
Potential Benefits of Dietary Plant Compounds on Normal and Tumor Brain Cells in Humans: In Silico and In Vitro Approaches
 , , , and
, , , and 
Abstract
:1. Introduction
2. Results
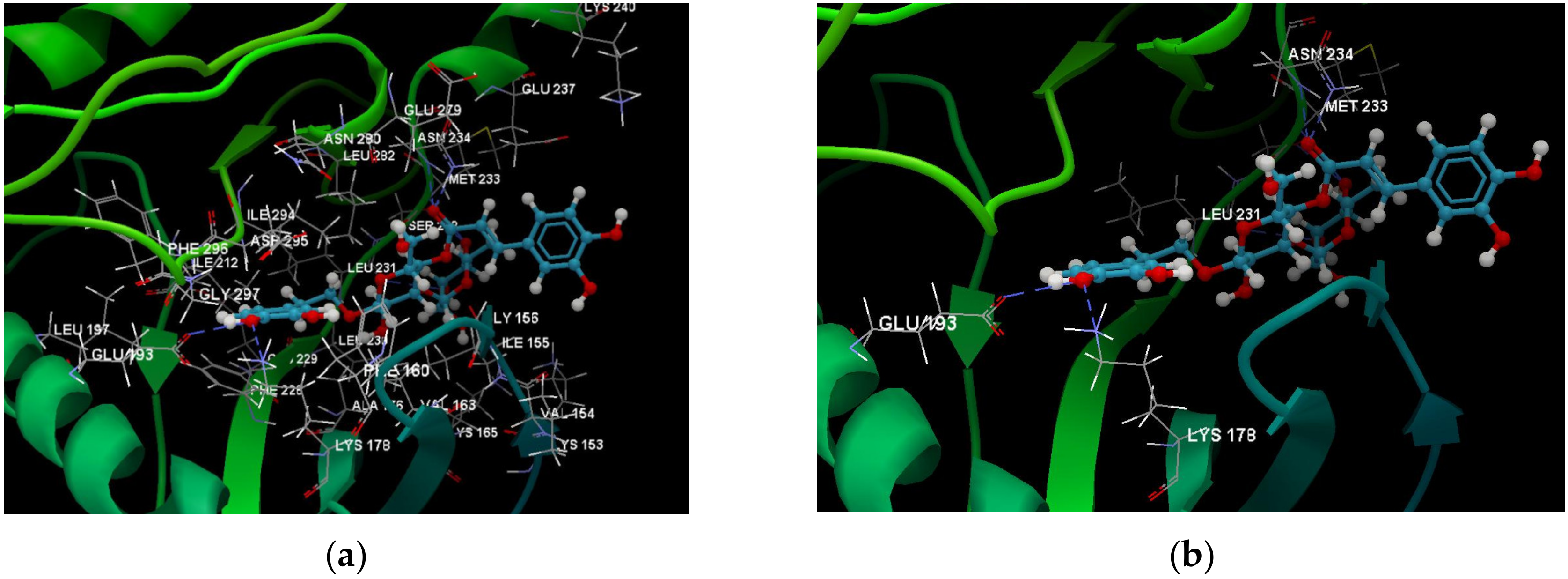
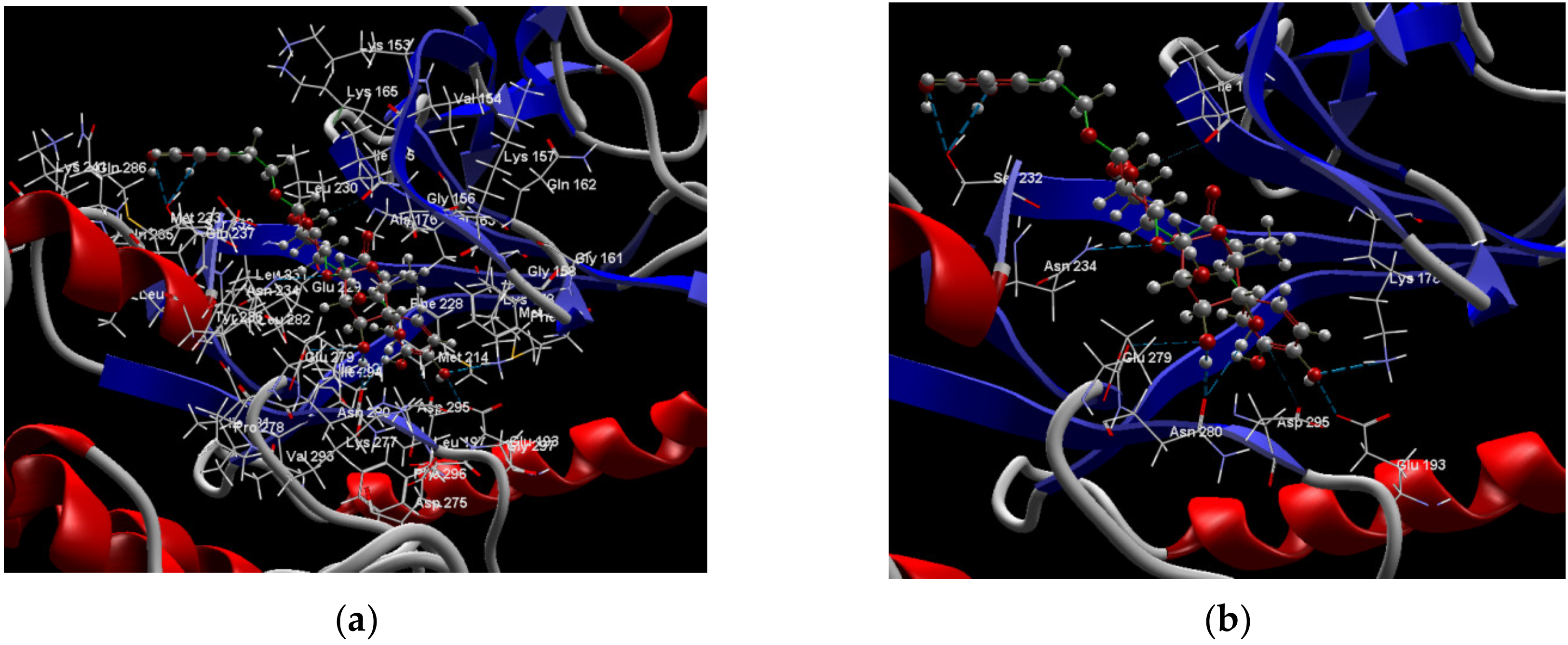
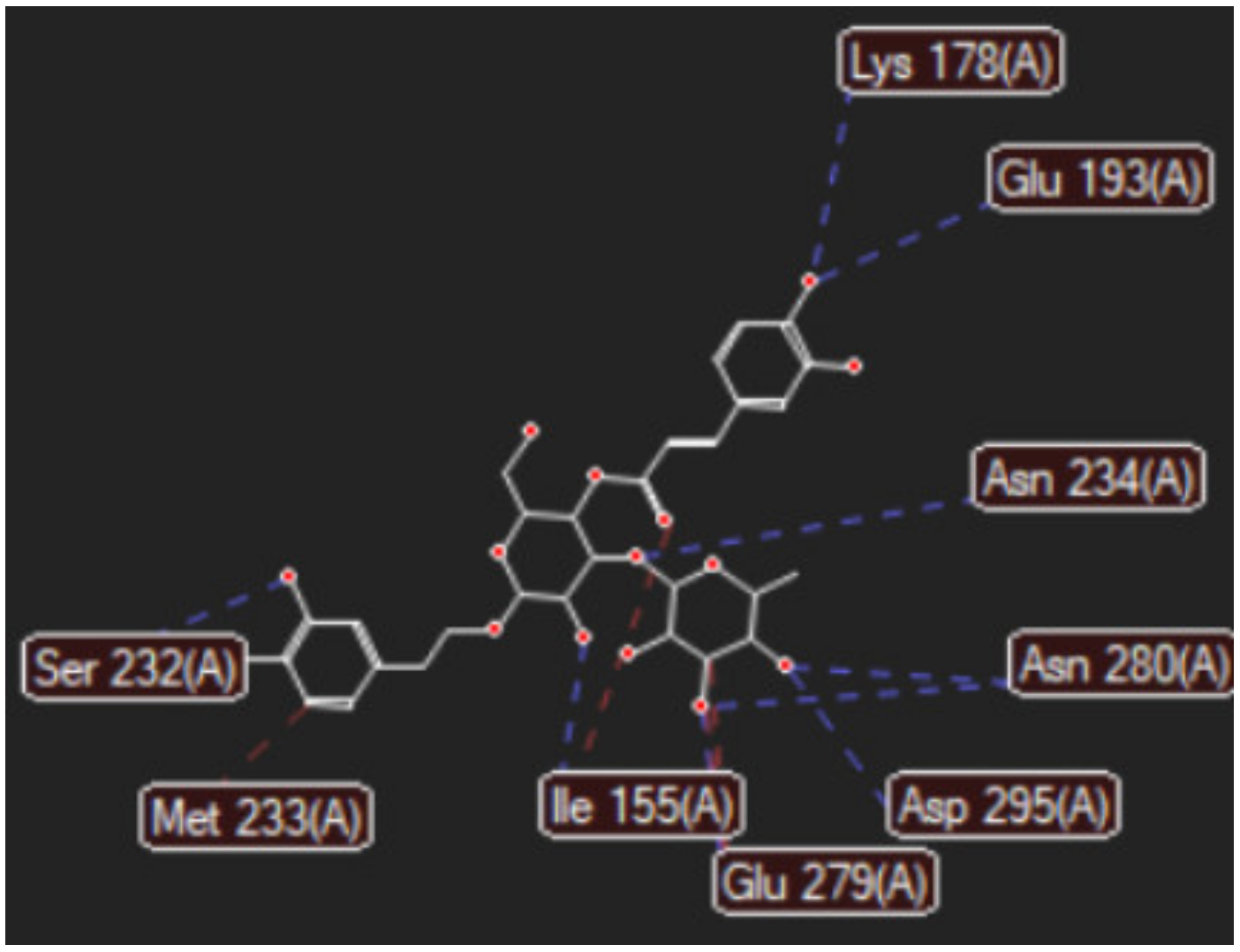
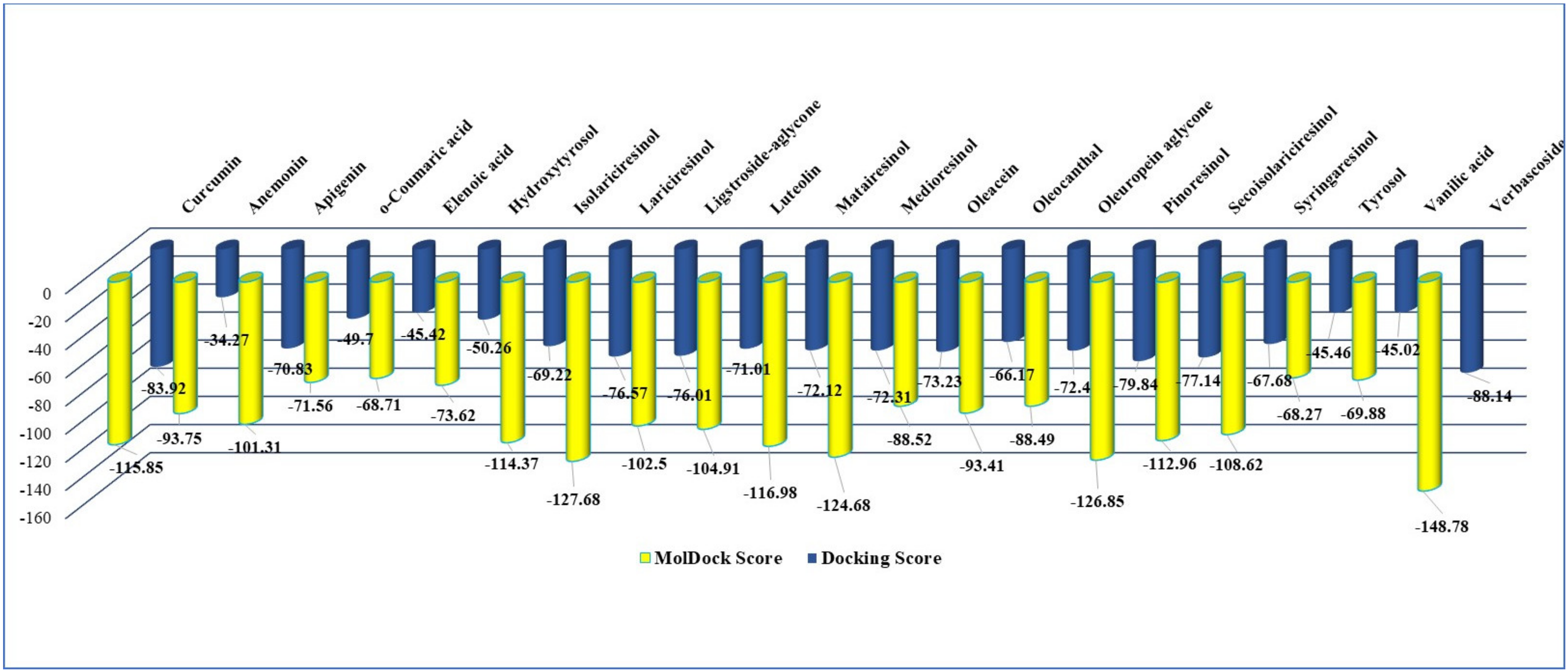
2.1. In Silico Studies Results
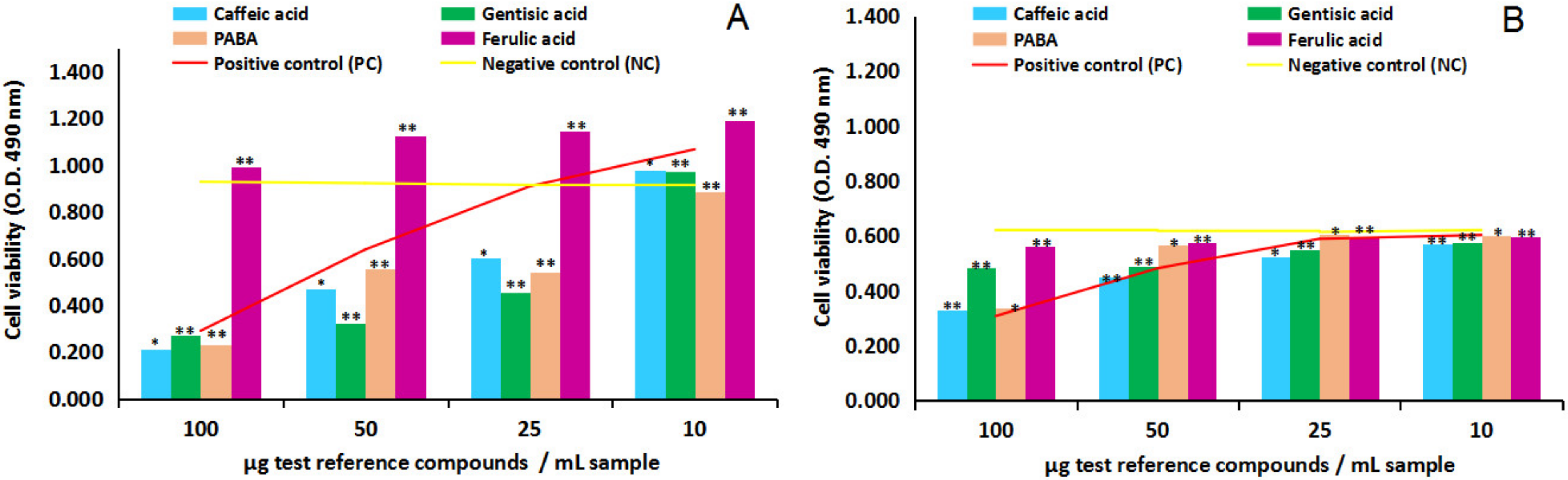
2.2. In Vitro Pharmacological Studies Results
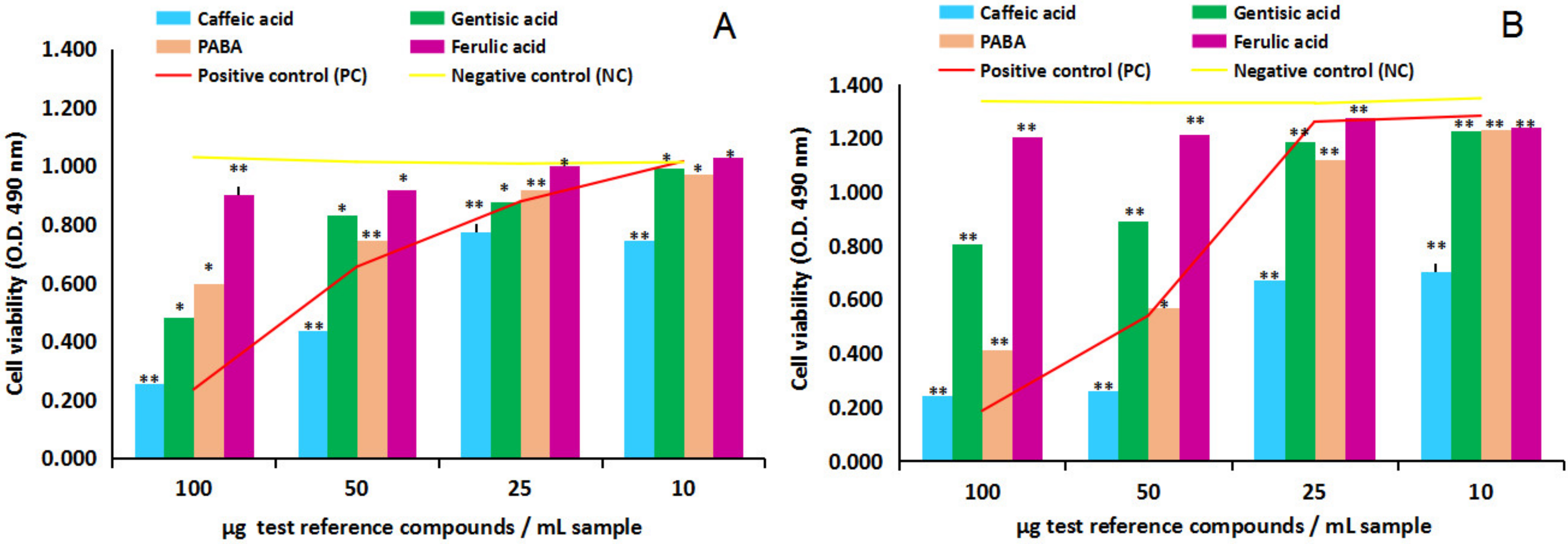
2.2.1. Cytotoxicity and Anti-Proliferative Activity of the Four Test Reference Compounds on Normal Human Astrocytes (NHA) and Human Glioma Cell Line (U87)
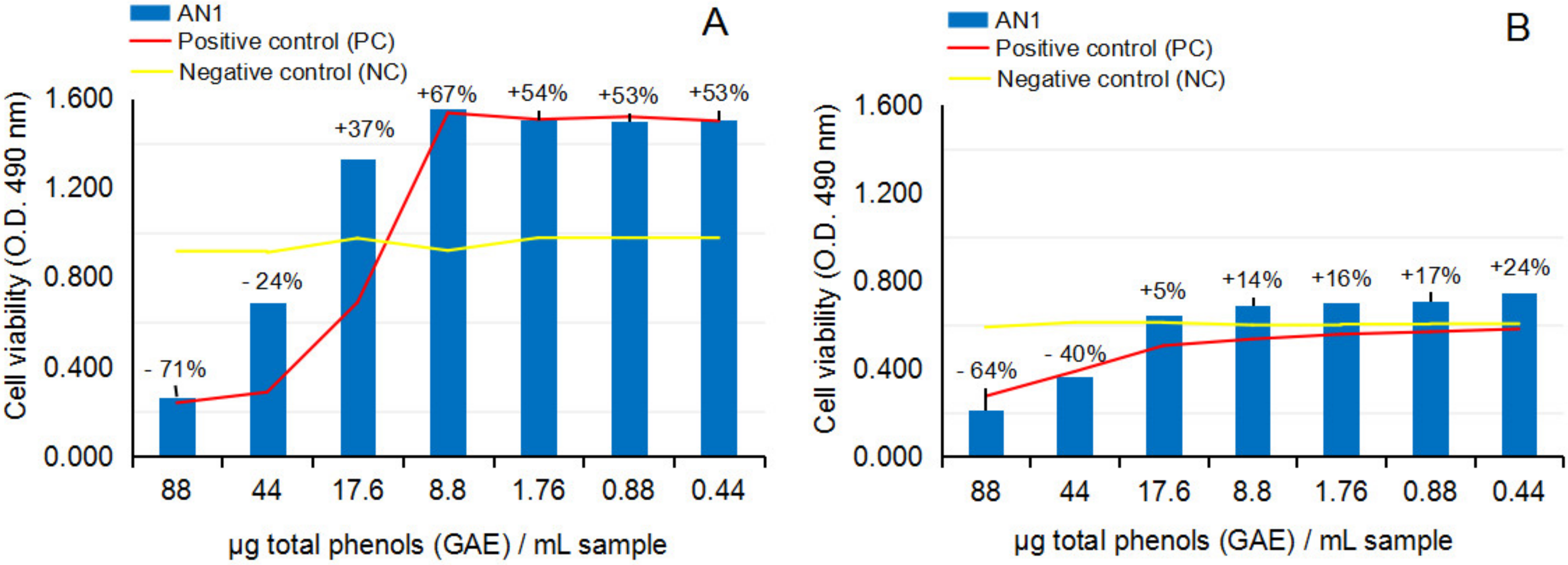
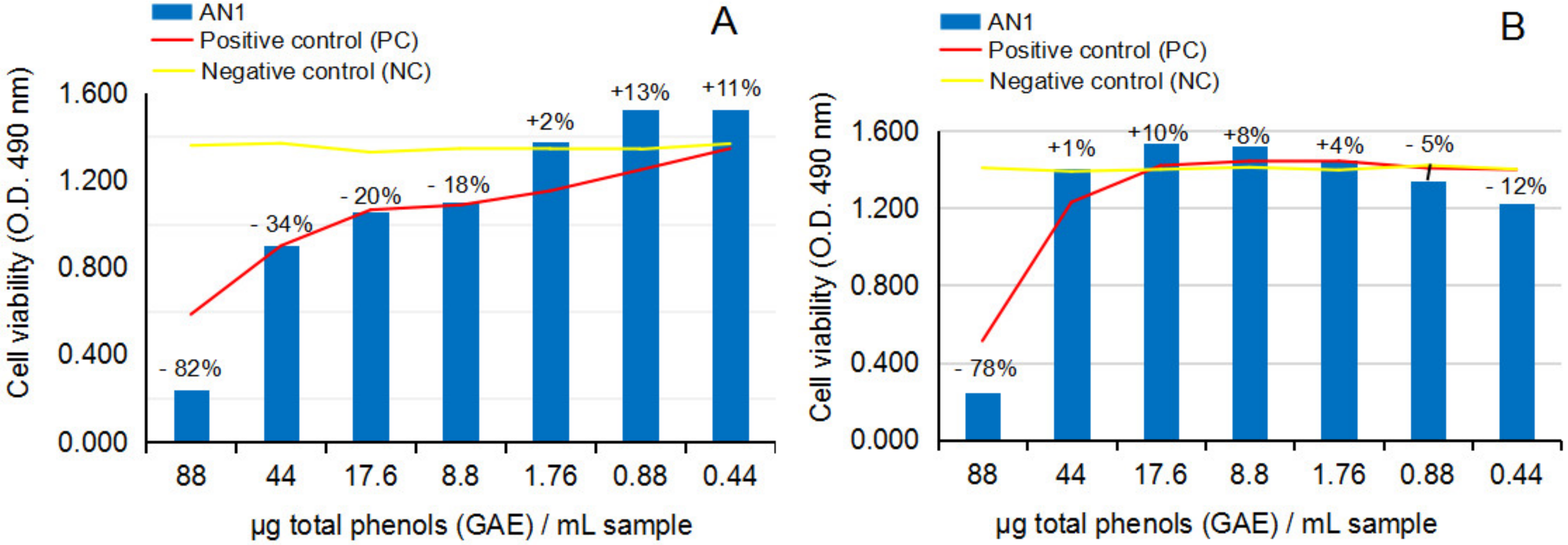
2.2.2. Cytotoxicity and Anti-Proliferative Activity of the Test Vegetal Extract AN1 on Normal Human Astrocytes (NHA) and Human Glioma Cell Line (U87)
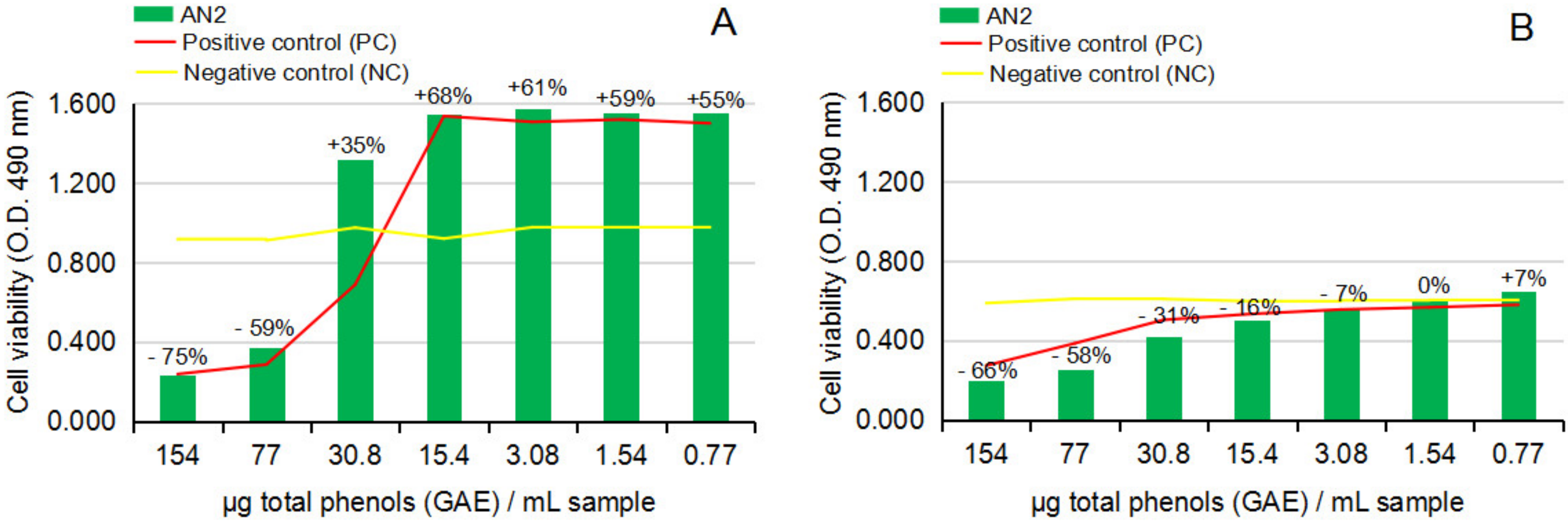
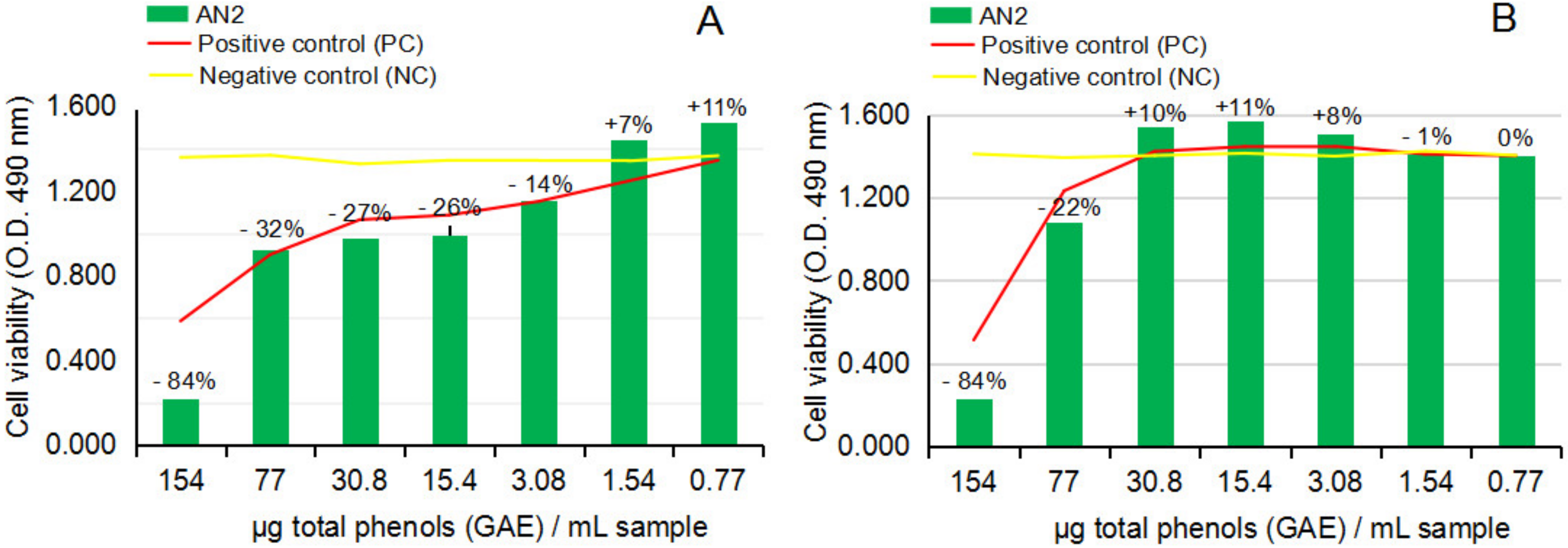
2.2.3. Cytotoxicity and Anti-Proliferative Activity of the Test Vegetal Extract AN2 on Normal Human Astrocytes (NHA) and Human Glioma Cell Line (U87)
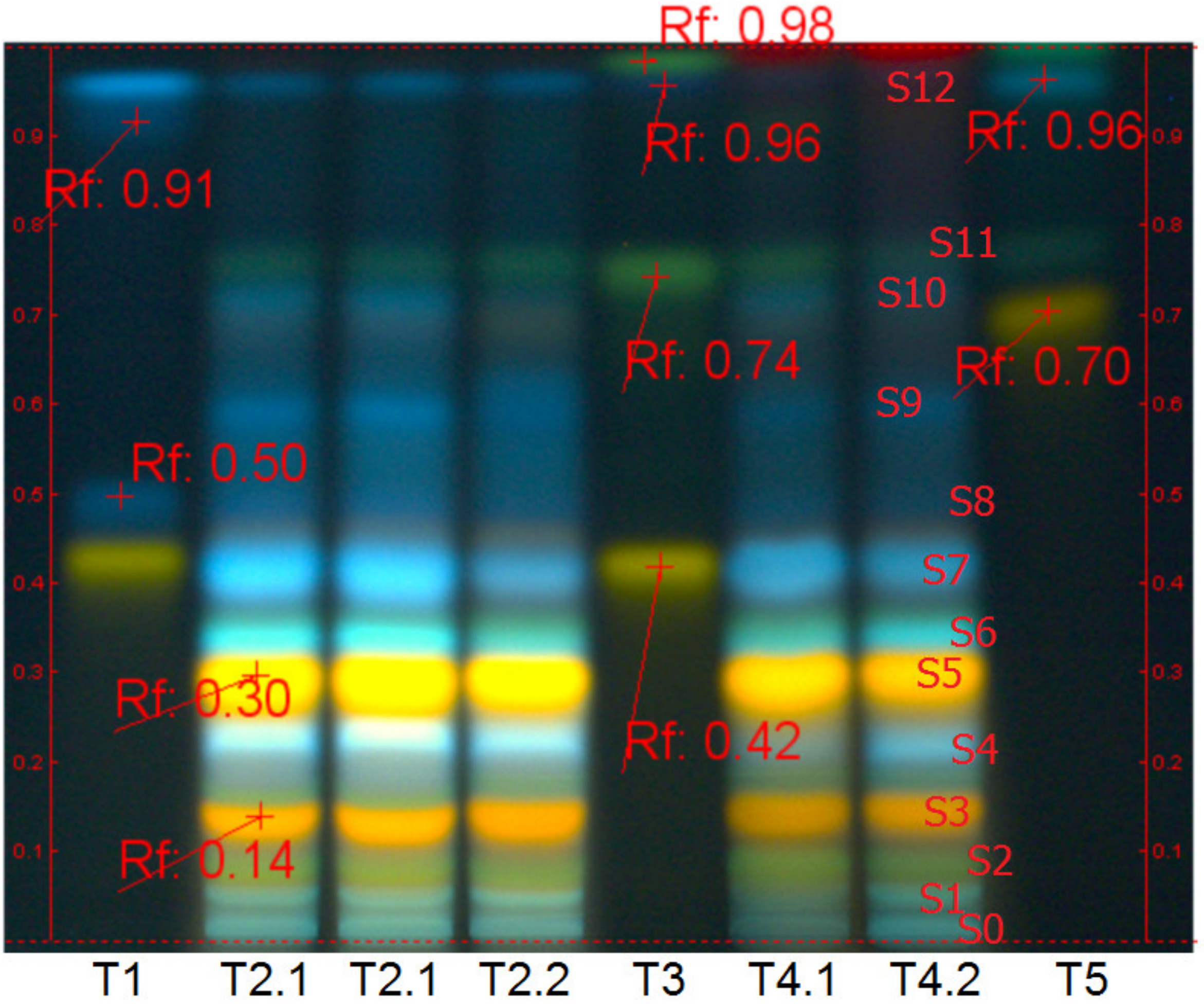
2.3. Plant Extraction and Their Chemical Characterization
3. Discussion



4. Materials and Methods
4.1. In Silico Assay
4.2. In Vitro Pharmacological Studies
4.3. Plant Extract Preparation
4.4. Plant Extracts Chemical Characterization
4.4.1. Chemical Qualitative Characterization of Plant Extracts
4.4.2. Chemical Quantitative Characterization of Plant Extracts
4.4.3. Chemicals, Reagents and Reference Compounds Description
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Park, J.R.; Eggert, A.; Caron, H. Neuroblastoma: Biology, prognosis, and treatment. Hematol. Oncol. Clin. North. Am. 2010, 24, 65–86. [Google Scholar] [CrossRef] [PubMed]
- Colon, N.C.; Chung, D.H. Neuroblastoma. Adv. Pediatr. 2011, 58, 297–311. [Google Scholar] [CrossRef] [PubMed]
- Maris, J.M. The biologic basis for neuroblastoma heterogeneity and risk stratification. Curr. Opin. Pediatr. 2005, 17, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Brodeur, G.M. Spontaneous regression of neuroblastoma. Cell Tissue Res. 2018, 372, 277–286. [Google Scholar] [CrossRef]
- Durazzo, A.; Lucarini, M.; Camilli, E.; Marconi, S.; Gabrielli, P.; Lisciani, S.; Gambelli, L.; Aguzzi, A.; Novellino, E.; Santini, A.; et al. Dietary Lignans: Definition, Description and Research Trends in Databases Development. Molecules 2018, 23, 3251. [Google Scholar] [CrossRef]
- Qiao, Y.; Sunada, N.K.; Hatada, A.E.; Lange, I.; Khutsishvili, M.; Alizade, V.; Atha, D.; Ko’omoa-Lange, D.-L.; Borris, R.P. Potential anti-neuroblastoma agents from Juniperus oblonga. Biochem. Biophys. Res. Commun. 2019, 516, 733–738. [Google Scholar] [CrossRef]
- Rao, J.M.; Srinivas, P.V.; Yadav, J.S.; Raghavan, K.V.; Saxena, A.K.; Shanmugavel, M.; Kampasi, H.; Qazi, G.N. A Novel Herbal Chemical Composition for the Treatment of Cancer. European Patent Office Publ. of Application with Search Report EP20010274887. Available online: https://patents.google.com/patent/CA2469093A1/en (accessed on 10 December 2022).
- López-Yerena, A.; Grases-Pintó, B.; Zhan-Dai, S.; Pérez-Cano, F.J.; Lamuela-Raventos, R.M.; Rodríguez-Lagunas, M.J.; Vallverdú-Queralt, A. Nutrition during pregnancy and lactation: New evidence for the vertical transmission of extra virgin olive oil phenolic compounds in rats. Food Chem. 2022, 391, 133211. [Google Scholar] [CrossRef]
- Shi, L.; Potts, M.; Kennelly, P.J. The serine, threonine, and/or tyrosine-specific protein kinases and protein phosphatases of prokaryotic organisms: A family portrait. FEMS Microbiol. Rev. 1998, 22, 229–253. [Google Scholar] [CrossRef]
- Bailey, J.J.; Schirrmacher, R.; Farrell, K.; Bernard-Gauthier, V. Tropomyosin receptor kinase inhibitors: An updated patent review for 2010–2016—Part II. Expert Opin. Ther. Pat. 2017, 27, 831–849. [Google Scholar] [CrossRef]
- Hay, N.; Sonenberg, N. Upstream and downstream of mTOR. Genes Dev. 2004, 18, 1926–1945. [Google Scholar] [CrossRef]
- King, D.; Yeomanson, D.; Bryant, H.E. PI3King the Lock: Targeting the PI3K/Akt/mTOR Pathway as a Novel Therapeutic Strategy in Neuroblastoma. J. Pediatr. Hematol. Oncol. 2015, 37, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Louis, C.U.; Shohet, J.M. Neuroblastoma: Molecular pathogenesis and therapy. Annu. Rev. Med. 2015, 66, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dube, C.; Gibert, M., Jr.; Cruickshanks, N.; Wang, B.; Coughlan, M.; Yang, Y.; Setiady, I.; Deveau, C.; Saoud, K.; et al. The p53 Pathway in Glioblastoma. Cancers 2018, 10, 297. [Google Scholar] [CrossRef] [PubMed]
- Van Maerken, T.; Rihani, A.; Dreidax, D.; De Clercq, S.; Yigit, N.; Marine, J.-C.; Westermann, F.; De Paepe, A.; Vandesompele, J.; Speleman, F. Functional Analysis of the p53 Pathway in Neuroblastoma Cells Using the Small-Molecule MDM2 Antagonist Nutlin-3. Mol. Cancer Ther. 2011, 10, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Fekry, B.; Jeffries, K.A.; Esmaeilniakooshkghazi, A.; Szulc, Z.M.; Knagge, K.J.; Kirchner, D.R.; Horita, D.A.; Krupenko, S.A.; Krupenko, N.I. C16-ceramide is a natural regulatory ligand of p53 in cellular stress response. Nat. Commun. 2018, 9, 4149. [Google Scholar] [CrossRef] [PubMed]
- Shangary, S.; Wang, S. Small-molecule inhibitors of the MDM2-p53 protein-protein interaction to reactivate p53 function: A novel approach for cancer therapy. Annu. Rev. Pharmacol. Toxicol. 2009, 49, 223–241. [Google Scholar] [CrossRef]
- Hannak, E.; Kirkham, M.; Hyman, A.A.; Oegema, K. Aurora-A kinase is required for centrosome maturation in Caenorhabditis elegans. J. Cell Biol. 2001, 155, 1109–1116. [Google Scholar] [CrossRef]
- Crane, R.; Gadea, B.; Littlepage, L.; Wu, H.; Ruderman, J.V. Aurora A, meiosis and mitosis. Biol. Cell. 2004, 96, 215–229. [Google Scholar] [CrossRef]
- Romain, C.; Paul, P.; Kim, K.W.; Lee, S.; Qiao, J.; Chung, D.H. Targeting Aurora kinase-A downregulates cell proliferation and angiogenesis in neuroblastoma. J. Pediatr. Surg. 2014, 49, 159–165. [Google Scholar] [CrossRef]
- Choudhury, S.R.; Karmakar, S.; Banik, N.L.; Ray, S.K. Targeting angiogenesis for controlling neuroblastoma. J. Oncol. 2012, 2012, 782020. [Google Scholar] [CrossRef]
- Meister, B.; Grünebach, F.; Bautz, F.; Brugger, W.; Fink, F.M.; Kanz, L.; Möhle, R. Expression of vascular endothelial growth factor (VEGF) and its receptors in human neuroblastoma. Eur. J. Cancer 1999, 35, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Slack, A.; Chen, Z.; Tonelli, R.; Pule, M.; Hunt, L.; Pession, A.; Shohet, J.M. The p53 regulatory gene MDM2 is a direct transcriptional target of MYCN in neuroblastoma. Proc. Natl. Acad. Sci. USA 2005, 102, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Zafar, A.; Wang, W.; Liu, G.; Xian, W.; McKeon, F.; Zhou, J.; Zhang, R. Targeting the p53-MDM2 pathway for neuroblastoma therapy: Rays of hope. Cancer Lett. 2021, 496, 16–29. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Rodriguez-Silva, M.; Acanda de la Rocha, A.M.; Wolf, A.L.; Lai, Y.; Liu, Y.; Reinhold, W.C.; Pommier, Y.; Chambers, J.W.; Tse-Dinh, Y.-C. Tyrosyl-DNA Phosphodiesterase 1 and Topoisomerase I Activities as Predictive Indicators for Glioblastoma Susceptibility to Genotoxic Agents. Cancers 2019, 11, 1416. [Google Scholar] [CrossRef]
- Swanepoel, B.; Venables, L.; Olaru, O.T.; Nitulescu, G.M.; van de Venter, M. In Vitro Anti-proliferative Activity and Mechanism of Action of Anemone nemorosa. Int. J. Mol. Sci. 2019, 20, 1217. [Google Scholar] [CrossRef] [PubMed]
- Jia, D.; Han, B.; Yang, S.; Zhao, J. Anemonin alleviates nerve injury after cerebral ischemia and reperfusion (i/r) in rats by improving antioxidant activities and inhibiting apoptosis pathway. J. Mol. Neurosci. 2014, 53, 271–279. [Google Scholar] [CrossRef]
- Lukianchuk, A.; Khropot, O.; Konechnyi, Y.; Konechna, R.; Novikov, V. Wood anemone. Anemone nemorosa L. Analytical review. Sci. Rise Pharm. Sci. 2017, 3, 34–38. [Google Scholar]
- Hao, D.-C.; Gu, X.; Xiao, P. Anemone medicinal plants: Ethnopharmacology, phytochemistry and biology. Acta Pharm. Sin. B 2017, 7, 146–158. [Google Scholar] [CrossRef]
- Banerjee, S.; Ji, C.; Mayfield, J.E.; Goel, A.; Xiao, J.; Dixon, J.E.; Guo, X. Ancient drug curcumin impedes 26S proteasome activity by direct inhibition of dual-specificity tyrosine-regulated kinase 2. Proc. Natl. Acad. Sci. USA 2018, 115, 8155–8160. [Google Scholar] [CrossRef]
- Limban, C.; Nuta, D.C.; Missir, A.V.; Roman, R.; Caproiu, M.T.; Dumitrascu, F.; Pintilie, L.; Stefaniu, A.; Chifiriuc, M.C.; Popa, M.; et al. Synthesis and Characterization of New Fluoro/Trifluoromethyl-Substituted Acylthiourea Derivatives with Promising Activity against Planktonic and Biofilm-Embedded Microbial Cells. Processes 2020, 8, 503. [Google Scholar] [CrossRef]
- Schroder, V.; Radu, N.; Cornea, P.C.; Coman, O.A.; Pirvu, L.C.; Mohammed, M.S.O.; Stefaniu, A.; Pintilie, L.; Bostan, M.; Caramihai, M.D.; et al. Studies Regarding the Antimicrobial Behavior of Clotrimazole and Limonene. Antibiotics 2022, 11, 1816. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://pubchem.ncbi.nlm.nih.gov (accessed on 30 January 2023).
- Halgren, T.A. Merck molecular force field. I. Basis, form, scope, parameterization, and performance of MMFF94. J. Comput. Chem. 1996, 17, 490–519. [Google Scholar] [CrossRef]
- Ghose, A.K.; Crippen, G.M. Atomic physicochemical parameters for three-dimensional-structure-directed quantitative structure-activity relationships. 2. Modeling dispersive and hydrophobic interactions. J. Chem. Inf. Comput. Sci. 1987, 27, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Leo, A.; Hansch, C.; Elkins, D. Partition Coefficients and their uses. Chem. Rev. 1971, 6, 525–616. [Google Scholar] [CrossRef]
- Proudfoot, J.R. Drugs, leads, and drug-likeness: An analysis of some recently launched drugs. Bioorg. Med. Chem. Lett. 2002, 12, 1647–1650. [Google Scholar] [CrossRef]
- Protocols & Applications Guide. Available online: www.promega.com (accessed on 8 August 2022).
- Zink, A.; Conrad, J.; Telugu, N.S.; Diecke, S.; Heinz, A.; Wanker, E.; Priller, J.; Prigione, A. Assessment of Ethanol-Induced Toxicity on iPSC-Derived Human Neurons Using a Novel High-Throughput Mitochondrial Neuronal Health (MNH) Assay. Front. Cell. Dev. Biol. 2020, 8, 590540. [Google Scholar] [CrossRef]
- Pirvu, L.; Stefaniu, A.; Neagu, G.; Pintilie, L. Studies on Anemone nemorosa L. extracts; polyphenols profile, antioxidant activity, and effects on Caco-2 cells by in vitro and in silico studies. Open Chem. 2022, 20, 299–312. [Google Scholar] [CrossRef]
- Wagner, H.; Bladt, S. Plant Drug Analysis. In A Thin Layer Chromatography Atlas, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 1996. [Google Scholar]
- Voges, D.; Zwickl, P.; Baumeister, W. The 26S proteasome: A molecular machine designed for controlled proteolysis. Annu. Rev. Biochem. 1999, 68, 1015–1068. [Google Scholar] [CrossRef]
- Adams, J. The proteasome: Structure, function, and role in the cell. Cancer Treat. Rev. 2003, 29 (Suppl. 1), 3–9. [Google Scholar] [CrossRef]
- Belizario, J.E.; Alves, J.; Garay-Malpartida, M.; Occhiucci, J.M. Coupling caspase cleavage and proteasomal degradation of proteins carrying PEST motif. Curr. Protein Pept. Sci. 2008, 9, 210–220. [Google Scholar] [CrossRef]
- Available online: https://www.medchemexpress.com/Bortezomib.html (accessed on 2 March 2023).
- Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Bortezomib (accessed on 2 March 2023).
- Łuczkowska, K.; Sokolowska, K.E.; Taryma-Lesniak, O.; Pastuszak, K.; Supernat, A.; Bybjerg-Grauholm, J.; Hansen, L.L.; Paczkowska, E.; Wojdacz, T.K.; Machaliński, B. Bortezomib induces methylation changes in neuroblastoma cells that appear to play a significant role in resistance development to this compound. Sci. Rep. 2021, 11, 9846. [Google Scholar] [CrossRef] [PubMed]
- Tandon, V.; de la Vega, L.; Banerjee, S. Emerging roles of DYRK2 in cancer. J. Biol. Chem. 2021, 296, 100233. [Google Scholar] [CrossRef] [PubMed]
- Correa-Sáez, A.; Jiménez-Izquierdo, R.; Garrido-Rodríguez, M.; Morrugares, R.; Muñoz, E.; Calzado, M.A. Updating dual-specificity tyrosine-phosphorylation-regulated kinase 2 (DYRK2): Molecular basis, functions and role in diseases. Cell. Mol. Life Sci. 2020, 77, 4747–4763. [Google Scholar] [CrossRef] [PubMed]
- Available online: http://www.chemspider.com/Chemical-Structure.4444445.html (accessed on 2 March 2023).
- Available online: http://www.chemspider.com/Chemical-Structure.839564.html (accessed on 2 March 2023).
- Berrougui, H.; Herrera-González, M.; Marhuenda, E.; Ettaib, A.; Hmamouch, M. Relaxant activity of methanolic extract from seeds of Peganum harmala on isolated rat aorta. Therapie 2022, 57, 236–241. [Google Scholar]
- Oodi, A.; Norouzi, H.; Amirizadeh, N.; Nikougoftar, M.; Vafaie, Z. Harmine, a Novel DNA Methyltransferase 1 Inhibitor in the Leukemia Cell Line. Indian J. Hematol. Blood Transfus. 2017, 33, 509–515. [Google Scholar] [CrossRef]
- Fuloria, S.; Mehta, J.; Chandel, A.; Sekar, M.; Rani, N.N.I.M.; Begum, M.Y.; Subramaniyan, V.; Chidambaram, K.; Thangavelu, L.; Nordin, R.; et al. A Comprehensive Review on the Therapeutic Potential of Curcuma longa Linn. in Relation to its Major Active Constituent Curcumin. Front. Pharmacol. 2022, 13, 820806. [Google Scholar] [CrossRef]
- Zhai, K.; Brockmüller, A.; Kubatka, P.; Shakibaei, M.; Büsselberg, D. Curcumin’s Beneficial Effects on Neuroblastoma: Mechanisms, Challenges, and Potential Solutions. Biomolecules 2020, 10, 1469. [Google Scholar] [CrossRef]
- Chen, J.; Marechal, V.; Levine, A.J. Mapping of the p53 and mdm-2 interaction domains. Mol. Cell Biol. 1993, 13, 4107–4114. [Google Scholar]
- Picksley, S.M.; Vojtesek, B.; Sparks, A.; Lane, D.P. Immunochemical analysis of the interaction of p53 with MDM2–fine mapping of the MDM2 binding site on p53 using synthetic peptides. Oncogene 1994, 9, 2523–2529. [Google Scholar]
- Stoll, R.; Renner, C.; Hansen, S.; Palme, S.; Klein, C.; Belling, A.; Holak, T.A. Chalcone derivatives antagonize interactions between the human oncoprotein MDM2 and p53. Biochemistry 2001, 40, 336–344. [Google Scholar] [CrossRef]
- Constantinescu, C.; Lungu, C.N. Anticancer Activity of Natural and Synthetic Chalcones. Int. J Mol. Sci. 2021, 2, 11306. [Google Scholar] [CrossRef] [PubMed]
- MDM2-p53 Interaction Inhibitors Library. Available online: https://www.chemdiv.com/ (accessed on 1 March 2023).
- Haupt, S.; Gamell, C.; Wolyniec, K.; Haupt, Y. Interplay between p53 and VEGF: How to prevent the guardian from becoming a villain. Cell Death Differ. 2013, 20, 852–854. [Google Scholar] [CrossRef]
- Ghahremani, M.F.; Gossens, S.; Nittner, D.; Bisteau, X.; Bartunkova, S.; Zwolinska, A.; Hulpiau, P.; Haigh, K.; Haenebalcke, L.; Drogat, B.; et al. p53 promotes VEGF expression and angiogenesis in the absence of an intact p21-Rb pathway. Cell Death Differ. 2013, 20, 888–897. [Google Scholar] [CrossRef] [PubMed]
- Vassal, G.; Pondarré, C.; Cappelli, C.; Terrier-Lacombe, M.J.; Boland, I.; Morizet, J.; Bénard, J.; Vénuat, A.M.; Ardouin, P.; Hartmann, O.; et al. DNA-topoisomerase I, a new target for the treatment of neuroblastoma. Eur. J. Cancer. 1997, 33, 2011–2015. [Google Scholar] [CrossRef] [PubMed]
- Vassal, G.; Terrier-Lacombe, M.J.; Bissery, M.C.; Vénuat, A.M.; Gyergyay, F.; Bénard, J.; Morizet, J.; Boland, I.; Ardouin, P.; Bressac-de-Paillerets, B.; et al. Therapeutic activity of CPT-11, a DNA-topoisomerase I inhibitor, against peripheral primitive neuroectodermal tumour and neuroblastoma xenografts. Br. J. Cancer 1996, 74, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Vassal, G.; Pondarré, C.; Boland, I.; Cappelli, C.; Santos, A.; Thomas, C.; Lucchi, E.; Imadalou, K.; Pein, F.; Morizet, J.; et al. Preclinical development of camptothecin derivatives and clinical trials in pediatric oncology. Biochimie 1998, 80, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Wang, J.; Sun, F.; Zhen, Z.; Chen, T.; Lu, S.; Huang, J.; Zhang, Y.; Sun, X. Vincristine, Irinotecan, and Temozolomide in Patientswithh Relapsed/Refractory Neuroblastoma. Front. Oncol. 2022, 12, 804310. [Google Scholar] [CrossRef]
- Kanzawa, F.; Koizumi, F.; Koh, Y.; Nakamura, T.; Tatsumi, Y.; Fukumoto, H.; Saijo, N.; Yoshioka, T.; Nishio, K. In Vitro Synergistic Interactions between the Cisplatin Analogue Nedaplatin and the DNA Topoisomerase I Inhibitor Irinotecan and the Mechanism of this Interaction. Clin. Cancer Res. 2001, 7, 202–209. [Google Scholar]
- Castañeda, A.M.; Meléndez, C.M.; Uribe, D.; Pedroza-Díaz, J. Synergistic effects of natural compounds and conventional chemotherapeutic agents: Recent insights for the development of cancer treatment strategies. Heliyon 2022, 8, e09519. [Google Scholar] [CrossRef]
- Limaye, A.; Sweta, J.; Madhavi, M.; Mudgal, U.; Mukherjee, S.; Sharma, S.; Hussain, T.; Nayarisseri, A.; Singh, S.K. In Silico Insights on GD2: A Potential Target for Pediatric Neuroblastoma. Curr. Top Med. Chem. 2019, 19, 2766–2781. [Google Scholar] [CrossRef]
- Sumera; Anwer, F.; Waseem, M.; Fatima, A.; Malik, N.; Ali, A.; Zahid, S. Molecular Docking and Molecular Dynamics Studies Reveal Secretory Proteins as Novel Targets of Temozolomide in Glioblastoma Multiforme. Molecules 2022, 27, 7198. [Google Scholar] [CrossRef] [PubMed]
- Arjmand, B.; Hamidpour, S.K.; Alavi-Moghadam, S.; Yavari, H.; Shahbazbadr, A.; Tavirani, M.R.; Gilany, K.; Larijani, B. Molecular Docking as a Therapeutic Approach for Targeting Cancer Stem Cell Metabolic Processes. Front. Pharmacol. 2022, 13, 768556. [Google Scholar] [CrossRef]
- Cardinali, A.; Pati, S.; Minervini, F.; D’Antuono, I.; Linsalata, V.; Lattanzio, V. Verbascoside, isoverbascoside, and their derivatives recovered from olive mill wastewater as possible food antioxidants. J. Agric. Food Chem. 2012, 60, 1822–1829. [Google Scholar] [CrossRef] [PubMed]
- Alipieva, K.; Korkina, L.; Orhan, I.E.; Georgiev, M.I. Verbascoside—A review of its occurrence, (bio)synthesis and pharmacological significance. Biotechnol. Adv. 2014, 32, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- During, A.; Debouche, C.; Raas, T.; Larondelle, Y. Among plant lignans, pinoresinol has the strongest anti-inflammatory properties in human intestinal Caco-2 cells. J. Nutr. 2012, 142, 1798–1805. [Google Scholar] [CrossRef]
- Shao, Y.; Molnar, L.F.; Jung, Y.; Kussmann, J.; Ochsenfeld, C.; Brown, S.T.; Gilbert, A.T.B.; Slipchenko, L.V.; Levchenko, S.V.; O’Neill, D.P.; et al. Advances in methods and algorithms in a modern quantum chemistry program package. Phys. Chem. Chem. Phys. 2006, 8, 3172–3191. [Google Scholar] [CrossRef]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Compound | Chemical Structure * | Docking Score CLC | MolDock Score MVD | Ranking Score CLC | Ranking Score MVD |
|---|---|---|---|---|---|
| Verbascoside |  | −88.14 | −148.78 | 1 | 1 |
| Pinoresinol |  | −79.04 | −126.85 | 2 | 3 |
| Co-crystallized CUR A501, CURCUMIN (5ZTN native ligand) |  | −83.92 | −115.85 | 3 | 6 |
| Secoisolariciresinol |  | −77.14 | −112.96 | 4 | 8 |
| Lariciresinol |  | −76.57 | −127.28 | 5 | 2 |
| Ligstroside aglycone |  | −76.01 | −102.50 | 6 | 11 |
| Oleuropein aglycone ** |  | −72.40 | −88.49 | 7 | 16 |
| Oleacein ** |  | −73.23 | −88.52 | 8 | 15 |
| Medioresinol |  | −72.31 | −124.68 | 9 | 4 |
| Matairesinol |  | −72.12 | −116.98 | 10 | 5 |
| Luteolin |  | −71.01 | −104.91 | 11 | 10 |
| Apigenin |  | −70.83 | −101.31 | 12 | 12 |
| Isolariciresinol ** |  | −69.22 | −114.37 | 13 | 7 |
| Syringaresinol |  | −67.68 | −108.62 | 14 | 9 |
| Tyrosol |  | −46.56 | −68.27 | 18 | 21 |
| Elenolic acid |  | −45.42 | −68.71 | 19 | 20 |
| Vanillic acid |  | −45.02 | −69.88 | 20 | 19 |
| o-Coumaric acid |  | −49.70 | −71.56 | 17 | 18 |
| Hydroxytyrosol |  | −50.26 | −73.62 | 16 | 17 |
| Oleocanthal |  | −66.17 | −93.41 | 15 | 14 |
| Anemonin |  | −34.27 | −93.75 | 21 | 13 |
| Test Compounds | No. of Atoms | Weight [Daltons] | Flexible Bonds | Lipinski’s Violations | Hydrogen Donors | Hydrogen Acceptors | logP * |
|---|---|---|---|---|---|---|---|
| Anemonin | 22 | 192.17 | 0 | 0 | 0 | 4 | 0.41 |
| Apigenin | 30 | 270.24 | 1 | 0 | 3 | 5 | 4.28 |
| o-Coumaric acid | 20 | 164.16 | 2 | 0 | 2 | 3 | 1.93 |
| CURCUMIN | 49 | 370.40 | 9 | 0 | 2 | 6 | 3.00 |
| Elenoic acid | 31 | 242.23 | 5 | 0 | 1 | 6 | 0.12 |
| Hydroxytyrosol | 21 | 154.16 | 2 | 0 | 3 | 3 | 0.79 |
| Isolariciresinol | 50 | 360.40 | 5 | 0 | 4 | 6 | 2.04 |
| Lariciresinol | 50 | 360.40 | 6 | 0 | 3 | 6 | 2.40 |
| Ligstroside-aglycone | 48 | 362.37 | 8 | 0 | 2 | 7 | 1.50 |
| Luteolin | 31 | 286.24 | 1 | 0 | 4 | 6 | 3.93 |
| Matairesinol | 48 | 358.39 | 6 | 0 | 2 | 6 | 3.25 |
| Medioresinol | 52 | 388.41 | 5 | 0 | 2 | 7 | 2.25 |
| Oleacein | 43 | 320.34 | 10 | 0 | 2 | 6 | 1.11 |
| Oleocanthal | 42 | 304.34 | 10 | 0 | 1 | 5 | 1.47 |
| Oleuropein aglycone | 49 | 378.37 | 8 | 0 | 3 | 8 | 1.14 |
| Pinoresinol | 48 | 358.39 | 4 | 0 | 2 | 6 | 2.28 |
| Secoisolariciresinol | 52 | 362.42 | 9 | 0 | 4 | 6 | 2.51 |
| Syringaresinol | 56 | 418.44 | 6 | 0 | 2 | 8 | 2.22 |
| Vanillic acid | 20 | 168.15 | 2 | 0 | 2 | 4 | 1.26 |
| Verbascoside | 80 | 624.59 | 11 | 3 | 9 | 15 | −0.51 |
| Tyrosol | 20 | 138.16 | 2 | 0 | 2 | 2 | 1.14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pirvu, L.C.; Neagu, G.; Albulescu, A.; Stefaniu, A.; Pintilie, L. Potential Benefits of Dietary Plant Compounds on Normal and Tumor Brain Cells in Humans: In Silico and In Vitro Approaches. Int. J. Mol. Sci. 2023, 24, 7404. https://doi.org/10.3390/ijms24087404
Pirvu LC, Neagu G, Albulescu A, Stefaniu A, Pintilie L. Potential Benefits of Dietary Plant Compounds on Normal and Tumor Brain Cells in Humans: In Silico and In Vitro Approaches. International Journal of Molecular Sciences. 2023; 24(8):7404. https://doi.org/10.3390/ijms24087404
Chicago/Turabian StylePirvu, Lucia Camelia, Georgeta Neagu, Adrian Albulescu, Amalia Stefaniu, and Lucia Pintilie. 2023. "Potential Benefits of Dietary Plant Compounds on Normal and Tumor Brain Cells in Humans: In Silico and In Vitro Approaches" International Journal of Molecular Sciences 24, no. 8: 7404. https://doi.org/10.3390/ijms24087404