
Interaction Network Construction and Functional Analysis of the Plasma Membrane H+-ATPase in Bangia fuscopurpurea (Rhodophyta)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
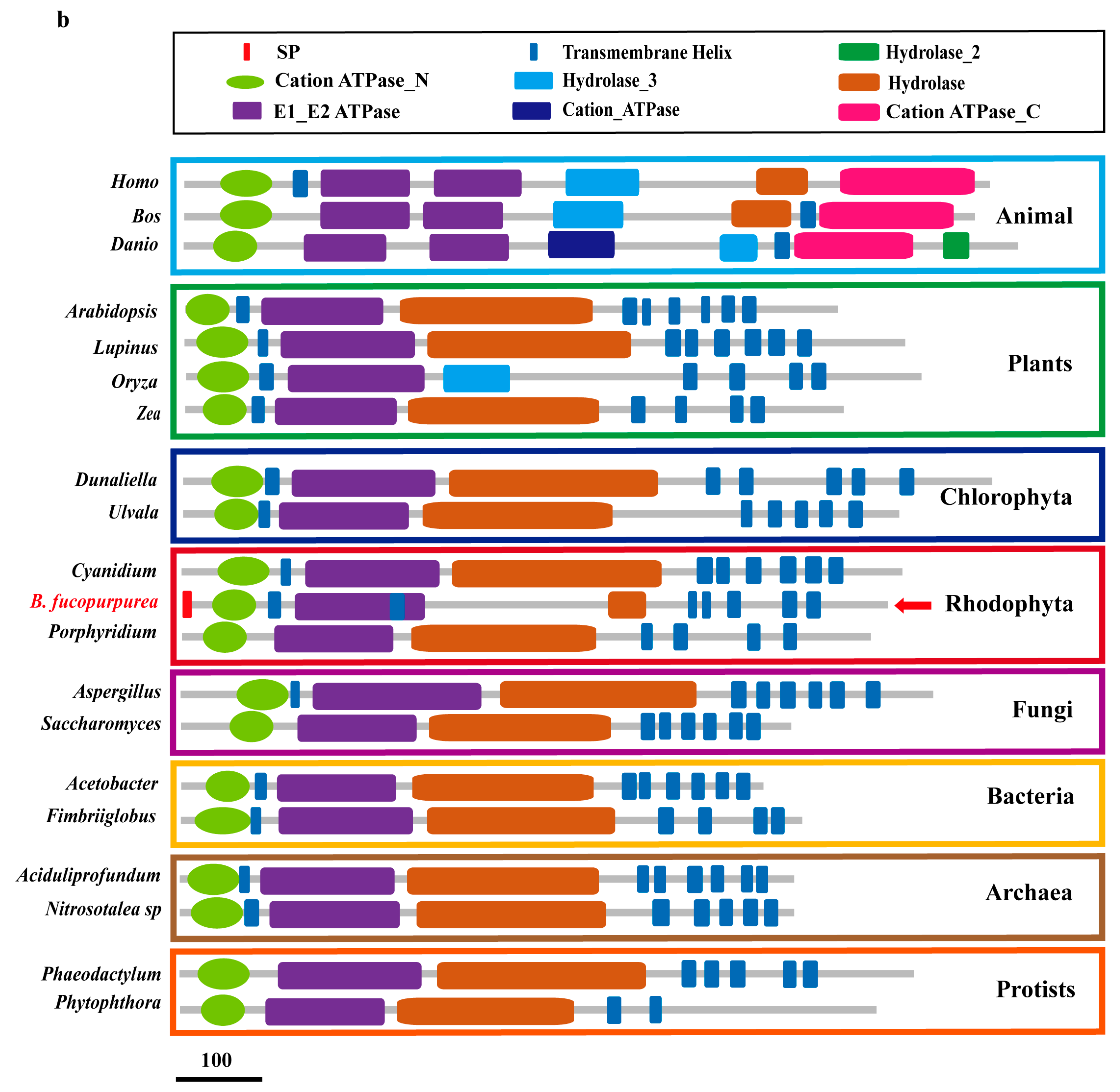
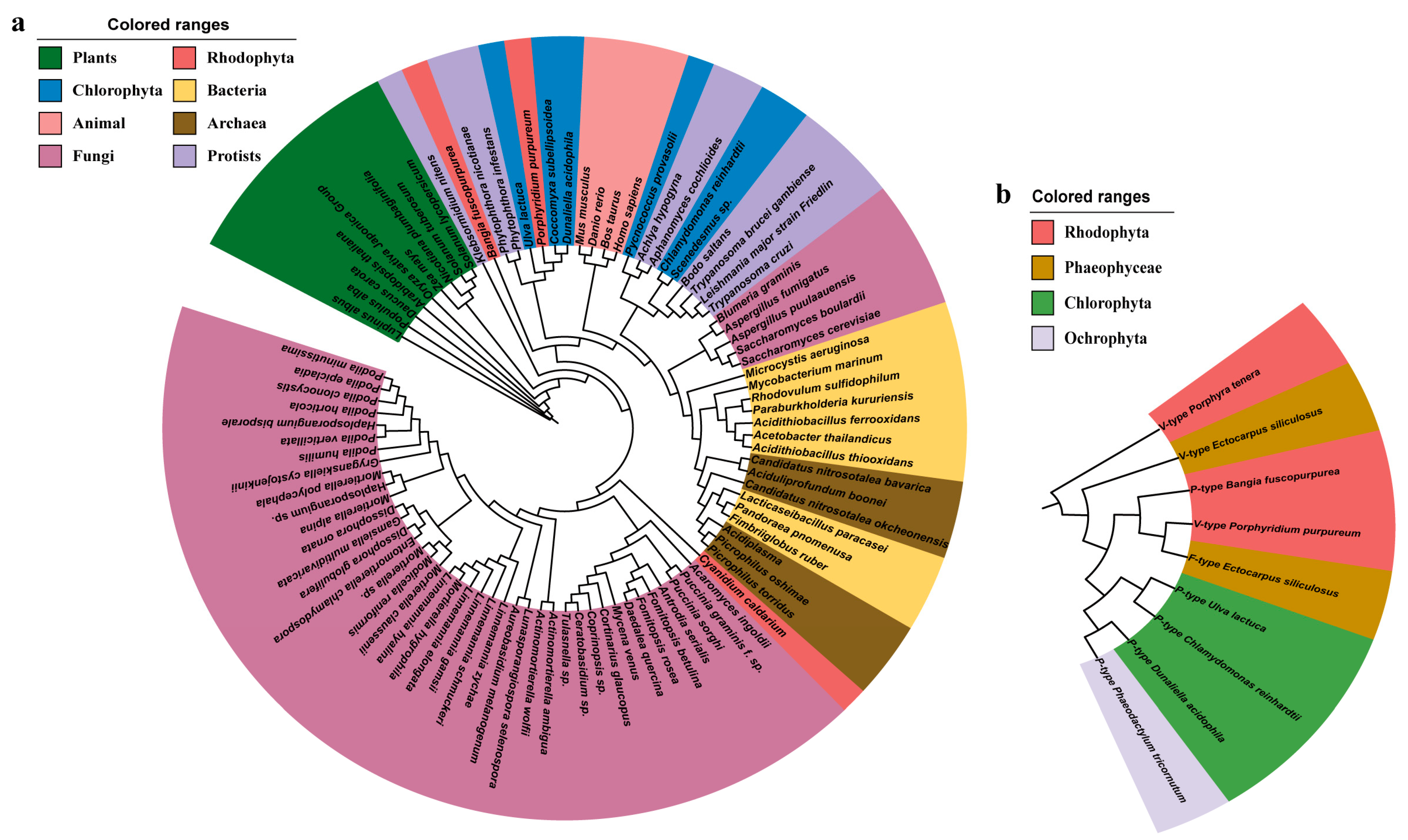
2.1. Features of BfPMHA
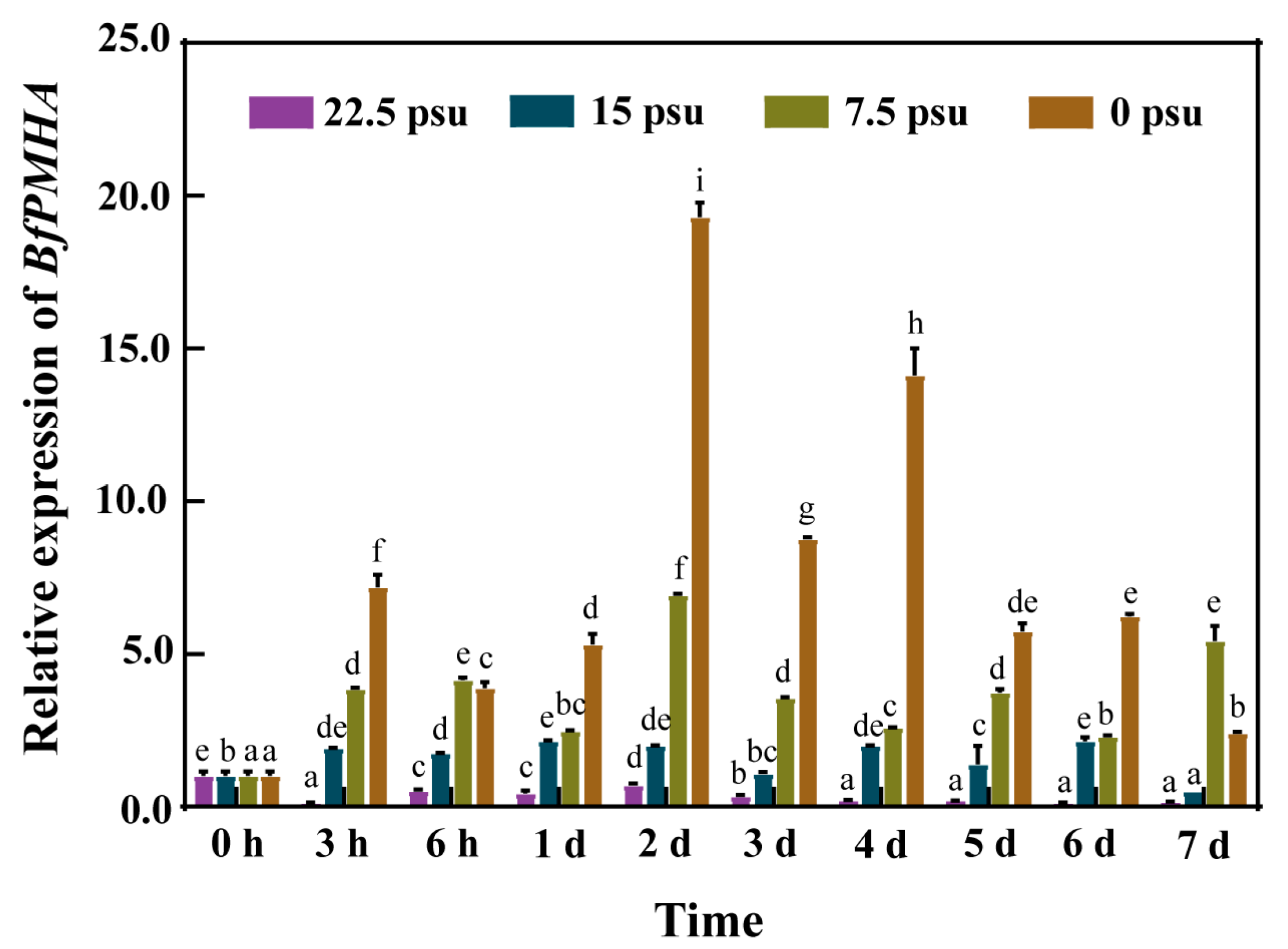
2.2. Expression Pattern of the BfPMHA Gene under Hypo-Salinity
2.3. Screening of Putative Interaction Proteins of BfPMHA
2.4. Characterization of BfMnSOD, BfFBA, and BfGAPDH
2.5. Analysis of Interaction between BfPMHA and the Three Interaction Proteins
2.6. Transgenic Yeast Tolerance to NaCl Stress
3. Discussion
4. Materials and Methods
4.1. Materials and Growth Conditions
4.2. Isolation and Sequence Analysis of BfPMHA
4.3. Prediction of BfPMHA Sequence Features and Protein Structure
4.4. RNA Isolation and qRT-PCR
4.5. Protein Interaction Analysis
4.6. NaCl Stress Tolerance Assay in Yeast
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Witzel, K.; Matros, A.; Bertsch, U.; Aftab, T.; Rutten, T.; Ramireddy, E.; Melzer, M.; Kunze, G.; Mock, H.P. The Jacalin-Related Lectin HvHorcH Is Involved in the Physiological Response of Barley Roots to Salt Stress. Int. J. Mol. Sci. 2021, 22, 10248. [Google Scholar] [CrossRef] [PubMed]
- Butcher, K.; Wick, A.F.; DeSutter, T.; Chatterjee, A.; Harmon, J. Soil salinity: A threat to global food security. Agron. J. 2016, 108, 2189–2200. [Google Scholar] [CrossRef]
- Rawoof, A.; Ramchiary, N.; Abdin, M.Z. A high-throughput RNA-Seq approach to elucidate the transcriptional response of Piriformospora indica to high salt stress. Sci. Rep. 2021, 11, 1–15. [Google Scholar]
- Karsten, U. Seaweed acclimation to salinity and desiccation stress. In Seaweed Biology; Springer: Berlin/Heidelberg, Germany, 2012; pp. 87–107. [Google Scholar]
- Wiencke, C. The response of Pyruvate Kinase From The Intertidal Red Alga Porphyra umbilicalis to Sodium and Potassium Ions. J. Plant Physiol. 1984, 116, 447–453. [Google Scholar] [CrossRef]
- Brawley, S.H.; Blouin, N.A.; Ficko-Blean, E.; Wheeler, G.L.; Lohr, M.; Goodson, H.V.; Jenkins, J.W.; Blaby-Haas, C.E.; Helliwell, K.E.; Chan, C.X. Insights into the red algae and eukaryotic evolution from the genome of Porphyra umbilicalis (Bangiophyceae, Rhodophyta). Proc. Natl. Acad. Sci. USA 2017, 114, E6361–E6370. [Google Scholar] [CrossRef]
- Broom, J.E.S.; Farr, T.J.; Nelson, W.A. Phylogeny of the Bangia flora of New Zealand suggests a southern origin for Porphyra and Bangia (Bangiales, Rhodophyta). Mol. Biol. Evol. 2004, 31, 1197–1207. [Google Scholar] [CrossRef]
- Geesink, R. Experimental investigations on marine and freshwater Bangia (Rhodophyta) from the Netherlands. J. Exp. Mar. Biol. Ecol. 1973, 11, 239–247. [Google Scholar] [CrossRef]
- Reed, R.H. On the conspecificity of marine and freshwater Bangia in Britain. Br. Phycol. J. 1980, 15, 411–416. [Google Scholar] [CrossRef]
- Wang, W.J.; Zhu, J.Y.; Xu, P.; Xu, J.R.; Lin, X.Z.; Huang, C.K.; Song, W.L.; Peng, G.; Wang, G.C. Characterization of the life history of Bangia fuscopurpurea (Bangiaceae, Rhodophyta) in connection with its cultivation in China. Aquaculture 2008, 278, 101–109. [Google Scholar] [CrossRef]
- Wang, W.J.; Li, X.L.; Zhu, J.Y.; Liang, Z.R.; Liu, F.L.; Sun, X.T.; Wang, F.J.; Shen, Z.G. Antioxidant response to salinity stress in freshwater and marine Bangia (Bangiales, Rhodophyta). Aquat. Bot. 2019, 154, 35–41. [Google Scholar] [CrossRef]
- Wei, J.; Li, X.L.; Ma, D.; Wang, W.; Liang, Z.; Li, G.; Liu, F.; Sun, X.; Wang, F. Effects of Salinity and Na3VO4 Stress on Photosynthetic Physiology of Bangia. Prog. Fish. Sci. 2020, 41, 141–149. [Google Scholar]
- Wang, W.; Shen, Z.; Sun, X.; Liu, F.; Liang, Z.; Wang, F.; Zhu, J. De novo transcriptomics analysis revealed a global reprogramming towards dehydration and hyposalinity in Bangia fuscopurpurea gametophytes (Rhodophyta). J. Appl. Phycol. 2018, 31, 637–651. [Google Scholar] [CrossRef]
- Ding, M.; Zhang, M.; Zeng, H.; Hayashi, Y.; Zhu, Y.; Kinoshita, T. Molecular basis of plasma membrane H+-ATPase function and potential application in the agricultural production. Plant Physiol. Biochem. 2021, 168, 10–16. [Google Scholar] [CrossRef]
- Palmgren, M.G. Plant plasma membrane H+-ATPases: Powerhouses for nutrient uptake. Annu. Rev. Plant Biol. 2001, 52, 817–845. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Shi, T.; Tang, X.; Tang, M.; Gong, J.; Yi, Y. Bacillus amyloliquefaciens PDR1 from root of karst adaptive plant enhances Arabidopsis thaliana resistance to alkaline stress through modulation of plasma membrane H+-ATPase activity. Plant Physiol. Biochem. 2020, 155, 472–482. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, Y.; Ma, L.; Yang, Z.; Dong, Q.; Li, Q.; Ni, X.; Kudla, J.; Song, C.; Guo, Y. The Ca2+ Sensor SCaBP3/CBL7 Modulates Plasma Membrane H+-ATPase Activity and Promotes Alkali Tolerance in Arabidopsis. Plant Cell 2019, 31, 1367–1384. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, Y.; Nakamura, S.; Takemiya, A.; Takahashi, Y.; Shimazaki, K.-i.; Kinoshita, T. Biochemical characterization of in vitro phosphorylation and dephosphorylation of the plasma membrane H+-ATPase. Plant Cell Physiol. 2010, 51, 1186–1196. [Google Scholar] [CrossRef]
- Cao, Y.; Zhang, M.; Liang, X.; Li, F.; Shi, Y.; Yang, X.; Jiang, C. Natural variation of an EF-hand Ca2+-binding-protein coding gene confers saline-alkaline tolerance in maize. Nat. Commun. 2020, 11, 186. [Google Scholar] [CrossRef]
- Manzoor, M.A.; Cheng, X.; Li, G.; Su, X.; Abdullah, M.; Cai, Y. Gene structure, evolution and expression analysis of the P-ATPase gene family in Chinese pear (Pyrus bretschneideri). Comput. Biol. Chem. 2020, 88, 107346. [Google Scholar] [CrossRef]
- Weiss, M.; Pick, U. Primary structure and effect of pH on the expression of the plasma membrane H+-ATPase from Dunaliella acidophila and Dunaliella salina. Plant Physiol. 1996, 112, 1693–1702. [Google Scholar] [CrossRef]
- Klenell, M.; Snoeijs, P.; Pedersén, M. The Involvement Of A Plasma Membrane H+-ATPase In The Blue-Light Enhancement Of Photosynthesis In Laminaria Digitata (Phaeophyta). J. Phycol. 2002, 38, 1143–1149. [Google Scholar] [CrossRef]
- Zhang, S.R.; Yang, Q. Effect of Low Temperature Stress on the Activities of Pm H+-ATPase in Spirulina (Arthrospira) Platensis. J. Inn. Mong. Med. Univ. 2015, 37, 1–5. [Google Scholar]
- Axelsen, K.; Venema, K.; Jahn, T.; Baunsgaard, L.; Palmgren, M. Molecular dissection of the C-terminal regulatory domain of the plant plasma membrane H+-ATPase AHA2: Mapping of residues that when altered give rise to an activated enzyme. Biochemistry 1999, 38, 7227–7234. [Google Scholar] [CrossRef] [PubMed]
- Morsomme, P.; Boutry, M. The plant plasma membrane H+-ATPase: Structure, function and regulation. Biochim. Biophys. Acta (BBA)-Biomembr. 2000, 1465, 1–16. [Google Scholar] [CrossRef]
- Tsirigos, K.D.; Govindarajan, S.; Bassot, C.; Västermark, Å.; Lamb, J.; Shu, N.; Elofsson, A. Topology of membrane proteins-predictions, limitations and variations. Curr. Opin. Struct. Biol. 2018, 50, 9–17. [Google Scholar] [CrossRef]
- Morsomme, P.; de Kerchove d’Exaerde, A.; De Meester, S.; Thines, D.; Goffeau, A.; Boutry, M. Single point mutations in various domains of a plant plasma membrane H+-ATPase expressed in Saccharomyces cerevisiae increase H+-pumping and permit yeast growth at low pH. EMBO J. 1996, 15, 5513–5526. [Google Scholar] [CrossRef]
- Falhof, J.; Pedersen, J.T.; Fuglsang, A.T.; Palmgren, M. Plasma membrane H+-ATPase regulation in the center of plant physiology. Mol. Plant 2016, 9, 323–337. [Google Scholar] [CrossRef]
- Ohta, H.; Shirakawa, H.; Uchida, K.; Yoshida, M.; Matuo, Y.; Enami, I. Cloning and sequencing of the gene encoding the plasma membrane H+-ATPase from an acidophilic red alga, Cyanidium caldarium. Biochim. Et Biophys. Acta (BBA)-Bioenerg. 1997, 1319, 9–13. [Google Scholar] [CrossRef]
- Chong, S.; Graham, T.G.; Dugast-Darzacq, C.; Dailey, G.M.; Darzacq, X.; Tjian, R. Tuning levels of low-complexity domain interactions to modulate endogenous oncogenic transcription. Mol. Cell 2022, 82, 2084–2097.e5. [Google Scholar] [CrossRef]
- Serrano, R.; Kielland-Brandt, M.C.; Fink, G.R. Yeast plasma membrane ATPase is essential for growth and has homology with (Na+ + K+), K+-and Ca2+-ATPases. Nature 1986, 319, 689–693. [Google Scholar] [CrossRef]
- Palmgren, M.; Sommarin, M.; Serrano, R.; Larsson, C. Identification of an autoinhibitory domain in the C-terminal region of the plant plasma membrane H+-ATPase. J. Biol. Chem. 1991, 266, 20470–20475. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Bin, J.-H.; Wang, X.-J. Effects of methyl jasmonate treatment on the hydrolytic activity and phosphorylation level of plasma membrane H+-ATPase in mung bean (Vigna radiata L.) hypocotyls. J. Plant Physiol. Mol. Biol. 2004, 30, 665–670. [Google Scholar]
- Ling, Q.; Xiang, Z.; Liu, H.; Shang, K. Regulation of PM H+-ATPase activity by its phosphorylation. J. Biochem. Mol. Biol. 1998, 14, 346–349. [Google Scholar]
- Chen, J.; Cui, Y.; Shu, W.; Zhao, Y.; Chen, X. Cloning of wheat PM H+-ATPase gene and analysis of its putative protein structure. J. Triticeae Crops 2009, 29, 947–953. [Google Scholar]
- Kluge, C.; Lahr, J.; Hanitzsch, M.; Bolte, S.; Golldack, D.; Dietz, K.-J. New insight into the structure and regulation of the plant vacuolar H+-ATPase. J. Bioenerg. Biomembr. 2003, 35, 377–388. [Google Scholar] [CrossRef]
- Li, Y.; Zeng, H.; Xu, F.; Yan, F.; Xu, W. H+-ATPases in Plant Growth and Stress Responses. Annu. Rev. Plant Biol. 2022, 73, 495–521. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chu, J.S.-C.; Chen, J.; Luo, Q.; Wang, H.; Lu, R.; Zhu, Z.; Yuan, G.; Yi, X.; Mao, Y. Insights into the ancient adaptation to intertidal environments by red algae based on a genomic and multiomics investigation of Neoporphyra haitanensis. Mol. Biol. Evol. 2022, 39, msab315. [Google Scholar] [CrossRef]
- Verweij, W.; Spelt, C.; Di Sansebastiano, G.-P.; Vermeer, J.; Reale, L.; Ferranti, F.; Koes, R.; Quattrocchio, F. An H+ P-ATPase on the tonoplast determines vacuolar pH and flower colour. Nat. Cell Biol. 2008, 10, 1456–1462. [Google Scholar] [CrossRef]
- Appelhagen, I.; Nordholt, N.; Seidel, T.; Spelt, K.; Koes, R.; Quattrochio, F.; Sagasser, M.; Weisshaar, B. TRANSPARENT TESTA 13 is a tonoplast P3A-ATP ase required for vacuolar deposition of proanthocyanidins in Arabidopsis thaliana seeds. Plant J. 2015, 82, 840–849. [Google Scholar] [CrossRef]
- Schumacher, K.; Krebs, M. The V-ATPase: Small cargo, large effects. Curr. Opin. Plant Biol. 2010, 13, 724–730. [Google Scholar] [CrossRef]
- Dettmer, J.; Liu, T.-Y.; Schumacher, K. Functional analysis of Arabidopsis V-ATPase subunit VHA-E isoforms. Eur. J. Cell Biol. 2010, 89, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Xu, J.; Chang, W.; Zhang, Z. Isolation and expression analysis of four members of the plasma membrane H+-ATPase gene family in Hevea brasiliensis. Trees 2015, 29, 1355–1363. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, Y.; Chen, X.; Xu, F.; Ding, M.; Ye, W.; Kawai, Y.; Toda, Y.; Hayashi, Y.; Suzuki, T. Plasma membrane H+-ATPase overexpression increases rice yield via simultaneous enhancement of nutrient uptake and photosynthesis. Nat. Commun. 2021, 12, 735. [Google Scholar] [CrossRef]
- Ravari, H.H.; Kavousi, H.R.; Mohammadi, F.; Pourseyedi, S. Partial cloning, characterization, and analysis of expression and activity of plasma membrane H+-ATPase in Kallar grass [Leptochloa fusca (L.) Kunth] under salt stress. Biol. Futura 2020, 71, 231–240. [Google Scholar] [CrossRef]
- Janicka, M.; Wdowikowska, A.; Klobus, G. Assay of Plasma Membrane H+-ATPase in Plant Tissues under Abiotic Stresses. Methods Mol. Biol. 2018, 1696, 205–215. [Google Scholar] [PubMed]
- Haruta, M.; Gray, W.M.; Sussman, M.R. Regulation of the plasma membrane proton pump H+-ATPase by phosphorylation. Curr. Opin. Plant Biol. 2015, 28, 68–75. [Google Scholar] [CrossRef]
- Uji, T.; Hirata, R.; Mikami, K.; Mizuta, H.; Saga, N. Molecular characterization and expression analysis of sodium pump genes in the marine red alga Porphyra yezoensis. Mol. Biol. Rep. 2012, 39, 7973–7980. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Zhou, X.; Yang, Q.; Liu, H.; Kurtenbach, R. Changes in H+-ATPase activity and conjugated polyamine contents in plasma membrane purified from developing wheat embryos under short-time drought stress. Plant Growth Regul. 2015, 75, 1–10. [Google Scholar] [CrossRef]
- Kalampanayil, B.D.; Wimmers, L.E. Identification and characterization of a salt-stress-induced plasma membrane H+-ATPase in tomato. Plant Cell Environ. 2001, 24, 999–1000. [Google Scholar] [CrossRef]
- Vitart, V.; Baxter, I.; Doerner, P.; Harper, J.F. Evidence for a role in growth and salt resistance of a plasma membrane H+-ATPase in the root endodermis. Plant J. 2001, 27, 191–201. [Google Scholar] [CrossRef]
- Elmore, J.M.; Coaker, G. The Role of the Plasma Membrane H+-ATPase in Plant–Microbe Interactions. Mol. Plant 2011, 4, 416–427. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, X.; Guo, W.; Liu, W.; Shao, W.; Zhao, J.; Li, J.; Dong, Q.; Ma, L.; He, Q.; et al. Testing the polar auxin transport model with a selective plasma membrane H+-ATPase inhibitor. J. Integr. Plant Biol. 2022, 64, 1229–1245. [Google Scholar] [CrossRef] [PubMed]
- Wei, J. Physiological Mechanism Research of Bangia fuscopurpurea and Bangia atropurpurea in Response to Salinity Stress. Master’s Thesis, Shanghai Ocean University, Shanghai, China, 2020. [Google Scholar]
- Bose, J.; Xie, Y.; Shen, W.; Shabala, S. Haem oxygenase modifies salinity tolerance in Arabidopsis by controlling K+ retention via regulation of the plasma membrane H+-ATPase and by altering SOS1 transcript levels in roots. J. Exp. Bot. 2013, 64, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Shen, Z.; Zhang, Y.; Wu, X.; Wang, J.; Sa, G.; Zhang, Y.; Zhang, H.; Deng, C.; Liu, J.; et al. Populus euphratica WRKY1 binds the promoter of H+-ATPase gene to enhance gene expression and salt tolerance. J. Exp. Bot. 2020, 71, 1527–1539. [Google Scholar] [CrossRef]
- Olfatmiri, H.; Alemzadeh, A.; Zakipour, Z. Up-regulation of plasma membrane H+-ATPase under salt stress may enable Aeluropus littoralis to cope with stress. Mol. Biol. Res. Commun. 2014, 3, 67. [Google Scholar]
- Miller, A.-F.; Padmakumar, K.; Sorkin, D.L.; Karapetian, A.; Vance, C.K. Proton-coupled electron transfer in Fe-superoxide dismutase and Mn-superoxide dismutase. J. Inorg. Biochem. 2003, 93, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, D.; Xiang, L.; Wang, Q. MnSOD functions as a thermoreceptor activated by low temperature. J. Inorg. Biochem. 2022, 229, 111745. [Google Scholar] [CrossRef] [PubMed]
- McAdam, M.E.; Fox, R.A.; Lavelle, F.; Fielden, E.M. A pulse-radiolysis study of the manganese-containing superoxide dismutase from Bacillus stearothermophilus. A kinetic model for the enzyme action. Biochem. J. 1977, 165, 71–79. [Google Scholar] [CrossRef]
- McAdam, M.E.; Levelle, F.; Fox, R.A.; Fielden, E.M. A pulse-radiolysis study of the manganese-containing superoxide dismutase from Bacillus stearothermophilus. Biochem. J. 1977, 165, 81–87. [Google Scholar] [CrossRef]
- Nounjan, N.; Nghia, P.T.; Theerakulpisut, P. Exogenous proline and trehalose promote recovery of rice seedlings from salt-stress and differentially modulate antioxidant enzymes and expression of related genes. J. Plant Physiol. 2012, 169, 596–604. [Google Scholar] [CrossRef]
- Kaminaka, H.; Morita, S.; Tokumoto, M.; Masumura, T.; Tanaka, K. Differential gene expressions of rice superoxide dismutase isoforms to oxidative and environmental stresses. Free Radic. Res. 1999, 31, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Sairam, R.K.; Srivastava, G.C.; Agarwal, S.; Meena, R.C. Differences in antioxidant activity in response to salinity stress in tolerant and susceptible wheat genotypes. Biol. Plant. 2005, 49, 85–91. [Google Scholar] [CrossRef]
- Tristan, C.; Shahani, N.; Sedlak, T.W.; Sawa, A. The diverse functions of GAPDH: Views from different subcellular compartments. Cell. Signal. 2011, 23, 317–323. [Google Scholar] [CrossRef]
- Sertsuvalkul, N.; DeMell, A.; Dinesh-Kumar, S.P. The complex roles of autophagy in plant immunity. FEBS Lett. 2022, 596, 2163–2171. [Google Scholar] [CrossRef]
- Sirover, M.A. New insights into an old protein: The functional diversity of mammalian glyceraldehyde-3-phosphate dehydrogenase. Biochim. Biophys. Acta 1999, 1432, 159–184. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Wang, Y.; Zheng, X.; Jia, Q.; Zhao, J.; Bai, F.; Hong, Y.; Liu, Y. Cytoplastic glyceraldehyde-3-phosphate dehydrogenases interact with ATG3 to negatively regulate autophagy and immunity in Nicotiana benthamiana. Plant Cell 2015, 27, 1316–1331. [Google Scholar] [CrossRef]
- Henry, E.; Fung, N.; Liu, J.; Drakakaki, G.; Coaker, G. Beyond glycolysis: GAPDHs are multi-functional enzymes involved in regulation of ROS, autophagy, and plant immune responses. PLoS Genet. 2015, 11, e1005199. [Google Scholar] [CrossRef] [PubMed]
- Wawer, I.; Bucholc, M.; Astier, J.; Anielska-Mazur, A.; Dahan, J.; Kulik, A.; Wyslouch-Cieszynska, A.; Zareba-Koziol, M.; Krzywinska, E.; Dadlez, M.; et al. Regulation of Nicotiana tabacum osmotic stress-activated protein kinase and its cellular partner GAPDH by nitric oxide in response to salinity. Biochem. J. 2010, 429, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, H.; Amano, W.; Kubo, T.; Fukuhara, A.; Ihara, H.; Azuma, Y.-T.; Tajima, H.; Inui, T.; Sawa, A.; Takeuchi, T. Glyceraldehyde-3-phosphate dehydrogenase aggregate formation participates in oxidative stress-induced cell death. J. Biol. Chem. 2009, 284, 34331–34341. [Google Scholar] [CrossRef]
- Cai, B.; Ning, Y.; Li, Q.; Li, Q.; Ai, X. Effects of the Chloroplast Fructose-1,6-Bisphosphate Aldolase Gene on Growth and Low-Temperature Tolerance of Tomato. Int. J. Mol. Sci. 2022, 23, 728. [Google Scholar] [CrossRef]
- Lu, W.; Tang, X.; Huo, Y.; Xu, R.; Qi, S.; Huang, J.; Zheng, C.; Wu, C.A. Identification and characterization of fructose 1,6-bisphosphate aldolase genes in Arabidopsis reveal a gene family with diverse responses to abiotic stresses. Gene 2012, 503, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Komori, T.; Hashimoto, A.; Kuwata, S.; Kubo, T. Differential expression of plastidic aldolase genes in Nicotiana plants under salt stress. Plant Sci. 2000, 154, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Li, Q.; Liu, F.; Bi, H.; Ai, X. Decreasing fructose-1, 6-bisphosphate aldolase activity reduces plant growth and tolerance to chilling stress in tomato seedlings. Physiol. Plant. 2018, 163, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Zhang, Z.; Zhang, Y. Cloning and molecular characterization of fructose-1,6-bisphosphate aldolase gene regulated by high-salinity and drought in Sesuvium portulacastrum. Plant Cell Rep. 2009, 28, 975–984. [Google Scholar] [CrossRef]
- Xue, G.-P.; McIntyre, C.L.; Glassop, D.; Shorter, R. Use of expression analysis to dissect alterations in carbohydrate metabolism in wheat leaves during drought stress. Plant Mol. Biol. 2008, 67, 197–214. [Google Scholar] [CrossRef] [PubMed]
- Shu, S.; Chen, L.; Lu, W.; Sun, J.; Guo, S.; Yuan, Y.; Li, J. Effects of exogenous spermidine on photosynthetic capacity and expression of Calvin cycle genes in salt-stressed cucumber seedlings. J. Plant Res. 2014, 127, 763–773. [Google Scholar] [CrossRef]
- Ozturk, Z.N.; Talamé, V.; Deyholos, M.; Michalowski, C.B.; Galbraith, D.W.; Gozukirmizi, N.; Tuberosa, R.; Bohnert, H.J. Monitoring large-scale changes in transcript abundance in drought-and salt-stressed barley. Plant Mol. Biol. 2002, 48, 551–573. [Google Scholar] [CrossRef] [PubMed]
- Kamal, A.H.; Cho, K.; Kim, D.E.; Uozumi, N.; Chung, K.Y.; Lee, S.Y.; Choi, J.S.; Cho, S.W.; Shin, C.S.; Woo, S.H. Changes in physiology and protein abundance in salt-stressed wheat chloroplasts. Mol. Biol. Rep. 2012, 39, 9059–9074. [Google Scholar] [CrossRef]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]
- Mu, J.; Fu, Y.; Liu, B.; Zhang, Y.; Wang, A.; Li, Y.; Zhu, J. SiFBA5, a cold-responsive factor from Saussurea involucrata promotes cold resilience and biomass increase in transgenic tomato plants under cold stress. BMC Plant Biol. 2021, 21, 75. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Sa, G.; Zhang, Y.; Deng, J.; Deng, S.; Wang, M.; Zhang, H.; Yao, J.; Ma, X. Populus euphratica J3 mediates root K+/Na+ homeostasis by activating plasma membrane H+-ATPase in transgenic Arabidopsis under NaCl salinity. Plant Cell Tissue Organ Cult. (PCTOC) 2017, 131, 75–88. [Google Scholar] [CrossRef]
- Yang, Y.; Qin, Y.; Xie, C.; Zhao, F.; Zhao, J.; Liu, D.; Chen, S.; Fuglsang, A.T.; Palmgren, M.G.; Schumaker, K.S. The Arabidopsis chaperone J3 regulates the plasma membrane H+-ATPase through interaction with the PKS5 kinase. Plant Cell 2010, 22, 1313–1332. [Google Scholar] [CrossRef]
- Fuglsang, A.T.; Borch, J.; Bych, K.; Jahn, T.P.; Roepstorff, P.; Palmgren, M.G. The binding site for regulatory 14-3-3 protein in plant plasma membrane H+-ATPase: Involvement of a region promoting phosphorylation-independent interaction in addition to the phosphorylation-dependent C-terminal end. J. Biol. Chem. 2003, 278, 42266–42272. [Google Scholar] [CrossRef]
- Jones, S.; Thornton, J.M. Principles of protein-protein interactions. Proc. Natl. Acad. Sci. USA 1996, 93, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Rao, V.S.; Srinivas, K.; Sujini, G.N.; Kumar, G.N. Protein-protein interaction detection: Methods and analysis. Int. J. Proteom. 2014, 2014, 147648. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Tang, N.; Zhang, H.; Li, X.; Xiao, J.; Xiong, L. Constitutive activation of transcription factor OsbZIP46 improves drought tolerance in rice. Plant Physiol. 2012, 158, 1755–1768. [Google Scholar] [CrossRef]
- Xie, K.; Minkenberg, B.; Yang, Y. Boosting CRISPR/Cas9 multiplex editing capability with the endogenous tRNA-processing system. Proc. Natl. Acad. Sci. USA 2015, 112, 3570–3575. [Google Scholar] [CrossRef]
- Hu, B.; Zhou, Y.; Zhou, Z.; Sun, B.; Zhou, F.; Yin, C.; Ma, W.; Chen, H.; Lin, Y. Repressed OsMESL expression triggers reactive oxygen species-mediated broad-spectrum disease resistance in rice. Plant Biotechnol. J. 2021, 19, 1511–1522. [Google Scholar] [CrossRef]
- Wang, P.; Guo, Y.; Wang, Y.; Gao, C. Vacuolar membrane H+-ATPase c” subunit gene (ThVHAc” 1) from Tamarix hispida Willd improves salt stress tolerance. Plant Physiol. Biochem. 2020, 157, 370–378. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, H.; Wang, W.; Cao, Y.; Liang, Z.; Zhang, P. Interaction Network Construction and Functional Analysis of the Plasma Membrane H+-ATPase in Bangia fuscopurpurea (Rhodophyta). Int. J. Mol. Sci. 2023, 24, 7644. https://doi.org/10.3390/ijms24087644
Yao H, Wang W, Cao Y, Liang Z, Zhang P. Interaction Network Construction and Functional Analysis of the Plasma Membrane H+-ATPase in Bangia fuscopurpurea (Rhodophyta). International Journal of Molecular Sciences. 2023; 24(8):7644. https://doi.org/10.3390/ijms24087644
Chicago/Turabian StyleYao, Haiqin, Wenjun Wang, Yuan Cao, Zhourui Liang, and Pengyan Zhang. 2023. "Interaction Network Construction and Functional Analysis of the Plasma Membrane H+-ATPase in Bangia fuscopurpurea (Rhodophyta)" International Journal of Molecular Sciences 24, no. 8: 7644. https://doi.org/10.3390/ijms24087644