
Modifying the Expression of Cysteine Protease Gene PCP Affects Pollen Development, Germination and Plant Drought Tolerance in Maize


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
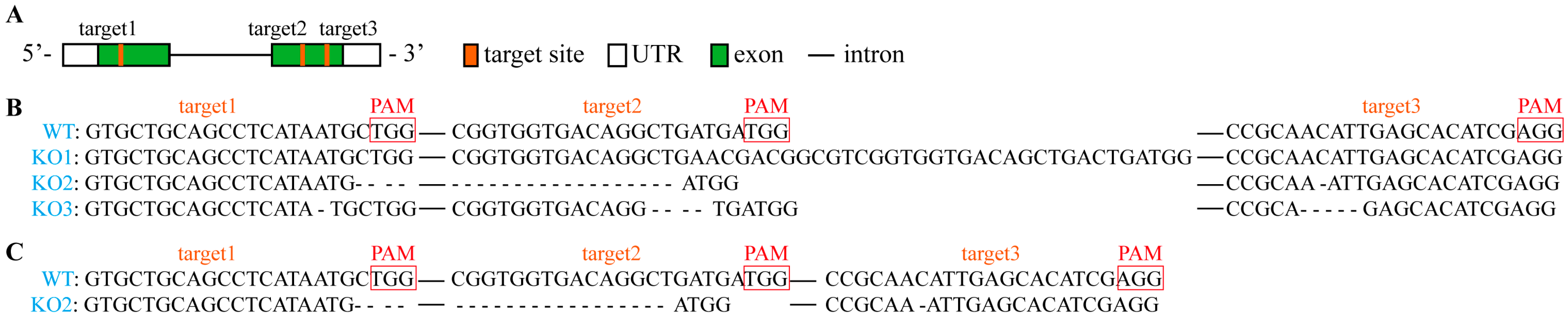
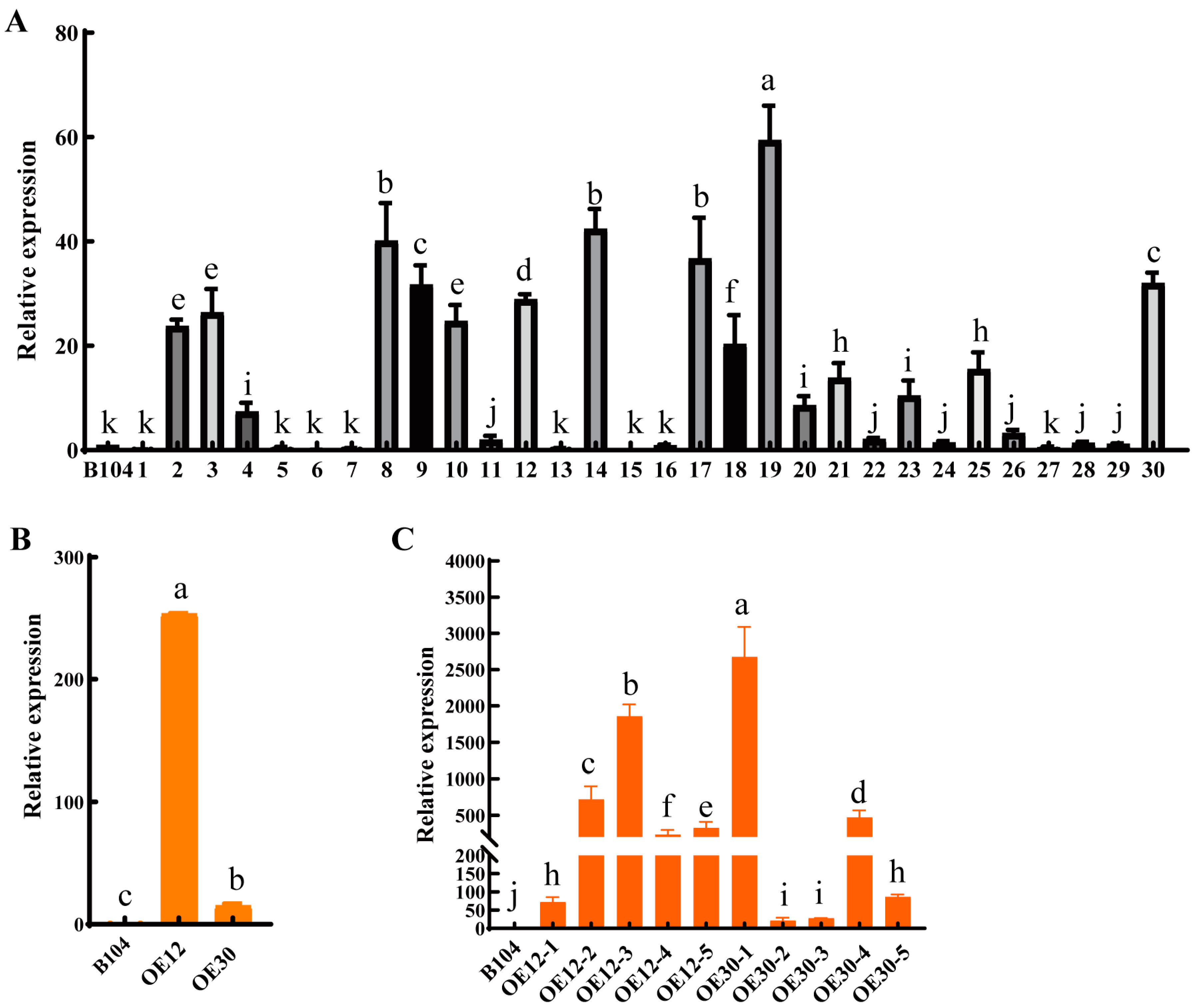
2.1. Identification of the pcp Mutants
2.2. Generation of PCP Overexpressed Lines
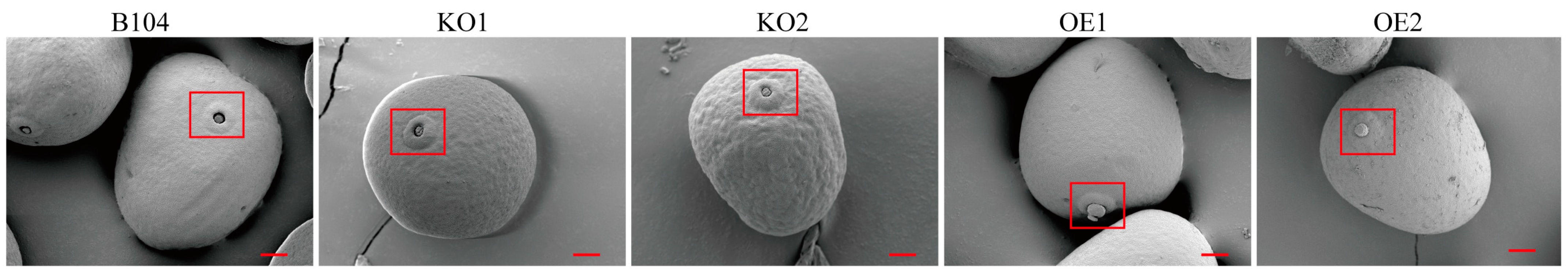
2.3. Pollen Morphology and Structure Comparison
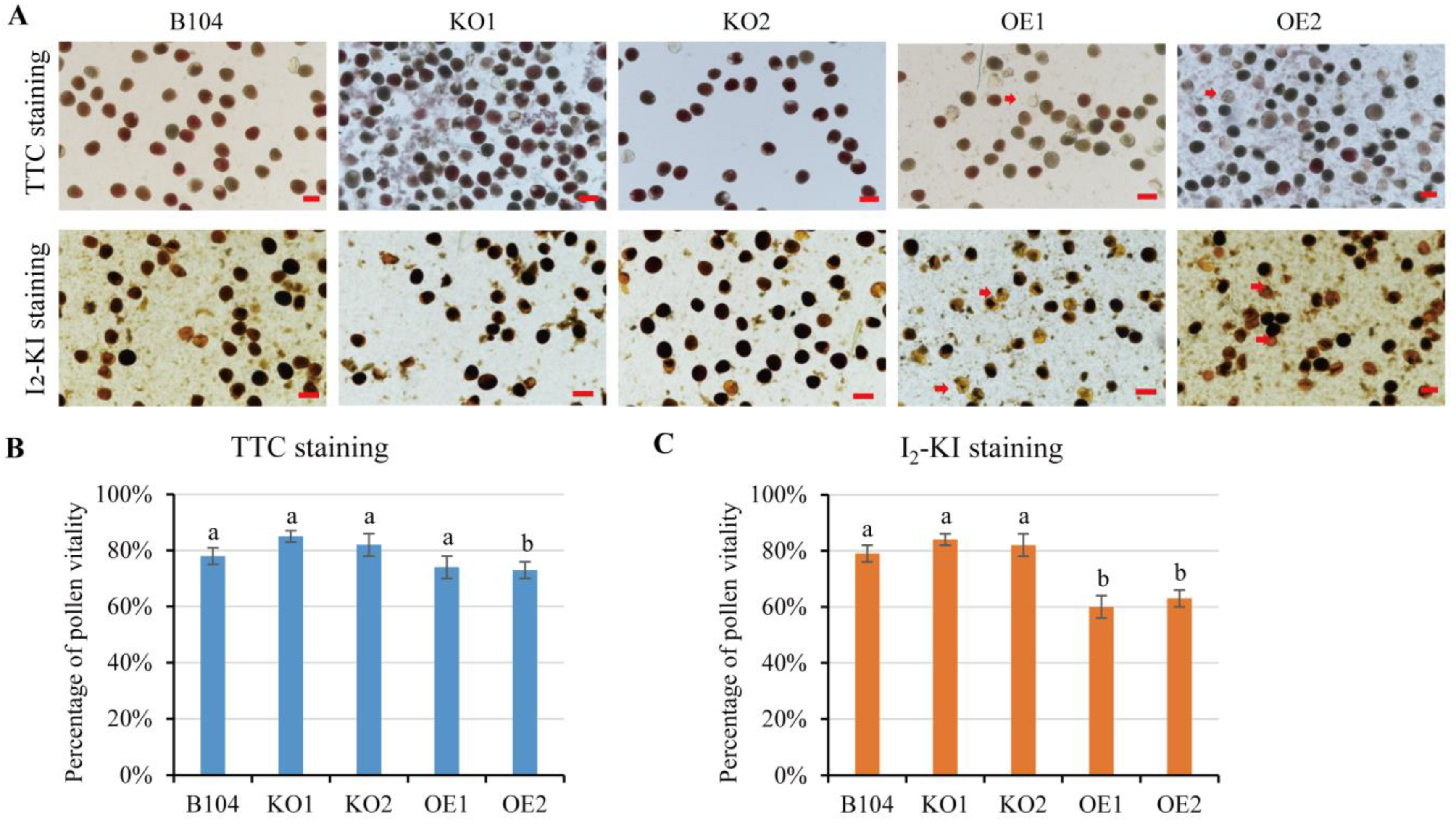
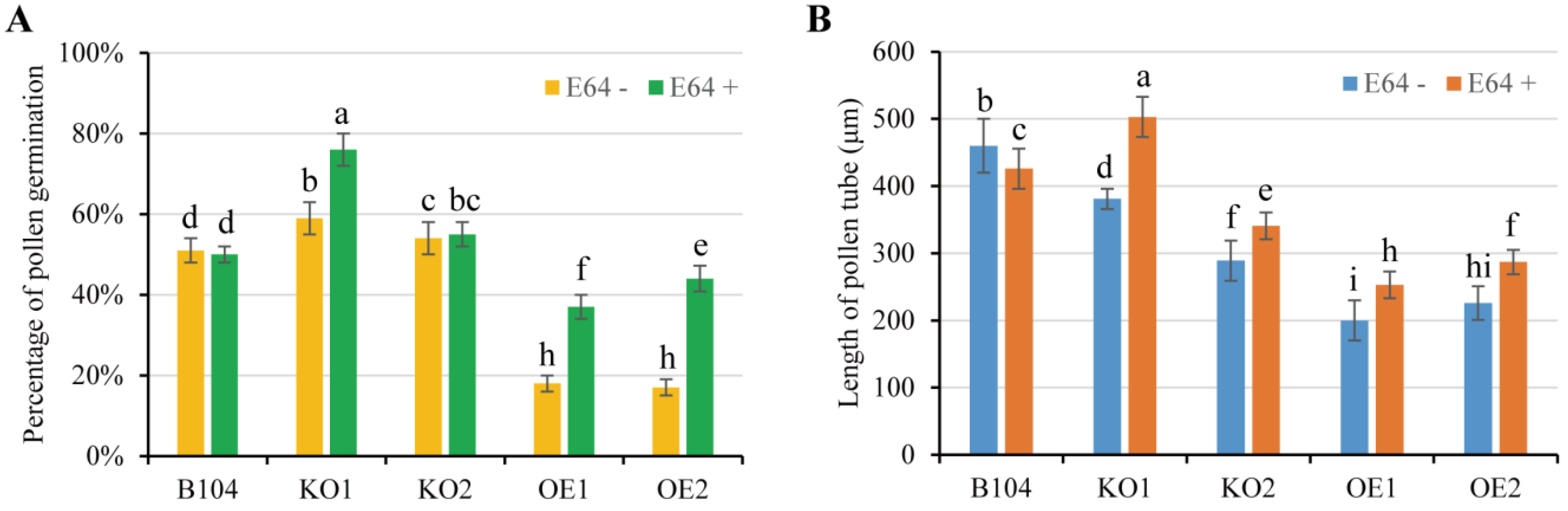
2.4. In Vitro Pollen Viability and Pollen Germination
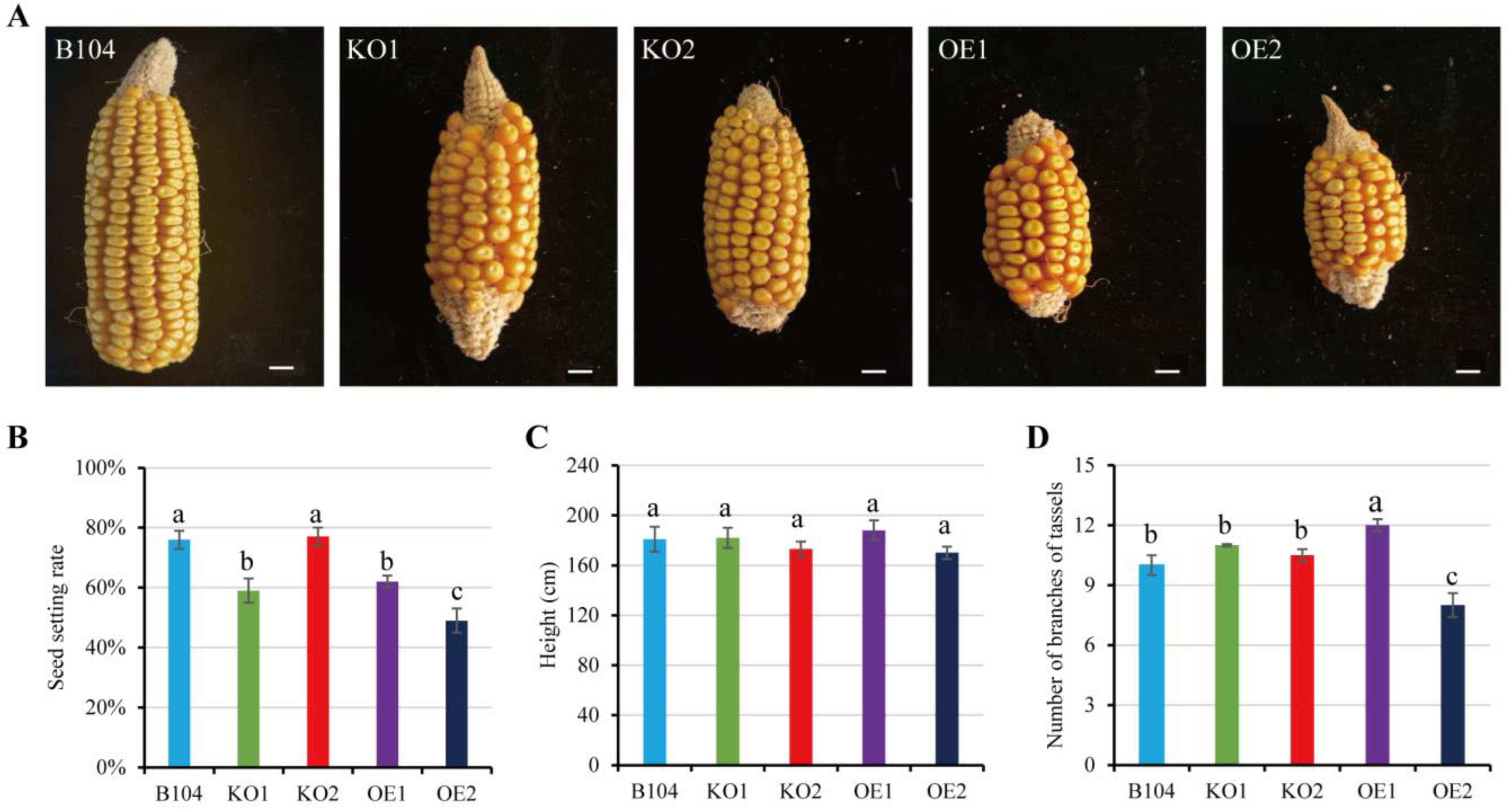
2.5. Comparison of Plant Phenotypes among WT and Transgenic Maize
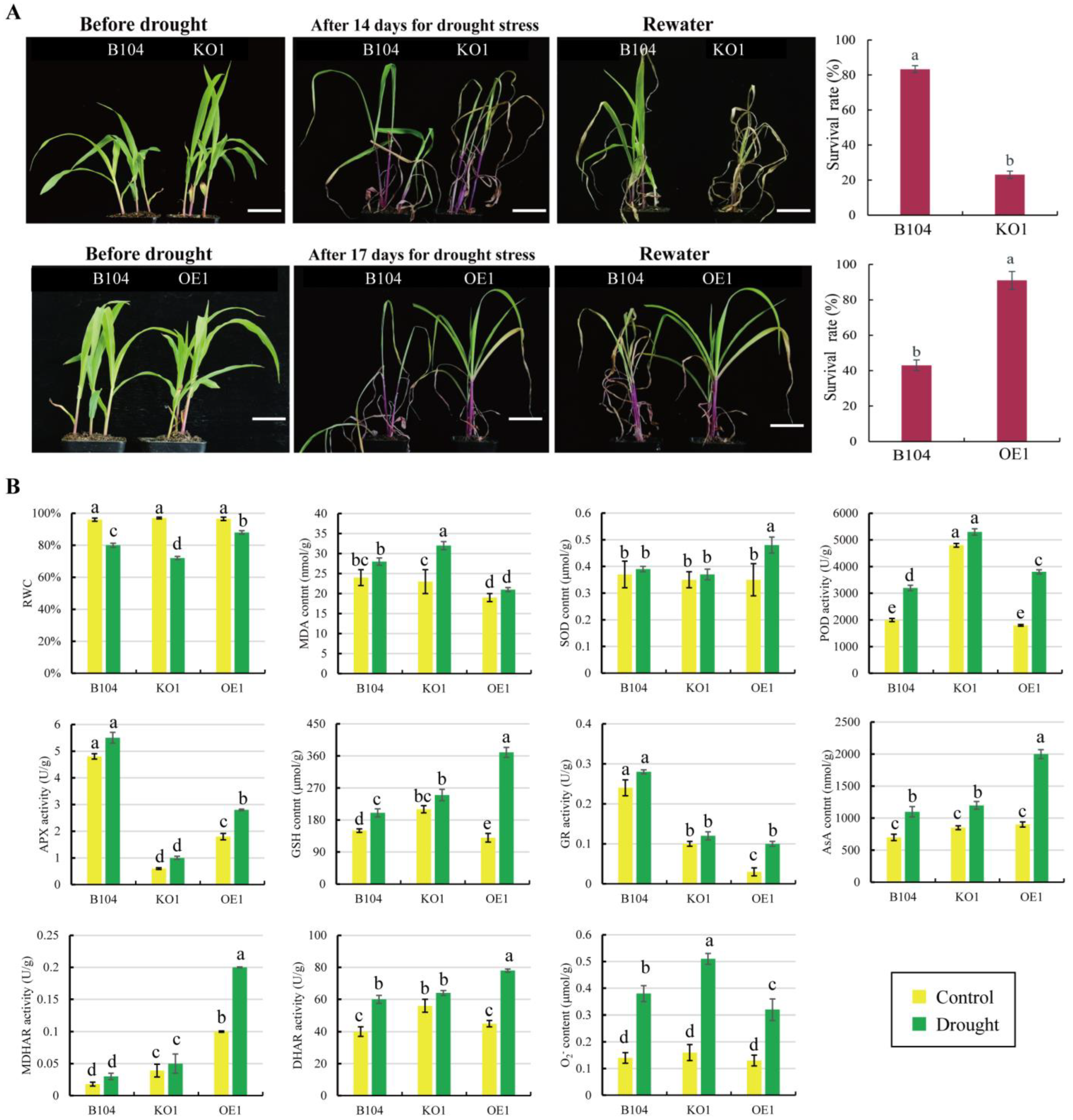
2.6. Comparison of Drought Tolerance between WT Maize and Transgenic Lines
3. Discussion
3.1. The Increased Expression of PCP Negatively Affects Maize Pollen Germination
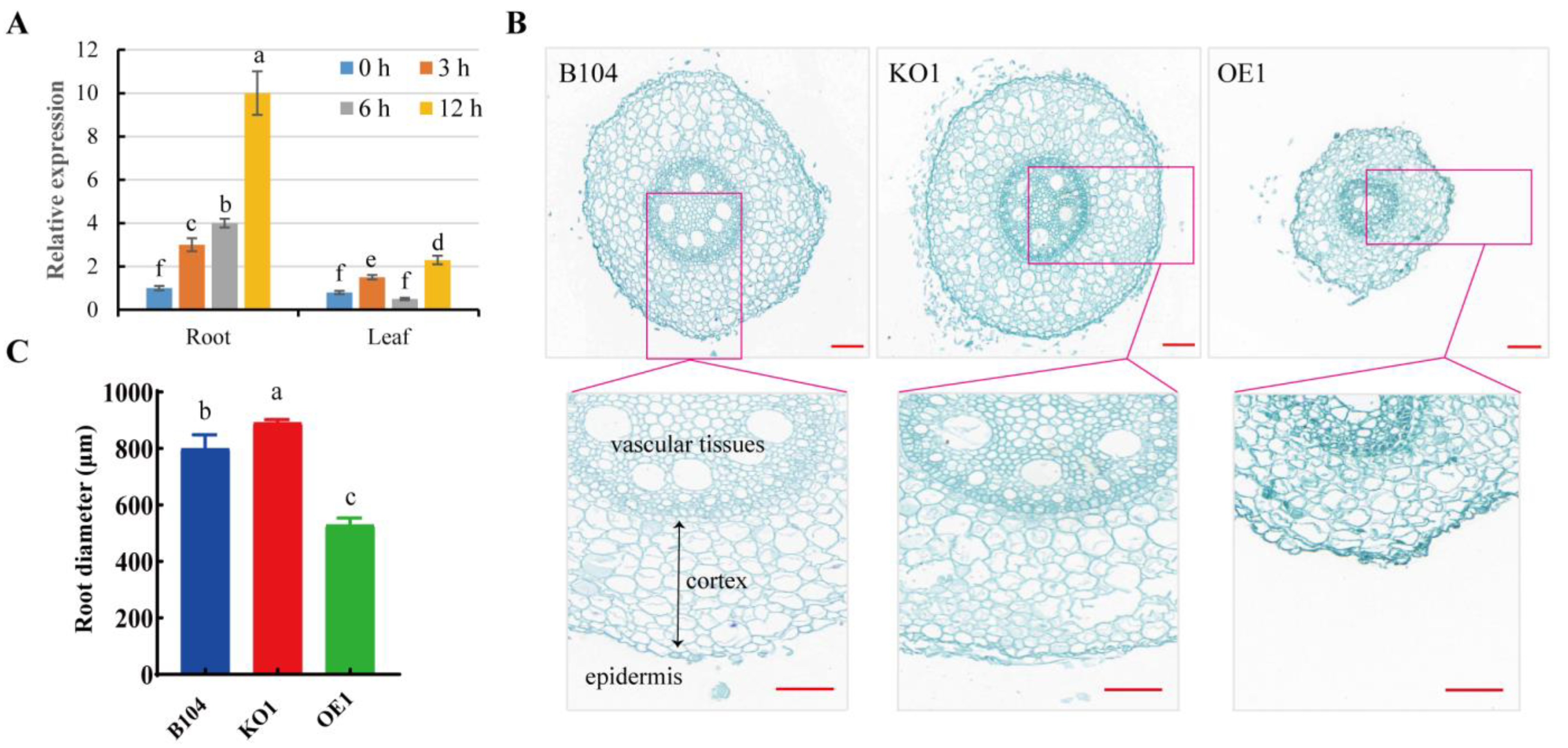
3.2. The Increased Expression of PCP Positively Enhances Maize Drought Tolerance
4. Materials and Methods
4.1. Plant Materials, Growth Conditions and Phenotyping
4.2. Drought Treatment
4.3. Generation of PCP Overexpressed Lines
4.4. Generation of CRISPR-Cas9 Mutants
4.5. Scanning Electron Microscopy (SEM) of Pollen Grains
4.6. Transmission Electron Microscopy (TEM) of Pollen Grains
4.7. Pollen Viability and Germination In Vitro
4.8. Physiological and Biochemical Assays
4.9. Preparation of Paraffin Section of Root
4.10. RT-qPCR
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Solomon, M.; Belenghi, B.; Delledonne, M.; Menachem, E.; Levine, A. The involvement of cysteine proteases and protease inhibitor genes in the regulation of programmed cell death in plants. Plant Cell 1999, 11, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Helm, M.; Schmid, M.; Hierl, G.; Terneus, K.; Tan, L.; Lottspeich, F.; Kieliszewski, M.J.; Gietl, C. KDEL-tailed cysteine endopeptidases involved in programmed cell death, intercalation of new cells, and dismantling of extensin scaffolds. Am. J. Bot. 2008, 95, 1049–1062. [Google Scholar] [CrossRef]
- Richau, K.H.; Kaschani, F.; Verdoes, M.; Pansuriya, T.C.; Niessen, S.; Stüber, K.; Colby, T.; Overkleeft, H.S.; Bogyo, M.; Van der Hoorn, R.A. Subclassification and biochemical analysis of plant papain-like cysteine proteases displays subfamily-specific characteristics. Plant Physiol. 2012, 158, 1583–1599. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Wang, R.; Xiao, J.; Zhu, S.; Li, X.; Han, S.; Li, Z.; Zhao, Y.; Shohag, M.J.I.; He, Z.; et al. An integrated physiology, cytology, and proteomics analysis reveals a network of sugarcane protoplast responses to enzymolysis. Front. Plant Sci. 2022, 13, 1066073. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Liu, D.; Lv, X.; Wang, Y.; Xun, Z.; Liu, Z.; Li, F.; Lu, H. The cysteine protease CEP1, a key executor involved in tapetal programmed cell death, regulates pollen development in Arabidopsis. Plant Cell 2014, 26, 2939–2961. [Google Scholar] [CrossRef]
- Li, Y.; Suen, D.F.; Huang, C.Y.; Kung, S.Y.; Huang, A.H. The maize tapetum employs diverse mechanisms to synthesize and store proteins and flavonoids and transfer them to the pollen surface. Plant Physiol. 2012, 158, 1548–1561. [Google Scholar] [CrossRef]
- Yang, Y.X.; Dong, C.H.; Yu, J.Y.; Shi, L.; Tong, C.B.; Li, Z.B.; Huang, J.Y.; Liu, S.Y. Cysteine Protease 51 (CP51), an anther-specific cysteine protease gene, is essential for pollen exine formation in Arabidopsis. Plant Cell Tissue Organ Cult. 2014, 119, 383–397. [Google Scholar] [CrossRef]
- Zhang, X.M.; Wang, Y.; Lv, X.M.; Li, H.; Sun, P.; Lu, H.; Li, F.L. NtCP56, a new cysteine protease in Nicotiana tabacum L., involved in pollen grain development. J. Exp. Bot. 2009, 60, 1569–1577. [Google Scholar] [CrossRef]
- Lee, S.; Jung, K.H.; An, G.; Chung, Y.Y. Isolation and characterization of a rice cysteine protease gene, OsCp1, using T-DNA gene-trap system. Plant Mol. Biol. 2004, 54, 755–765. [Google Scholar] [CrossRef]
- Edlund, A.F.; Swanson, R.; Preuss, D. Pollen and stigma structure and function: The role of diversity in pollination. Plant Cell 2002, 16, S84–S97. [Google Scholar] [CrossRef]
- Suen, D.F.; Wu, S.S.H.; Chang, H.C.; Dhugga, K.S.; Huang, A.H.C. Cell wall reactive proteins in the coat and wall of maize pollen: Potential role in pollen tube growth on the stigma and through the style. J. Biol. Chem. 2003, 278, 43672–43681. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Cai, G.; Gong, F.P.; An, S.; Cresti, M.; Wang, W. Proteome profiling of maize pollen coats reveals novel protein components. Plant Mol. Biol. Rep. 2015, 33, 975–986. [Google Scholar] [CrossRef]
- Li, Y.; Niu, L.; Wu, X.; Faleri, C.; Tai, F.; Zhang, M.; Liu, H.; Wang, W.; Cai, G. Genome-wide identification and comparison of cysteine proteases in the pollen coat and other tissues in maize. Front. Plant Sci. 2021, 12, 709534. [Google Scholar] [CrossRef] [PubMed]
- Mayfield, J.A.; Fiebig, A.; Johnstone, S.E.; Preuss, D. Gene families from the Arabidopsis thaliana pollen coat proteome. Science 2001, 292, 2482–2485. [Google Scholar] [CrossRef]
- Gong, F.; Wu, X.; Wang, W. Diversity and function of maize pollen coat proteins: From biochemistry to proteomics. Front. Plant Sci. 2015, 6, 199. [Google Scholar] [CrossRef]
- Bih, F.Y.; Wu, S.S.; Ratnayake, C.; Walling, L.L.; Nothnagel, E.A.; Huang, A.H. The predominant protein on the surface of maize pollen is an endoxylanase synthesized by a tapetum mRNA with a long 5′ leader. J. Biol. Chem. 1999, 274, 22884–22894. [Google Scholar] [CrossRef]
- Liu, H.; Hu, M.; Wang, Q.; Cheng, L.; Zhang, Z. Role of papain-like cysteine proteases in plant development. Front. Plant Sci. 2018, 9, 1717. [Google Scholar] [CrossRef]
- Chen, H.J.; Su, C.T.; Lin, C.H.; Huang, G.J.; Lin, Y.H. Expression of sweet potato cysteine protease SPCP2 altered developmental characteristics and stress responses in transgenic Arabidopsis plants. J. Plant Physiol. 2010, 167, 838–847. [Google Scholar] [CrossRef]
- Zang, Q.W.; Wang, C.X.; Li, X.Y.; Guo, Z.A.; Jing, R.L.; Zhao, J.; Chang, X.P. Isolation and characterization of a gene encoding a polyethylene glycol-induced cysteine protease in common wheat. J. Biosci. 2010, 35, 379–388. [Google Scholar] [CrossRef]
- Simova-Stoilova, L.; Vaseva, I.; Grigorova, B.; Demirevska, K.; Feller, U. Proteolytic activity and cysteine protease expression in wheat leaves under severe soil drought and recovery. Plant Physiol. Bioch. 2010, 48, 200–206. [Google Scholar] [CrossRef]
- Yingqi, H.; Lv, Y.; Zhang, J.; Ahmad, N.; Li, Y.; Wang, N.; Xiuming, L.; Na, Y.; Li, X. Identification and functional characterization of safflower cysteine protease 1 as negative regulator in response to low-temperature stress in transgenic Arabidopsis. Planta 2022, 255, 106. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Cui, G.; Wang, Y.; Hao, Y.; Du, J.; Zhang, H.; Wang, C.; Zhang, H.; Wu, S.B.; Sun, Y. Expression of foreign genes demonstrates the effectiveness of pollen-mediated transformation in Zea mays. Front. Plant Sci. 2017, 8, 383. [Google Scholar] [CrossRef] [PubMed]
- Furness, C.A.; Rudall, P.J. The operculum in pollen of monocotyledons. Aliso 2006, 22, 191–196. [Google Scholar] [CrossRef]
- Wang, W.; Milanesi, C.; Faleri, C.; Cresti, M. Localization of group-1 allergen Zea m 1 in the coat and wall of maize pollen. Acta Histochem. 2006, 108, 395–400. [Google Scholar] [CrossRef]
- Yennawar, N.H.; Li, L.C.; Dudzinski, D.M.; Tabuchi, A.; Cosgrove, D.J. Crystal structure and activities of EXPB1 (Zea m 1), a beta-expansin and group-1 pollen allergen from maize. Proc. Natl. Acad. Sci. USA 2006, 103, 14664–14671. [Google Scholar] [CrossRef]
- Katifori, E.; Alben, S.; Cerda, E.; Nelson, D.R.; Dumais, J. Foldable structures and the natural design of pollen grains. Proc. Natl. Acad. Sci. USA 2010, 107, 7635–7639. [Google Scholar] [CrossRef]
- Dobritsa, A.A.; Kirkpatrick, A.B.; Reeder, S.H.; Li, P.; Owen, H.A. Pollen aperture factor INP1 acts late in aperture formation by excluding specific membrane domains from exine deposition. Plant Physiol. 2018, 176, 326–339. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, G.; Tan, Q.; Yuan, H.; Betts, N.; Zhu, L.; Zhang, D.; Liang, W. Rice pollen aperture formation is regulated by the interplay between OsINP1 and OsDAF1. Nat. Plants 2020, 6, 394–403. [Google Scholar] [CrossRef]
- Dobritsa, A.A.; Coerper, D. The novel plant protein INAPERTURATE POLLEN1 marks distinct cellular domains and controls formation of apertures in the Arabidopsis pollen exine. Plant Cell 2012, 24, 4452–4464. [Google Scholar] [CrossRef]
- Li, P.; Ben-Menni Schuler, S.; Reeder, S.H.; Wang, R.; Suárez Santiago, V.N.; Dobritsa, A.A. INP1 involvement in pollen aperture formation is evolutionarily conserved and may require species-specific partners. J. Exp. Bot. 2018, 69, 983–996. [Google Scholar] [CrossRef]
- Albert, B.; Ressayre, A.; Dillmann, C.; Carlson, A.L.; Swanson, R.J.; Gouyon, P.H.; Dobritsa, A.A. Effect of aperture number on pollen germination, survival and reproductive success in Arabidopsis thaliana. Ann. Bot. 2018, 121, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tao, H.; Tian, B.; Sheng, D.; Xu, C.; Zhou, H.; Huang, S.; Wang, P. Flowering dynamics, pollen, and pistil contribution to grain yield in response to high temperature during maize flowering. Environ. Exp. Bot. 2019, 158, 80–88. [Google Scholar] [CrossRef]
- Xu, Y.; Yang, J.; Wang, Y.; Wang, J.; Yu, Y.; Long, Y.; Wang, Y.; Zhang, H.; Ren, Y.; Chen, J.; et al. OsCNGC13 promotes seed-setting rate by facilitating pollen tube growth in stylar tissues. PLoS Genet. 2017, 13, e1006906. [Google Scholar] [CrossRef] [PubMed]
- Nazish, M.; Althobaiti, A.T. Palyno-morphological characteristics as a systematic approach in the identification of halophytic Poaceae species from a saline environment. Plants 2022, 11, 2618. [Google Scholar] [CrossRef]
- Datta, R.; Chamusco, K.C.; Chourey, P.S. Starch biosynthesis during pollen maturation is associated with altered patterns of gene expression in maize. Plant Physiol. 2002, 130, 1645–1656. [Google Scholar] [CrossRef]
- Lee, S.K.; Kim, H.; Cho, J.I.; Nguyen, C.D.; Moon, S.; Park, J.E.; Park, H.R.; Huh, J.H.; Jung, K.H.; Guiderdoni, E.; et al. Deficiency of rice hexokinase HXK5 impairs synthesis and utilization of starch in pollen grains and causes male sterility. J. Exp. Bot. 2020, 71, 116–125. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, X.; Chen, R.; Yang, L.; Fan, K.; Liu, Y.; Wang, G.; Ren, Z.; Liu, Y. Generation of transgene-free semidwarf maize plants by gene editing of Gibberellin-oxidase20-3 using CRISPR/Cas9. Front. Plant Sci. 2020, 11, 1048. [Google Scholar] [CrossRef]
- Liu, W.Y.; Lin, H.H.; Yu, C.P.; Chang, C.K.; Chen, H.J.; Lin, J.J.; Lu, M.J.; Tu, S.L.; Shiu, S.H.; Wu, S.H.; et al. Maize ANT1 modulates vascular development, chloroplast development, photosynthesis, and plant growth. Proc. Natl. Acad. Sci. USA 2020, 117, 21747–21756. [Google Scholar] [CrossRef]
- Luo, Y.; Zhang, M.; Liu, Y.; Liu, J.; Li, W.; Chen, G.; Peng, Y.; Jin, M.; Wei, W.; Jian, L.; et al. Genetic variation in YIGE1 contributes to ear length and grain yield in maize. New Phytol. 2022, 234, 513–526. [Google Scholar] [CrossRef]
- Li, Q.; Wu, G.; Zhao, Y.; Wang, B.; Zhao, B.; Kong, D.; Wei, H.; Chen, C.; Wang, H. CRISPR/Cas9-mediated knockout and overexpression studies reveal a role of maize phytochrome C in regulating flowering time and plant height. Plant Biotechnol. J. 2020, 18, 2520–2532. [Google Scholar] [CrossRef]
- Wang, W.; Zhao, P.; Zhou, X.M.; Xiong, H.X.; Sun, M.X. Genome-wide identification and characterization of cystatin family genes in rice (Oryza sativa L.). Plant Cell Rep. 2015, 34, 1579–1592. [Google Scholar] [CrossRef] [PubMed]
- D’Ippólito, S.; Rey-Burusco, M.F.; Feingold, S.E.; Guevara, M.G. Role of proteases in the response of plants to drought. Plant Physiol. Bioch. 2021, 168, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Golldack, D.; Li, C.; Mohan, H.; Probst, N. Tolerance to drought and salt stress in plants: Unraveling the signaling networks. Front. Plant Sci. 2014, 5, 151. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.A.; Tanveer, M.; Ashraf, U.; Hussain, S.; Shahzad, B.; Khan, I.; Wang, L. Effect of progressive drought stress on growth, leaf gas exchange, and antioxidant production in two maize cultivars. Environ. Sci. Pollut. Res. 2016, 23, 17132–17141. [Google Scholar] [CrossRef] [PubMed]
- Scharwies, J.D.; Dinneny, J.R. Water transport, perception, and response in plants. J. Plant Res. 2019, 132, 311–324. [Google Scholar] [CrossRef]
- Ranjan, A.; Sinha, R.; Singla-Pareek, S.L.; Pareek, A.; Singh, A.K. Shaping the root system architecture in plants for adaptation to drought stress. Physiol. Plant 2022, 174, e13651. [Google Scholar] [CrossRef]
- Lopez-Valdivia, I.; Perkins, A.C.; Schneider, H.M.; Vallebueno-Estrada, M.; Burridge, J.D.; González-Orozco, E.; Montufar, A.; Montiel, R.; Lynch, J.P.; Vielle-Calzada, J.P. Gradual domestication of root traits in the earliest maize from Tehuacán. Proc. Natl. Acad. Sci. USA 2022, 119, e2110245119. [Google Scholar] [CrossRef]
- Li, Z.; Liu, C.; Zhang, Y.; Wang, B.; Ran, Q.; Zhang, J. The bHLH family member ZmPTF1 regulates drought tolerance in maize by promoting root development and abscisic acid synthesis. J. Exp. Bot. 2019, 70, 5471–5486. [Google Scholar] [CrossRef]
- Jaramillo, R.E.; Nord, E.A.; Chimungu, J.G.; Brown, K.M.; Lynch, J.P. Root cortical burden influences drought tolerance in maize. Ann. Bot. 2013, 112, 429–437. [Google Scholar] [CrossRef]
- Hazman, M.Y.; Kabil, F.F. Maize root responses to drought stress depend on root class and axial position. J. Plant Res. 2022, 135, 105–120. [Google Scholar] [CrossRef]
- Zhu, J.; Brown, K.M.; Lynch, J.P. Root cortical aerenchyma improves the drought tolerance of maize (Zea mays L.). Plant Cell Environ. 2010, 33, 740–749. [Google Scholar] [PubMed]
- Chimungu, J.G.; Brown, K.M.; Lynch, J.P. Large root cortical cell size improves drought tolerance in maize. Plant Physiol. 2014, 166, 2166–2178. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Ramírez, C.; Guillotin, B.; Xu, X.; Rahni, R.; Zhang, S.; Yan, Z.; Coqueiro Dias Araujo, P.; Demesa-Arevalo, E.; Lee, L.; Van Eck, J.; et al. Ground tissue circuitry regulates organ complexity in maize and Setaria. Science 2021, 374, 1247–1252. [Google Scholar] [CrossRef] [PubMed]
- Namyslov, J.; Bauriedlová, Z.; Janoušková, J.; Soukup, A.; Tylová, E. Exodermis and endodermis respond to nutrient deficiency in nutrient-specific and localized manner. Plants 2020, 9, 201. [Google Scholar] [CrossRef] [PubMed]
- Gong, F.; Takahashi, H.; Omori, F.; Wang, W.; Mano, Y.; Nakazono, M. QTLs for constitutive aerenchyma from Zea nicaraguensis improve tolerance of maize to root-zone oxygen deficiency. J. Exp. Bot. 2019, 70, 6475–6487. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, H.; Wu, X.; Wang, W. Identification of drought tolerant mechanisms in a drought-tolerant maize mutant based on physiological, biochemical and transcriptomic analyses. BMC Plant Biol. 2020, 20, 315. [Google Scholar] [CrossRef]
- Frame, B.R.; Shou, H.; Chikwamba, R.K.; Zhang, Z.; Xiang, C.; Fonger, T.M.; Pegg, S.E.; Li, B.; Nettleton, D.S.; Pei, D.; et al. Agrobacterium tumefaciens-mediated transformation of maize embryos using a standard binary vector system. Plant Physiol. 2002, 129, 13–22. [Google Scholar] [CrossRef]
- Bae, S.; Park, J.; Kim, J.S. Cas-OFFinder: A fast and versatile algorithm that searches for potential off-target sites of Cas9 RNA-guided endonucleases. Bioinformatics 2014, 30, 1473–1475. [Google Scholar] [CrossRef]
- Kaefer, K.A.C.; Chiapetti, R.; Fogaça, L.; Muller, A.L.; Calixto, G.B.; Chaves, E.I.D. Viability of maize pollen grains in vitro collected at different times of the day. Afr. J. Agr. Res. 2016, 11, 1040–1047. [Google Scholar]
- Du, G.; Xu, J.; Gao, C.; Lu, J.; Li, Q.; Du, J.; Lv, M.; Sun, X. Effect of low storage temperature on pollen viability of fifteen herbaceous peonies. Biotechnol. Rep. 2019, 21, e00309. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhao, P.; Wu, X.; Wang, W. Proteomic identification of differentially expressed proteins in mature and germinated maize pollen. Acta Physiol. Plant 2011, 33, 1467–1474. [Google Scholar] [CrossRef]
- Matsumoto, K.; Mizoue, K.; Kitamura, K.; Tse, W.C.; Huber, C.P.; Ishida, T. Structural basis of inhibition of cysteine proteases by E-64 and its derivatives. Biopolymers 1999, 51, 99–107. [Google Scholar] [CrossRef]
- Chebli, Y.; Pujol, L.; Shojaeifard, A.; Brouwer, I.; van Loon, J.J.; Geitmann, A. Cell wall assembly and intracellular trafficking in plant cells are directly affected by changes in the magnitude of gravitational acceleration. PLoS ONE 2013, 8, e58246. [Google Scholar] [CrossRef] [PubMed]
- Pryor, W.A. On the detection of lipid hydroperoxides in biological samples. Free. Radic. Biol. Med. 1989, 7, 177–178. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I.; Kode, A.; Biswas, S. Assay for quantitative determination of glutathione and glutathione disulfide levels using enzymatic recycling method. Nat. Protoc. 2006, 1, 3159–3165. [Google Scholar] [CrossRef]
- Hu, X.; Liu, R.; Li, Y.; Wang, W.; Tai, F.; Xue, R.; Li, C. Heat shock protein 70 regulates the abscisic acid-induced antioxidant response of maize to combined drought and heat stress. Plant Growth Regul. 2010, 60, 225–235. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Niu, L.; Zhou, X.; Liu, H.; Tai, F.; Wang, W. Modifying the Expression of Cysteine Protease Gene PCP Affects Pollen Development, Germination and Plant Drought Tolerance in Maize. Int. J. Mol. Sci. 2023, 24, 7406. https://doi.org/10.3390/ijms24087406
Li Y, Niu L, Zhou X, Liu H, Tai F, Wang W. Modifying the Expression of Cysteine Protease Gene PCP Affects Pollen Development, Germination and Plant Drought Tolerance in Maize. International Journal of Molecular Sciences. 2023; 24(8):7406. https://doi.org/10.3390/ijms24087406
Chicago/Turabian StyleLi, Yanhua, Liangjie Niu, Xiaoli Zhou, Hui Liu, Fuju Tai, and Wei Wang. 2023. "Modifying the Expression of Cysteine Protease Gene PCP Affects Pollen Development, Germination and Plant Drought Tolerance in Maize" International Journal of Molecular Sciences 24, no. 8: 7406. https://doi.org/10.3390/ijms24087406