
Endocannabinoid System Components of the Female Mouse Reproductive Tract Are Modulated during Reproductive Aging


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. ECS Component Quantification in Female Reproductive Tract during Aging
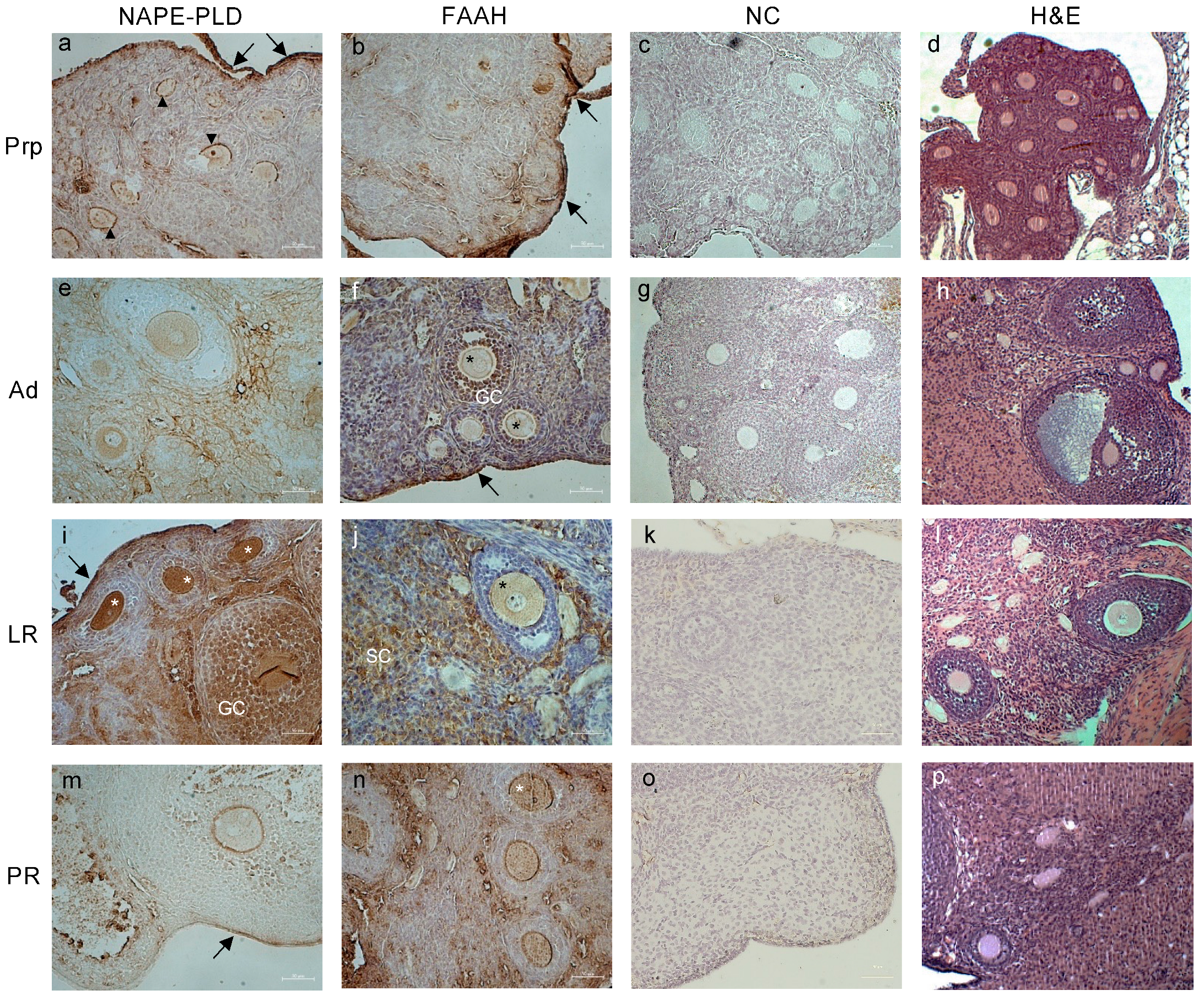
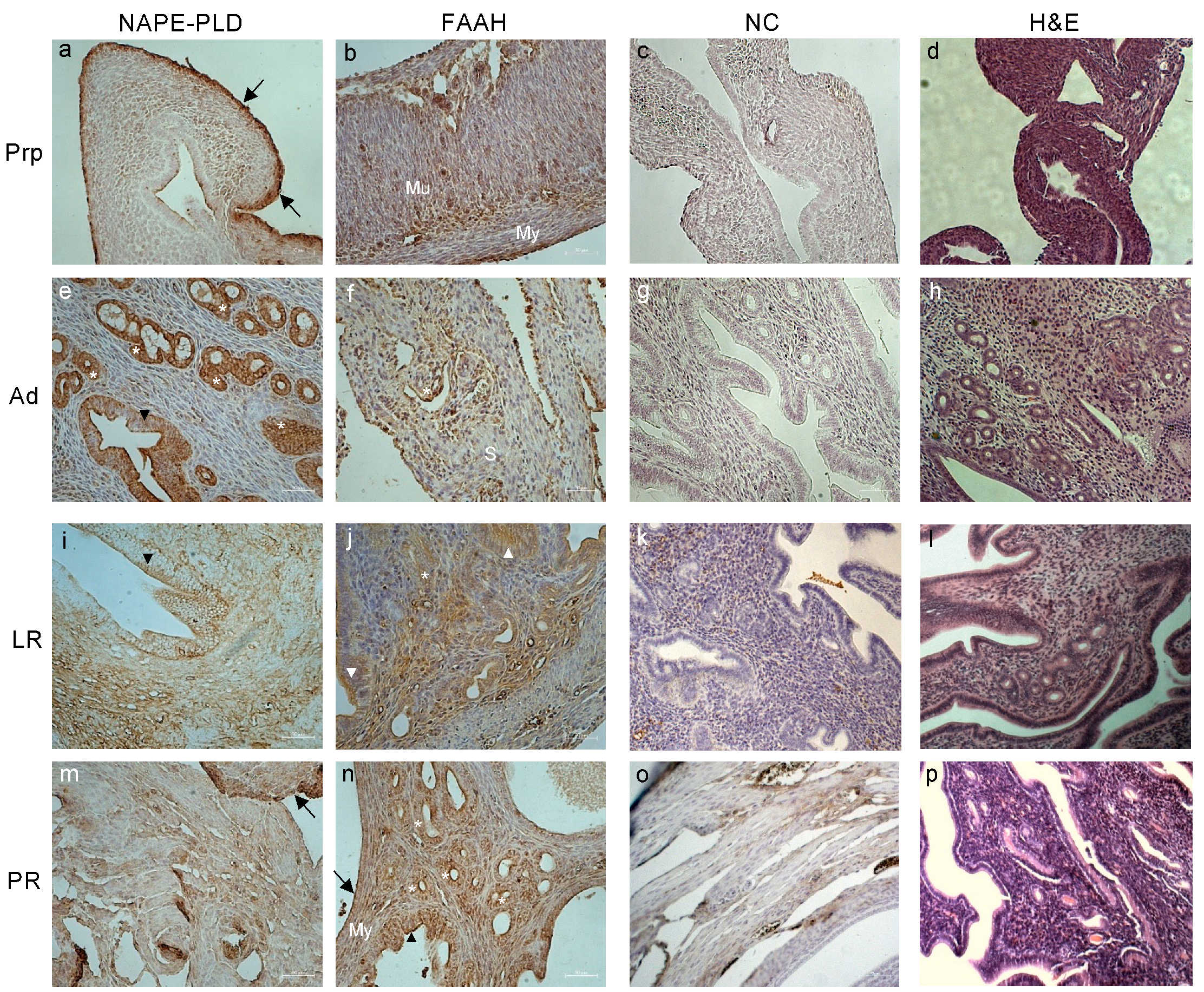
2.2. Tissue Localization of ECS Components in Female Reproductive Tract during Aging
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animals and Sample Collection
4.3. ELISA Assay
4.4. Hematoxylin–Eosin (H&E) and Immunohistochemistry (IHC)
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- ESHRE Capri Workshop Group Fertility and Ageing. Hum. Reprod. Update 2005, 11, 261–276. [CrossRef]
- Bendarska-Czerwińska, A.; Zmarzły, N.; Morawiec, E.; Panfil, A.; Bryś, K.; Czarniecka, J.; Ostenda, A.; Dziobek, K.; Sagan, D.; Boroń, D.; et al. Endocrine Disorders and Fertility and Pregnancy: An Update. Front. Endocrinol. 2023, 13, 970439. [Google Scholar] [CrossRef] [PubMed]
- Cecconi, S.; Rossi, G.; Castellucci, A.; D’Andrea, G.; Maccarrone, M. Endocannabinoid Signaling in Mammalian Ovary in the Memory of Riccardo Del Gratta (1966-2013). Eur. J. Obstet. Gynecol. Reprod. Biol. 2014, 178, 6–11. [Google Scholar] [CrossRef]
- Cecconi, S.; Rapino, C.; Di Nisio, V.; Rossi, G.; Maccarrone, M. The (Endo)Cannabinoid Signaling in Female Reproduction: What Are the Latest Advances? Prog. Lipid Res. 2020, 77, 101019. [Google Scholar] [CrossRef] [PubMed]
- El-Talatini, M.R.; Taylor, A.H.; Elson, J.C.; Brown, L.; Davidson, A.C.; Konje, J.C. Localisation and Function of the Endocannabinoid System in the Human Ovary. PLoS ONE 2009, 4, e4579. [Google Scholar] [CrossRef] [PubMed]
- El-Talatini, M.R.; Taylor, A.H.; Konje, J.C. Fluctuation in Anandamide Levels from Ovulation to Early Pregnancy in In-Vitro Fertilization-Embryo Transfer Women, and Its Hormonal Regulation. Hum. Reprod. 2009, 24, 1989–1998. [Google Scholar] [CrossRef]
- Cecconi, S.; Rossi, G.; Oddi, S.; Di Nisio, V.; Maccarrone, M. Role of Major Endocannabinoid-Binding Receptors during Mouse Oocyte Maturation. Int. J. Mol. Sci. 2019, 20, 2866. [Google Scholar] [CrossRef]
- Bagavandoss, P.; Grimshaw, S. Temporal and Spatial Distribution of the Cannabinoid Receptors (CB 1, CB2) and Fatty Acid Amide Hydroxylase in the Rat Ovary. Anat. Rec. 2010, 293, 1425–1432. [Google Scholar] [CrossRef]
- Pirone, A.; Lenzi, C.; Briganti, A.; Abbate, F.; Levanti, M.; Abramo, F.; Miragliotta, V. Spatial Distribution of Cannabinoid Receptor 1 and Fatty Acid Amide Hydrolase in the Cat Ovary and Oviduct. Acta Histochem. 2017, 119, 417–422. [Google Scholar] [CrossRef]
- De Domenico, E.; Todaro, F.; Rossi, G.; Dolci, S.; Geremia, R.; Rossi, P.; Grimaldi, P. Overactive Type 2 Cannabinoid Receptor Induces Meiosis in Fetal Gonads and Impairs Ovarian Reserve. Cell Death Dis. 2017, 8, e3085. [Google Scholar] [CrossRef]
- McKinney, M.K.; Cravatt, B.E. Structure and Function of Fatty Acid Amide Hydrolase. Annu. Rev. Biochem. 2005, 74, 411–432. [Google Scholar] [CrossRef] [PubMed]
- Schuel, H. Tuning the Oviduct to the Anandamide Tone. J. Clin. Investig. 2006, 116, 2087–2090. [Google Scholar] [CrossRef] [PubMed]
- Maccarrone, M.; Valensise, H.; Bari, M.; Lazzarin, N.; Romanini, C.; Finazzi-Agrò, A. Relation between Decreased Anandamide Hydrolase Concentrations in Human Lymphocytes and Miscarriage. Lancet 2000, 355, 1326–1329. [Google Scholar] [CrossRef] [PubMed]
- Gervasi, M.G.; Osycka-Salut, C.; Sanchez, T.; Alonso, C.A.I.; Llados, C.; Castellano, L.; Franchi, A.M.; Villalõn, M.; Perez-Martinez, S. Sperm Release from the Oviductal Epithelium Depends on Ca2+ Influx Upon Activation of CB1 and TRPV1 by Anandamide. J. Cell Biochem. 2016, 117, 320–333. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Wang, H.; Okabe, M.; Mackie, K.; Kingsley, P.J.; Marnett, L.J.; Cravatt, B.F.; Dey, S.K. Genetic Loss of Faah Compromises Male Fertility in Mice. Biol. Reprod. 2009, 80, 235–242. [Google Scholar] [CrossRef]
- Fonseca, B.M.; Correia-da-Silva, G.; Taylor, A.H.; Lam, P.M.W.; Marczylo, T.H.; Bell, S.C.; Konje, J.C.; Teixeira, N.A. The Endocannabinoid 2-Arachidonoylglycerol (2-AG) and Metabolizing Enzymes during Rat Fetoplacental Development: A Role in Uterine Remodelling. Int. J. Biochem. Cell Biol. 2010, 42, 1884–1892. [Google Scholar] [CrossRef]
- Fonseca, B.M.; Correia-da-Silva, G.; Taylor, A.H.; Lam, P.M.W.; Marczylo, T.H.; Konje, J.C.; Bell, S.C.; Teixeira, N.A. N-Acylethanolamine Levels and Expression of Their Metabolizing Enzymes during Pregnancy. Endocrinology 2010, 151, 3965–3974. [Google Scholar] [CrossRef]
- Maia, J.; Fonseca, B.M.; Teixeira, N.; Correia-Da-Silva, G. The Fundamental Role of the Endocannabinoid System in Endometrium and Placenta: Implications in Pathophysiological Aspects of Uterine and Pregnancy Disorders. Hum. Reprod. Update 2020, 26, 586–602. [Google Scholar] [CrossRef]
- Ashton, J.C.; Dowie, M.J.; Glass, M. The Endocannabinoid System and Human Brain Functions: Insight From Memory, Motor, and Mood Pathologies. In The Endocannabinoid System: Genetics, Biochemistry, Brain Disorders, and Therapy; Elsevier Inc.: Amsterdam, The Netherlands, 2017; pp. 115–186. ISBN 9780128096673. [Google Scholar]
- Kendall, D.A.; Yudowski, G.A. Cannabinoid Receptors in the Central Nervous System: Their Signaling and Roles in Disease. Front. Cell. Neurosci. 2017, 10, 294. [Google Scholar] [CrossRef]
- Rahaman, O.; Ganguly, D. Endocannabinoids in Immune Regulation and Immunopathologies. Immunology 2021, 164, 242–252. [Google Scholar] [CrossRef]
- Brents, L.K. Marijuana, the Endocannabinoid System and the Female Reproductive System. Yale J. Biol. Med. 2016, 89, 175–191. [Google Scholar] [PubMed]
- López-Cardona, A.P.; Pérez-Cerezales, S.; Fernández-González, R.; Laguna-Barraza, R.; Pericuesta, E.; Agirregoitia, N.; Gutiérrez-Adán, A.; Agirregoitia, E. CB1 Cannabinoid Receptor Drives Oocyte Maturation and Embryo Development via PI3K/Akt and MAPK Pathways. FASEB J. 2017, 31, 3372–3382. [Google Scholar] [CrossRef] [PubMed]
- Totorikaguena, L.; Olabarrieta, E.; Lolicato, F.; Romero-Aguirregomezcorta, J.; Smitz, J.; Agirregoitia, N.; Agirregoitia, E. The Endocannabinoid System Modulates the Ovarian Physiology and Its Activation Can Improve in Vitro Oocyte Maturation. J. Cell. Physiol. 2020, 235, 7580–7591. [Google Scholar] [CrossRef]
- Francou, M.M.; Girela, J.L.; de Juan, A.; Ten, J.; Bernabeu, R.; de Juan, J. Human Sperm Motility, Capacitation and Acrosome Reaction Are Impaired by 2-Arachidonoylglycerol Endocannabinoid. Histol. Histopathol. 2017, 32, 1351–1358. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xie, H.; Guo, Y.; Zhang, H.; Takahashi, T.; Kingsley, P.J.; Marnett, L.J.; Das, S.K.; Cravatt, B.F.; Dey, S.K. Fatty Acid Amide Hydrolase Deficiency Limits Early Pregnancy Events. J. Clin. Investig. 2006, 116, 2122–2131. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, B.M.; Correia-da-Silva, G.; Taylor, A.H.; Konje, J.C.; Bell, S.C.; Teixeira, N.A. Spatio-Temporal Expression Patterns of Anandamide-Binding Receptors in Rat Implantation Sites: Evidence for a Role of the Endocannabinoid System during the Period of Placental Development. Reprod. Biol. Endocrinol. 2009, 7, 121. [Google Scholar] [CrossRef] [PubMed]
- Scotchie, J.G.; Savaris, R.F.; Martin, C.E.; Young, S.L. Endocannabinoid Regulation in Human Endometrium across the Menstrual Cycle. Reprod. Sci. 2015, 22, 113–123. [Google Scholar] [CrossRef]
- Gebeh, A.K.; Willets, J.M.; Marczylo, E.L.; Taylor, A.H.; Konje, J.C. Ectopic Pregnancy Is Associated with High Anandamide Levels and Aberrant Expression of FAAH and CB1 in Fallopian Tubes. J. Clin. Endocrinol. Metab. 2012, 97, 2827–2835. [Google Scholar] [CrossRef]
- Rava, A.; Trezza, V. Emerging Roles of Endocannabinoids as Key Lipid Mediators for a Successful Pregnancy. Int. J. Mol. Sci. 2023, 24, 5220. [Google Scholar] [CrossRef]
- Gervasi, M.G.; Marczylo, T.H.; Lam, P.M.; Rana, S.; Franchi, A.M.; Konje, J.C.; Perez-Martinez, S. Anandamide Levels Fluctuate in the Bovine Oviduct during the Oestrous Cycle. PLoS ONE 2013, 8, e72521. [Google Scholar] [CrossRef]
- Wang, H.; Xie, H.; Sun, X.; Kingsley, P.J.; Marnett, L.J.; Cravatt, B.F.; Dey, S.K. Differential Regulation of Endocannabinoid Synthesis and Degradation in the Uterus during Embryo Implantation. Prostaglandins Other Lipid Mediat. 2007, 83, 62–74. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, B.M.; Cunha, S.C.; Gonçalves, D.; Mendes, A.; Braga, J.; Correia-Da-Silva, G.; Teixeira, N.A. Decidual NK Cell-Derived Conditioned Medium from Miscarriages Affects Endometrial Stromal Cell Decidualisation: Endocannabinoid Anandamide and Tumour Necrosis Factor-α Crosstalk. Hum. Reprod. 2020, 35, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xie, H.; Dey, S.K. Loss of Cannabinoid Receptor CB1 Induces Preterm Birth. PLoS ONE 2008, 3, e3320. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.H.; Tortolani, D.; Ayakannu, T.; Konje, J.C.; Maccarrone, M. (Endo)Cannabinoids and Gynaecological Cancers. Cancers 2021, 13, 37. [Google Scholar] [CrossRef]
- Oto, T.; Urata, K.; Hayashi, Y.; Hitomi, S.; Shibuta, I.; Iwata, K.; Iinuma, T.; Shinoda, M. Age-Related Differences in Transient Receptor Potential Vanilloid 1 and 2 Expression Patterns in the Trigeminal Ganglion Neurons Contribute to Changes in the Palatal Mucosal Heat Pain Sensitivity. Tohoku J. Exp. Med. 2022, 256, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Wanner, S.P.; Garami, A.; Pakai, E.; Oliveira, D.L.; Gavva, N.R.; Coimbra, C.C.; Romanovsky, A.A. Aging Reverses the Role of the Transient Receptor Potential Vanilloid-1 Channel in Systemic Inflammation from Anti-Inflammatory to Proinflammatory. Cell Cycle 2012, 11, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Xiao, T.; Sun, M.; Zhao, C.; Kang, J. TRPV1: A Promising Therapeutic Target for Skin Aging and Inflammatory Skin Diseases. Front. Pharmacol. 2023, 14, 1037925. [Google Scholar] [CrossRef]
- Siregar, A.S.; Nyiramana, M.M.; Kim, E.-J.; Shin, E.-J.; Kim, C.-W.; Lee, D.K.; Hong, S.-G.; Han, J.; Kang, D. TRPV1 Is Associated with Testicular Apoptosis in Mice. J. Anim. Reprod. Biotechnol. 2019, 34, 311–317. [Google Scholar] [CrossRef]
- Shin, M.; Buckner, A.; Prince, J.; Bullock, T.N.J.; Hsu, K.L. Diacylglycerol Lipase-β Is Required for TNF-α Response but Not CD8+ T Cell Priming Capacity of Dendritic Cells. Cell Chem. Biol. 2019, 26, 1036–1041.e3. [Google Scholar] [CrossRef]
- Lipina, C.; Walsh, S.K.; Mitchell, S.E.; Speakman, J.R.; Wainwright, C.L.; Hundal, H.S. GPR55 Deficiency Is Associated with Increased Adiposity and Impaired Insulin Signaling in Peripheral Metabolic Tissues. FASEB J. 2019, 33, 1299–1312. [Google Scholar] [CrossRef]
- Mancuso, P.; Bouchard, B. The Impact of Aging on Adipose Function and Adipokine Synthesis. Front. Endocrinol. 2019, 10, 137. [Google Scholar] [CrossRef] [PubMed]
- Kurauti, M.A.; Soares, G.M.; Marmentini, C.; Bronczek, G.A.; Branco, R.C.S.; Boschero, A.C. Insulin and Aging. In Vitamins and Hormones; Academic Press Inc.: Cambridge, MA, USA, 2021; Volume 115, pp. 185–219. ISBN 9780323855488. [Google Scholar]
- Asano, Y. Age-Related Accumulation of Non-Heme Ferric and Ferrous Iron in Mouse Ovarian Stroma Visualized by Sensitive Non-Heme Iron Histochemistry. J. Histochem. Cytochem. 2012, 60, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Di Nisio, V.; Rossi, G.; Chiominto, A.; Pompili, E.; Cecconi, S. Do Aging and Parity Affect VEGF-A/VEGFR Content and Signaling in the Ovary?-A Mouse Model Study. Int. J. Mol. Sci. 2023, 24, 3318. [Google Scholar] [CrossRef] [PubMed]
- Rossi, G.; Dufrusine, B.; Lizzi, A.R.; Luzi, C.; Piccoli, A.; Fezza, F.; Iorio, R.; D’andrea, G.; Dainese, E.; Cecconi, S.; et al. Bisphenol a Deranges the Endocannabinoid System of Primary Sertoli Cells with an Impact on Inhibin b Production. Int. J. Mol. Sci. 2020, 21, 8986. [Google Scholar] [CrossRef] [PubMed]
- Paradisi, A.; Oddi, S.; Maccarrone, M.; Mondino, I.C.; Vergata-Santa, M.-T. The Endocannabinoid System in Ageing: A New Target for Drug Development. Curr. Drug Targets 2006, 7, 1539–1552. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossi, G.; Di Nisio, V.; Chiominto, A.; Cecconi, S.; Maccarrone, M. Endocannabinoid System Components of the Female Mouse Reproductive Tract Are Modulated during Reproductive Aging. Int. J. Mol. Sci. 2023, 24, 7542. https://doi.org/10.3390/ijms24087542
Rossi G, Di Nisio V, Chiominto A, Cecconi S, Maccarrone M. Endocannabinoid System Components of the Female Mouse Reproductive Tract Are Modulated during Reproductive Aging. International Journal of Molecular Sciences. 2023; 24(8):7542. https://doi.org/10.3390/ijms24087542
Chicago/Turabian StyleRossi, Gianna, Valentina Di Nisio, Alessandro Chiominto, Sandra Cecconi, and Mauro Maccarrone. 2023. "Endocannabinoid System Components of the Female Mouse Reproductive Tract Are Modulated during Reproductive Aging" International Journal of Molecular Sciences 24, no. 8: 7542. https://doi.org/10.3390/ijms24087542