
Genome-Wide Analysis of the Expansin Gene Family in Populus and Characterization of Expression Changes in Response to Phytohormone (Abscisic Acid) and Abiotic (Low-Temperature) Stresses


Abstract
:1. Introduction
2. Results
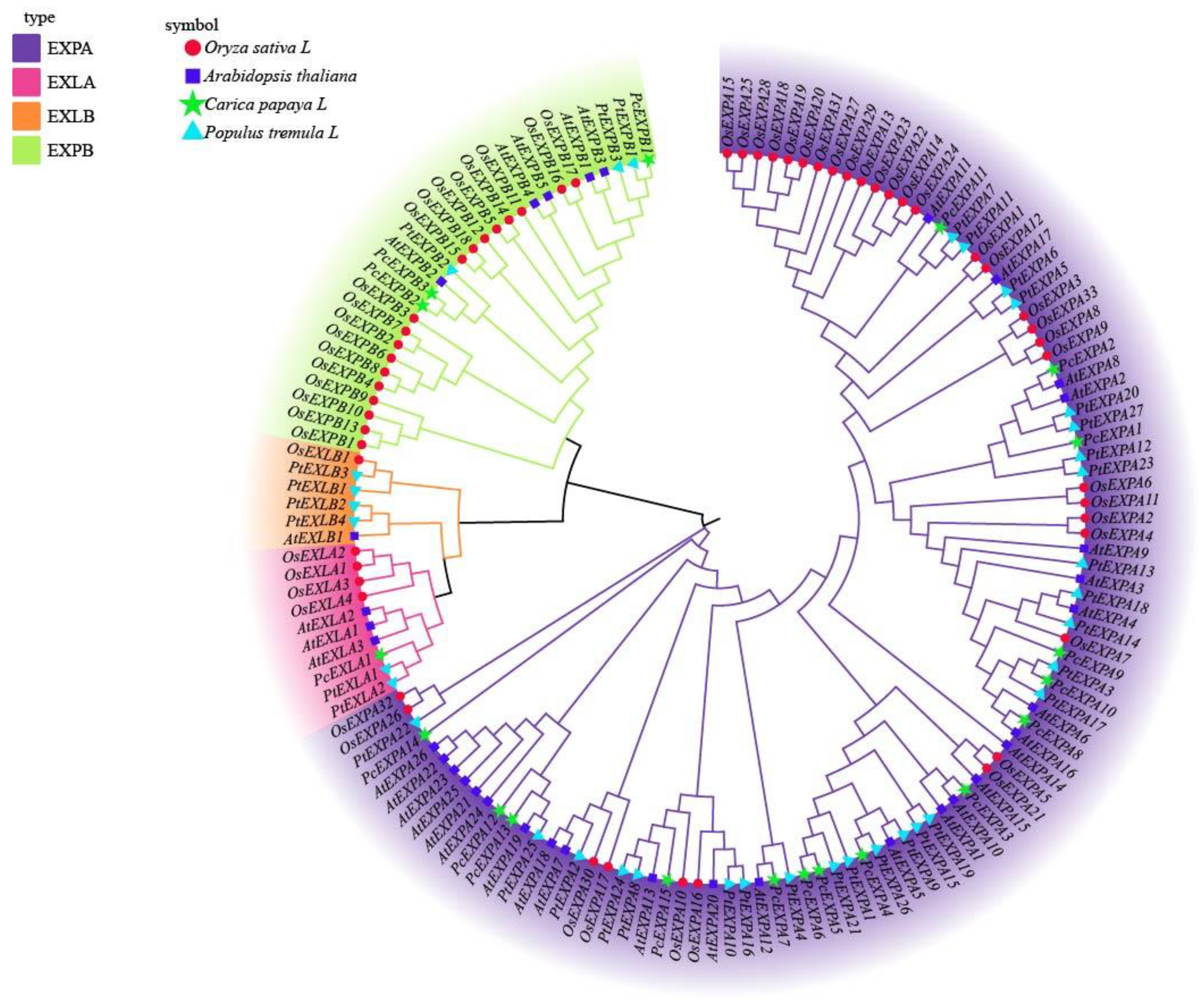
2.1. Identification of Expansin Genes in Populus and Phylogenetic Relationships
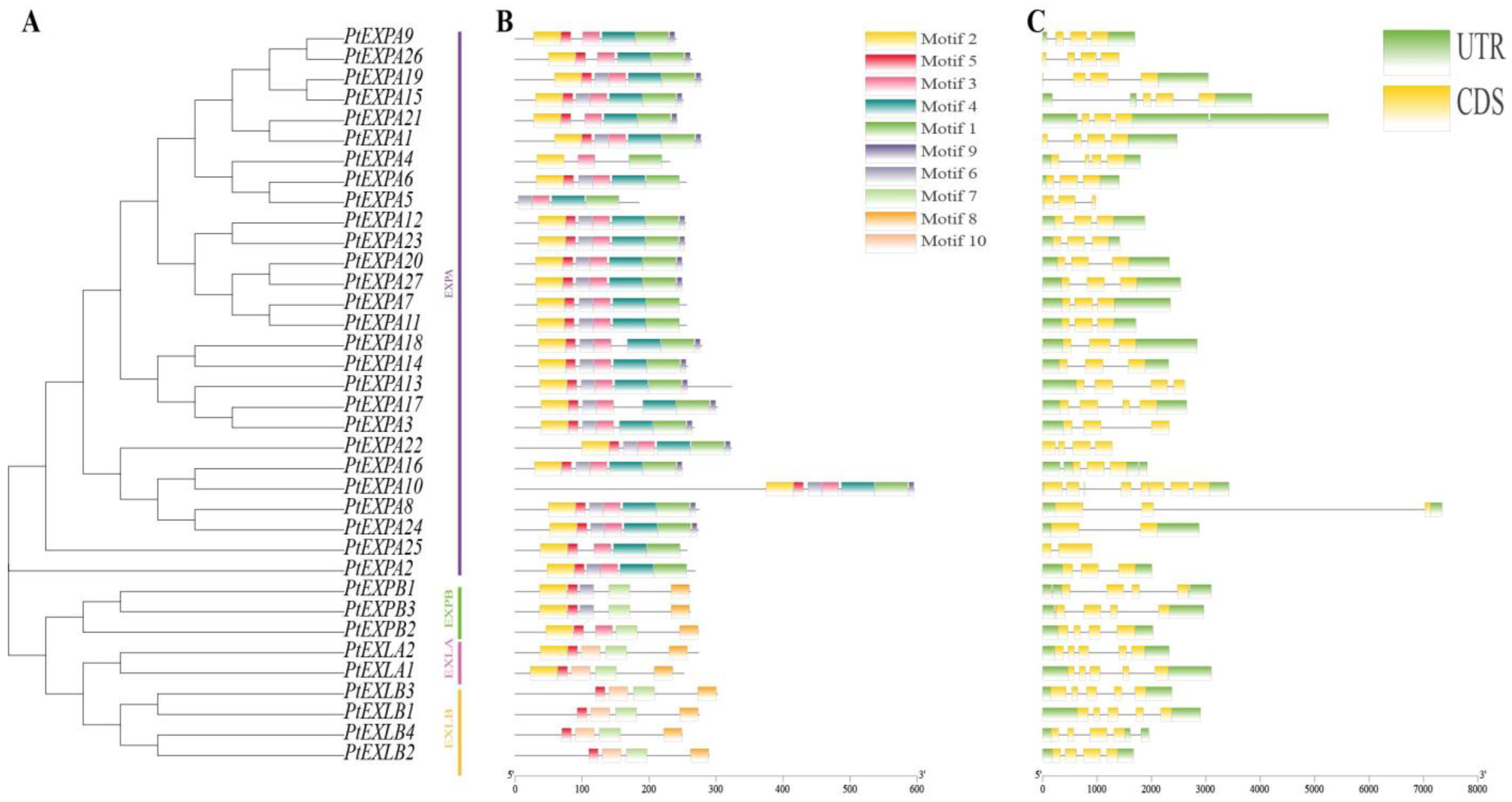
2.2. Gene Structures and Conserved Protein Motifs of PtEXs
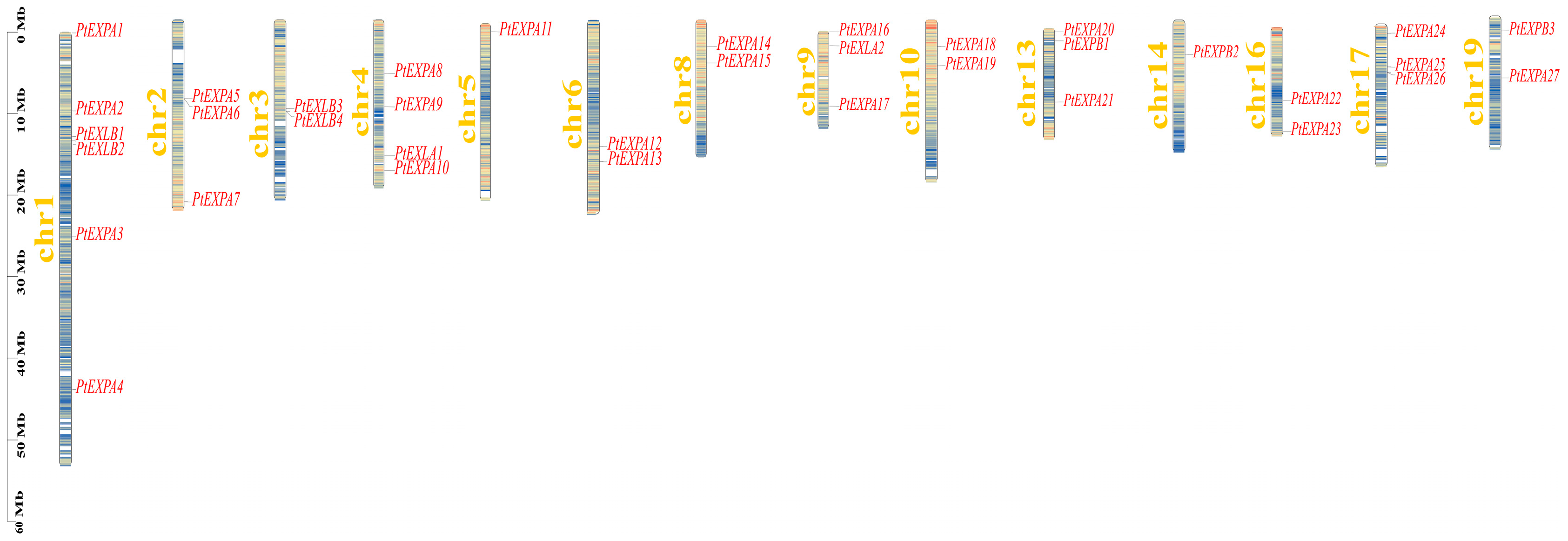
2.3. Chromosome Distributions of the PtEXs
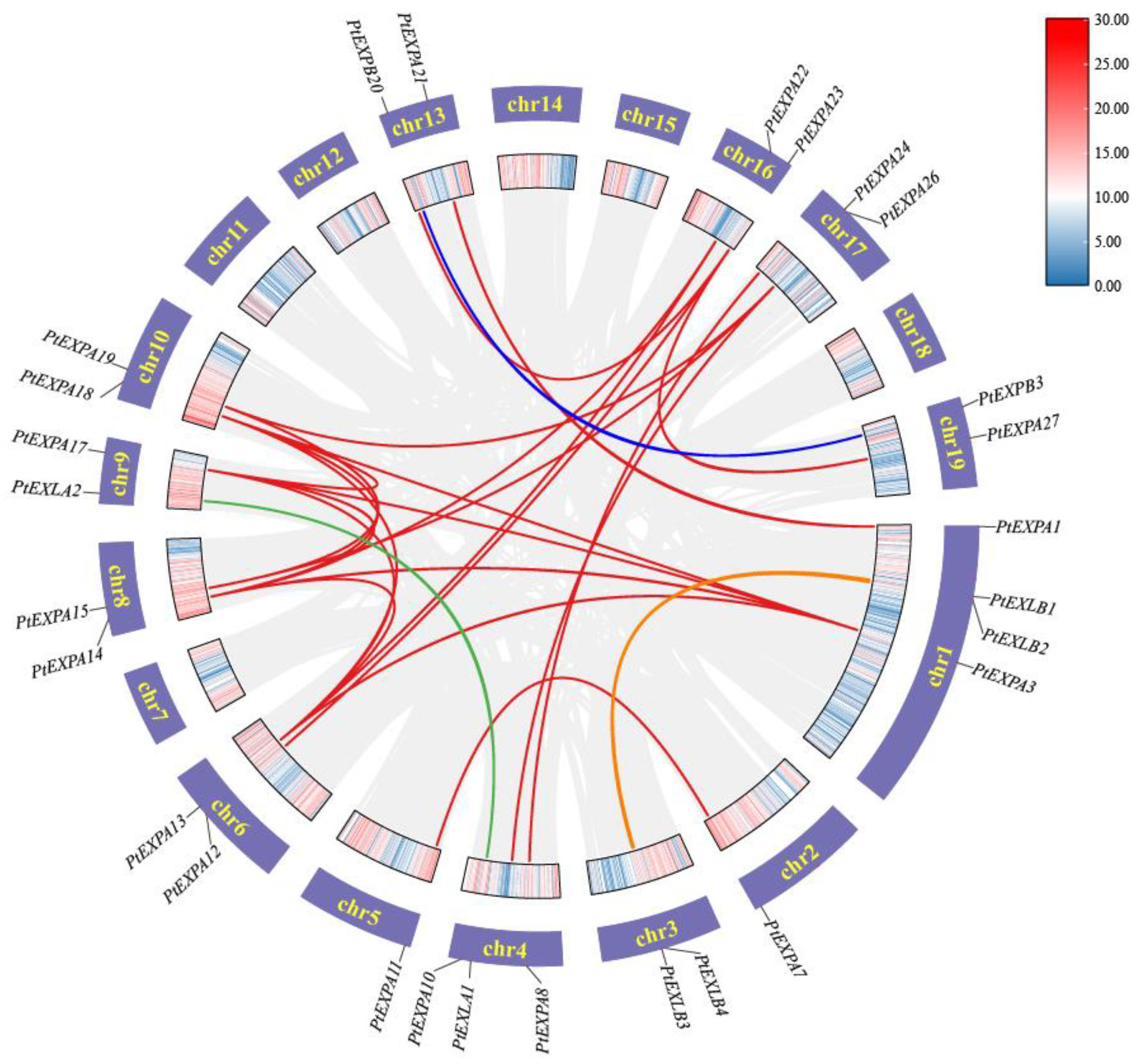
2.4. Duplication Events of Expansins
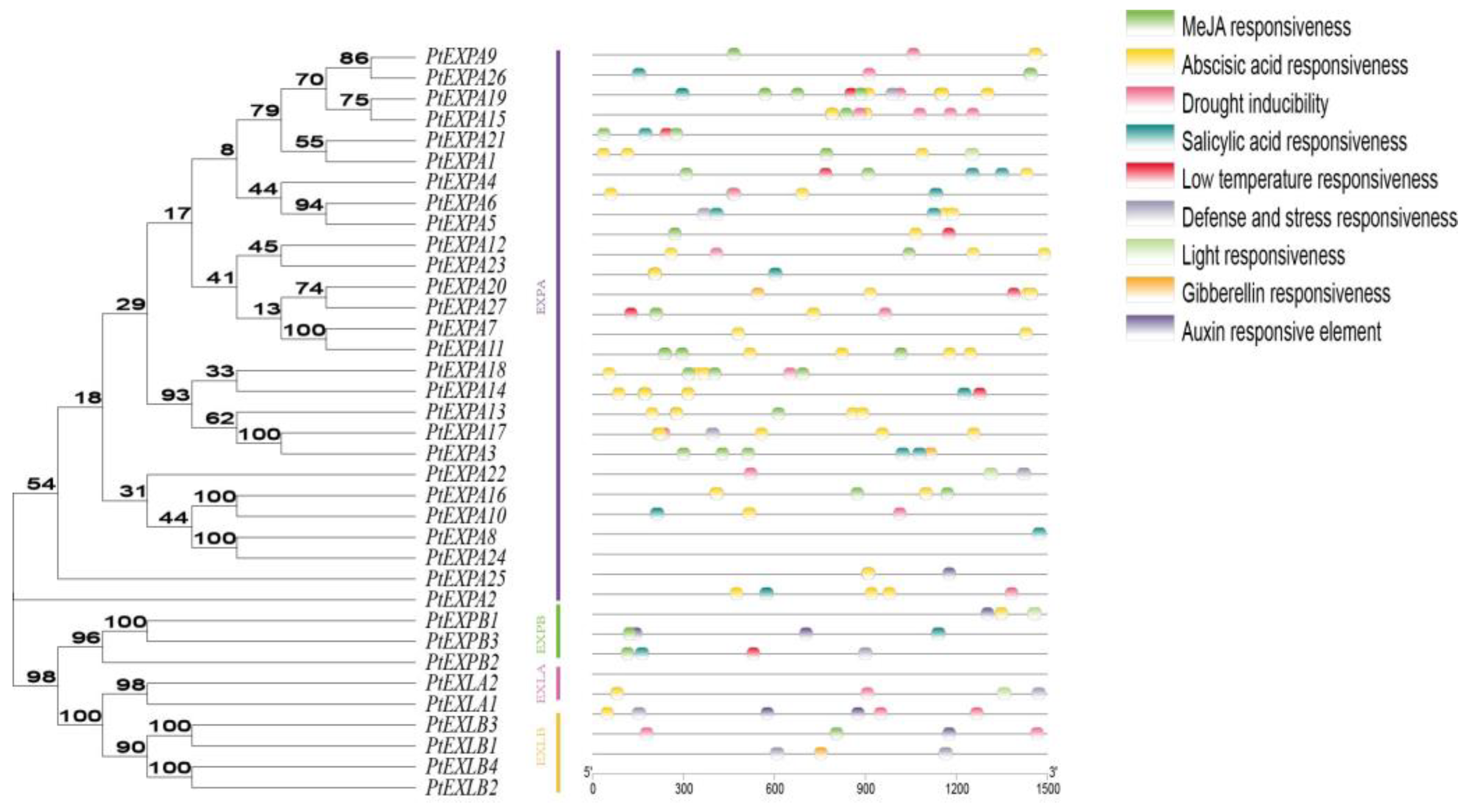
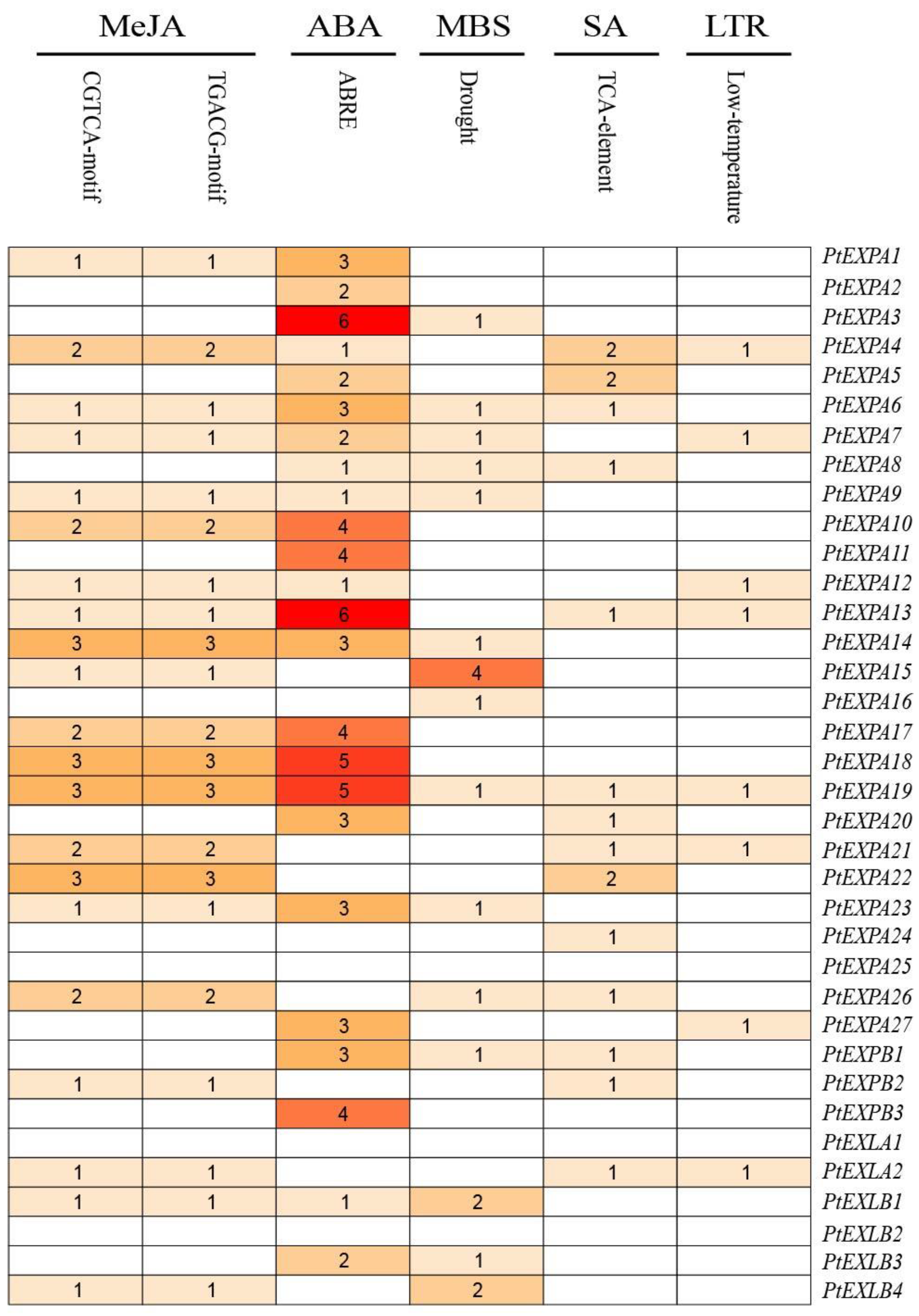
2.5. Analysis of Cis-Acting Elements
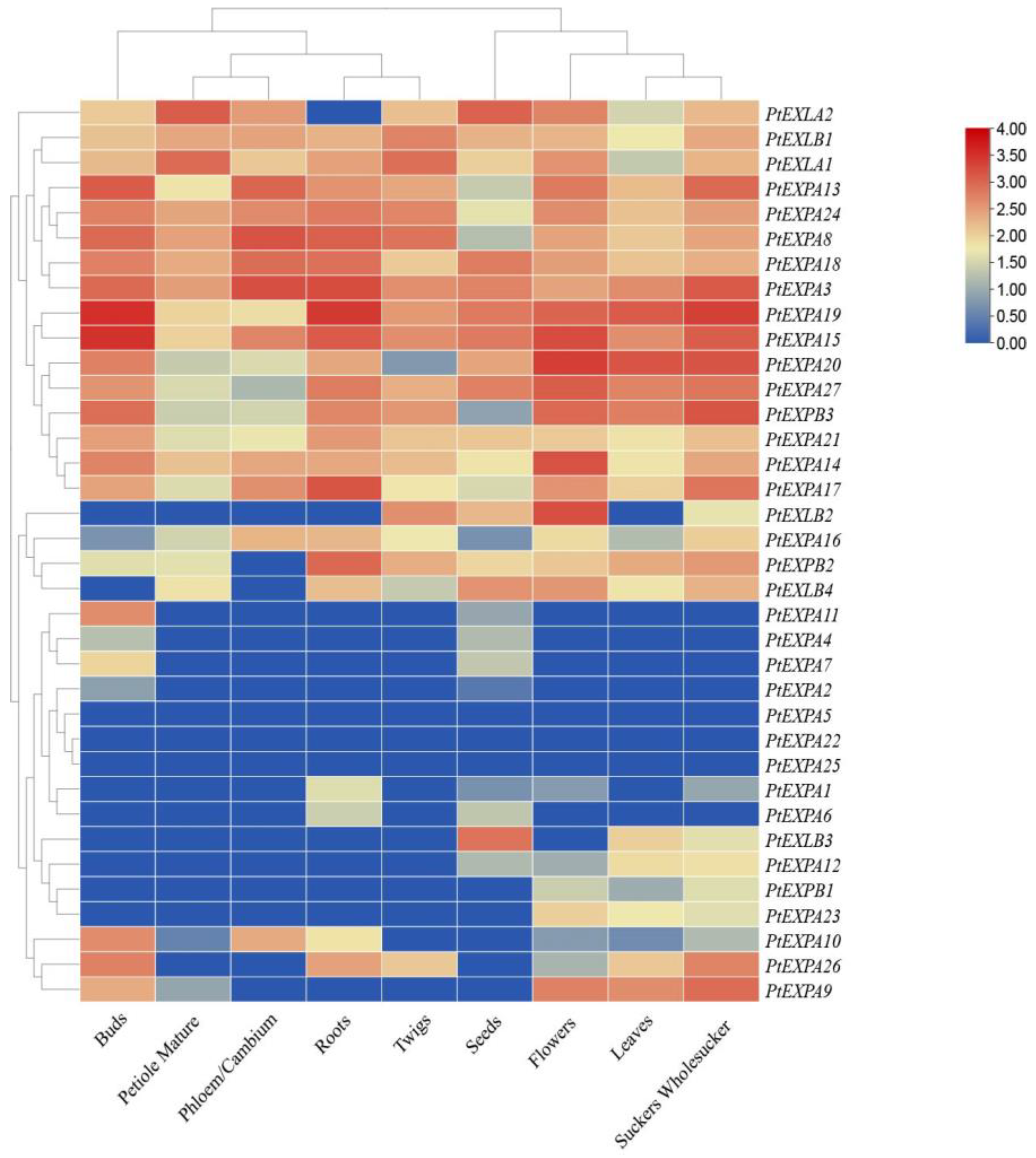
2.6. Analysis of PtEXs Expression in Populus
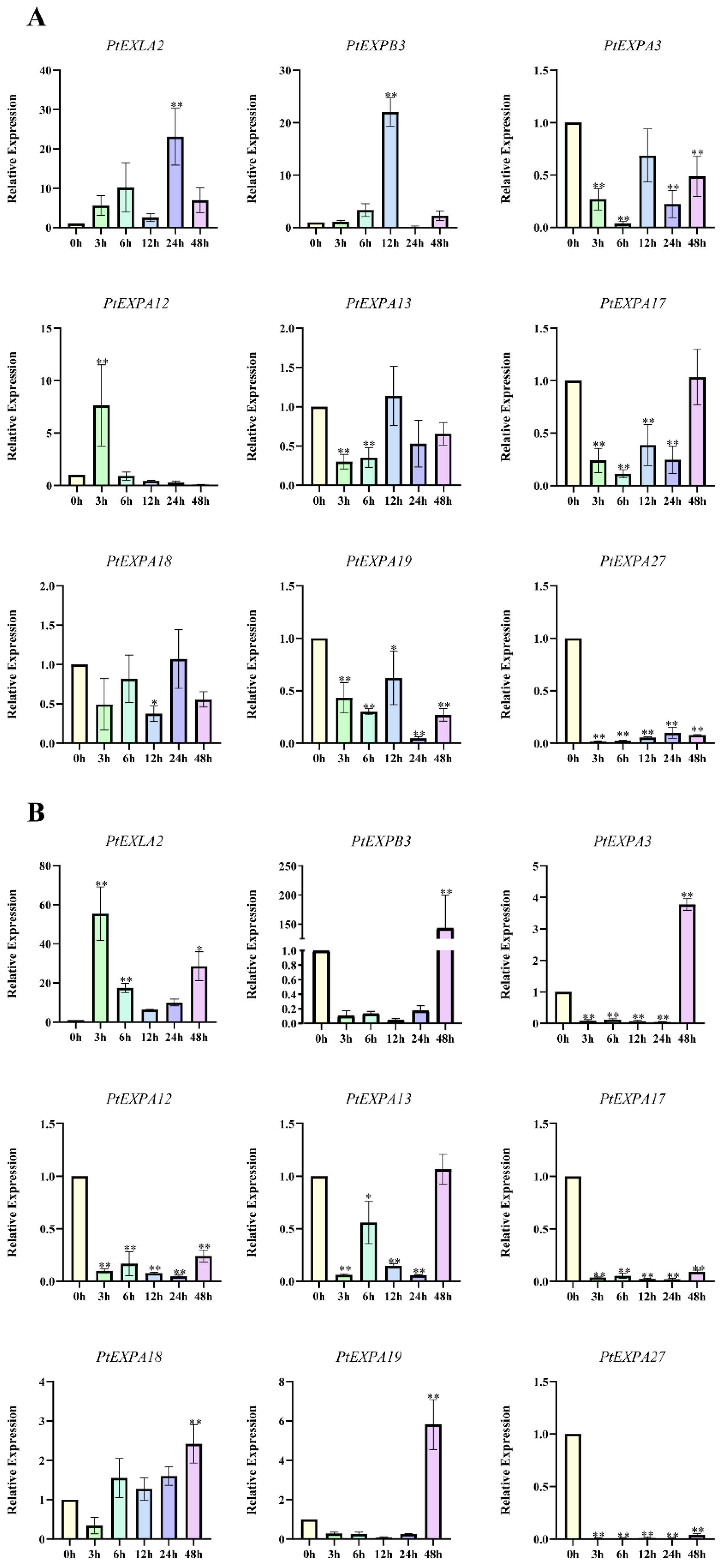
2.7. The Expression of PtEXs in Response to Phytohormone and Abiotic Stresses
3. Discussion
4. Materials and Methods
4.1. Identification of the Expansin Gene Family in Populus
4.2. Phylogenetic Analysis
4.3. Gene Structure, Chromosomal Locations and Cis-Regulatory Elements
4.4. Gene Duplication Analyses
4.5. Transcriptome Data Analysis
4.6. Plant Materials and Treatments
4.7. RNA Extraction and qRT-PCR Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McQueen-Mason, S.; Cosgrove, D.J. Disruption of hydrogen bonding between plant cell wall polymers by proteins that induce wall extension. Proc. Natl. Acad. Sci. USA 1994, 91, 6574–6578. [Google Scholar] [CrossRef] [Green Version]
- Cosgrove, D.J. Characterization of long-term extension of isolated cell walls from growing cucumber hypocotyls. Planta 1989, 177, 121–130. [Google Scholar] [CrossRef]
- Lee, Y.; Choi, D.; Kende, H. Expansins: Ever-expanding numbers and functions. Curr. Opin. Plant Biol. 2001, 4, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Georgelis, N.; Yennawar, N.H.; Cosgrove, D.J. Structural basis for entropy-driven cellulose binding by a type-A cellulose-binding module (CBM) and bacterial expansin. Proc. Natl. Acad. Sci. USA 2012, 109, 14830–14835. [Google Scholar] [CrossRef] [Green Version]
- Kende, H.; Bradford, K.; Brummell, D.; Cho, H.-T.; Cosgrove, D.; Fleming, A.; Gehring, C.; Lee, Y.; Mcqueen-Mason, S.; Rose, J.; et al. Nomenclature for members of the expansin superfamily of genes and proteins. Plant Mol. Biol. 2004, 55, 311–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, E.; Cosgrove, D.J. Expansins in growing tomato leaves. Plant J. 1995, 8, 795–802. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.-T.; Cosgrove, D.J. Altered expression of expansin modulates leaf growth and pedicel abscission in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2000, 97, 9783–9788. [Google Scholar] [CrossRef] [Green Version]
- Pien, S.; Wyrzykowska, J.; McQueen-Mason, S.; Smart, C.; Fleming, A. Local expression of expansin induces the entire process of leaf development and modifies leaf shape. Proc. Natl. Acad. Sci. USA 2001, 98, 11812–11817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.-T.; Kende, H. Expansins and Internodal Growth of Deepwater Rice. Plant Physiol. 1997, 113, 1145–1151. [Google Scholar] [CrossRef] [Green Version]
- Choi, D.; Lee, Y.; Cho, H.-T.; Kende, H. Regulation of expansin gene expression affects growth and development in transgenic rice plants. Plant Cell 2003, 15, 1386–1398. [Google Scholar] [CrossRef] [Green Version]
- Choi, D.; Cho, H.-T.; Lee, Y. Expansins: Expanding importance in plant growth and development. Physiol. Plant. 2006, 126, 511–518. [Google Scholar] [CrossRef]
- Cho, H.-T.; Cosgrove, D.J. Regulation of root hair initiation and expansin gene expression in arabidopsis. Plant Cell 2002, 14, 3237–3253. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Thorne, E.T.; Sharp, R.E.; Cosgrove, D.J. Modification of expansin transcript levels in the maize primary root at low water potentials. Plant Physiol. 2001, 126, 1471–1479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.; Zhao, J.; Li, X.; Qin, L.; Yan, X.; Liao, H. A soybean β-expansin gene GmEXPB2 intrinsically involved in root system architecture responses to abiotic stresses. Plant J. 2011, 66, 541–552. [Google Scholar] [CrossRef]
- Pezzotti, M.; Feron, R.; Mariani, C. Pollination modulates expression of the PPAL gene, a pistil-specific β-expansin. Plant Mol. Biol. 2002, 49, 187–197. [Google Scholar] [CrossRef]
- Rose, J.K.C.; Lee, H.H.; Bennett, A.B. Expression of a divergent expansin gene is fruit-specific and ripening-regulated. Proc. Natl. Acad. Sci. USA 1997, 94, 5955–5960. [Google Scholar] [CrossRef] [Green Version]
- Geilfus, C.-M.; Zörb, C.; Mühling, K.H. Salt stress differentially affects growth-mediating β-expansins in resistant and sensitive maize (Zea mays L.). Plant Physiol. Biochem. 2010, 48, 993–998. [Google Scholar] [CrossRef]
- Han, Y.Y.; Li, A.X.; Li, F.; Zhao, M.R.; Wang, W. Characterization of a wheat (Triticum aestivum L.) expansin gene, TaEXPB23, involved in the abiotic stress response and phytohormone regulation. Plant Physiol. Biochem. 2012, 54, 49–58. [Google Scholar] [CrossRef]
- Kuluev, B.; Avalbaev, A.; Mikhaylova, E.; Nikonorov, Y.; Berezhneva, Z.; Chemeris, A. Expression profiles and hormonal regulation of tobacco expansin genes and their involvement in abiotic stress response. J. Plant Physiol. 2016, 206, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Dotto, M.; Pombo, M.A.; Martínez, G.A.; Civello, P.M. Heat treatments and expansin gene expression in strawberry fruit. Sci. Hortic. 2011, 130, 775–780. [Google Scholar] [CrossRef]
- Sampedro, J.; Cosgrove, D.J. The expansin superfamily. Genome Biol. 2005, 6, 242. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Wu, N.; Song, W.; Yin, G.; Qin, Y.; Yan, Y.; Hu, Y. Soybean (Glycine max) expansin gene superfamily origins: Segmental and tandem duplication events followed by divergent selection among subfamilies. BMC Plant Biol. 2014, 14, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.-C.; Wang, J.; Delhomme, N.; Schiffthaler, B.; Sundström, G.; Zuccolo, A.; Nystedt, B.; Hvidsten, T.R.; De La Torre, A.; Cossu, R.M.; et al. Functional and evolutionary genomic inferences in Populus through genome and population sequencing of American and European aspen. Proc. Natl. Acad. Sci. USA 2018, 115, E10970–E10978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, F.; Chen, Y.; Wu, H.; Yin, T. Genome-Wide Comparative Analysis of R2R3 MYB Gene Family in Populus and Salix and Identification of Male Flower Bud Development-Related Genes. Front. Plant Sci. 2021, 12, 721558. [Google Scholar] [CrossRef] [PubMed]
- Biselli, C.; Vietto, L.; Rosso, L.; Cattivelli, L.; Nervo, G.; Fricano, A. Advanced Breeding for Biotic Stress Resistance in Poplar. Plants 2022, 11, 2032. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, F.; Wang, H.; Chen, Y.; Yin, T.; Wu, H. Genome-Wide Comparative Analysis of the Fasciclin-like Arabinogalactan Proteins (FLAs) in Salicacea and Identification of Secondary Tissue Development-Related Genes. Int. J. Mol. Sci. 2023, 24, 1481. [Google Scholar] [CrossRef]
- Santner, A.; A Calderon-Villalobos, L.I.; Estelle, M. Plant hormones are versatile chemical regulators of plant growth. Nat. Chem. Biol. 2009, 5, 301–307. [Google Scholar] [CrossRef]
- Popko, J.; Hänsch, R.; Mendel, R.-R.; Polle, A.; Teichmann, T. The role of abscisic acid and auxin in the response of poplar to abiotic stress. Plant Biol. 2010, 12, 242–258. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Su, H.; Cao, H.; Wei, H.; Fu, X.; Jiang, X.; Song, Q.; He, X.; Xu, C.; Luo, K. AUXIN RESPONSE FACTOR7 integrates gibberellin and auxin signaling via interactions between DELLA and AUX/IAA proteins to regulate cambial activity in poplar. Plant Cell 2022, 34, 2688–2707. [Google Scholar] [CrossRef] [PubMed]
- Ullah, C.; Tsai, C.; Unsicker, S.B.; Xue, L.; Reichelt, M.; Gershenzon, J.; Hammerbacher, A. Salicylic acid activates poplar defense against the biotrophic rust fungus Melampsora larici-populina via increased biosynthesis of catechin and proanthocyanidins. New Phytol. 2019, 221, 960–975. [Google Scholar] [CrossRef] [Green Version]
- Mauriat, M.; Petterle, A.; Bellini, C.; Moritz, T. Gibberellins inhibit adventitious rooting in hybrid aspen and Arabidopsis by affecting auxin transport. Plant J. 2014, 78, 372–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Shao, Y.; Chaffin, T.A.; Lee, J.H.; Poindexter, M.R.; Ahkami, A.H.; Blumwald, E.; Stewart, C.N.J. Performance of abiotic stress-inducible synthetic promoters in genetically engineered hybrid poplar (Populus tremula × Populus alba). Front. Plant Sci. 2022, 13, 1011939. [Google Scholar] [CrossRef]
- Harfouche, A.; Meilan, R.; Altman, A. Molecular and physiological responses to abiotic stress in forest trees and their relevance to tree improvement. Tree Physiol. 2014, 34, 1181–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, N.; Li, Z.; Wu, F.; Tang, M. Comparative photochemistry activity and antioxidant responses in male and female Populus cathayana cuttings inoculated with arbuscular mycorrhizal fungi under salt. Sci. Rep. 2016, 6, 37663. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Kang, Y.; Leng, J.; Xu, Q. Genome-Wide Analysis of the miRNA–mRNAs Network Involved in Cold Tolerance in Populus simonii × P. nigra. Genes 2019, 10, 430. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Duan, B.; Chen, J.; Korpelainen, H.; Niinemets, Ü.; Li, C. Males exhibit competitive advantages over females of Populus deltoides under salinity stress. Tree Physiol. 2016, 36, 1573–1584. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-Q.; Wei, P.-C.; Xiong, Y.-M.; Yang, Y.; Chen, J.; Wang, X.-C. Overexpression of the Arabidopsis α-expansin gene AtEXPA1 accelerates stomatal opening by decreasing the volumetric elastic modulus. Plant Cell Rep. 2011, 30, 27–36. [Google Scholar] [CrossRef]
- Lee, H.W.; Kim, J. EXPANSINA17 up-regulated by LBD18/ASL20 promotes lateral root formation during the auxin response. Plant Cell Physiol. 2013, 54, 1600–1611. [Google Scholar] [CrossRef] [Green Version]
- Goh, H.-H.; Sloan, J.; Dorca-Fornell, M.D.C.; Fleming, A. Inducible repression of multiple expansin genes leads to growth suppression during leaf development. Plant Physiol. 2012, 159, 1759–1770. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.M.; Kang, B.; He, X.W.; Lv, S.L.; Bai, Y.H.; Ding, W.N.; Chen, M.; Cho, H.-T.; Wu, P. Root hair-specific expansins modulate root hair elongation in rice. Plant J. 2011, 66, 725–734. [Google Scholar] [CrossRef]
- Bai, S.; Wu, H.; Zhang, J.; Pan, Z.; Zhao, W.; Li, Z.; Tong, C. Genome Assembly of Salicaceae Populus deltoides (Eastern Cottonwood) I-69 Based on Nanopore Sequencing and Hi-C Technologies. J. Hered. 2021, 112, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Yao, D.; Chen, Y.; Yang, W.; Zhao, W.; Gao, H.; Tong, C. De Novo Genome Assembly of Populus simonii Further Supports That Populus simonii and Populus trichocarpa Belong to Different Sections. G3 Genes Genomes Genet. 2020, 10, 455–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sjödin, A.; Street, N.R.; Sandberg, G.; Gustafsson, P.; Jansson, S. The Populus Genome Integrative Explorer (PopGenIE): A new resource for exploring the Populus genome. New Phytol. 2009, 182, 1013–1025. [Google Scholar] [CrossRef]
- Sampedro, J.; Lee, Y.; Carey, R.E.; DePamphilis, C.; Cosgrove, D.J. Use of genomic history to improve phylogeny and understanding of births and deaths in a gene family. Plant J. 2005, 44, 409–419. [Google Scholar] [CrossRef]
- Hou, L.; Zhang, Z.; Dou, S.; Zhang, Y.; Pang, X.; Li, Y. Genome-wide identification, characterization, and expression analysis of the expansin gene family in Chinese jujube (Ziziphus jujuba Mill.). Planta 2019, 249, 815–829. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xu, R.; Gao, Z.; Chen, C.; Jiang, Z.; Shu, H. A genome-wide analysis of the expansin genes in Malus × Domestica. Mol. Genet. Genom. 2014, 289, 225–236. [Google Scholar] [CrossRef]
- Zhang, W.; Yan, H.; Chen, W.; Liu, J.; Jiang, C.; Jiang, H.; Zhu, S.; Cheng, B. Genome-wide identification and characterization of maize expansin genes expressed in endosperm. Mol. Genet. Genom. 2014, 289, 1061–1074. [Google Scholar] [CrossRef]
- Feng, X.; Li, C.; He, F.; Xu, Y.; Li, L.; Wang, X.; Chen, Q.; Li, F. Genome-Wide Identification of Expansin Genes in Wild Soybean (Glycine soja) and Functional Characterization of Expansin B1 (GsEXPB1) in Soybean Hair Root. Int. J. Mol. Sci. 2022, 23, 5407. [Google Scholar] [CrossRef]
- Santo, S.D.; Vannozzi, A.; Tornielli, G.B.; Fasoli, M.; Venturini, L.; Pezzotti, M.; Zenoni, S. Genome-wide analysis of the expansin gene superfamily reveals grapevine-specific structural and functional characteristics. PLoS ONE 2013, 8, e62206. [Google Scholar] [CrossRef]
- Lu, Y.; Liu, L.; Wang, X.; Han, Z.; Ouyang, B.; Zhang, J.; Li, H. Genome-wide identification and expression analysis of the expansin gene family in tomato. Mol. Genet. Genom. 2016, 291, 597–608. [Google Scholar] [CrossRef]
- Ding, A.; Marowa, P.; Kong, Y. Genome-wide identification of the expansin gene family in tobacco (Nicotiana tabacum). Mol. Genet. Genom. 2016, 291, 1891–1907. [Google Scholar] [CrossRef]
- Moore, R.C.; Purugganan, M.D. The early stages of duplicate gene evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Han, B.; Jiao, Y. Genetic Contribution of Paleopolyploidy to Adaptive Evolution in Angiosperms. Mol. Plant 2020, 13, 59–71. [Google Scholar] [CrossRef]
- Ma, Z.; Bielenberg, D.G.; Brown, K.M.; Lynch, J.P. Regulation of root hair density by phosphorus availability in Arabidopsis thaliana. Plant Cell Environ. 2001, 24, 459–467. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.-K.; Ahn, J.H.; Song, S.-K.; Choi, Y.D.; Lee, J.S. Expression of an expansin gene is correlated with root elongation in soybean. Plant Physiol. 2003, 131, 985–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lü, S.; Gu, H.; Yuan, X.; Wang, X.; Wu, A.-M.; Qu, L.; Liu, J.-Y. The GUS reporter-aided analysis of the promoter activities of a rice metallothionein gene reveals different regulatory regions responsible for tissue-specific and inducible expression in transgenic Arabidopsis. Transgenic Res. 2007, 16, 177–191. [Google Scholar] [CrossRef] [PubMed]
- Walther, D.; Brunnemann, R.; Selbig, J. The regulatory code for transcriptional response diversity and its relation to genome structural properties in a. thaliana. PLoS Genet. 2007, 3, e11. [Google Scholar] [CrossRef]
- Le, D.T.; Nishiyama, R.; Watanabe, Y.; Vankova, R.; Tanaka, M.; Seki, M.; Ham, L.H.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.-S.P. Identification and expression analysis of cytokinin metabolic genes in soybean under normal and drought conditions in relation to cytokinin levels. PLoS ONE 2012, 7, e42411. [Google Scholar] [CrossRef] [Green Version]
- Hutchison, K.W.; Singer, P.B.; McInnis, S.; Diaz-Sala, C.; Greenwood, M.S. Expansins are conserved in conifers and expressed in hypocotyls in response to exogenous auxin1. Plant Physiol. 1999, 120, 827–832. [Google Scholar] [CrossRef] [Green Version]
- Vriezen, W.H.; De Graaf, B.; Mariani, C.; Voesenek, L. Submergence induces expansin gene expression in flooding-tolerant Rumex palustris and not in flooding-intolerant R. acetosa. Planta 2000, 210, 956–963. [Google Scholar] [CrossRef]
- Xue, Z.; Kou, X.; Luo, Y.; Zhu, B.; Xu, W. Effect of ethylene on polygalacturonase, lipoxygenase and expansin in ripening of tomato fruits. Trans. Tianjin Univ. 2009, 15, 173–177. [Google Scholar] [CrossRef]
- Lynch, M.; Conery, J.S. The evolutionary fate and consequences of duplicate genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.; Wu, M.; Liu, H.; Gao, Y.; Chen, J.; Yan, H.; Xiang, Y. Genome-wide identification and expression analysis of the NF-Y transcription factor family in Populus. Physiol. Plant. 2021, 171, 309–327. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wang, W.; Wang, D.; Hu, S.; Zhang, Q.; Wang, Z.; Cui, L. Genome-Wide Identification and Characterization of the WRKY Gene Family in Scutellaria baicalensis Georgi under Diverse Abiotic Stress. Int. J. Mol. Sci. 2022, 23, 4225. [Google Scholar] [CrossRef]
- Kong, R.S.; Way, D.A.; Henry, H.A.L.; Smith, N.G. Stomatal conductance, not biochemistry, drives low temperature acclimation of photosynthesis in Populus balsamifera, regardless of nitrogen availability. Plant Biol. 2022, 24, 766–779. [Google Scholar] [CrossRef]
- Eom, S.H.; Ahn, M.-A.; Kim, E.; Lee, H.J.; Lee, J.H.; Wi, S.H.; Kim, S.K.; Bin Lim, H.; Hyun, T.K. Plant Response to Cold Stress: Cold Stress Changes Antioxidant Metabolism in Heading Type Kimchi Cabbage (Brassica rapa L. ssp. Pekinensis). Antioxidants 2022, 11, 700. [Google Scholar] [CrossRef]
- Lv, T.; Liu, Q.; Xiao, H.; Fan, T.; Zhou, Y.; Wang, J.; Tian, C.-E. Genome-wide identification and analysis of the IQM gene family in soybean. Front. Plant Sci. 2022, 13, 1093589. [Google Scholar] [CrossRef]
- Gutierrez, L.; Mauriat, M.; Guénin, S.; Pelloux, J.; Lefebvre, J.-F.; Louvet, R.; Rusterucci, C.; Moritz, T.; Guerineau, F.; Bellini, C.; et al. The lack of a systematic validation of reference genes: A serious pitfall undervalued in reverse transcription-polymerase chain reaction (RT-PCR) analysis in plants. Plant Biotechnol. J. 2008, 6, 609–618. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Id | pI | Mw/kD | AAs | |
|---|---|---|---|---|---|
| EXPA | PtEXPA1 | Potra2n1c11.1 | 8.99 | 30.19 | 278 |
| EXPA | PtEXPA2 | Potra2n1c960.1 | 9.49 | 29.15 | 269 |
| EXPA | PtEXPA3 | Potra2n1c2087.1 | 9.48 | 29.02 | 267 |
| EXPA | PtEXPA4 | Potra2n1c3505.1 | 9.69 | 25.33 | 231 |
| EXPA | PtEXPA5 | Potra2n2c4734.1 | 10.22 | 20.64 | 185 |
| EXPA | PtEXPA6 | Potra2n2c4737.1 | 8.98 | 27.76 | 256 |
| EXPA | PtEXPA7 | Potra2n2c6293.1 | 9.1 | 27.83 | 256 |
| EXPA | PtEXPA8 | Potra2n4c9142.1 | 8.88 | 30.02 | 275 |
| EXPA | PtEXPA9 | Potra2n4c9551.1 | 8.66 | 25.81 | 241 |
| EXPA | PtEXPA10 | Potra2n4c10239.1 | 8.77 | 67.81 | 596 |
| EXPA | PtEXPA11 | Potra2n5c10644.1 | 9.27 | 27.79 | 256 |
| EXPA | PtEXPA12 | Potra2n6c14375.1 | 9.14 | 27.62 | 255 |
| EXPA | PtEXPA13 | Potra2n6c14588.1 | 9.62 | 34.96 | 323 |
| EXPA | PtEXPA14 | Potra2n8c17125.1 | 9.5 | 2.79 | 258 |
| EXPA | PtEXPA15 | Potra2n8c17409.1 | 9.53 | 26.72 | 251 |
| EXPA | PtEXPA16 | Potra2n9c18642.1 | 8.09 | 27.80 | 250 |
| EXPA | PtEXPA17 | Potra2n9c19851.1 | 9.4 | 33.06 | 302 |
| EXPA | PtEXPA18 | Potra2n10c20623.1 | 9.37 | 30.40 | 279 |
| EXPA | PtEXPA19 | Potra2n10c20953.1 | 9.9 | 30.34 | 279 |
| EXPA | PtEXPA20 | Potra2n13c24992.1 | 7.52 | 26.70 | 250 |
| EXPA | PtEXPA21 | Potra2n13c25752.1 | 9.01 | 25.59 | 242 |
| EXPA | PtEXPA22 | Potra2n16c30169.1 | 9.34 | 35.69 | 323 |
| EXPA | PtEXPA23 | Potra2n16c30491.1 | 8.37 | 27.38 | 254 |
| EXPA | PtEXPA24 | Potra2n17c30722.1 | 8.89 | 29.94 | 274 |
| EXPA | PtEXPA25 | Potra2n17c31117.1 | 8.69 | 28.32 | 257 |
| EXPA | PtEXPA26 | Potra2n17c31179.1 | 9.44 | 28.55 | 264 |
| EXPA | PtEXPA27 | Potra2n19c33898.1 | 6.01 | 26.54 | 250 |
| EXPB | PtEXPB1 | Potra2n13c25121.1 | 7.61 | 28.71 | 262 |
| EXPB | PtEXPB2 | Potra2n14c26821.1 | 5.68 | 28.91 | 274 |
| EXPB | PtEXPB3 | Potra2n19c33436.1 | 8.51 | 28.60 | 262 |
| EXLA | PtEXLA1 | Potra2n4c10039.1 | 8.25 | 27.74 | 252 |
| EXLA | PtEXLA2 | Potra2n9c18880.1 | 8.96 | 29.88 | 274 |
| EXLB | PtEXLB1 | Potra2n1c1267.1 | 5.26 | 30.20 | 275 |
| EXLB | PtEXLB2 | Potra2n1c1318.1 | 6.57 | 31.54 | 290 |
| EXLB | PtEXLB3 | Potra2n3c7743.1 | 5.71 | 33.60 | 302 |
| EXLB | PtEXLB4 | Potra2n3c7783.1 | 6.71 | 27.61 | 250 |
| Gene 1 | Gene 2 | Ka | Ks | Ka/Ks | Duplication Type | T (MYA) 1 |
|---|---|---|---|---|---|---|
| PtEXLB2 | PtEXLB4 | 0.078312 | 0.340365 | 0.230082 | Segmental Duplication | 18.7 |
| PtEXPA19 | PtEXPA15 | 0.028716 | 0.369511 | 0.077713 | Segmental Duplication | 20.3 |
| PtEXLB1 | PtEXLB3 | 0.108542 | 0.365474 | 0.296989 | Segmental Duplication | 20.08 |
| PtEXPA7 | PtEXPA11 | 0.051896 | 0.325983 | 0.1592 | Segmental Duplication | 17.91 |
| PtEXLA1 | PtEXLA2 | 0.075097 | 0.431483 | 0.174043 | Segmental Duplication | 23.7 |
| PtEXPA3 | PtEXPA14 | 0.099992 | 1.26139 | 0.079272 | Segmental Duplication | 69.3 |
| PtEXPA14 | PtEXPA17 | 0.097609 | 1.549521 | 0.062993 | Segmental Duplication | 85.13 |
| PtEXPA3 | PtEXPA17 | 0.037618 | 0.367091 | 0.102475 | Segmental Duplication | 20.16 |
| PtEXPA23 | PtEXPA27 | 0.094301 | 2.399905 | 0.039294 | Segmental Duplication | 131.86 |
| PtEXPA23 | PtEXPA12 | 0.063563 | 0.48916 | 0.129944 | Segmental Duplication | 26.87 |
| PtEXPB1 | PtEXPB3 | 0.037749 | 0.220531 | 0.171174 | Segmental Duplication | 12.11 |
| PtEXPA3 | PtEXPA18 | 0.131003 | 1.817119 | 0.072094 | Segmental Duplication | 99.84 |
| PtEXPA18 | PtEXPA14 | 0.081269 | 0.406443 | 0.199952 | Segmental Duplication | 22.33 |
| PtEXPA18 | PtEXPA17 | 0.179824 | 1.564093 | 0.11497 | Segmental Duplication | 85.93 |
| PtEXPA19 | PtEXPA26 | 0.145877 | 1.0302 | 0.141601 | Segmental Duplication | 56.6 |
| PtEXPA26 | PtEXPA9 | 0.055452 | 0.448207 | 0.123719 | Segmental Duplication | 24.62 |
| PtEXPA26 | PtEXPA15 | 0.108133 | 1.471218 | 0.073499 | Segmental Duplication | 80.83 |
| PtEXPA3 | PtEXPA13 | 0.119706 | 1.685795 | 0.071008 | Segmental Duplication | 92.62 |
| PtEXPA13 | PtEXPA17 | 0.124216 | 1.591113 | 0.078069 | Segmental Duplication | 87.42 |
| PtEXPA22 | PtEXPA14 | 0.404278 | 2.402618 | 0.168266 | Segmental Duplication | 132.01 |
| Species | EXPA | EXPB | EXLA | EXLB | Total | Reference |
|---|---|---|---|---|---|---|
| Populus tremula L. | 27 (75%) | 3 (8.3%) | 2 (5.5%) | 4 (11.1%) | 36 | In this study |
| Arabidopsis | 25 (71.4%) | 6 (17.1%) | 3 (8.6%) | 1 (2.9%) | 35 | [44] |
| Oryza sativa | 34 (58.6%) | 19(32.8%) | 4 (6.9%) | 1 (1.7%) | 58 | [44] |
| Chinese jujube | 19 (63.3%) | 3 (10.0%) | 1 (3.3%) | 7 (23.3%) | 30 | [45] |
| Apple | 34 (82.9%) | 1 (2.4%) | 2 (4.9%) | 4 (9.8%) | 41 | [46] |
| maize | 36 (40.9%) | 48 (54.5%) | 4 (4.5%) | 0 (0%) | 88 | [47] |
| Soybean | 49 (65.3%) | 9 (12.0%) | 12(2.7%) | 25(20.0%) | 72 | [48] |
| Grapevine | 20 (69.0%) | 4 (13.8%) | 1 (3.4%) | 4 (13.8%) | 29 | [49] |
| Tomato | 25 (65.8%) | 8 (21.1%) | 1 (2.6%) | 4 (10.5%) | 38 | [50] |
| Tobacco | 36 (69.2%) | 6 (11.5%) | 3 (5.8%) | 7 (13.5%) | 52 | [51] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, Z.; Zhou, F.; Chen, Y.; Wu, H.; Yin, T. Genome-Wide Analysis of the Expansin Gene Family in Populus and Characterization of Expression Changes in Response to Phytohormone (Abscisic Acid) and Abiotic (Low-Temperature) Stresses. Int. J. Mol. Sci. 2023, 24, 7759. https://doi.org/10.3390/ijms24097759
Yin Z, Zhou F, Chen Y, Wu H, Yin T. Genome-Wide Analysis of the Expansin Gene Family in Populus and Characterization of Expression Changes in Response to Phytohormone (Abscisic Acid) and Abiotic (Low-Temperature) Stresses. International Journal of Molecular Sciences. 2023; 24(9):7759. https://doi.org/10.3390/ijms24097759
Chicago/Turabian StyleYin, Zhihui, Fangwei Zhou, Yingnan Chen, Huaitong Wu, and Tongming Yin. 2023. "Genome-Wide Analysis of the Expansin Gene Family in Populus and Characterization of Expression Changes in Response to Phytohormone (Abscisic Acid) and Abiotic (Low-Temperature) Stresses" International Journal of Molecular Sciences 24, no. 9: 7759. https://doi.org/10.3390/ijms24097759