
Identification and Characterization of Proteins That Are Involved in RTP1S-Dependent Transport of Olfactory Receptors


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
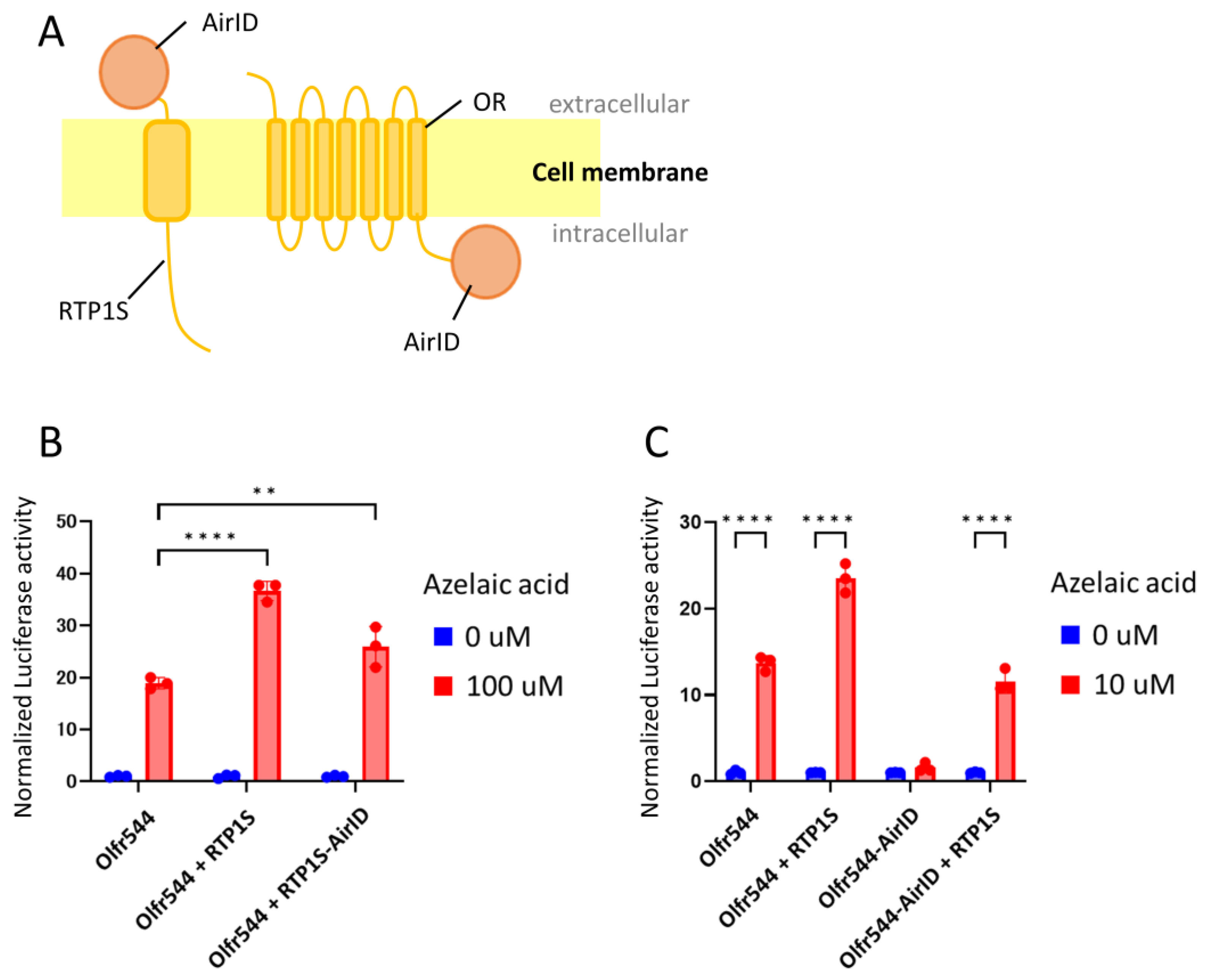
2.1. Design and Construction of RTP1S and OR Fused with Biotin Ligase
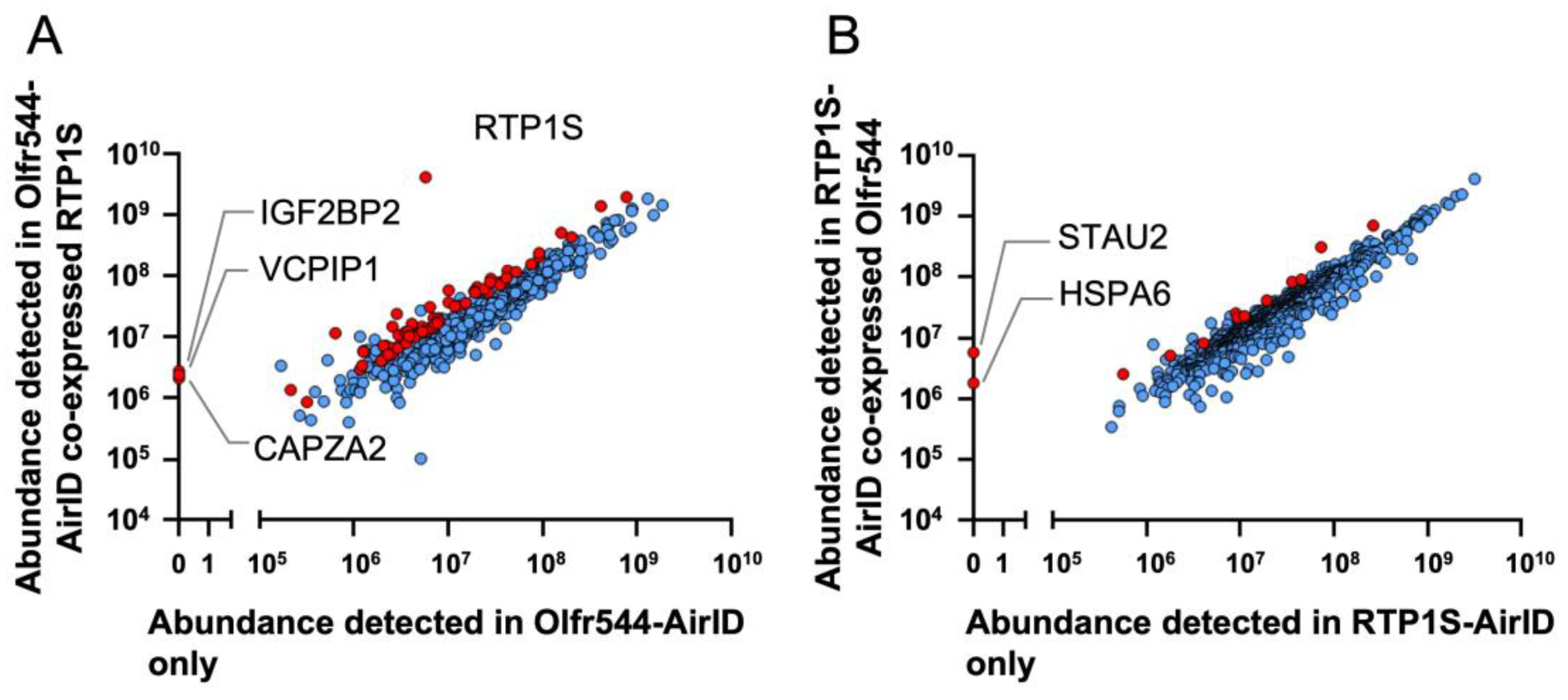
2.2. Proximity Labeling-Coupled Mass Spectrometry in HEK293T Cells
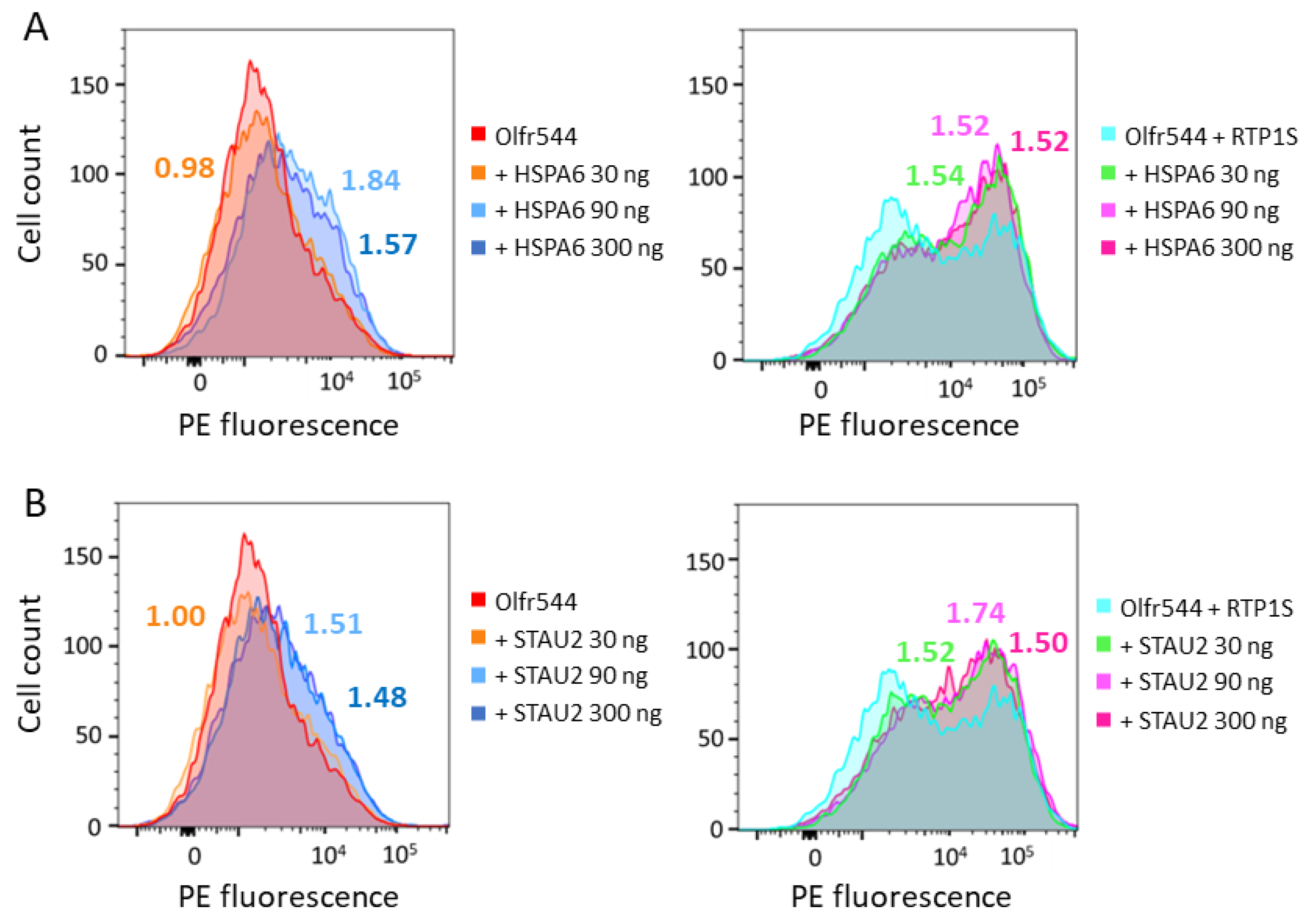
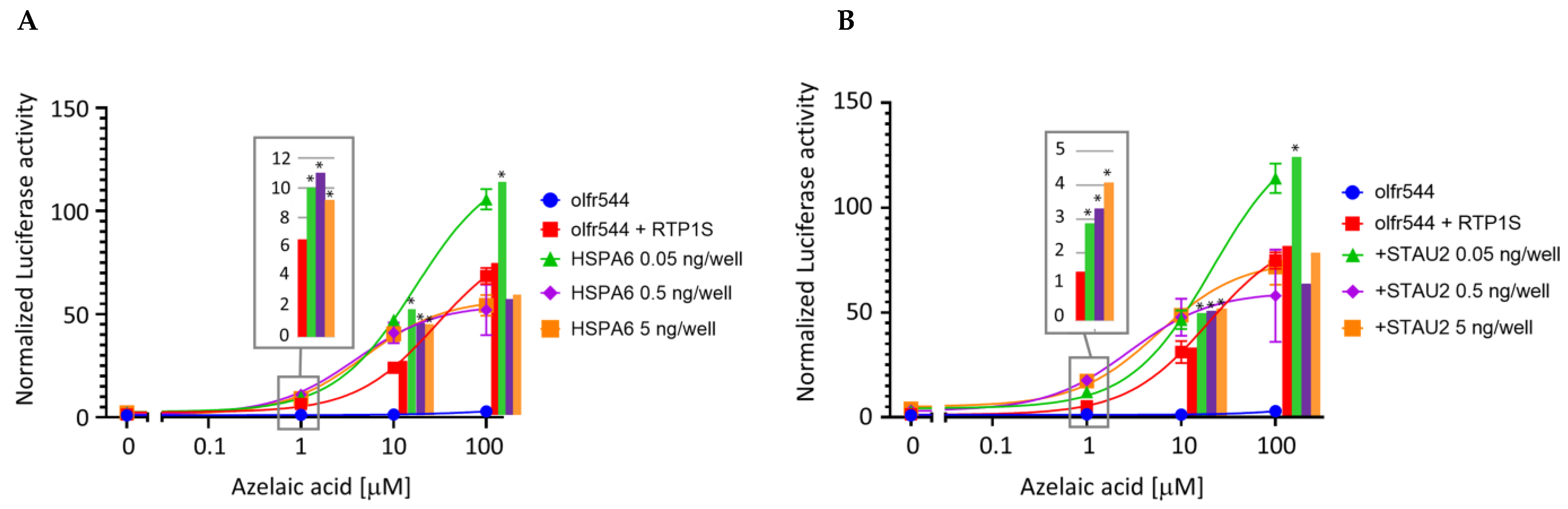
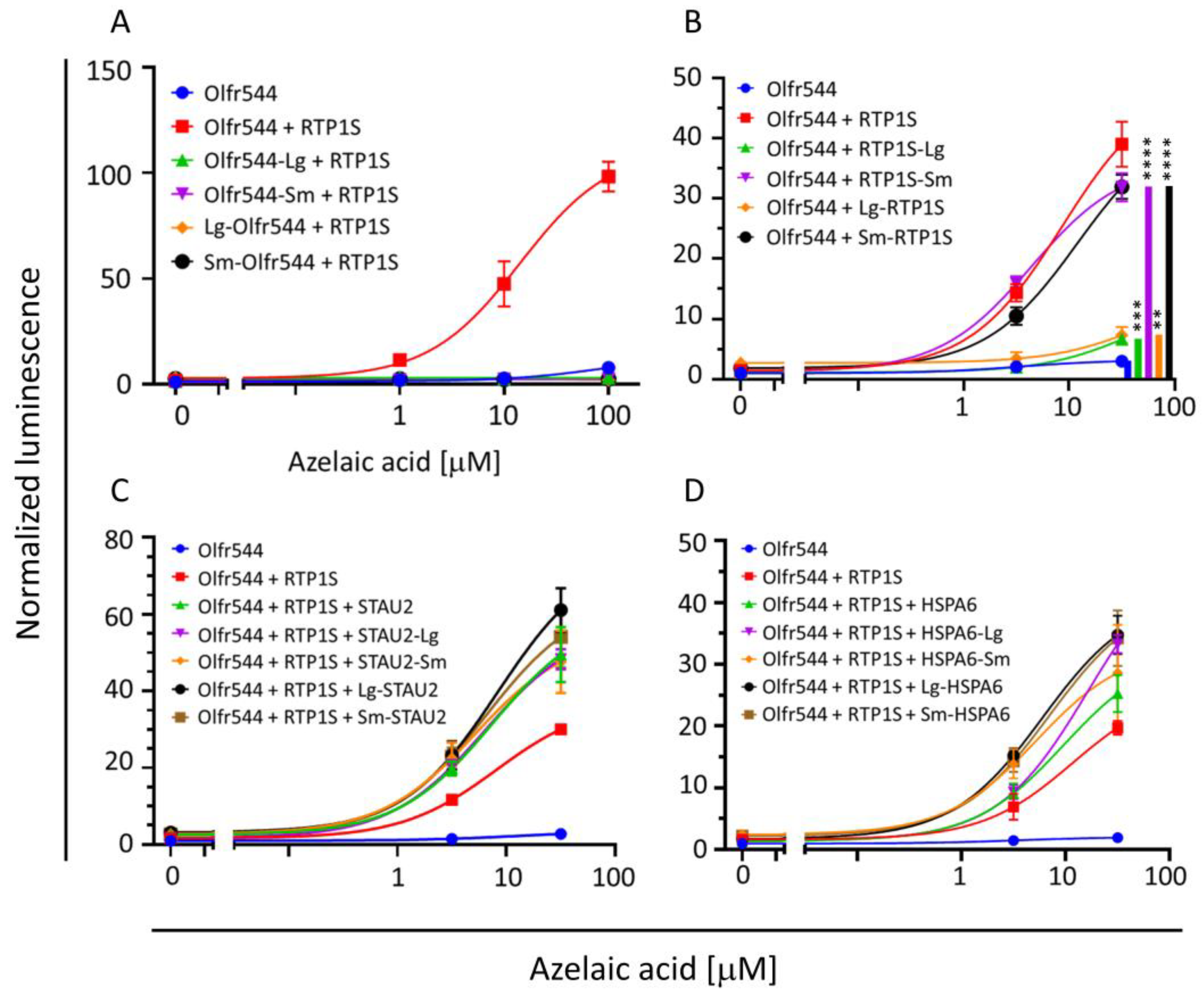
2.3. HSPA6 and STAU2 Facilitate Surface Expression and Ligand Response of Olfr544
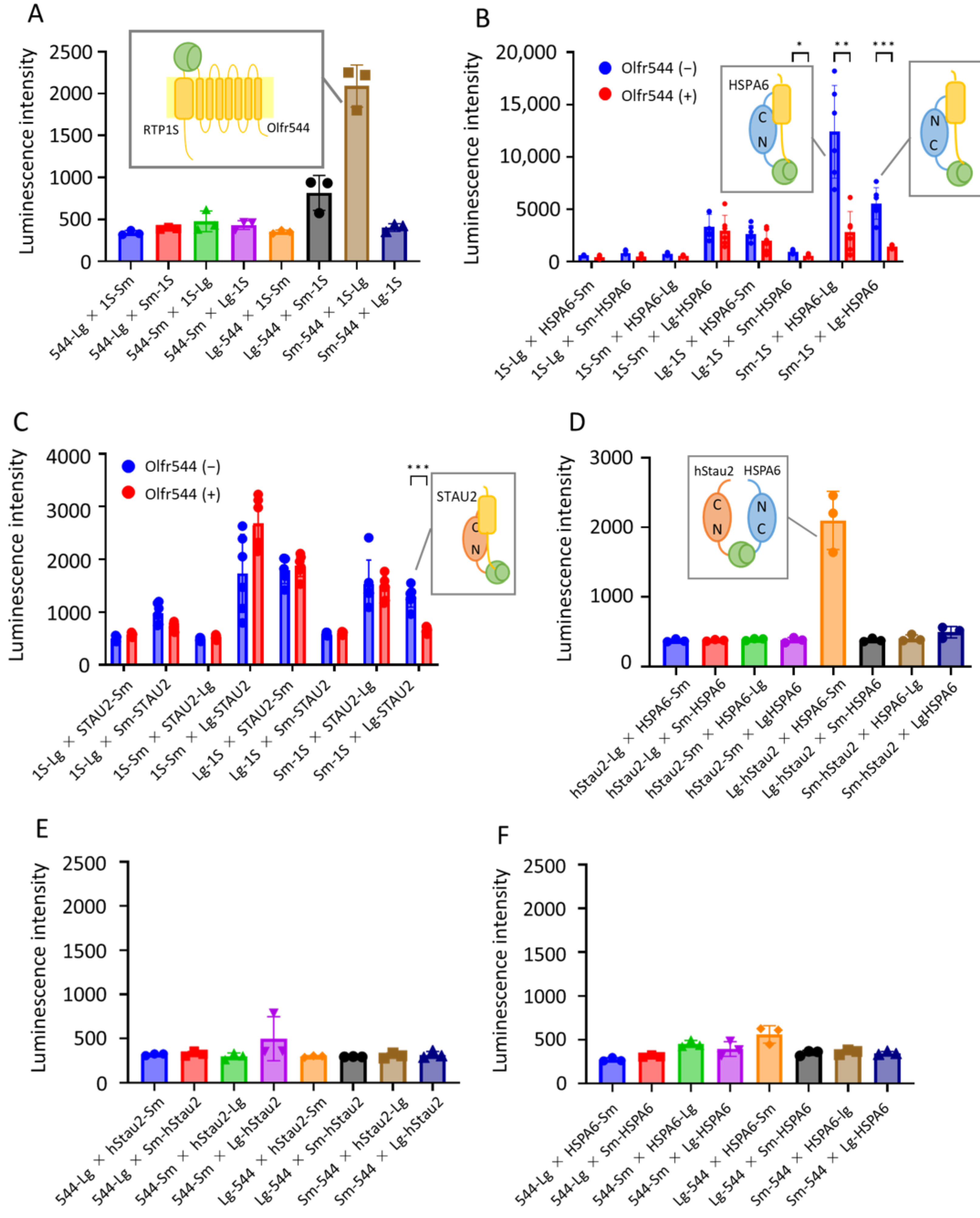
2.4. Intracellular Interaction Analysis Using NanoBiT Assay
3. Discussion
4. Materials and Methods
4.1. DNA and Vector Preparation
4.2. Cell Culture
4.3. Luciferase Assay
4.4. Affinity Purification of Biotinylated Proteins
4.5. MS Sample Preparation
4.6. LC-MS/MS Analysis
4.7. Data Analysis
4.8. Flow Cytometry Analyses
4.9. Split Luciferase Reconstruction Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buck, L.; Axel, R. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell 1991, 65, 175–187. [Google Scholar] [CrossRef]
- DeMaria, S.; Ngai, J. The cell biology of smell. J. Cell Biol. 2010, 191, 443–452. [Google Scholar] [CrossRef]
- Zou, D.J.; Chesler, A.; Firestein, S. How the olfactory bulb got its glomeruli: A just so story? Nat. Rev. Neurosci. 2009, 10, 611–618. [Google Scholar] [CrossRef]
- Niimura, Y. Evolutionary dynamics of olfactory receptor genes in chordates: Interaction between environments and genomic contents. Hum. Genom. 2009, 4, 107–118. [Google Scholar] [CrossRef]
- Zhang, X.; Firestein, S. The olfactory receptor gene superfamily of the mouse. Nat. Neurosci. 2002, 5, 124–133. [Google Scholar] [CrossRef]
- Serizawa, S.; Miyamichi, K.; Sakano, H. One neuron–one receptor rule in the mouse olfactory system. Trends Genet. 2004, 20, 648–653. [Google Scholar] [CrossRef]
- Lyons, D.B.; Allen, W.E.; Goh, T.; Tsai, L.; Barnea, G.; Lomvardas, S. An epigenetic trap stabilizes singular olfactory receptor expression. Cell 2013, 154, 325–336. [Google Scholar] [CrossRef]
- McClintock, T.S.; Landers, T.M.; Gimelbrant, A.A.; Fuller, L.Z.; Jackson, B.A.; Jayawickreme, C.K.; Lerner, M.R. Functional expression of olfactory–adrenergic receptor chimeras and intracellular retention of heterologously expressed olfactory receptors. Brain Res. Mol. Brain Res. 1997, 48, 270–278. [Google Scholar] [CrossRef]
- Lu, M.; Echeverri, F.; Moyer, B.D. Endoplasmic reticulum retention, degradation, and aggregation of olfactory G-protein–coupled receptors. Traffic 2003, 4, 416–433. [Google Scholar] [CrossRef]
- Saito, H.; Kubota, M.; Roberts, R.W.; Chi, Q.; Matsunami, H. RTP family members induce functional expression of mammalian odorant receptors. Cell 2004, 119, 679–691. [Google Scholar] [CrossRef]
- Zhuang, H.; Matsunami, H. Synergism of accessory factors in functional expression of mammalian odorant receptors. J. Biol. Chem. 2007, 282, 15284–15293. [Google Scholar] [CrossRef]
- Sharma, R.; Ishimaru, Y.; Davison, I.; Ikegami, K.; Chien, M.S.; You, H.; Chi, Q.; Kubota, M.; Yohda, M.; Ehlers, M.; et al. Olfactory receptor accessory proteins play crucial roles in receptor function and gene choice. Elife 2017, 6, e21895. [Google Scholar] [CrossRef]
- Fukutani, Y.; Hori, A.; Tsukada, S.; Sato, R.; Ishii, J.; Kondo, A.; Matsunami, H.; Yohda, M. Improving the odorant sensitivity of olfactory receptor-expressing yeast with accessory proteins. Anal. Biochem. 2015, 471, 1–8. [Google Scholar] [CrossRef]
- Saito, H.; Chi, Q.; Zhuang, H.; Matsunami, H.; Mainland, J.D. Odor coding by a mammalian receptor repertoire. Sci. Signal. 2009, 2, ra9. [Google Scholar] [CrossRef]
- Zhuang, H.; Matsunami, H. Evaluating cell-surface expression and measuring activation of mammalian odorant receptors in heterologous cells. Nat. Protoc. 2008, 3, 1402–1413. [Google Scholar] [CrossRef]
- Mainland, J.D.; Keller, A.; Li, Y.R.; Zhou, T.; Trimmer, C.; Snyder, L.L.; Moberly, A.H.; Adipietro, K.A.; Liu, W.L.; Zhuang, H.; et al. The missense of smell: Functional variability in the human odorant receptor repertoire. Nat. Neurosci. 2014, 17, 114–120. [Google Scholar] [CrossRef]
- Asakawa, M.; Fukutani, Y.; Savangsuksa, A.; Noguchi, K.; Matsunami, H.; Yohda, M. Modification of the response of olfactory receptorsto acetophenone by CYP1a2. Sci. Rep. 2017, 7, 10167. [Google Scholar] [CrossRef]
- Keller, A.; Zhuang, H.; Chi, Q.; Vosshall, L.B.; Matsunami, H. Genetic variation in a human odorant receptor alters odour perception. Nature 2007, 449, 468–472. [Google Scholar] [CrossRef]
- Behrens, M.; Bartelt, J.; Reichling, C.; Winnig, M.; Kuhn, C.; Meyerhof, W. Members of RTP and REEP gene families influence functional bitter taste receptor expression. J. Biol. Chem. 2006, 281, 20650–20659. [Google Scholar] [CrossRef]
- Décaillot, F.M.; Rozenfeld, R.; Gupta, A.; Devi, L.A. Cell surface targeting of opioid receptor heterodimers by RTP4. Proc. Natl. Acad. Sci. USA 2008, 105, 16045–16050. [Google Scholar] [CrossRef]
- Neuhaus, E.M.; Mashukova, A.; Zhang, W.; Barbour, J.; Hatt, H. A specific heat shock protein enhances the expression of mammalian olfactory receptor proteins. Chem. Senses 2006, 31, 445–452. [Google Scholar] [CrossRef]
- Wu, L.; Pan, Y.; Chen, G.Q.; Matsunami, H.; Zhuang, H. Receptor- transporting protein 1 short (RTP1S) mediates translocation and activation of odorant receptors by acting through multiple steps. J. Biol. Chem. 2012, 287, 22287–22294. [Google Scholar] [CrossRef]
- Fukutani, Y.; Tamaki, R.; Inoue, R.; Koshizawa, T.; Sakashita, S.; Ikegami, K.; Ohsawa, I.; Matsunami, H.; Yohda, M. The N-terminal region of RTP1S plays important roles in dimer formation and odorant receptor-trafficking. J. Biol. Chem. 2019, 294, 14661–14673. [Google Scholar] [CrossRef]
- Ikegami, K.; de March, C.A.; Nagai, M.H.; Ghosh, S.; Do, M.; Sharma, R.; Bruguera, E.S.; Lu, Y.E.; Fukutani, Y.; Vaidehi, N.; et al. Structural instability and divergence from conserved residues underlie intracellular retention of mammalian odorant receptors. Proc. Natl. Acad. Sci. USA 2020, 117, 2957–2967. [Google Scholar] [CrossRef]
- Kido, K.; Yamanaka, S.; Nakano, S.; Motani, K.; Shinohara, S.; Nozawa, A.; Kosako, H.; Ito, S.; Sawasaki, T. AirID, a novel proximity biotinylation enzyme, for analysis of protein-protein interactions. Elife 2020, 9, e54983. [Google Scholar] [CrossRef]
- Daugaard, M.; Rohde, M.; Jäättelä, M. The heat shock protein 70 family: Highly homologous proteins with overlapping and distinct functions. FEBS Lett. 2007, 581, 3702–3710. [Google Scholar] [CrossRef]
- St Johnston, D.; Beuchle, D.; Nüsslein-Volhard, C. Staufen, a gene required to localize maternal RNAs in the Drosophila egg. Cell 1991, 66, 51–63. [Google Scholar] [CrossRef]
- Schupbach, T.; Wieschaus, E. Germline autonomy of maternal-effect mutations altering the embryonic body pattern of Drosophila. Dev. Biol. 1986, 113, 443–448. [Google Scholar] [CrossRef]
- Micklem, D.R.; Adams, J.; Grünert, S.; St Johnston, D. Distinct roles of two conserved Staufen domains in oskar mRNA localization and translation. EMBO J. 2000, 19, 1366–1377. [Google Scholar] [CrossRef]
- Marión, R.M.; Fortes, P.; Beloso, A.; Dotti, C.; Ortín, J. A human sequence homologue of Staufen is an RNA-binding protein that is associated with polysomes and localizes to the rough endoplasmic reticulum. Mol. Cell. Biol. 1999, 19, 2212–2219. [Google Scholar] [CrossRef]
- Tang, S.J.; Meulemans, D.; Vazquez, L.; Colaco, N.; Schuman, E. A role for a rat homolog of staufen in the transport of RNA to neuronal dendrites. Neuron 2001, 32, 463–475. [Google Scholar] [CrossRef]
- Dixon, A.S.; Schwinn, M.K.; Hall, M.P.; Zimmerman, K.; Otto, P.; Lubben, T.H.; Butler, B.L.; Binkowski, B.F.; Machleidt, T.; Kirkland, T.A.; et al. NanoLuc Complementation Reporter Optimized for Accurate Measurement of Protein Interactions in Cells. ACS Chem. Biol. 2016, 11, 400–408. [Google Scholar] [CrossRef]
- Kuhlmann, K.; Tschapek, A.; Wiese, H.; Eisenacher, M.; Meyer, H.E.; Hatt, H.H.; Oeljeklaus, S.; Warscheid, B. The membrane proteome of sensory cilia to the depth of olfactory receptors. Mol Cell. Proteom. 2014, 13, 1828–1843. [Google Scholar] [CrossRef]
- Tholanikunnel, B.G.; Joseph, K.; Kandasamy, K.; Baldys, A.; Raymond, J.R.; Luttrell, L.M.; McDermott, P.J.; Fernandes, D.J. Novel mechanisms in the regulation of G protein-coupled receptor trafficking to the plasma membrane. J. Biol. Chem. 2010, 285, 33816–33825. [Google Scholar] [CrossRef]
- Almasi, S.; Jasmin, B.J. The multifunctional RNA-binding protein Staufen1: An emerging regulator of oncogenesis through its various roles in key cellular events. Cell Mol. Life Sci. 2021, 78, 7145–7160. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Inoue, R.; Fukutani, Y.; Niwa, T.; Matsunami, H.; Yohda, M. Identification and Characterization of Proteins That Are Involved in RTP1S-Dependent Transport of Olfactory Receptors. Int. J. Mol. Sci. 2023, 24, 7829. https://doi.org/10.3390/ijms24097829
Inoue R, Fukutani Y, Niwa T, Matsunami H, Yohda M. Identification and Characterization of Proteins That Are Involved in RTP1S-Dependent Transport of Olfactory Receptors. International Journal of Molecular Sciences. 2023; 24(9):7829. https://doi.org/10.3390/ijms24097829
Chicago/Turabian StyleInoue, Ryosuke, Yosuke Fukutani, Tatsuya Niwa, Hiroaki Matsunami, and Masafumi Yohda. 2023. "Identification and Characterization of Proteins That Are Involved in RTP1S-Dependent Transport of Olfactory Receptors" International Journal of Molecular Sciences 24, no. 9: 7829. https://doi.org/10.3390/ijms24097829