrane Conductance Regulator
6.3. GABA-Activated Cl− Channels
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nehrke, K. Membrane ion transport in non-excitable tissues. WormBook 2014, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssen, L.J. Calcium Handling in Airway Smooth Muscle: Mechanisms and Therapeutic Implications. Can. Respir. J. 1998, 5, 491–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadsby, D.C. Ion channels versus ion pumps: The principal difference, in principle. Nat. Rev. Mol. Cell Biol. 2009, 10, 344–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dascal, N. Ion-channel regulation by G proteins. Trends Endocrinol. Metab. 2001, 12, 391–398. [Google Scholar] [CrossRef]
- Song, S.; Luo, L.; Sun, B.; Sun, D. Roles of glial ion transporters in brain diseases. Glia 2020, 68, 472–494. [Google Scholar] [CrossRef]
- Rosati, B.; Mckinnon, D. Regulation of Ion Channel Expression. Circ. Res. 2004, 94, 874–883. [Google Scholar] [CrossRef]
- Cole, T.J.; Short, K.L.; Hooper, S.B. The science of steroids. Semin. Fetal Neonatal Med. 2019, 24, 170–175. [Google Scholar] [CrossRef]
- Miller, W.L.; Auchus, R.J. The Molecular Biology, Biochemistry, and Physiology of Human Steroidogenesis and Its Disorders. Endocr. Rev. 2011, 32, 81–151. [Google Scholar] [CrossRef] [Green Version]
- Martin, Y.N.; Pabelick, C.M. Sex differences in the pulmonary circulation: Implications for pulmonary hypertension. Am. J. Physiol.-Heart Circ. Physiol. 2014, 306, H1253–H1264. [Google Scholar] [CrossRef] [Green Version]
- Sathish, V.; Martin, Y.N.; Prakash, Y.S. Sex steroid signaling: Implications for lung diseases. Pharmacol. Ther. 2015, 150, 94–108. [Google Scholar] [CrossRef] [Green Version]
- Townsend, E.A.; Miller, V.M.; Prakash, Y.S. Sex Differences and Sex Steroids in Lung Health and Disease. Endocr. Rev. 2012, 33, 1–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, J.; Shen, Y.; Li, R. Estrogen synthesis and signaling pathways during aging: From periphery to brain. Trends Mol. Med. 2013, 19, 197–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurokawa, J.; Kodama, M.; Clancy, C.E.; Furukawa, T. Sex hormonal regulation of cardiac ion channels in drug-induced QT syndromes. Pharmacol. Ther. 2016, 168, 23–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Ma, H.; Barman, S.A.; Liu, A.T.; Sellers, M.; Stallone, J.N.; Prossnitz, E.R.; White, R.E.; Han, G. Activation of G protein-coupled estrogen receptor induces endothelium-independent relaxation of coronary artery smooth muscle. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E882–E888. [Google Scholar] [CrossRef] [Green Version]
- Tran, Q.K. Reciprocality Between Estrogen Biology and Calcium Signaling in the Cardiovascular System. Front. Endocrinol. 2020, 11, 568203. [Google Scholar] [CrossRef]
- Mah, V.; Seligson, D.B.; Li, A.; Márquez, D.C.; Wistuba, I.I.; Elshimali, Y.; Fishbein, M.C.; Chia, D.; Pietras, R.J.; Goodglick, L. Aromatase Expression Predicts Survival in Women with Early-Stage Non–Small Cell Lung Cancer. Cancer Res. 2007, 67, 10484–10490. [Google Scholar] [CrossRef] [Green Version]
- Mair, K.M.; Wright, A.F.; Duggan, N.; Rowlands, D.J.; Hussey, M.J.; Roberts, S.; Fullerton, J.; Nilsen, M.; Loughlin, L.; Thomas, M.; et al. Sex-Dependent Influence of Endogenous Estrogen in Pulmonary Hypertension. Am. J. Respir. Crit. Care Med. 2014, 190, 456–467. [Google Scholar] [CrossRef] [Green Version]
- Martin, Y.N.; Manlove, L.; Dong, J.; Carey, W.A.; Thompson, M.A.; Pabelick, C.M.; Pandya, H.C.; Martin, R.J.; Wigle, D.A.; Prakash, Y.S. Hyperoxia-induced changes in estradiol metabolism in postnatal airway smooth muscle. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2015, 308, L141–L146. [Google Scholar] [CrossRef] [Green Version]
- Heldring, N.; Pike, A.; Andersson, S.; Matthews, J.; Cheng, G.; Hartman, J.; Tujague, M.; Ström, A.; Treuter, E.; Warner, M.; et al. Estrogen receptors: How do they signal and what are their targets. Physiol. Rev. 2007, 87, 905–931. [Google Scholar] [CrossRef] [Green Version]
- Townsend, E.A.; Thompson, M.A.; Pabelick, C.M.; Prakash, Y.S. Rapid effects of estrogen on intracellular Ca2+ regulation in human airway smooth muscle. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2010, 298, L521–L530. [Google Scholar] [CrossRef] [Green Version]
- Aravamudan, B.; Goorhouse, K.J.; Unnikrishnan, G.; Thompson, M.A.; Pabelick, C.M.; Hawse, J.R.; Prakash, Y.S.; Sathish, V. Differential Expression of Estrogen Receptor Variants in Response to Inflammation Signals in Human Airway Smooth Muscle. J. Cell. Physiol. 2017, 232, 1754–1760. [Google Scholar] [CrossRef]
- Kow, L.-M.; Pfaff, D.W. Rapid estrogen actions on ion channels: A survey in search for mechanisms. Steroids 2016, 111, 46–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berridge, M.J.; Bootman, M.D.; Roderick, H.L. Calcium signalling: Dynamics, homeostasis and remodelling. Nat. Rev. Mol. Cell Biol. 2003, 4, 517–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyes-García, J.; Flores-Soto, E.; Carbajal-García, A.; Sommer, B.; Montaño, L. Maintenance of intracellular Ca2+ basal concentration in airway smooth muscle (Review). Int. J. Mol. Med. 2018, 42, 2998–3008. [Google Scholar] [CrossRef] [Green Version]
- Janssen, L.J. T-type and L-type Ca2+ currents in canine bronchial smooth muscle: Characterization and physiological roles. Am. J. Physiol. 1997, 272, C1757–C1765. [Google Scholar] [CrossRef] [PubMed]
- Montaño, L.M.; Barajas-Lopez, C.; Daniel, E.E. Canine bronchial sustained contraction in Ca2+-free medium: Role of intracellular Ca2+. Can. J. Physiol. Pharmacol. 1996, 74, 1236–1248. [Google Scholar] [CrossRef]
- Sommer, B.; Flores-Soto, E.; Reyes-García, J.; Díaz-Hernández, V.; Carbajal, V.; Montaño, L.M. Na+ permeates through L-type Ca2+ channel in bovine airway smooth muscle. Eur. J. Pharmacol. 2016, 782, 77–88. [Google Scholar] [CrossRef]
- Bean, B.P. Classes of calcium channels in vertebrate cells. Annu. Rev. Physiol. 1989, 51, 367–384. [Google Scholar] [CrossRef]
- Yu, J.; Bose, R. Calcium channels in smooth muscle. Gastroenterology 1991, 100, 1448–1460. [Google Scholar] [CrossRef]
- Green, K.A.; Small, R.C.; Foster, R.W. The properties of voltage-operated Ca2+-channels in bovine isolated trachealis cells. Pulm. Pharmacol. 1993, 6, 49–62. [Google Scholar] [CrossRef]
- Hisada, T.; Kurachi, Y.; Sugimoto, T. Properties of membrane currents in isolated smooth muscle cells from guinea-pig trachea. Pflug. Arch. Eur. J. Physiol. 1990, 416, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Kotlikoff, M.I. Calcium currents in isolated canine airway smooth muscle cells. Am. J. Physiol. 1988, 254, C793–C801. [Google Scholar] [CrossRef] [PubMed]
- Marthan, R.; Martin, C.; Amédée, T.; Mironneau, J. Calcium channel currents in isolated smooth muscle cells from human bronchus. J. Appl. Physiol. 1989, 66, 1706–1714. [Google Scholar] [CrossRef] [PubMed]
- Flores-Soto, E.; Reyes-García, J.; Carbajal-García, A.; Campuzano-González, E.; Perusquía, M.; Sommer, B.; Montaño, L.M. Sex steroids effects on guinea pig airway smooth muscle tone and intracellular Ca2+ basal levels. Mol. Cell. Endocrinol. 2017, 439, 444–456. [Google Scholar] [CrossRef]
- Bhallamudi, S.; Connell, J.; Pabelick, C.M.; Prakash, Y.S.; Sathish, V. Estrogen receptors differentially regulate intracellular calcium handling in human nonasthmatic and asthmatic airway smooth muscle cells. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2020, 318, L112–L124. [Google Scholar] [CrossRef] [PubMed]
- Tsang, S.Y.; Yao, X.; Wong, C.M.; Chan, F.L.; Chen, Z.Y.; Huang, Y. Differential regulation of K+ and Ca2+ channel gene expression by chronic treatment with estrogen and tamoxifen in rat aorta. Eur. J. Pharmacol. 2004, 483, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Krebs, J.; Agellon, L.B.; Michalak, M. Ca2+ homeostasis and endoplasmic reticulum (ER) stress: An integrated view of calcium signaling. Biochem. Biophys. Res. Commun. 2015, 460, 114–121. [Google Scholar] [CrossRef]
- Peel, S.E.; Liu, B.; Hall, I.P. A key role for STIM1 in store operated calcium channel activation in airway smooth muscle. Respir. Res. 2006, 7, 119. [Google Scholar] [CrossRef] [Green Version]
- Peel, S.E.; Liu, B.; Hall, I.P. ORAI and Store-Operated Calcium Influx in Human Airway Smooth Muscle Cells. Am. J. Respir. Cell Mol. Biol. 2008, 38, 744–749. [Google Scholar] [CrossRef] [Green Version]
- Palty, R.; Raveh, A.; Kaminsky, I.; Meller, R.; Reuveny, E. SARAF Inactivates the Store Operated Calcium Entry Machinery to Prevent Excess Calcium Refilling. Cell 2012, 149, 425–438. [Google Scholar] [CrossRef] [Green Version]
- Albarran, L.; Regodón, S.; Salido, G.M.; Lopez, J.J.; Rosado, J.A. Role of STIM1 in the surface expression of SARAF. Channels 2017, 11, 84–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Erxleben, C.; Yildirim, E.; Abramowitz, J.; Armstrong, D.L.; Birnbaumer, L. Orai proteins interact with TRPC channels and confer responsiveness to store depletion. Proc. Natl. Acad. Sci. USA 2007, 104, 4682–4687. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.-H.; Zheng, Y.-M.; Liao, B.; Wang, Y.-X. Functional Role of Canonical Transient Receptor Potential 1 and Canonical Transient Receptor Potential 3 in Normal and Asthmatic Airway Smooth Muscle Cells. Am. J. Respir. Cell Mol. Biol. 2010, 43, 17–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yocum, G.T.; Chen, J.; Choi, C.H.; Townsend, E.A.; Zhang, Y.; Xu, D.; Fu, X.W.; Sanderson, M.J.; Emala, C.W. Role of transient receptor potential vanilloid 1 in the modulation of airway smooth muscle tone and calcium handling. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2017, 312, L812–L821. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Zhang, X.; Kuang, H.; Wu, J.; Guo, Y.; Ma, L. Effect of TRPV1 channel on the proliferation and apoptosis in asthmatic rat airway smooth muscle cells. Exp. Lung Res. 2013, 39, 283–294. [Google Scholar] [CrossRef]
- Jia, Y.; Wang, X.; Varty, L.; Rizzo, C.; Yang, R.; Correll, C.; Phelps, P.; Egan, R.; Hey, J. Functional TRPV4 channels are expressed in human airway smooth muscle cells. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2004, 287, L272–L278. [Google Scholar] [CrossRef] [PubMed]
- Uchida, Y.; Izumizaki, M. Effect of menstrual cycle and female hormones on TRP and TREK channels in modifying thermosensitivity and physiological functions in women. J. Therm. Biol. 2021, 100, 103029. [Google Scholar] [CrossRef]
- Flores-Soto, E.; Reyes-García, J.; Sommer, B.; Montaño, L.M. Sarcoplasmic reticulum Ca2+ refilling is determined by L-type Ca2+ and store operated Ca2+ channels in guinea pig airway smooth muscle. Eur. J. Pharmacol. 2013, 721, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Kalidhindi, R.; Ambhore, N.; Thompson, M.; Pabelick, C.; Prakash, Y.; Venkatachalem, S. Differential estrogen receptor signaling regulates store operated calcium entry in human airway smooth muscle. Am. J. Respir. Crit. Care Med. 2018, 197, A7259. [Google Scholar]
- Sathish, V.; Freeman, M.R.; Long, E.; Thompson, M.A.; Pabelick, C.M.; Prakash, Y.S. Cigarette Smoke and Estrogen Signaling in Human Airway Smooth Muscle. Cell. Physiol. Biochem. 2015, 36, 1101–1115. [Google Scholar] [CrossRef]
- Sheridan, J.T.; Gilmore, R.C.; Watson, M.J.; Archer, C.B.; Tarran, R. 17β-Estradiol Inhibits Phosphorylation of Stromal Interaction Molecule 1 (STIM1) Protein. J. Biol. Chem. 2013, 288, 33509–33518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, C.K.; So, W.Y.; Law, S.K.; Leung, F.P.; Yau, K.L.; Yao, X.; Huang, Y.; Li, X.; Tsang, S.Y. Estrogen controls embryonic stem cell proliferation via store-operated calcium entry and the nuclear factor of activated T-cells (NFAT). J. Cell. Physiol. 2012, 227, 2519–2530. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Miao, C.; Liu, M.; Wang, X.; Wang, L.; Wang, D.J. 17β-Estradiol via Orai1 activates calcium mobilization to induce cell proliferation in epithelial ovarian cancer. J. Biochem. Mol. Toxicol. 2020, 34, e22603. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Jiang, K.; Li, J.; Hao, X.; Chu, W.; Luo, C.; Zhu, Y.; Xie, R.; Chen, B. Estrogen enhances the proliferation and migration of ovarian cancer cells by activating transient receptor potential channel C3. J. Ovarian Res. 2020, 13, 20. [Google Scholar] [CrossRef] [Green Version]
- Méndez-Reséndiz, K.A.; Enciso-Pablo, Ó.; González-Ramírez, R.; Juárez-Contreras, R.; Rosenbaum, T.; Morales-Lázaro, S.L. Steroids and TRP Channels: A Close Relationship. Int. J. Mol. Sci. 2020, 21, 3819. [Google Scholar] [CrossRef]
- Yang, H.; Choi, K.C.; Hyun, S.H.; Jeung, E.B. Coexpression and estrogen-mediated regulation of TRPV6 and PMCA1 in the human endometrium during the menstrual cycle. Mol. Reprod. Dev. 2011, 78, 274–282. [Google Scholar] [CrossRef]
- Tran, D.N.; Jung, E.-M.; Ahn, C.; Lee, J.-H.; Yoo, Y.-M.; Jeung, E.-B. Effects of Bisphenol A and 4-tert-Octylphenol on Embryo Implantation Failure in Mouse. Int. J. Environ. Res. Public Health 2018, 15, 1614. [Google Scholar] [CrossRef] [Green Version]
- Lifshitz, L.M.; Carmichael, J.D.; Lai, F.A.; Sorrentino, V.; Bellvé, K.; Fogarty, K.E.; Zhuge, R. Spatial organization of RYRs and BK channels underlying the activation of STOCs by Ca2+ sparks in airway myocytes. J. Gen. Physiol. 2011, 138, 195–209. [Google Scholar] [CrossRef] [Green Version]
- Jude, J.A.; Dileepan, M.; Panettieri, R.A.; Walseth, T.F.; Kannan, M.S. Altered CD38/Cyclic ADP-Ribose Signaling Contributes to the Asthmatic Phenotype. J. Allergy 2012, 2012, 289468. [Google Scholar] [CrossRef] [Green Version]
- Fritz, N.; Macrez, N.; Mironneau, J.; Jeyakumar, L.H.; Fleischer, S.; Morel, J.-L. Ryanodine receptor subtype 2 encodes Ca2+ oscillations activated by acetylcholine via the M2 muscarinic receptor/cADP-ribose signalling pathway in duodenum myocytes. J. Cell Sci. 2005, 118, 2261–2270. [Google Scholar] [CrossRef] [Green Version]
- Deshpande, D.A.; Walseth, T.F.; Panettieri, R.A.; Kannan, M.S. CD38-cyclic ADP-ribose-mediated Ca2+ signaling contributes to airway smooth muscle hyperresponsiveness. FASEB J. 2003, 17, 452–454. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Liang, Q.; Chen, Y.; Wang, H.-S. Molecular Mechanisms Underlying the Rapid Arrhythmogenic Action of Bisphenol A in Female Rat Hearts. Endocrinology 2013, 154, 4607–4617. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Ma, J.; Chen, Y.; Wang, H. Rapid responses and mechanism of action for low-dose bisphenol S on ex vivo rat hearts and isolated myocytes: Evidence of female-specific proarrhythmic effects. Environ. Health Perspect. 2015, 123, 571–578. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Guo, Y.; Huang, W.; Deng, K.Y.; Qian, Y.; Xin, H.B. 17β-Estradiol Promotes Apoptosis in Airway Smooth Muscle Cells Through CD38/SIRT1/p53 Pathway. Front. Endocrinol. 2018, 9, 770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.-Q.; Song, R.; Romero, M.; Dasgupta, C.; Huang, X.; Holguin, M.A.; Williams, V.; Xiao, D.; Wilson, S.M.; Zhang, L. Pregnancy Increases Ca2+ Sparks/Spontaneous Transient Outward Currents and Reduces Uterine Arterial Myogenic Tone. Hypertension 2019, 73, 691–702. [Google Scholar] [CrossRef] [PubMed]
- Jiao, L.; Machuki, J.; Wu, Q.; Shi, M.; Fu, L.; Adekunle, A.; Tao, X.; Xu, C.; Hu, X.; Yin, Z.; et al. Estrogen and calcium handling proteins: New discoveries and mechanisms in cardiovascular diseases. Am. J. Physiol.-Heart Circ. Physiol. 2020, 318, H820–H829. [Google Scholar] [CrossRef]
- Tappia, P.S.; Dent, M.R.; Aroutiounova, N.; Babick, A.P.; Weiler, H. Gender differences in the modulation of cardiac gene expression by dietary conjugated linoleic acid isomers. Can. J. Physiol. Pharmacol. 2007, 3–4, 465–475. [Google Scholar] [CrossRef]
- Yaras, N.; Tuncay, E.; Purali, N.; Sahinoglu, B.; Vassort, G.; Turan, B. Sex-related effects on diabetes-induced alterations in calcium release in the rat heart. Am. J. Physiol.-Heart Circ. Physiol. 2007, 293, H3584–H3592. [Google Scholar] [CrossRef] [Green Version]
- Bell, J.R.; Raaijmakers, A.J.; Curl, C.L.; Reichelt, M.E.; Harding, T.W.; Bei, A.; Ng, D.C.; Erickson, J.R.; Vila Petroff, M.; Harrap, S.B.; et al. Cardiac CaMKIIδ splice variants exhibit target signaling specificity and confer sex-selective arrhythmogenic actions in the ischemic-reperfused heart. Int. J. Cardiol. 2015, 181, 288–296. [Google Scholar] [CrossRef]
- Farrell, S.R.; Ross, J.L.; Howlett, S.E. Sex differences in mechanisms of cardiac excitation-contraction coupling in rat ventricular myocytes. Am. J. Physiol.-Heart Circ. Physiol. 2010, 299, H36–H45. [Google Scholar] [CrossRef] [Green Version]
- Rybalchenko, V.; Grillo, M.A.; Gastinger, M.J.; Rybalchenko, N.; Payne, A.J.; Koulen, P. The unliganded long isoform of estrogen receptor beta stimulates brain ryanodine receptor single channel activity alongside with cytosolic Ca2+. J. Recept. Signal Transduct. Res. 2009, 29, 326–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narayanan, D.; Adebiyi, A.; Jaggar, J.H. Inositol trisphosphate receptors in smooth muscle cells. Am. J. Physiol.-Heart Circ. Physiol. 2012, 302, H2190–H2210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, T.; Hao, Q.; Zheng, Y.M.; Liu, Q.H.; Wang, Y.X. Inositol 1,4,5-trisphosphate activates TRPC3 channels to cause extracellular Ca2+ influx in airway smooth muscle cells. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2015, 309, L1455–L1466. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.X.; Zheng, Y.M.; Mei, Q.B.; Wang, Q.S.; Collier, M.L.; Fleischer, S.; Xin, H.B.; Kotlikoff, M.I. FKBP12.6 and cADPR regulation of Ca2+ release in smooth muscle cells. Am. J. Physiol. Cell Physiol. 2004, 286, C538–C546. [Google Scholar] [CrossRef]
- Montaño, L.M.; Flores-Soto, E.; Reyes-García, J.; Díaz-Hernández, V.; Carbajal-García, A.; Campuzano-González, E.; Ramírez-Salinas, G.L.; Velasco-Velázquez, M.A.; Sommer, B. Testosterone induces hyporesponsiveness by interfering with IP3 receptors in guinea pig airway smooth muscle. Mol. Cell. Endocrinol. 2018, 473, 17–30. [Google Scholar] [CrossRef]
- Romero-Martínez, B.S.; Montaño, L.M.; Solís-Chagoyán, H.; Sommer, B.; Ramírez-Salinas, G.L.; Pérez-Figueroa, G.E.; Flores-Soto, E. Possible Beneficial Actions of Caffeine in SARS-CoV-2. Int. J. Mol. Sci. 2021, 22, 5460. [Google Scholar] [CrossRef]
- Deshpande, D.A.; Wang, W.C.H.; Mcilmoyle, E.L.; Robinett, K.S.; Schillinger, R.M.; An, S.S.; Sham, J.S.K.; Liggett, S.B. Bitter taste receptors on airway smooth muscle bronchodilate by localized calcium signaling and reverse obstruction. Nat. Med. 2010, 16, 1299–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reuquén, P.; Oróstica, M.L.; Rojas, I.; Díaz, P.; Parada-Bustamante, A.; Orihuela, P.A. Estradiol increases IP3 by a nongenomic mechanism in the smooth muscle cells from the rat oviduct. Reproduction 2015, 150, 331–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marino, M.; Pallottini, V.; Trentalance, A. Estrogens cause rapid activation of IP3-PKC-α signal transduction pathway in HEPG2 cells. Biochem. Biophys. Res. Commun. 1998, 245, 254–258. [Google Scholar] [CrossRef]
- Ekstein, J.; Nasatzky, E.; Boyan, B.D.; Ornoy, A.; Schwartz, Z. Growth-plate chondrocytes respond to 17β-estradiol with sex-specific increases in IP3 and intracellular calcium ion signalling via a capacitative entry mechanism. Steroids 2005, 70, 775–786. [Google Scholar] [CrossRef]
- Le Mellay, V.; Grosse, B.; Lieberherr, M. Phospholipase C β and membrane action of calcitriol and estradiol. J. Biol. Chem. 1997, 272, 11902–11907. [Google Scholar] [CrossRef] [Green Version]
- Micevych, P.; Soma, K.K.; Sinchak, K. Neuroprogesterone: Key to estrogen positive feedback? Brain Res. Rev. 2008, 57, 470–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkwood, K.L.; Homick, K.; Dragon, M.B.; Bradford, P.G. Cloning and characterization of the type I inositol 1,4,5-trisphosphate receptor gene promoter. Regulation by 17β-estradiol in osteoblasts. J. Biol. Chem. 1997, 272, 22425–22431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomás, J.; Santos, C.R.A.; Duarte, A.C.; Maltez, M.; Quintela, T.; Lemos, M.C.; Gonçalves, I. Bitter taste signaling mediated by Tas2r144 is down-regulated by 17β-estradiol and progesterone in the rat choroid plexus. Mol. Cell. Endocrinol. 2019, 495, 110521. [Google Scholar] [CrossRef]
- Sommer, B.; Flores-Soto, E.; Gonzalez-Avila, G. Cellular Na+ handling mechanisms involved in airway smooth muscle contraction (Review). Int. J. Mol. Med. 2017, 40, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiPolo, R.; Beaugé, L. Sodium/calcium exchanger: Influence of metabolic regulation on ion carrier interactions. Physiol. Rev. 2006, 86, 155–203. [Google Scholar] [CrossRef] [Green Version]
- Philipson, K.D.; Nicoll, D.A. Sodium-calcium exchange: A molecular perspective. Annu. Rev. Physiol. 2000, 62, 111–133. [Google Scholar] [CrossRef]
- Lytton, J. Na+/Ca2+ exchangers: Three mammalian gene families control Ca2+ transport. Biochem. J. 2007, 406, 365–382. [Google Scholar] [CrossRef]
- Algara-Suárez, P.; Mejía-Elizondo, R.; Sims, S.; Saavedra-Alanis, V.; Espinosa-Tanguma, R. The 1.3 isoform of Na+-Ca2+ exchanger expressed in guinea pig tracheal smooth muscle is less sensitive to KB-R7943. J. Physiol. Biochem. 2010, 66, 117–125. [Google Scholar] [CrossRef]
- Janssen, L.J.; Walters, D.K.; Wattie, J. Regulation of [Ca2+]i in canine airway smooth muscle by Ca2+-ATPase and Na+/Ca2+ exchange mechanisms. Am. J. Physiol. 1997, 273, L322–L330. [Google Scholar] [CrossRef]
- Wen, J.; Meng, X.; Xuan, B.; Zhou, T.; Gao, H.; Dong, H.; Wang, Y. Na+/Ca2+ Exchanger 1 in Airway Smooth Muscle of Allergic Inflammation Mouse Model. Front. Pharmacol. 2018, 9, 1471. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Zhao, P.; Zhang, W.-J.; Qiu, J.-Y.; Tan, L.; Liu, X.-C.; Wang, Q.; Luo, X.; She, Y.-S.; Zang, D.-A.; et al. Generation and Role of Oscillatory Contractions in Mouse Airway Smooth Muscle. Cell. Physiol. Biochem. 2018, 47, 1546–1555. [Google Scholar] [CrossRef] [PubMed]
- Sathish, V.; Delmotte, P.F.; Thompson, M.A.; Pabelick, C.M.; Sieck, G.C.; Prakash, Y.S. Sodium-calcium exchange in intracellular calcium handling of human airway smooth muscle. PLoS ONE 2011, 6, e23662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.; Inman, M.; Kiss, L.; Janssen, L.J. Reverse-mode NCX current in mouse airway smooth muscle: Na+ and voltage dependence, contributions to Ca2+ influx and contraction, and altered expression in a model of allergen-induced hyperresponsiveness. Acta Physiol. 2012, 205, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Lee, D.; Ahn, C.; Kang, H.Y.; An, B.S.; Seong, Y.H.; Jeung, E.B. Effects of estrogen on esophageal function through regulation of Ca2+-related proteins. J. Gastroenterol. 2017, 52, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Chu, S.H.; Goldspink, P.; Kowalski, J.; Beck, J.; Schwertz, D.W. Effect of estrogen on calcium-handling proteins, β-adrenergic receptors, and function in rat heart. Life Sci. 2006, 79, 1257–1267. [Google Scholar] [CrossRef]
- Sims, C.; Reisenweber, S.; Viswanathan, P.C.; Choi, B.R.; Walker, W.H.; Salama, G. Sex, age, and regional differences in L-type calcium current are important determinants of arrhythmia phenotype in rabbit hearts with drug-induced long QT type 2. Circ. Res. 2008, 102, e86–e100. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Yang, X.; Alber, S.; Shusterman, V.; Salama, G. Regional genomic regulation of cardiac sodium-calcium exchanger by oestrogen. J. Physiol. 2011, 589, 1061–1080. [Google Scholar] [CrossRef]
- Kravtsov, G.M.; Kam, K.W.; Liu, J.; Wu, S.; Wong, T.M. Altered Ca2+ handling by ryanodine receptor and Na+-Ca2+ exchange in the heart from ovariectomized rats: Role of protein kinase A. Am. J. Physiol. Cell Physiol. 2007, 292, C1625–C1635. [Google Scholar] [CrossRef]
- Yang, H.Y.; Firth, J.M.; Francis, A.J.; Alvarez-Laviada, A.; MacLeod, K.T. Effect of ovariectomy on intracellular Ca2+ regulation in guinea pig cardiomyocytes. Am. J. Physiol.-Heart Circ. Physiol. 2017, 313, H1031–H1043. [Google Scholar] [CrossRef] [Green Version]
- Sugishita, K.; Su, Z.; Li, F.; Philipson, K.D.; Barry, W.H. Gender influences [Ca2+]i during metabolic inhibition in myocytes overexpressing the Na+-Ca2+ exchanger. Circulation 2001, 104, 2101–2106. [Google Scholar] [CrossRef] [Green Version]
- Cross, H.R.; Lu, L.; Steenbergen, C.; Philipson, K.D.; Murphy, E. Overexpression of the Cardiac Na+/Ca2+ Exchanger Increases Susceptibility to Ischemia/Reperfusion Injury in Male, but Not Female, Transgenic Mice. Circ. Res. 1998, 83, 1215–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, J.C.; López-Zapata, D.F.; Francis, L.; De Los Reyes, L. Effects of estradiol and IGF-1 on the sodium calcium exchanger in rat cultured cortical neurons. Cell. Mol. Neurobiol. 2011, 31, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Jeffs, G.J.; Meloni, B.P.; Bakker, A.J.; Knuckey, N.W. The role of the Na+/Ca2+ exchanger (NCX) in neurons following ischaemia. J. Clin. Neurosci. Off. J. Neurosurg. Soc. Australas. 2007, 14, 507–514. [Google Scholar] [CrossRef]
- Chen, Y.F.; Cao, J.; Zhong, J.N.; Chen, X.; Cheng, M.; Yang, J.; Gao, Y.D. Plasma membrane Ca2+-ATPase regulates Ca2+ signaling and the proliferation of airway smooth muscle cells. Eur. J. Pharmacol. 2014, 740, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Varga, K.; Hollósi, A.; Pászty, K.; Hegedűs, L.; Szakács, G.; Tímár, J.; Papp, B.; Enyedi, Á.; Padányi, R. Expression of calcium pumps is differentially regulated by histone deacetylase inhibitors and estrogen receptor alpha in breast cancer cells. BMC Cancer 2018, 18, 1029. [Google Scholar] [CrossRef] [Green Version]
- El-Beialy, W.; Galal, N.; Deyama, Y.; Yoshimura, Y.; Suzuki, K.; Tei, K.; Totsuka, Y. Effects of Estrogen on PMCA 2 and 4 in Human Fibroblast-like Synovial Cells and Mouse Macrophage-like Cells. Endocr. J. 2010, 57, 93–97. [Google Scholar] [CrossRef] [Green Version]
- Dick, I.M.; Liu, J.; Glendenning, P.; Prince, R.L. Estrogen and androgen regulation of plasma membrane calcium pump activity in immortalized distal tubule kidney cells. Mol. Cell. Endocrinol. 2003, 212, 11–18. [Google Scholar] [CrossRef]
- Khariv, V.; Acioglu, C.; Ni, L.; Ratnayake, A.; Li, L.; Tao, Y.-X.; Heary, R.F.; Elkabes, S. A link between plasma membrane calcium ATPase 2 (PMCA2), estrogen and estrogen receptor α signaling in mechanical pain. Sci. Rep. 2018, 8, 17260. [Google Scholar] [CrossRef] [Green Version]
- Bobe, R.; Bredoux, R.; Corvazier, E.; Andersen, J.P.; Clausen, J.D.; Dode, L.; Kovács, T.; Enouf, J. Identification, Expression, Function, and Localization of a Novel (Sixth) Isoform of the Human Sarco/Endoplasmic Reticulum Ca2+ ATPase 3 Gene. J. Biol. Chem. 2004, 279, 24297–24306. [Google Scholar] [CrossRef] [Green Version]
- Mahn, K.; Hirst, S.J.; Ying, S.; Holt, M.R.; Lavender, P.; Ojo, O.O.; Siew, L.; Simcock, D.E.; Mcvicker, C.G.; Kanabar, V.; et al. Diminished sarco/endoplasmic reticulum Ca2+ ATPase (SERCA) expression contributes to airway remodelling in bronchial asthma. Proc. Natl. Acad. Sci. USA 2009, 106, 10775–10780. [Google Scholar] [CrossRef] [Green Version]
- Prakash, Y.S.; Sathish, V.; Thompson, M.A.; Pabelick, C.M.; Sieck, G.C. Asthma and sarcoplasmic reticulum Ca2+ reuptake in airway smooth muscle. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2009, 297, L794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbajal, V.; Vargas, M.H.; lores-Soto, E.F.; Martínez-Cordero, E.; Bazán-Perkins, B.; Montaño, L.M. LTD4 induces hyperresponsiveness to histamine in bovine airway smooth muscle: Role of SR-ATPase Ca2+ pump and tyrosine kinase. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2005, 288, L84–L92. [Google Scholar] [CrossRef] [PubMed]
- Kotlikoff, M.I. Potassium channels in airway smooth muscle: A tale of two channels. Pharmacol. Ther. 1993, 58, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Adda, S.; Fleischmann, B.K.; Freedman, B.D.; Yu, M.; Hay, D.W.; Kotlikoff, M.I. Expression and function of voltage-dependent potassium channel genes in human airway smooth muscle. J. Biol. Chem. 1996, 271, 13239–13243. [Google Scholar] [CrossRef] [Green Version]
- Knox, A.J.; Tattersfield, A.E. Airway smooth muscle relaxation. Thorax 1995, 50, 894–901. [Google Scholar] [CrossRef] [Green Version]
- Brueggemann, L.I.; Kakad, P.P.; Love, R.B.; Solway, J.; Dowell, M.L.; Cribbs, L.L.; Byron, K.L. Kv7 potassium channels in airway smooth muscle cells: Signal transduction intermediates and pharmacological targets for bronchodilator therapy. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2012, 302, L120–L132. [Google Scholar] [CrossRef] [Green Version]
- Isaac, L.; Mcardle, S.; Miller, N.M.; Foster, R.W.; Small, R.C. Effects of some K+-channel inhibitors on the electrical behaviour of guinea-pig isolated trachealis and on its responses to spasmogenic drugs. Br. J. Pharmacol. 1996, 117, 1653–1662. [Google Scholar] [CrossRef] [Green Version]
- Janssen, L.J.; Nana, R. Na+/K+ ATPase mediates rhythmic spontaneous relaxations in canine airway smooth muscle. Respir. Physiol. 1997, 108, 187–194. [Google Scholar] [CrossRef]
- Dodson, A.M.; Rhoden, K.J. Bradykinin increases Na+-K+pump activity in cultured guinea-pig tracheal smooth muscle cells. Br. J. Pharmacol. 2001, 133, 1339–1345. [Google Scholar] [CrossRef] [Green Version]
- Rhoden, K.J.; Douglas, J.S. Evidence of Na-K-Cl cotransport in airway smooth muscle. Am. J. Physiol. 1995, 268, L551–L557. [Google Scholar] [CrossRef] [PubMed]
- Kume, H.; Hall, I.P.; Washabau, R.J.; Takagi, K.; Kotlikoff, M.I. β-adrenergic agonists regulate KCa channels in airway smooth muscle by cAMP-dependent and -independent mechanisms. J. Clin. Investig. 1994, 93, 371–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, G.; O’Connell, R.J.; Pietrzykowski, A.Z.; Treistman, S.N.; Ethier, M.F.; Madison, J.M. Interleukin-4 activates large-conductance, calcium-activated potassium (BKCa) channels in human airway smooth muscle cells. Exp. Physiol. 2008, 93, 908–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perkins, C.; Yanase, N.; Smulian, G.; Gildea, L.; Orekov, T.; Potter, C.; Brombacher, F.; Aronow, B.; Wills-Karp, M.; Finkelman, F.D. Selective stimulation of IL-4 receptor on smooth muscle induces airway hyperresponsiveness in mice. J. Exp. Med. 2011, 208, 853–867. [Google Scholar] [CrossRef]
- Shepherd, M.C.; Duffy, S.M.; Harris, T.; Cruse, G.; Schuliga, M.; Brightling, C.E.; Neylon, C.B.; Bradding, P.; Stewart, A.G. KCa3.1 Ca2+-Activated K+ Channels Regulate Human Airway Smooth Muscle Proliferation. Am. J. Respir. Cell Mol. Biol. 2007, 37, 525–531. [Google Scholar] [CrossRef]
- Yu, Z.H.; Wang, Y.X.; Song, Y.; Lu, H.Z.; Hou, L.N.; Cui, Y.Y.; Chen, H.Z. Up-regulation of KCa3.1 promotes human airway smooth muscle cell phenotypic modulation. Pharmacol. Res. 2013, 77, 30–38. [Google Scholar] [CrossRef]
- Yu, Z.-H.; Xu, J.-R.; Wang, Y.-X.; Xu, G.-N.; Xu, Z.-P.; Yang, K.; Wu, D.-Z.; Cui, Y.-Y.; Chen, H.-Z. Targeted Inhibition of KCa3.1 Channel Attenuates Airway Inflammation and Remodeling in Allergic Asthma. Am. J. Respir. Cell Mol. Biol. 2013, 48, 685–693. [Google Scholar] [CrossRef]
- Dimitropoulou, C.; White, R.E.; Ownby, D.R.; Catravas, J.D. Estrogen Reduces Carbachol-Induced Constriction of Asthmatic Airways by Stimulating Large-Conductance Voltage and Calcium-Dependent Potassium Channels. Am. J. Respir. Cell Mol. Biol. 2005, 32, 239–247. [Google Scholar] [CrossRef]
- Seibold, M.A.; Wang, B.; Eng, C.; Kumar, G.; Beckman, K.B.; Sen, S.; Choudhry, S.; Meade, K.; Lenoir, M.; Watson, H.G.; et al. An african-specific functional polymorphism in KCNMB1 shows sex-specific association with asthma severity. Hum. Mol. Genet. 2008, 17, 2681–2690. [Google Scholar] [CrossRef] [Green Version]
- Tsang, S.Y.; Yao, X.; Chan, H.Y.; Wong, C.M.; Chen, Z.Y.; Au, C.L.; Huang, Y. Contribution of K+ channels to relaxation induced by 17β-estradiol but not by progesterone in isolated rat mesenteric artery rings. J. Cardiovasc. Pharmacol. 2003, 41, 4–13. [Google Scholar] [CrossRef]
- Wong, C.M.; Tsang, S.Y.; Yao, X.; Chan, F.L.; Huang, Y. Differential effects of estrogen and progesterone on potassium channels expressed in Xenopus oocytes. Steroids 2008, 73, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.-Q.; Xiao, D.; Zhu, R.; Huang, X.; Yang, S.; Wilson, S.; Zhang, L. Pregnancy Upregulates Large-Conductance Ca2+-Activated K+-Channel Activity and Attenuates Myogenic Tone in Uterine Arteries. Hypertension 2011, 58, 1132–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.T.; Qiu, X.Y. 17β-Estradiol Upregulated Expression of α and β Subunits of Larger-Conductance Calcium-Activated K+ Channels (BK) via Estrogen Receptor β. J. Mol. Neurosci. 2015, 56, 799–807. [Google Scholar] [CrossRef]
- Nishimura, I.; Ui-Tei, K.; Saigo, K.; Ishii, H.; Sakuma, Y.; Kato, M. 17β-Estradiol at Physiological Concentrations Augments Ca2+-Activated K+ Currents via Estrogen Receptor β in the Gonadotropin-Releasing Hormone Neuronal Cell Line GT1-7. Endocrinology 2008, 149, 774–782. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Jin, L.; Leng, J.; Lang, J. Response of potassium channels to estrogen and progesterone in the uterine smooth muscle cells of adenomyosis in vitro. Zhonghua Fu Chan Ke Za Zhi 2015, 50, 843–847. [Google Scholar]
- Restrepo-Angulo, I.; Bañuelos, C.; Camacho, J. Ion Channel Regulation by Sex Steroid Hormones and Vitamin D in Cancer: A Potential Opportunity for Cancer Diagnosis and Therapy. Front. Pharmacol. 2020, 11, 152. [Google Scholar] [CrossRef]
- Coiret, G.; Matifat, F.; Hague, F.; Ouadid-Ahidouch, H. 17-β-Estradiol activates maxi-K channels through a non-genomic pathway in human breast cancer cells. FEBS Lett. 2005, 579, 2995–3000. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, A.; Vera, E.; Gamboa-Domínguez, A.; Lambert, P.; Gariglio, P.; Camacho, J. Calcium-activated potassium channels as potential early markers of human cervical cancer. Oncol. Lett. 2018, 15, 7249–7254. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Pinna, J.; Marroqui, L.; Hmadcha, A.; Lopez-Beas, J.; Soriano, S.; Villar-Pazos, S.; Alonso-Magdalena, P.; Dos Santos, R.S.; Quesada, I.; Martin, F.; et al. Oestrogen receptor β mediates the actions of bisphenol-A on ion channel expression in mouse pancreatic beta cells. Diabetologia 2019, 62, 1667–1680. [Google Scholar] [CrossRef] [Green Version]
- Marroqui, L.; Martinez-Pinna, J.; Castellano-Muñoz, M.; Dos Santos, R.S.; Medina-Gali, R.M.; Soriano, S.; Quesada, I.; Gustafsson, J.-A.; Encinar, J.A.; Nadal, A. Bisphenol-S and Bisphenol-F alter mouse pancreatic β-cell ion channel expression and activity and insulin release through an estrogen receptor ERβ mediated pathway. Chemosphere 2021, 265, 129051. [Google Scholar] [CrossRef]
- Mohr, C.J.; Steudel, F.A.; Gross, D.; Ruth, P.; Lo, W.-Y.; Hoppe, R.; Schroth, W.; Brauch, H.; Huber, S.M.; Lukowski, R. Cancer-Associated Intermediate Conductance Ca2+-Activated K+ Channel KCa3.1. Cancers 2019, 11, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sticht, C.; De La Torre, C.; Parveen, A.; Gretz, N. miRWalk: An online resource for prediction of microRNA binding sites. PLoS ONE 2018, 13, e0206239. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zhang, X.; Dhakal, I.B.; Beggs, M.; Kadlubar, S.; Luo, D. Induction of cell proliferation and survival genes by estradiol-repressed microRNAs in breast cancer cells. BMC Cancer 2012, 12, 29. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.S.; Sun, M.; Kho, A.T.; Moore, K.G.; Sylvia, J.M.; Weiss, S.T.; Lu, Q.; Tantisira, K.G. Circulating microRNAs and association with methacholine PC20 in the Childhood Asthma Management Program (CAMP) cohort. PLoS ONE 2017, 12, e0180329. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Yao, L.; Liu, C.; Tang, L.; Xing, T. Upregulation of microRNA-16 alters the response to inhaled β-agonists in patients with asthma though modulating expression of ADRB2. Mol. Med. Rep. 2019, 19, 4027–4034. [Google Scholar] [CrossRef] [Green Version]
- Taura, J.; Kircher, D.M.; Gameiro-Ros, I.; Slesinger, P.A. Comparison of K+ Channel Families. In Pharmacology of Potassium Channels; Gamper, N., Wang, K., Eds.; Springer: Cham, Switzerland, 2021; Volume 267. [Google Scholar]
- Evseev, A.I.; Semenov, I.; Archer, C.R.; Medina, J.L.; Dube, P.H.; Shapiro, M.S.; Brenner, R. Functional effects of KCNQ K+ channels in airway smooth muscle. Front. Physiol. 2013, 4, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drici, M.D.; Burklow, T.R.; Haridasse, V.; Glazer, R.I.; Woosley, R.L. Sex hormones prolong the QT interval and downregulate potassium channel expression in the rabbit heart. Circulation 1996, 94, 1471–1474. [Google Scholar] [CrossRef]
- Roepke, T.A.; Malyala, A.; Bosch, M.A.; Kelly, M.J.; Rønnekleiv, O.K. Estrogen Regulation of Genes Important for K+ Channel Signaling in the Arcuate Nucleus. Endocrinology 2007, 148, 4937–4951. [Google Scholar] [CrossRef]
- Roepke, T.A.; Qiu, J.; Smith, A.W.; Ronnekleiv, O.K.; Kelly, M.J. Fasting and 17β-Estradiol Differentially Modulate the M-Current in Neuropeptide Y Neurons. J. Neurosci. 2011, 31, 11825–11835. [Google Scholar] [CrossRef] [Green Version]
- Anneken, L.; Baumann, S.; Vigneault, P.; Biliczki, P.; Friedrich, C.; Xiao, L.; Girmatsion, Z.; Takac, I.; Brandes, R.P.; Kissler, S.; et al. Estradiol regulates human QT-interval: Acceleration of cardiac repolarization by enhanced KCNH2 membrane trafficking. Eur. Heart J. 2016, 37, 640–650. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.G.; Leem, Y.-E.; Kwon, I.; Kang, J.-S.; Bae, Y.M.; Cho, H. Estrogen modulates serotonin effects on vasoconstriction through Src inhibition. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Díaz, L.; Ceja-Ochoa, I.; Restrepo-Angulo, I.; Larrea, F.; Avila-Chávez, E.; García-Becerra, R.; Borja-Cacho, E.; Barrera, D.; Ahumada, E.; Gariglio, P.; et al. Estrogens and Human Papilloma Virus Oncogenes Regulate Human Ether-à-go-go-1 Potassium Channel Expression. Cancer Res. 2009, 69, 3300–3307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlson, A.E.; Brelidze, T.I.; Zagotta, W.N. Flavonoid regulation of EAG1 channels. J. Gen. Physiol. 2013, 141, 347–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souhrada, M.; Souhrada, J.F.; Cherniack, R.M. Evidence for a sodium electrogenic pump in airway smooth muscle. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1981, 51, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Chideckel, E.W.; Frost, J.L.; Mike, P.; Fedan, J.S. The effect of ouabain on tension in isolated respiratory tract smooth muscle of humans and other species. Br. J. Pharmacol. 1987, 92, 609–614. [Google Scholar] [CrossRef] [Green Version]
- Gunst, S.J.; Stropp, J.Q. Effect of Na-K adenosinetriphosphatase activity on relaxation of canine tracheal smooth muscle. J. Appl. Physiol. 1988, 64, 635–641. [Google Scholar] [CrossRef]
- Obradovic, M.; Zafirovic, S.; Jovanovic, A.; Milovanovic, E.S.; Mousa, S.A.; Labudovic-Borovic, M.; Isenovic, E.R. Effects of 17β-estradiol on cardiac Na+/K+-ATPase in high fat diet fed rats. Mol. Cell. Endocrinol. 2015, 416, 46–56. [Google Scholar] [CrossRef]
- Liu, C.G.; Xu, K.Q.; Xu, X.; Huang, J.J.; Xiao, J.C.; Zhang, J.P.; Song, H.P. 17β-oestradiol regulates the expression of Na+/K+-ATPase β1-subunit, sarcoplasmic reticulum Ca2+-ATPase and carbonic anhydrase iv in H9C2 cells. Clin. Exp. Pharmacol. Physiol. 2007, 34, 998–1004. [Google Scholar] [CrossRef]
- Obradovic, M.; Stewart, A.J.; Pitt, S.J.; Labudovic-Borovic, M.; Sudar, E.; Petrovic, V.; Zafirovic, S.; Maravic-Stojkovic, V.; Vasic, V.; Isenovic, E.R. In vivo effects of 17β-estradiol on cardiac Na+/K+-ATPase expression and activity in rat heart. Mol. Cell. Endocrinol. 2014, 388, 58–68. [Google Scholar] [CrossRef]
- Li, Y.; Yang, J.; Li, S.; Zhang, J.; Zheng, J.; Hou, W.; Zhao, H.; Guo, Y.; Liu, X.; Dou, K.; et al. N-myc Downstream-regulated Gene 2, a Novel Estrogen-targeted Gene, Is Involved in the Regulation of Na+/K+-ATPase. J. Biol. Chem. 2011, 286, 32289–32299. [Google Scholar] [CrossRef] [Green Version]
- Melis, M.G.; Troffa, C.; Manunta, P.; Pinna Parpaglia, P.; Soro, A.; Pala, F.; Madeddu, P.; Pazzola, A.; Tonolo, G.; Patteri, G. Influenze degli ormoni del ciclo mestruale sui trasporti cationici di membrana dei globuli rossi [Effect of menstrual cycle hormones on cation transport in the red-cell membrane]. Boll. Della Soc. Ital. Di Biol. Sper. 1990, 66, 679–684. [Google Scholar]
- Palacios, J.; Marusic, E.T.; Lopez, N.C.; Gonzalez, M.; Michea, L. Estradiol-induced expression of N+-K+-ATPase catalytic isoforms in rat arteries: Gender differences in activity mediated by nitric oxide donors. Am. J. Physiol.-Heart Circ. Physiol. 2004, 286, H1793–H1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro Junior, R.F.; Fiorim, J.; Marques, V.B.; de Sousa Ronconi, K.; Botelho, T.; Grando, M.D.; Bendhack, L.M.; Vassallo, D.V.; Stefanon, I. Vascular activation of K+ channels and Na+-K+ ATPase activity of estrogen-deficient female rats. Vasc. Pharmacol. 2017, 99, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, L.M.; Fujiwara, N.; Nakamura, K.T.; Wada, R.K. Na-K-2Cl cotransporter inhibition impairs human lung cellular proliferation. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2004, 287, L510–L514. [Google Scholar] [CrossRef]
- Wang, S.; Xiang, Y.Y.; Ellis, R.; Wattie, J.; Feng, M.; Inman, M.D.; Lu, W.Y. Effects of furosemide on allergic asthmatic responses in mice. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2011, 41, 1456–1467. [Google Scholar] [CrossRef]
- O’Donnell, M.E.; Lam, T.I.; Tran, L.Q.; Foroutan, S.; Anderson, S.E. Estradiol Reduces Activity of the Blood–Brain Barrier Na–K–Cl Cotransporter and Decreases Edema Formation in Permanent Middle Cerebral Artery Occlusion. J. Cereb. Blood Flow Metab. 2006, 26, 1234–1249. [Google Scholar] [CrossRef] [Green Version]
- Chang, E.; O’Donnell, M.E.; Barakat, A.I. Shear stress and 17β-estradiol modulate cerebral microvascular endothelial Na-K-Cl cotransporter and Na/H exchanger protein levels. Am. J. Physiol. Cell Physiol. 2008, 294, C363–C371. [Google Scholar] [CrossRef]
- Nakamura, N.H.; Rosell, D.R.; Akama, K.T.; McEwen, B.S. Estrogen and ovariectomy regulate mRNA and protein of glutamic acid decarboxylases and cation-chloride cotransporters in the adult rat hippocampus. Neuroendocrinology 2004, 80, 308–323. [Google Scholar] [CrossRef]
- Nugent, B.M.; Valenzuela, C.V.; Simons, T.J.; Mccarthy, M.M. Kinases SPAK and OSR1 Are Upregulated by Estradiol and Activate NKCC1 in the Developing Hypothalamus. J. Neurosci. 2012, 32, 593–598. [Google Scholar] [CrossRef] [Green Version]
- Palacios, J.; Espinoza, F.; Munita, C.; Cifuentes, F.; Michea, L. Na+-K+-2Cl− cotransporter is implicated in gender differences in the response of the rat aorta to phenylephrine. Br. J. Pharmacol. 2006, 148, 964–972. [Google Scholar] [CrossRef] [Green Version]
- Bers, D. Intracellular Na+ regulation in cardiac myocytes. Cardiovasc. Res. 2003, 57, 897–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradley, E.; Webb, T.I.; Hollywood, M.A.; Sergeant, G.P.; McHale, N.G.; Thornbury, K.D. The cardiac sodium current Na(v)1.5 is functionally expressed in rabbit bronchial smooth muscle cells. Am. J. Physiol. Cell Physiol. 2013, 305, C427–C435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snetkov, V.; Hirst, S.; Ward, J. Ion channels in freshly isolated and cultured human bronchial smooth muscle cells. Exp. Physiol. 1996, 81, 791–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, T.; Nagata, T.; Iida, H.; Imuta, H.; Iwasawa, K.; Ma, J.; Hara, K.; Omata, M.; Nagai, R.; Takizawa, H.; et al. Voltage-gated sodium channel expressed in cultured human smooth muscle cells: Involvement of SCN9A. FEBS Lett. 2004, 567, 339–343. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, T.; Jo, T.; Meguro, K.; Oonuma, H.; Ma, J.; Kubota, N.; Imuta, H.; Takano, H.; Iida, H.; Nagase, T.; et al. Effect of dexamethasone on voltage-gated Na+ channel in cultured human bronchial smooth muscle cells. Life Sci. 2008, 82, 1210–1215. [Google Scholar] [CrossRef]
- Fraser, S.P.; Pardo, L.A. Ion channels: Functional expression and therapeutic potential in cancer. Colloquium on Ion Channels and Cancer. EMBO Rep. 2008, 9, 512–515. [Google Scholar] [CrossRef] [Green Version]
- Fraser, S.P.; Ozerlat-Gunduz, I.; Onkal, R.; Diss, J.K.; Latchman, D.S.; Djamgoz, M.B. Estrogen and non-genomic upregulation of voltage-gated Na+ channel activity in MDA-MB-231 human breast cancer cells: Role in adhesion. J. Cell. Physiol. 2010, 224, 527–539. [Google Scholar] [CrossRef]
- Fouda, M.A.; Ruben, P.C. Protein Kinases Mediate Anti-Inflammatory Effects of Cannabidiol and Estradiol Against High Glucose in Cardiac Sodium Channels. Front. Pharmacol. 2021, 12, 668657. [Google Scholar] [CrossRef]
- Hu, F.; Wang, Q.; Wang, P.; Wang, W.; Qian, W.; Xiao, H.; Wang, L. 17β-Estradiol regulates the gene expression of voltage-gated sodium channels: Role of estrogen receptor α and estrogen receptor β. Endocrine 2012, 41, 274–280. [Google Scholar] [CrossRef]
- Bi, R.-Y.; Meng, Z.; Zhang, P.; Wang, X.-D.; Ding, Y.; Gan, Y.-H. Estradiol upregulates voltage-gated sodium channel 1.7 in trigeminal ganglion contributing to hyperalgesia of inflamed TMJ. PLoS ONE 2017, 12, e0178589. [Google Scholar] [CrossRef] [Green Version]
- Kow, L.M.; Devidze, N.; Pataky, S.; Shibuya, I.; Pfaff, D.W. Acute estradiol application increases inward and decreases outward whole-cell currents of neurons in rat hypothalamic ventromedial nucleus. Brain Res. 2006, 1116, 1–11. [Google Scholar] [CrossRef]
- Gallos, G.; Yim, P.; Emala, C.W. Chloride in airway smooth muscle: The ignored anion no longer? Am. J. Physiol.-Lung Cell. Mol. Physiol. 2012, 302, L733–L735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulley, S.; Jaggar, J.H. Cl− channels in smooth muscle cells. Pflügers Arch.-Eur. J. Physiol. 2014, 466, 861–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssen, L.J.; Sims, S.M. Ca2+-dependent Cl− current in canine tracheal smooth muscle cells. Am. J. Physiol. 1995, 269, C163–C169. [Google Scholar] [CrossRef]
- Kotlikoff, M.I.; Wang, Y.-X. Calcium Release and Calcium-Activated Chloride Channels in Airway Smooth Muscle Cells. Am. J. Respir. Crit. Care Med. 1998, 158, S109–S114. [Google Scholar] [CrossRef] [PubMed]
- Danielsson, J.; Yim, P.; Rinderspacher, A.; Fu, X.W.; Zhang, Y.; Landry, D.W.; Emala, C.W. Chloride channel blockade relaxes airway smooth muscle and potentiates relaxation by β-agonists. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2014, 307, L273–L282. [Google Scholar] [CrossRef] [Green Version]
- Hirota, S.; Trimble, N.; Pertens, E.; Janssen, L.J. Intracellular Cl− fluxes play a novel role in Ca2+ handling in airway smooth muscle. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2006, 290, L1146–L1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallos, G.; Remy, K.E.; Danielsson, J.; Funayama, H.; Fu, X.W.; Chang, H.Y.; Yim, P.; Xu, D.; Emala, C.W., Sr. Functional expression of the TMEM16 family of calcium-activated chloride channels in airway smooth muscle. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2013, 305, L625–L634. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.; Zhang, H.; Wu, M.; Yang, H.; Kudo, M.; Peters, C.J.; Woodruff, P.G.; Solberg, O.D.; Donne, M.L.; Huang, X.; et al. Calcium-activated chloride channel TMEM16A modulates mucin secretion and airway smooth muscle contraction. Proc. Natl. Acad. Sci. USA 2012, 109, 16354–16359. [Google Scholar] [CrossRef] [Green Version]
- Danielsson, J.; Kuforiji, A.S.; Yocum, G.T.; Zhang, Y.; Xu, D.; Gallos, G.; Emala, C.W. Agonism of the TMEM16A calcium-activated chloride channel modulates airway smooth muscle tone. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2020, 318, L287–L295. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, M.; Takayama, Y.; Sunagawa, M. The Calcium-Activated Chloride Channel TMEM16A is Inhibitied by Liquiritigenin. Front. Pharmacol. 2021, 12, 628968. [Google Scholar] [CrossRef] [PubMed]
- Coakley, R.D.; Sun, H.; Clunes, L.A.; Rasmussen, J.E.; Stackhouse, J.R.; Okada, S.F.; Fricks, I.; Young, S.L.; Tarran, R. 17β-Estradiol inhibits Ca2+-dependent homeostasis of airway surface liquid volume in human cystic fibrosis airway epithelia. J. Clin. Investig. 2008, 118, 4025–4035. [Google Scholar] [CrossRef]
- Imberti, R.; Garavaglia, M.L.; Verduci, I.; Cannavale, G.; Balduzzi, G.; Papetti, S.; Mazzanti, M. Antiestrogen- and tamoxifen-induced effects on calcium-activated chloride currents in epithelial cells carrying the ∆F508-CFTR point mutation. Respir. Res. 2018, 19, 198. [Google Scholar] [CrossRef]
- Vandebrouck, C.; Melin, P.; Norez, C.; Robert, R.; Guibert, C.; Mettey, Y.; Becq, F. Evidence that CFTR is expressed in rat tracheal smooth muscle cells and contributes to bronchodilation. Respir. Res. 2006, 7, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michoud, M.-C.; Robert, R.; Hassan, M.; Moynihan, B.; Haston, C.; Govindaraju, V.; Ferraro, P.; Hanrahan, J.W.; Martin, J.G. Role of the Cystic Fibrosis Transmembrane Conductance Channel in Human Airway Smooth Muscle. Am. J. Respir. Cell Mol. Biol. 2009, 40, 217–222. [Google Scholar] [CrossRef]
- Cook, D.P.; Rector, M.V.; Bouzek, D.C.; Michalski, A.S.; Gansemer, N.D.; Reznikov, L.R.; Li, X.; Stroik, M.R.; Ostedgaard, L.S.; Abou Alaiwa, M.H.; et al. Cystic Fibrosis Transmembrane Conductance Regulator in Sarcoplasmic Reticulum of Airway Smooth Muscle. Implications for Airway Contractility. Am. J. Respir. Crit. Care Med. 2016, 193, 417–426. [Google Scholar] [CrossRef] [Green Version]
- Norez, C.; Jayle, C.; Becq, F.; Vandebrouck, C. Bronchorelaxation of the human bronchi by CFTR activators. Pulm. Pharmacol. Ther. 2014, 27, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Bazett, M.; Haston, C.K. Airway hyperresponsiveness in FVB/N delta F508 cystic fibrosis transmembrane conductance regulator mice. J. Cyst. Fibros. Off. J. Eur. Cyst. Fibros. Soc. 2014, 13, 378–383. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Lam, H.; Koziol-White, C.; Limjunyawong, N.; Kim, D.; Kim, N.; Karmacharya, N.; Rajkumar, P.; Firer, D.; Dalesio, N.M.; et al. The odorant receptor OR2W3 on airway smooth muscle evokes bronchodilation via a cooperative chemosensory tradeoff between TMEM16A and CFTR. Proc. Natl. Acad. Sci. USA 2020, 117, 28485–28495. [Google Scholar] [CrossRef]
- Johannesson, M.; Lúdvíksdóttir, D.; Janson, C. Lung function changes in relation to menstrual cycle in females with cystic fibrosis. Respir. Med. 2000, 94, 1043–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.K.; Schultz, B.D.; Katzenellenbogen, J.A.; Price, E.M.; Bridges, R.J.; Bradbury, N.A. Estrogen inhibition of cystic fibrosis transmembrane conductance regulator-mediated chloride secretion. J. Pharmacol. Exp. Ther. 2000, 295, 195–204. [Google Scholar] [PubMed]
- Ajonuma, L.C.; Tsang, L.L.; Zhang, G.H.; Wong, C.H.Y.; Lau, M.C.; Ho, L.S.; Rowlands, D.K.; Zhou, C.X.; Ng, C.P.; Chen, J.; et al. Estrogen-Induced Abnormally High Cystic Fibrosis Transmembrane Conductance Regulator Expression Results in Ovarian Hyperstimulation Syndrome. Mol. Endocrinol. 2005, 19, 3038–3044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, H.; Wen, G.; Deng, S.; Wan, S.; Xu, J.; Liu, X.; Xie, R.; Dong, H.; Tuo, B. Oestrogen upregulates the expression levels and functional activities of duodenal mucosal CFTR and SLC26A6. Exp. Physiol. 2016, 101, 1371–1382. [Google Scholar] [CrossRef] [Green Version]
- Jin, P.-Y.; Lu, Y.-C.; Li, L.; Han, Q.-F. Co action of CFTR and AQP1 increases permeability of peritoneal epithelial cells on estrogen-induced ovarian hyper stimulation syndrome. BMC Cell Biol. 2012, 13, 23. [Google Scholar] [CrossRef] [Green Version]
- Sweezey, N.B.; Gauthier, C.; Gagnon, S.; Ferretti, E.; Kopelman, H. Progesterone and estradiol inhibit CFTR-mediated ion transport by pancreatic epithelial cells. Am. J. Physiol. 1996, 271, G747–G754. [Google Scholar] [CrossRef]
- Goodstadt, L.; Powell, T.; Figtree, G.A. 17β-estradiol potentiates the cardiac cystic fibrosis transmembrane conductance regulator chloride current in guinea-pig ventricular myocytes. J. Physiol. Sci. 2006, 56, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Mizuta, K.; Xu, D.; Pan, Y.; Comas, G.; Sonett, J.R.; Zhang, Y.; Panettieri, R.A.; Yang, J.; Emala, C.W. GABA<sub>A</sub>receptors are expressed and facilitate relaxation in airway smooth muscle. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2008, 294, L1206–L1216. [Google Scholar] [CrossRef] [Green Version]
- Gallos, G.; Yim, P.; Chang, S.; Zhang, Y.; Xu, D.; Cook, J.M.; Gerthoffer, W.T.; Emala, C.W., Sr. Targeting the restricted α-subunit repertoire of airway smooth muscle GABAA receptors augments airway smooth muscle relaxation. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2012, 302, L248–L256. [Google Scholar] [CrossRef] [Green Version]
- Gallos, G.; Yocum, G.T.; Siviski, M.E.; Yim, P.D.; Fu, X.W.; Poe, M.M.; Cook, J.M.; Harrison, N.; Perez-Zoghbi, J.; Emala, C.W., Sr. Selective targeting of the α5-subunit of GABAA receptors relaxes airway smooth muscle and inhibits cellular calcium handling. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2015, 308, L931–L942. [Google Scholar] [CrossRef] [Green Version]
- Yocum, G.T.; Turner, D.L.; Danielsson, J.; Barajas, M.B.; Zhang, Y.; Xu, D.; Harrison, N.L.; Homanics, G.E.; Farber, D.L.; Emala, C.W. GABAA receptor α4 -subunit knockout enhances lung inflammation and airway reactivity in a murine asthma model. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2017, 313, L406–L415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbison, A.; Fenelon, V. Estrogen regulation of GABAA receptor subunit mRNA expression in preoptic area and bed nucleus of the stria terminalis of female rat brain. J. Neurosci. 1995, 15, 2328–2337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maggi, A.; Perez, J. Estrogen-induced up-regulation of gamma-aminobutyric acid receptors in the CNS of rodents. J. Neurochem. 1986, 47, 1793–1797. [Google Scholar] [CrossRef]
- François-Bellan, A.M.; Segu, L.; Héry, M. Regulation by estradiol of GABAA and GABAB binding sites in the diencephalon of the rat: An autoradiographic study. Brain Res. 1989, 503, 144–147. [Google Scholar] [CrossRef] [PubMed]
- Locci, A.; Porcu, P.; Talani, G.; Santoru, F.; Berretti, R.; Giunti, E.; Licheri, V.; Sanna, E.; Concas, A. Neonatal estradiol exposure to female rats changes GABAA receptor expression and function, and spatial learning during adulthood. Horm. Behav. 2017, 87, 35–46. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romero-Martínez, B.S.; Sommer, B.; Solís-Chagoyán, H.; Calixto, E.; Aquino-Gálvez, A.; Jaimez, R.; Gomez-Verjan, J.C.; González-Avila, G.; Flores-Soto, E.; Montaño, L.M. Estrogenic Modulation of Ionic Channels, Pumps and Exchangers in Airway Smooth Muscle. Int. J. Mol. Sci. 2023, 24, 7879. https://doi.org/10.3390/ijms24097879
Romero-Martínez BS, Sommer B, Solís-Chagoyán H, Calixto E, Aquino-Gálvez A, Jaimez R, Gomez-Verjan JC, González-Avila G, Flores-Soto E, Montaño LM. Estrogenic Modulation of Ionic Channels, Pumps and Exchangers in Airway Smooth Muscle. International Journal of Molecular Sciences. 2023; 24(9):7879. https://doi.org/10.3390/ijms24097879
Chicago/Turabian StyleRomero-Martínez, Bianca S., Bettina Sommer, Héctor Solís-Chagoyán, Eduardo Calixto, Arnoldo Aquino-Gálvez, Ruth Jaimez, Juan C. Gomez-Verjan, Georgina González-Avila, Edgar Flores-Soto, and Luis M. Montaño. 2023. "Estrogenic Modulation of Ionic Channels, Pumps and Exchangers in Airway Smooth Muscle" International Journal of Molecular Sciences 24, no. 9: 7879. https://doi.org/10.3390/ijms24097879