
Morphological and Physiological Mechanisms of Melatonin on Delaying Drought-Induced Leaf Senescence in Cotton
 and
and
Abstract
:1. Introduction
2. Results
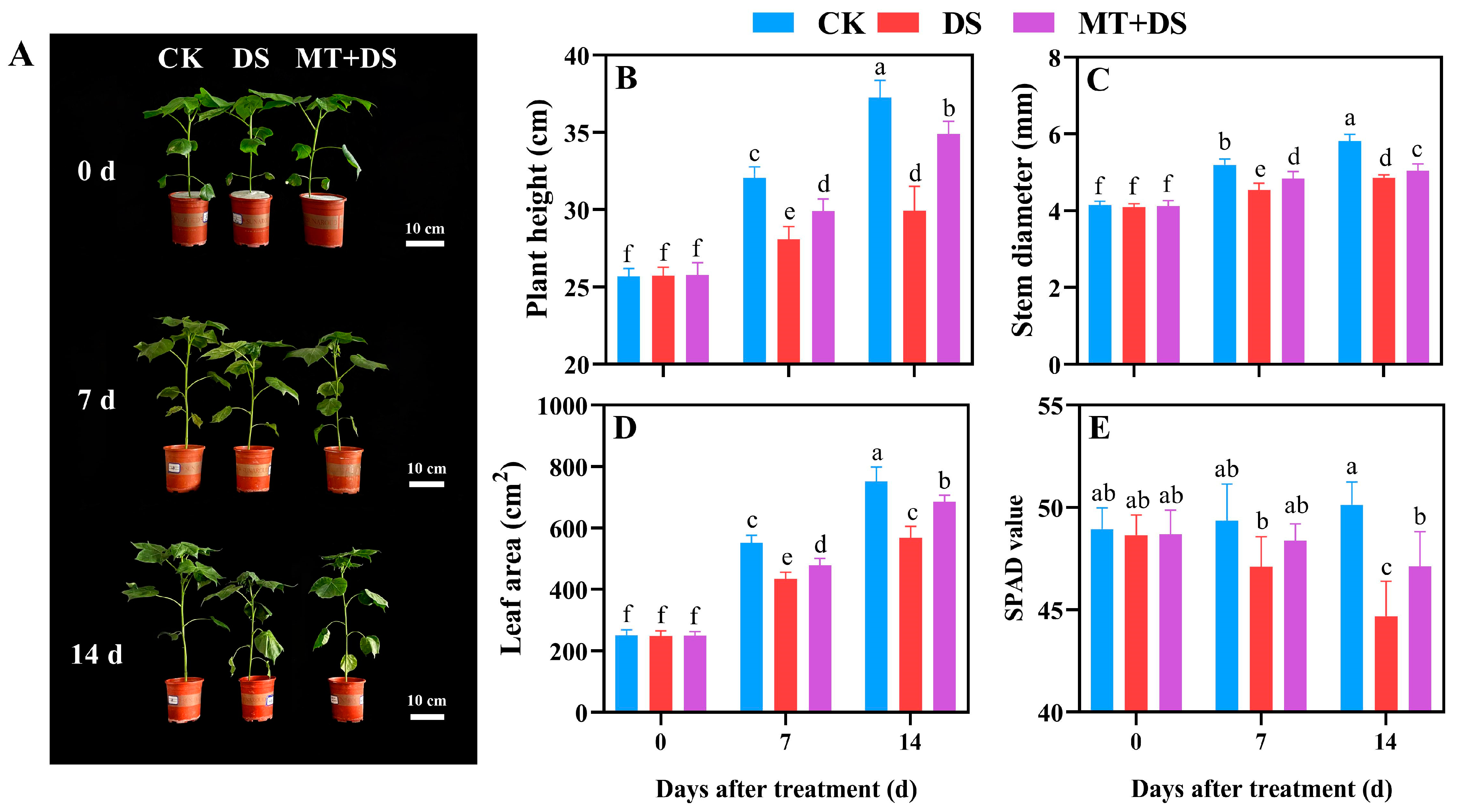
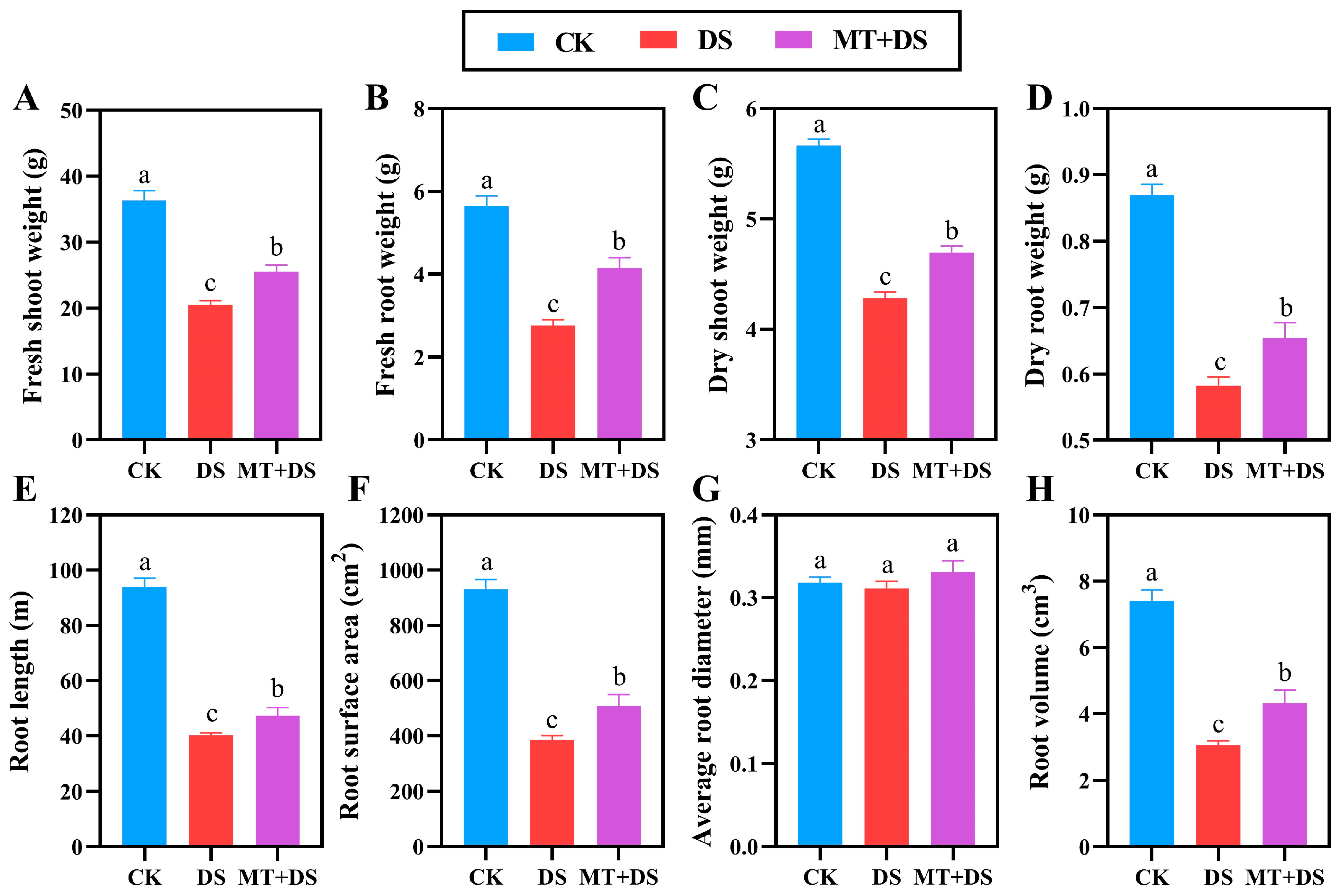
2.1. Effects of MT on the Growth and Development of Cotton Seedlings under DS
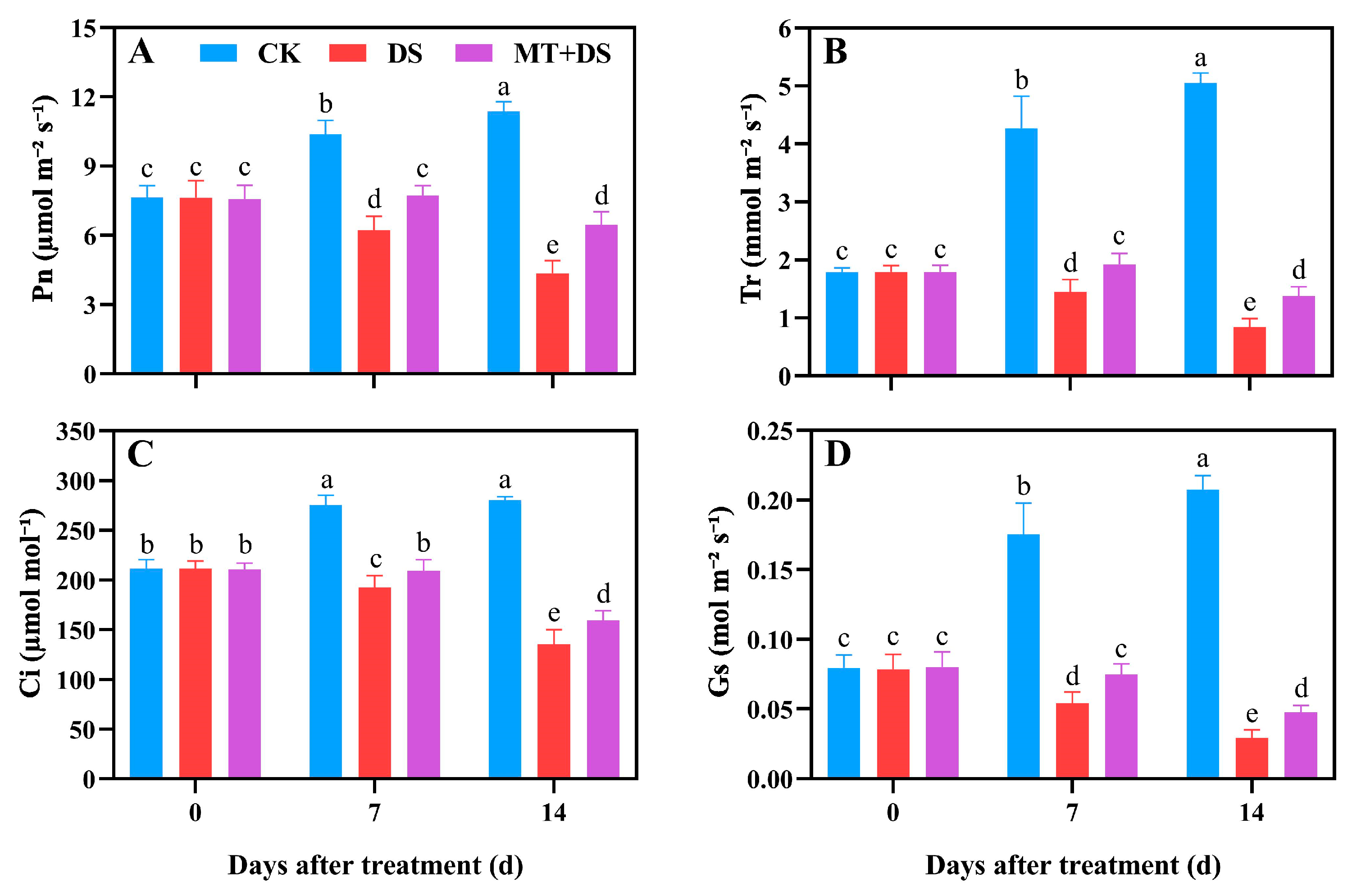
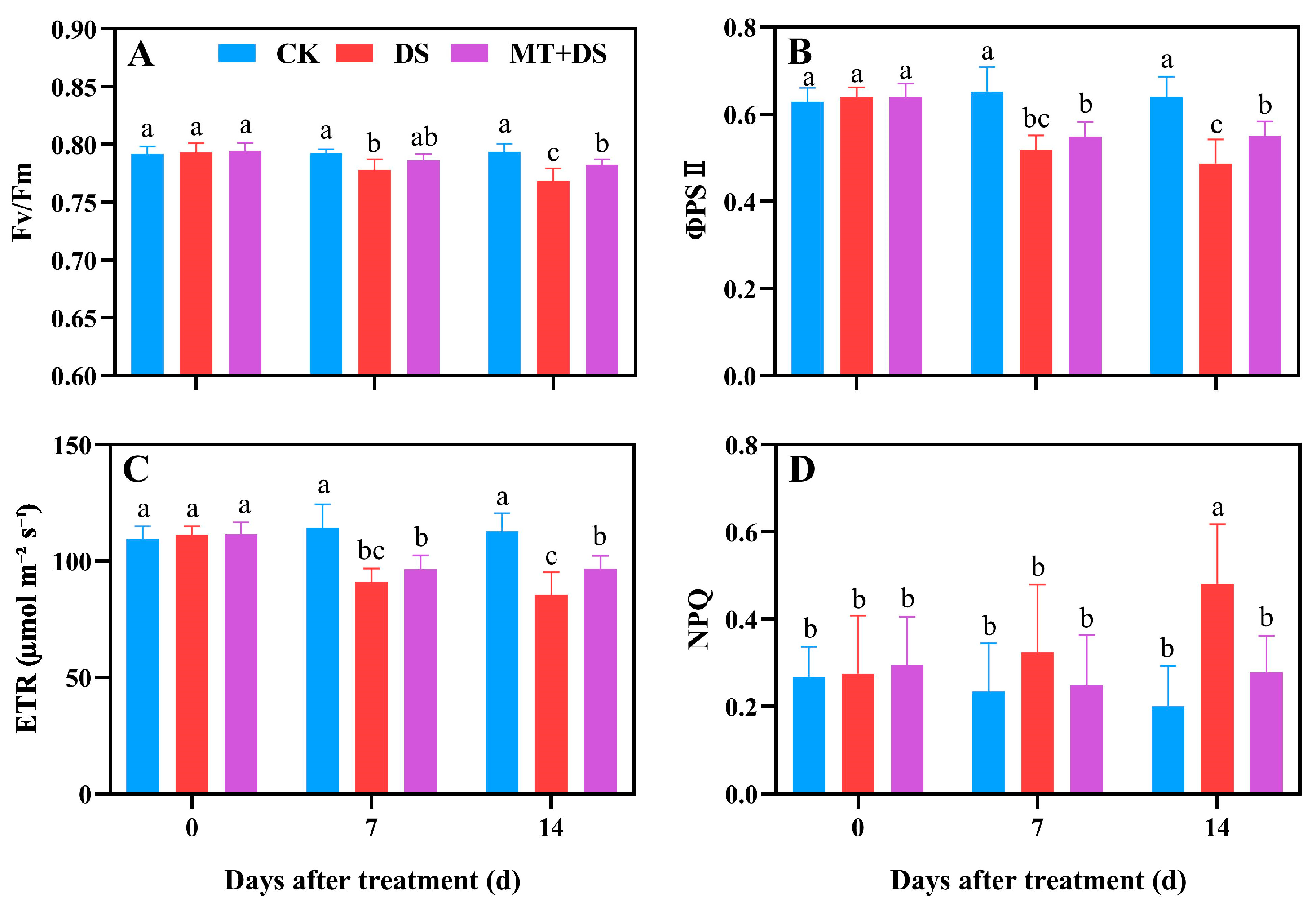
2.2. Effects of MT on the Photosynthetic Characteristics
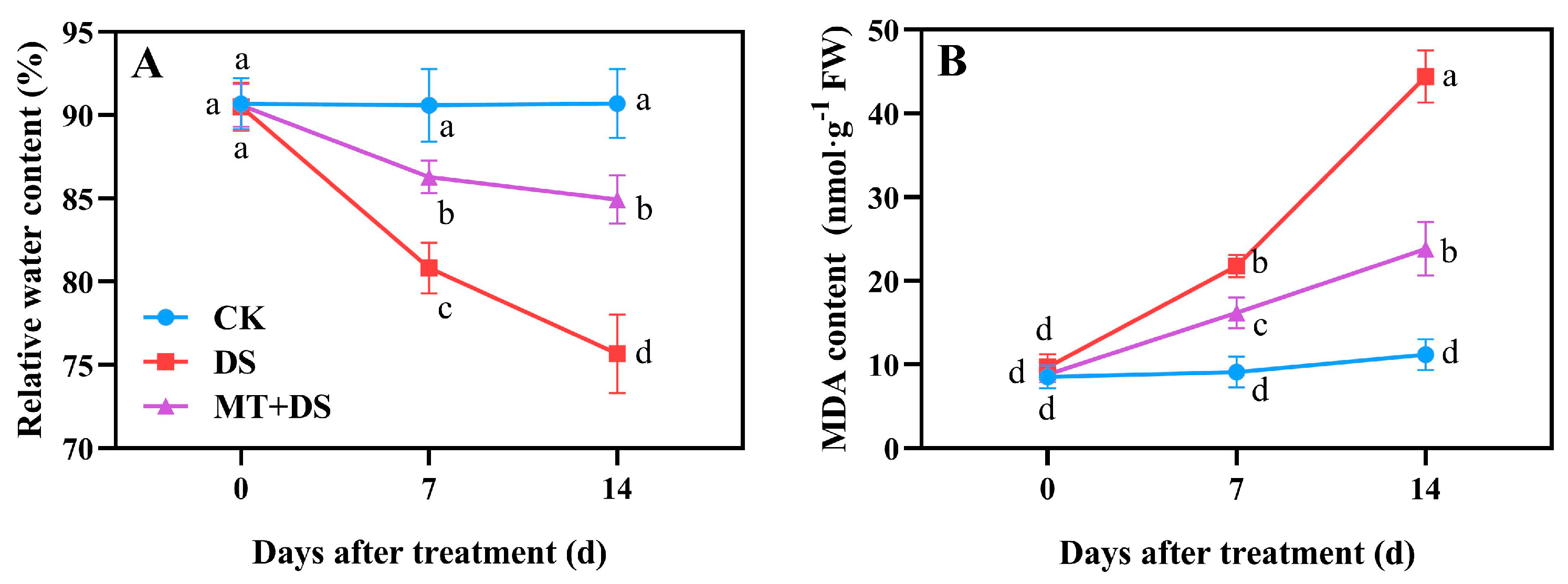
2.3. Effects of MT on Relative Water Content (RWC) and Malondialdehyde (MDA) Content
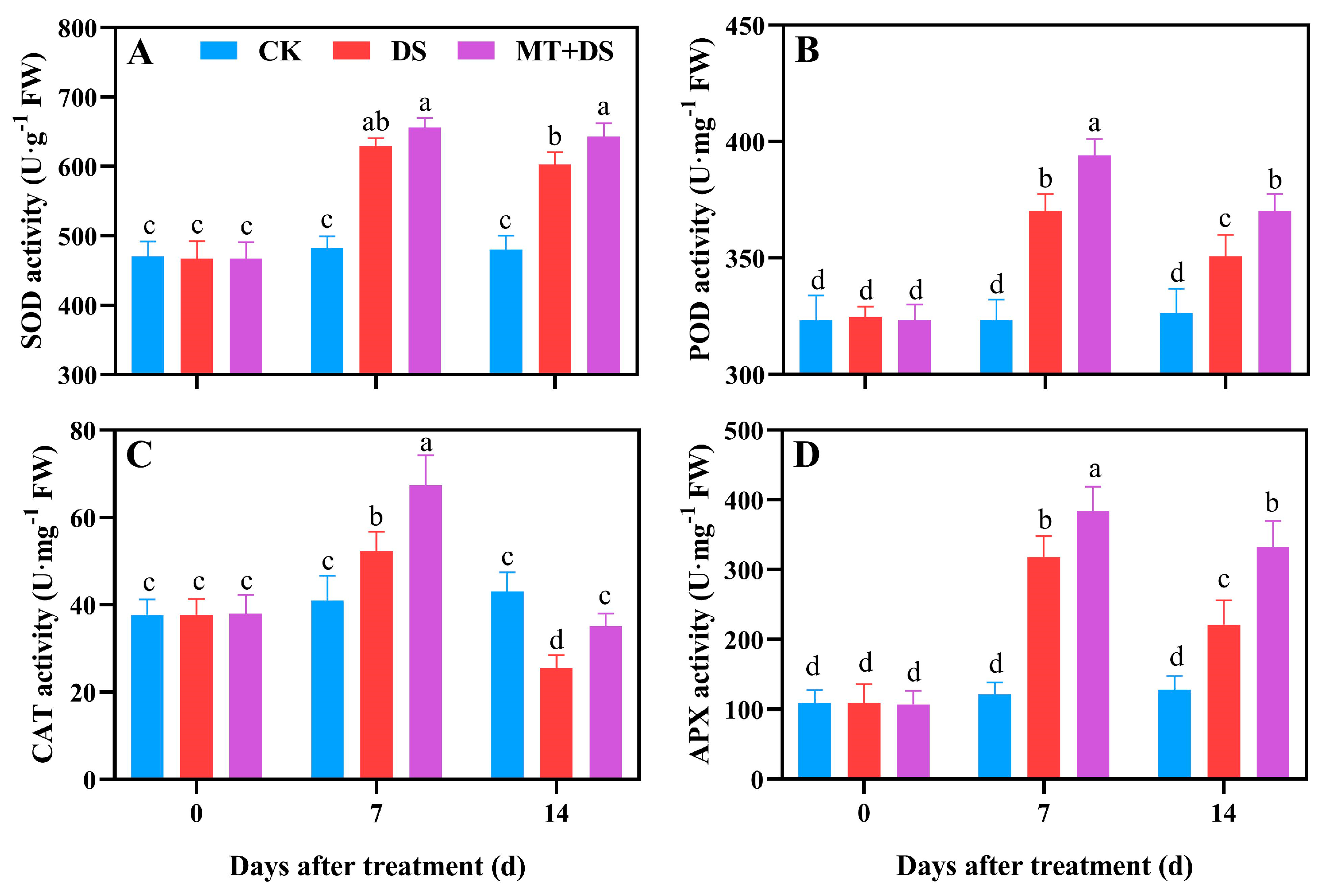
2.4. Effects of MT on the Antioxidant Enzyme Activity
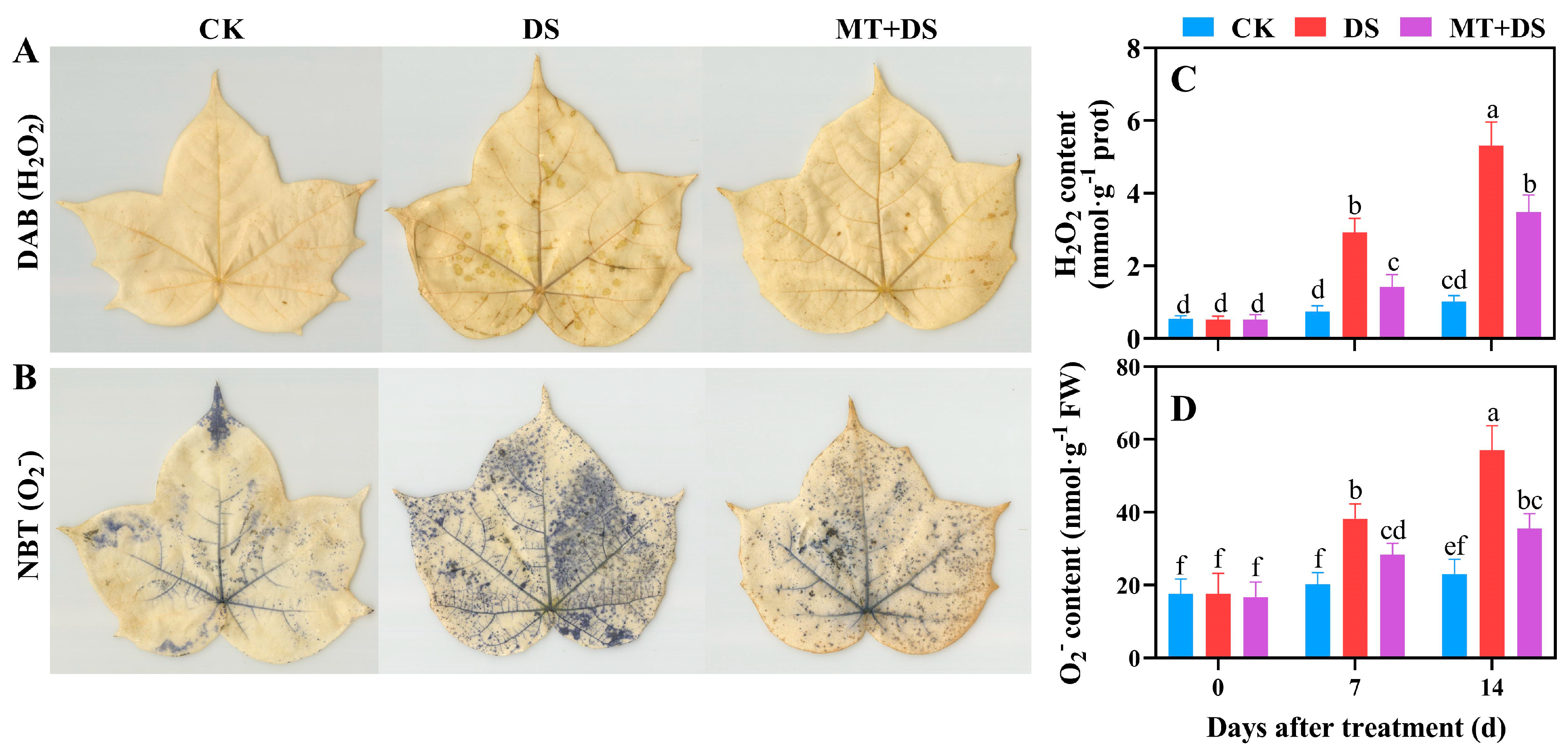
2.5. Effect of MT on the ROS Content
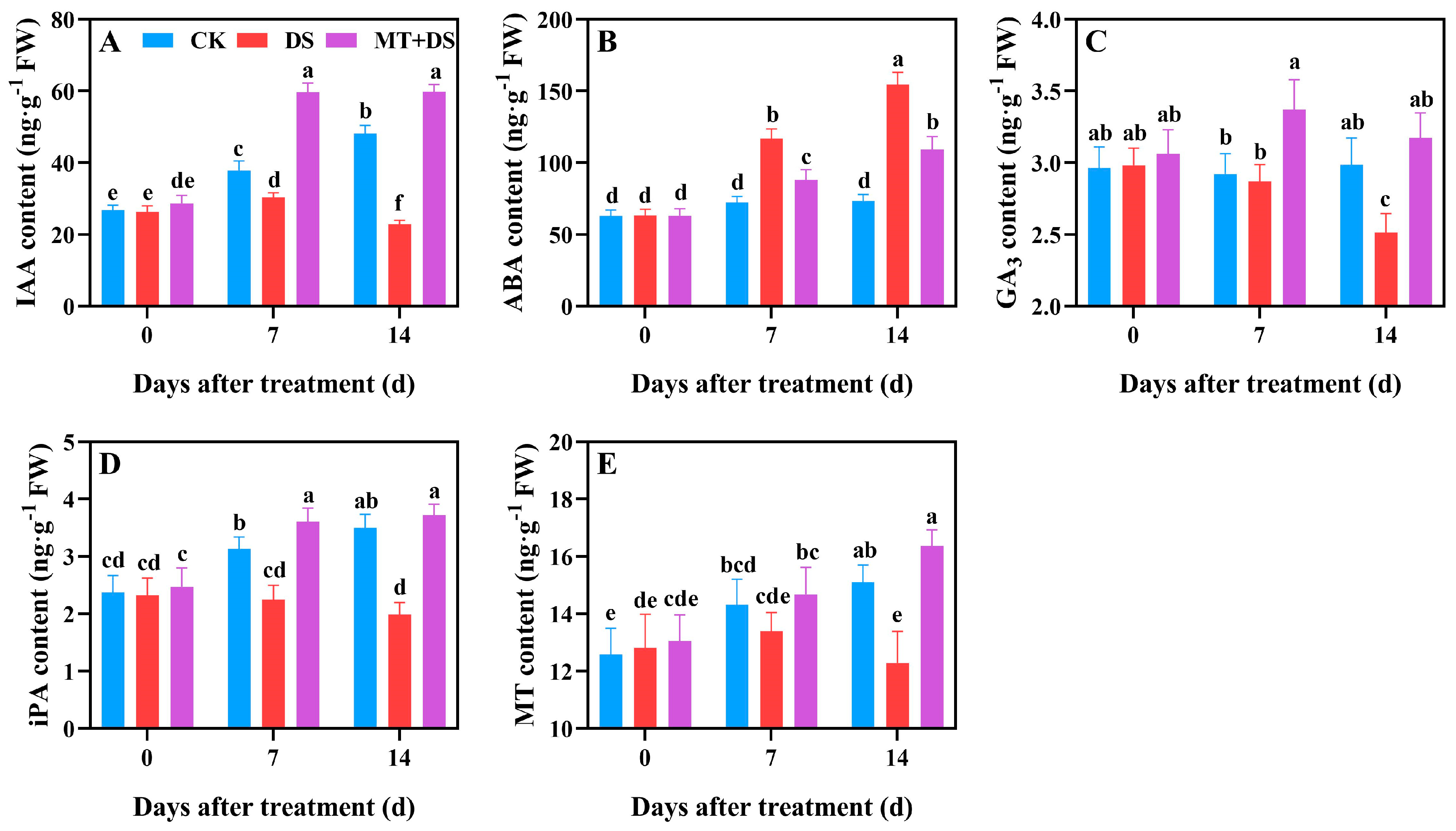
2.6. Effects of MT on the Endogenous Hormone Content
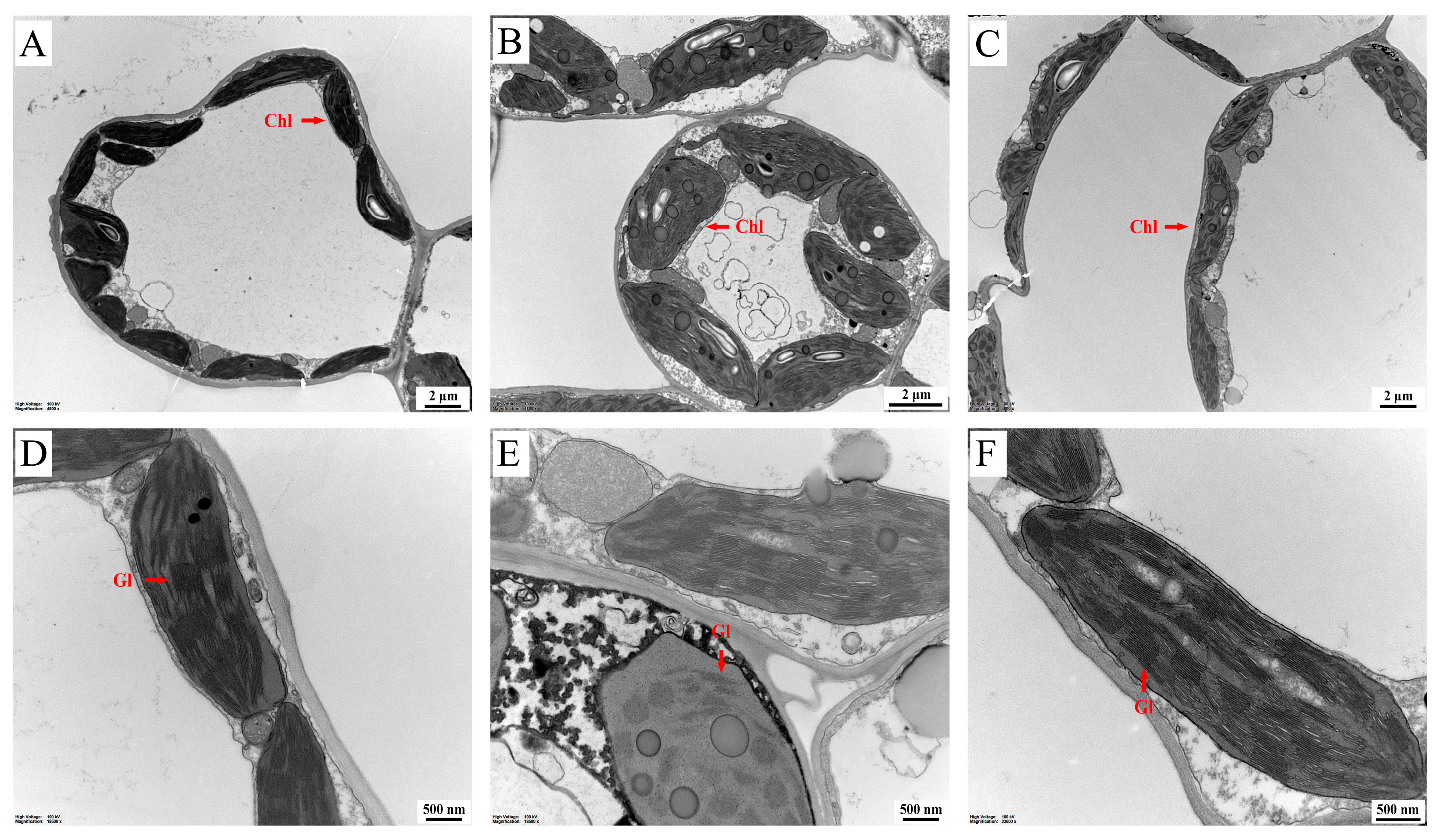
2.7. Effects of MT on the Ultrastructural Alterations in the Chloroplast
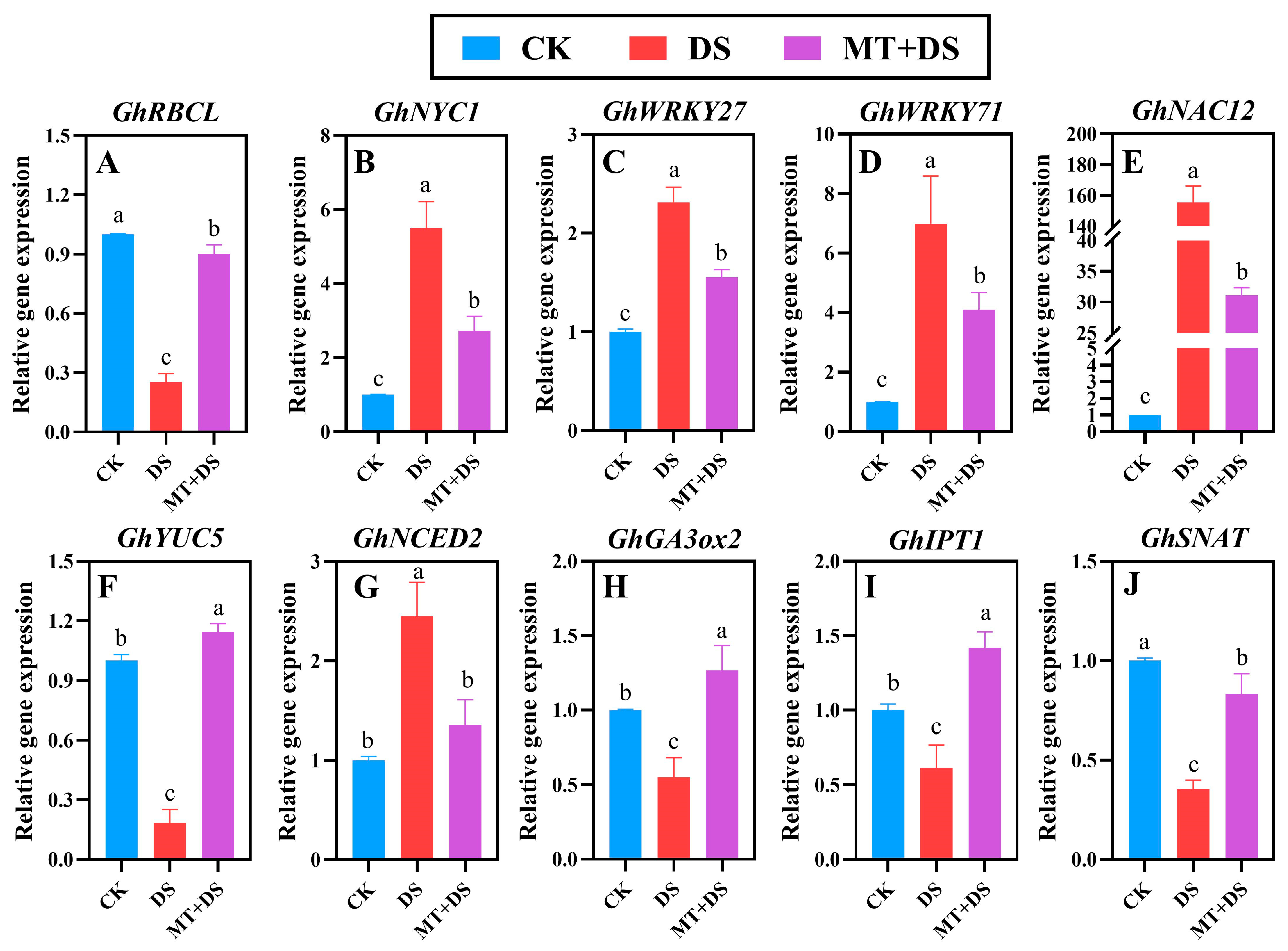
2.8. Effects of MT on the Transcript Abundance of Chlorophyll Degradation Genes, Senescence Marker Genes, and Hormone Synthesis Genes during Leaf Senescence
2.9. Correlation and Principal Component Analyses
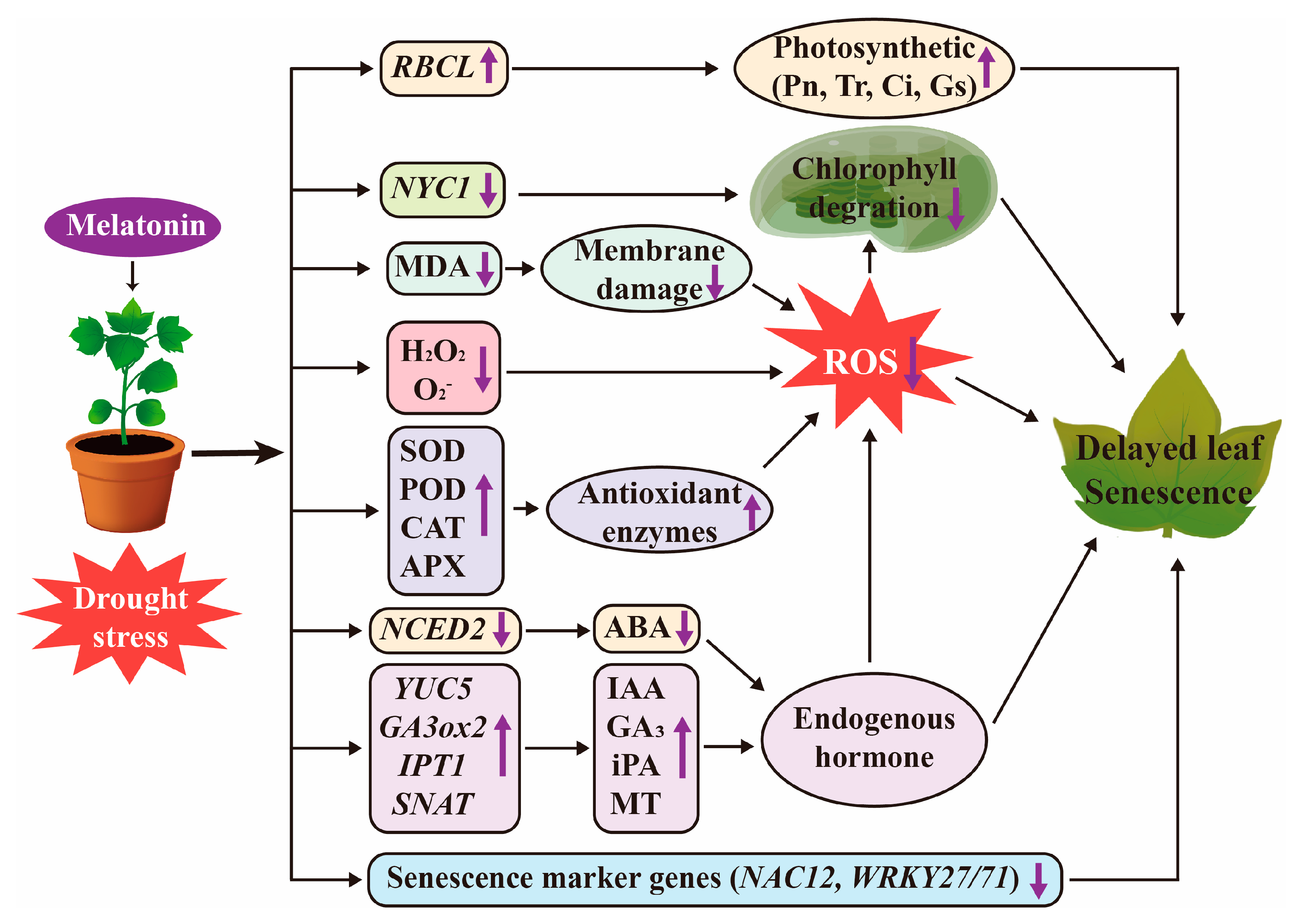
3. Discussion
3.1. MT Delays Leaf Senescence by Regulating Phytohormone Biosynthesis and Expression of Senescence Marker Genes
3.2. MT Delays Leaf Senescence by Inhibiting Chlorophyll Degradation
3.3. MT Delays Leaf Senescence by Regulating Redox Homeostasis
3.4. MT Delays Leaf Senescence and Promotes Plant Growth by Increasing Photosynthetic Efficiency
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Experimental Design
4.3. Plant Growth and Root Morphology
4.4. Evaluation of the Photosynthesis Parameters
4.5. Relative Water Content of the Leaves
4.6. The Antioxidant Enzyme Activity and Contents of Reactive Oxygen Species (ROS) and Malondialdehyde (MDA)
4.7. Determination of the Hormone Content
4.8. Transmission Electron Microscope (TEM) Analysis
4.9. qRT-PCR
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Woo, H.R.; Masclaux-Daubresse, C.; Lim, P.O. Plant Senescence: How Plants Know When and How to Die. J. Exp. Bot. 2018, 69, 715–718. [Google Scholar] [CrossRef]
- Sade, N.; del Mar Rubio-Wilhelmi, M.; Umnajkitikorn, K.; Blumwald, E. Stress-Induced Senescence and Plant Tolerance to Abiotic Stress. J. Exp. Bot. 2018, 69, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Pettigrew, W.T.; McCarty, J.C.; Vaughn, K.C. Leaf Senescence-like Characteristics Contribute to Cotton’s Premature Photosynthetic Decline. Photosynth. Res. 2000, 65, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Djanaguiraman, M.; Prasad, P.V.V.; Djanaguiraman, M.; Prasad, P.V.V. Ethylene Production under High Temperature Stress Causes Premature Leaf Senescence in Soybean. Funct. Plant Biol. 2010, 37, 1071–1084. [Google Scholar] [CrossRef]
- Constable, G.A.; Bange, M.P. The Yield Potential of Cotton (Gossypium hirsutum L.). Field Crops Res. 2015, 182, 98–106. [Google Scholar] [CrossRef]
- Abdelraheem, A.; Adams, N.; Zhang, J. Effects of Drought on Agronomic and Fiber Quality in an Introgressed Backcross Inbred Line Population of Upland Cotton under Field Conditions. Field Crops Res. 2020, 254, 107850. [Google Scholar] [CrossRef]
- Qi, F.; Jian, G.; Li, J. Discrimination on the Relationship among Cotton Premature Senescence, Red Leaf Disease and Alternaria Leaf Spot Disease. Cotton Sci. 2013, 25, 81–85. (In Chinese) [Google Scholar]
- Radin, J.W. Water Relations of Cotton Plants under Nitrogen Deficiency. IV. Leaf Senescence during Drought and Its Relation to Stomatal Closure. Physiol. Plant. 1981, 51, 145–149. [Google Scholar] [CrossRef]
- Abdelraheem, A.; Esmaeili, N.; O’Connell, M.; Zhang, J. Progress and Perspective on Drought and Salt Stress Tolerance in Cotton. Ind. Crops Prod. 2019, 130, 118–129. [Google Scholar] [CrossRef]
- Panigrahi, P.K.; Biswal, U.C. Ageing of Chloroplasts in Vitro I. Quantitative Analysis of the Degradation of Pigments, Proteins and Nucleic Acids. Plant Cell Physiol. 1979, 20, 775–779. [Google Scholar] [CrossRef]
- Hörtensteiner, S. Chlorophyll Degradation during Senescence. Annu. Rev. Plant Biol. 2006, 57, 55–77. [Google Scholar] [CrossRef]
- Rajcan, I.; Dwyer, L.M.; Tollenaar, M. Note on Relationship between Leaf Soluble Carbohydrate and Chlorophyll Concentrations in Maize during Leaf Senescence. Field Crops Res. 1999, 63, 13–17. [Google Scholar] [CrossRef]
- Sakuraba, Y. Light-Mediated Regulation of Leaf Senescence. Int. J. Mol. Sci. 2021, 22, 3291. [Google Scholar] [CrossRef]
- Khanna-Chopra, R. Leaf Senescence and Abiotic Stresses Share Reactive Oxygen Species-Mediated Chloroplast Degradation. Protoplasma 2012, 249, 469–481. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Xu, S.; Mu, D.; Sadeghnezhad, E.; Li, Q.; Ma, Z.; Zhao, L.; Zhang, Q.; Wang, L. Exogenous Melatonin Delays Dark-Induced Grape Leaf Senescence by Regulation of Antioxidant System and Senescence Associated Genes (SAGs). Plants 2019, 8, 366. [Google Scholar] [CrossRef] [Green Version]
- Jahan, M.S.; Shu, S.; Wang, Y.; Hasan, M.d.M.; El-Yazied, A.A.; Alabdallah, N.M.; Hajjar, D.; Altaf, M.A.; Sun, J.; Guo, S. Melatonin Pretreatment Confers Heat Tolerance and Repression of Heat-Induced Senescence in Tomato through the Modulation of ABA- and GA-Mediated Pathways. Front. Plant Sci. 2021, 12, 650955. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Zhao, Y.; Shan, W.; Kuang, J.; Lu, W.; Su, X.; Tao, N.; Lakshmanan, P.; Chen, J. Melatonin Delays Leaf Senescence of Postharvest Chinese Flowering Cabbage through ROS Homeostasis. Food Res. Int. 2020, 138, 109790. [Google Scholar] [CrossRef] [PubMed]
- Jibran, R.; Hunter, D.A.; Dijkwel, P.P. Hormonal Regulation of Leaf Senescence through Integration of Developmental and Stress Signals. Plant Mol. Biol. 2013, 82, 547–561. [Google Scholar] [CrossRef]
- Asad, M.; Zakari, S.; Zhao, Q.; Zhou, L.; Ye, Y.; Cheng, F. Abiotic Stresses Intervene with ABA Signaling to Induce Destructive Metabolic Pathways Leading to Death: Premature Leaf Senescence in Plants. Int. J. Mol. Sci. 2019, 20, 256. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.I.; Murphy, A.S.; Baek, D.; Lee, S.-W.; Yun, D.-J.; Bressan, R.A.; Narasimhan, M.L. YUCCA6 Over-Expression Demonstrates Auxin Function in Delaying Leaf Senescence in Arabidopsis thaliana. J. Exp. Bot. 2011, 62, 3981–3992. [Google Scholar] [CrossRef]
- Singh, S.; Letham, D.S.; Palni, L.M.S. Cytokinin Biochemistry in Relation to Leaf Senescence. VII. Endogenous Cytokinin Levels and Exogenous Applications of Cytokinins in Relation to Sequential Leaf Senescence of Tobacco. Physiol. Plant. 1992, 86, 388–397. [Google Scholar] [CrossRef]
- Fletcher, R.A.; Osborne, D.J. Regulation of Protein and Nucleic Acid Synthesis by Gibberellin during Leaf Senescence. Nature 1965, 207, 1176–1177. [Google Scholar] [CrossRef]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY Transcription Factors. Trends Plant Sci. 2010, 15, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Nam, H.G.; Lim, P.O. Regulatory Network of NAC Transcription Factors in Leaf Senescence. Curr. Opin. Plant Biol. 2016, 33, 48–56. [Google Scholar] [CrossRef]
- Bhattacharya, A. Soil Water Deficit and Physiological Issues in Plants; Springer: Singapore, 2021; ISBN 978-981-336-275-8. [Google Scholar]
- Pic, E.; de la Serve, B.T.; Tardieu, F.; Turc, O. Leaf Senescence Induced by Mild Water Deficit Follows the Same Sequence of Macroscopic, Biochemical, and Molecular Events as Monocarpic Senescence in Pea. Plant Physiol. 2002, 128, 236–246. [Google Scholar] [CrossRef]
- Bandurska, H. Drought Stress Responses: Coping Strategy and Resistance. Plants 2022, 11, 922. [Google Scholar] [CrossRef]
- Dong, H.; Li, W.; Eneji, A.E.; Zhang, D. Nitrogen Rate and Plant Density Effects on Yield and Late-Season Leaf Senescence of Cotton Raised on a Saline Field. Field Crops Res. 2012, 126, 137–144. [Google Scholar] [CrossRef]
- Chen, Y.; Dong, H. Mechanisms and Regulation of Senescence and Maturity Performance in Cotton. Field Crops Res. 2016, 189, 1–9. [Google Scholar] [CrossRef]
- Cook, B.I.; Smerdon, J.E.; Seager, R.; Coats, S. Global Warming and 21st Century Drying. Clim. Dyn. 2014, 43, 2607–2627. [Google Scholar] [CrossRef] [Green Version]
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of Extreme Weather Disasters on Global Crop Production. Nature 2016, 529, 84–87. [Google Scholar] [CrossRef] [Green Version]
- Rivero, R.M.; Kojima, M.; Gepstein, A.; Sakakibara, H.; Mittler, R.; Gepstein, S.; Blumwald, E. Delayed Leaf Senescence Induces Extreme Drought Tolerance in a Flowering Plant. Proc. Natl. Acad. Sci. USA 2007, 104, 19631–19636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubbels, R.; Reiter, R.J.; Klenke, E.; Goebel, A.; Schnakenberg, E.; Ehlers, C.; Schiwara, H.W.; Schloot, W. Melatonin in Edible Plants Identified by Radioimmunoassay and by High Performance Liquid Chromatography-Mass Spectrometry. J. Pineal Res. 1995, 18, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Marta, B.; Szafrańska, K.; Posmyk, M.M. Exogenous Melatonin Improves Antioxidant Defense in Cucumber Seeds (Cucumis sativus L.) Germinated under Chilling Stress. Front. Plant Sci. 2016, 7, 575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelagio-Flores, R.; Muñoz-Parra, E.; Ortiz-Castro, R.; López-Bucio, J. Melatonin Regulates Arabidopsis Root System Architecture Likely Acting Independently of Auxin Signaling: Role of Melatonin in Root Architecture and Auxin Responses. J. Pineal Res. 2012, 53, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ruiz, J.; Cano, A.; Arnao, M.B. Melatonin Acts as a Growth-Stimulating Compound in Some Monocot Species: Melatonin as Growth Promoter in Plants. J. Pineal Res. 2005, 39, 137–142. [Google Scholar] [CrossRef]
- Ahmad, S.; Su, W.; Kamran, M.; Ahmad, I.; Meng, X.; Wu, X.; Javed, T.; Han, Q. Foliar Application of Melatonin Delay Leaf Senescence in Maize by Improving the Antioxidant Defense System and Enhancing Photosynthetic Capacity under Semi-Arid Regions. Protoplasma 2020, 257, 1079–1092. [Google Scholar] [CrossRef]
- Martínez-Lorente, S.E.; Pardo-Hernández, M.; Martí-Guillén, J.M.; López-Delacalle, M.; Rivero, R.M. Interaction between Melatonin and NO: Action Mechanisms, Main Targets, and Putative Roles of the Emerging Molecule NOmela. Int. J. Mol. Sci. 2022, 23, 6646. [Google Scholar] [CrossRef]
- Wang, P.; Sun, X.; Li, C.; Wei, Z.; Liang, D.; Ma, F. Long-Term Exogenous Application of Melatonin Delays Drought-Induced Leaf Senescence in Apple. J. Pineal Res. 2013, 54, 292–302. [Google Scholar] [CrossRef]
- Zhang, J.; Li, H.; Xu, B.; Li, J.; Huang, B. Exogenous Melatonin Suppresses Dark-Induced Leaf Senescence by Activating the Superoxide Dismutase-Catalase Antioxidant Pathway and Down-Regulating Chlorophyll Degradation in Excised Leaves of Perennial Ryegrass (Lolium perenne L.). Front. Plant Sci. 2016, 7, 1500. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Jing, T.; Wang, Y.; Ai, X.; Bi, H. Melatonin Delays Leaf Senescence and Improves Cucumber Yield by Modulating Chlorophyll Degradation and Photoinhibition of PSII and PSI. Environ. Exp. Bot. 2022, 200, 104915. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin and Its Relationship to Plant Hormones. Ann. Bot. 2018, 121, 195–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Jia, W.; Li, S.; Xu, J.; Xu, Z. Enhancement of Nicotiana tabacum Resistance against Dehydration-Induced Leaf Senescence via Metabolite/Phytohormone-Gene Regulatory Networks Modulated by Melatonin. Front. Plant Sci. 2021, 12, 686062. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shi, Y.; Zhang, X.; Du, H.; Xu, B.; Huang, B. Melatonin Suppression of Heat-Induced Leaf Senescence Involves Changes in Abscisic Acid and Cytokinin Biosynthesis and Signaling Pathways in Perennial Ryegrass (Lolium perenne L.). Environ. Exp. Bot. 2017, 138, 36–45. [Google Scholar] [CrossRef]
- Zhao, Y.-Q.; Zhang, Z.-W.; Chen, Y.-E.; Ding, C.-B.; Yuan, S.; Reiter, R.J.; Yuan, M. Melatonin: A Potential Agent in Delaying Leaf Senescence. Crit. Rev. Plant Sci. 2021, 40, 1–22. [Google Scholar] [CrossRef]
- Hörtensteiner, S. Stay-Green Regulates Chlorophyll and Chlorophyll-Binding Protein Degradation during Senescence. Trends Plant Sci. 2009, 14, 155–162. [Google Scholar] [CrossRef]
- Kamran, M.; Ahmad, S.; Ahmad, I.; Hussain, I.; Meng, X.; Zhang, X.; Javed, T.; Ullah, M.; Ding, R.; Xu, P.; et al. Paclobutrazol Application Favors Yield Improvement of Maize under Semiarid Regions by Delaying Leaf Senescence and Regulating Photosynthetic Capacity and Antioxidant System during Grain-Filling Stage. Agronomy 2020, 10, 187. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Seo, P.J.; Lee, H.-J.; Park, C.-M. A NAC Transcription Factor NTL4 Promotes Reactive Oxygen Species Production during Drought-Induced Leaf Senescence in Arabidopsis: NTL4 in Drought-Induced Leaf Senescence. Plant J. 2012, 70, 831–844. [Google Scholar] [CrossRef]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The Physiology of Plant Responses to Drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Kanwar, M.K.; Yu, J.; Zhou, J. Phytomelatonin: Recent Advances and Future Prospects. J. Pineal Res. 2018, 65, e12526. [Google Scholar] [CrossRef] [Green Version]
- Shimamura, S.; Yoshioka, T.; Yamamoto, R.; Hiraga, S.; Nakamura, T.; Shimada, S.; Komatsu, S. Role of Abscisic Acid in Flood-Induced Secondary Aerenchyma Formation in Soybean (Glycine max) Hypocotyls. Plant Prod. Sci. 2014, 17, 131–137. [Google Scholar] [CrossRef]
- Dong, H.; Niu, Y.; Li, W.; Zhang, D. Effects of Cotton Rootstock on Endogenous Cytokinins and Abscisic Acid in Xylem Sap and Leaves in Relation to Leaf Senescence. J. Exp. Bot. 2008, 59, 1295–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Y.; Xu, X.; Li, L.; Sun, Q.; Wang, Q.; Huang, H.; Tong, Z.; Zhang, J. Melatonin-Mediated Development and Abiotic Stress Tolerance in Plants. Front. Plant Sci. 2023, 14, 1100827. [Google Scholar] [CrossRef] [PubMed]
- Smart, C.M. Gene Expression during Leaf Senescence. New Phytol. 1994, 126, 419–448. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.T.; Pang, C.; Fan, S.; Song, M.; Arain, S.; Yu, S. Isolation and Expression Profiling of GhNAC Transcription Factor Genes in Cotton (Gossypium hirsutum L.) during Leaf Senescence and in Response to Stresses. Gene 2013, 531, 220–234. [Google Scholar] [CrossRef]
- Bengoa Luoni, S.; Astigueta, F.H.; Nicosia, S.; Moschen, S.; Fernandez, P.; Heinz, R. Transcription Factors Associated with Leaf Senescence in Crops. Plants 2019, 8, 411. [Google Scholar] [CrossRef] [Green Version]
- Matile, P.; Hörtensteiner, S.; Thomas, H. Chlorophyll Degradation. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 67–95. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Zhang, J.; Burgess, P.; Rossi, S.; Huang, B. Interactive Effects of Melatonin and Cytokinin on Alleviating Drought-Induced Leaf Senescence in Creeping Bentgrass (Agrostis stolonifera). Environ. Exp. Bot. 2018, 145, 1–11. [Google Scholar] [CrossRef]
- Ackerson, R.C.; Hebert, R.R. Osmoregulation in Cotton in Response to Water Stress: I. Alterations in Photosynthesis, Leaf Conductance, Translocation, and Ultrastructure. Plant Physiol. 1981, 67, 484–488. [Google Scholar] [CrossRef] [Green Version]
- Jajic, I.; Sarna, T.; Strzalka, K. Senescence, Stress, and Reactive Oxygen Species. Plants 2015, 4, 393–411. [Google Scholar] [CrossRef] [Green Version]
- Imran, M.; Aaqil Khan, M.; Shahzad, R.; Bilal, S.; Khan, M.; Yun, B.-W.; Khan, A.L.; Lee, I.-J. Melatonin Ameliorates Thermotolerance in Soybean Seedling through Balancing Redox Homeostasis and Modulating Antioxidant Defense, Phytohormones and Polyamines Biosynthesis. Molecules 2021, 26, 5116. [Google Scholar] [CrossRef]
- Jiang, D.; Lu, B.; Liu, L.; Duan, W.; Meng, Y.; Li, J.; Zhang, K.; Sun, H.; Zhang, Y.; Dong, H.; et al. Exogenous Melatonin Improves the Salt Tolerance of Cotton by Removing Active Oxygen and Protecting Photosynthetic Organs. BMC Plant Biol. 2021, 21, 331. [Google Scholar] [CrossRef] [PubMed]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.-J. The Role of the Plant Antioxidant System in Drought Tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farouk, S.; Al-Amri, S.M. Ameliorative Roles of Melatonin and/or Zeolite on Chromium-Induced Leaf Senescence in Marjoram Plants by Activating Antioxidant Defense, Osmolyte Accumulation, and Ultrastructural Modification. Ind. Crops Prod. 2019, 142, 111823. [Google Scholar] [CrossRef]
- Balazadeh, S.; Parlitz, S.; Mueller-Roeber, B.; Meyer, R.C. Natural Developmental Variations in Leaf and Plant Senescence in Arabidopsis thaliana. Plant Biol. 2008, 10, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Krieger-Liszkay, A.; Krupinska, K.; Shimakawa, G. The Impact of Photosynthesis on Initiation of Leaf Senescence. Physiol. Plant. 2019, 166, 148–164. [Google Scholar] [CrossRef]
- Wang, R.; Gao, M.; Ji, S.; Wang, S.; Meng, Y.; Zhou, Z. Carbon Allocation, Osmotic Adjustment, Antioxidant Capacity and Growth in Cotton under Long-Term Soil Drought during Flowering and Boll-Forming Period. Plant Physiol. Biochem. 2016, 107, 137–146. [Google Scholar] [CrossRef]
- Altaf, M.A.; Shahid, R.; Ren, M.-X.; Altaf, M.M.; Khan, L.U.; Shahid, S.; Jahan, M.S. Melatonin Alleviates Salt Damage in Tomato Seedling: A Root Architecture System, Photosynthetic Capacity, Ion Homeostasis, and Antioxidant Enzymes Analysis. Sci. Hortic. 2021, 285, 110145. [Google Scholar] [CrossRef]
- Meng, J.-F.; Xu, T.-F.; Wang, Z.-Z.; Fang, Y.-L.; Xi, Z.-M.; Zhang, Z.-W. The Ameliorative Effects of Exogenous Melatonin on Grape Cuttings under Water-Deficient Stress: Antioxidant Metabolites, Leaf Anatomy, and Chloroplast Morphology. J. Pineal Res. 2014, 57, 200–212. [Google Scholar] [CrossRef]
- Kong, X.; Luo, Z.; Dong, H.; Eneji, A.E.; Li, W.; Lu, H. Gene Expression Profiles Deciphering Leaf Senescence Variation between Early- and Late-Senescence Cotton Lines. PLoS ONE 2013, 8, e69847. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Zhang, J.; Yan, K.; Zhou, Z.; Zhao, W.; Zhang, X.; Pu, Y.; Yu, R. Beneficial Effects of Abscisic Acid and Melatonin in Overcoming Drought Stress in Cotton (Gossypium hirsutum L.). Physiol. Plant. 2021, 173, 2041–2054. [Google Scholar] [CrossRef]
- Khan, M.N.; Zhang, J.; Luo, T.; Liu, J.; Rizwan, M.; Fahad, S.; Xu, Z.; Hu, L. Seed Priming with Melatonin Coping Drought Stress in Rapeseed by Regulating Reactive Oxygen Species Detoxification: Antioxidant Defense System, Osmotic Adjustment, Stomatal Traits and Chloroplast Ultrastructure Perseveration. Ind. Crops Prod. 2019, 140, 111597. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: A New Plant Hormone and/or a Plant Master Regulator? Trends Plant Sci. 2019, 24, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Oukarroum, A.; Bussotti, F.; Goltsev, V.; Kalaji, H.M. Correlation between Reactive Oxygen Species Production and Photochemistry of Photosystems I and II in Lemna gibba L. Plants under Salt Stress. Environ. Exp. Bot. 2015, 109, 80–88. [Google Scholar] [CrossRef]
- Sperdouli, I.; Moustaka, J.; Ouzounidou, G.; Moustakas, M. Leaf Age-Dependent Photosystem II Photochemistry and Oxidative Stress Responses to Drought Stress in Arabidopsis thaliana Are Modulated by Flavonoid Accumulation. Molecules 2021, 26, 4157. [Google Scholar] [CrossRef]
- Talla, S.K.; Panigrahy, M.; Kappara, S.; Nirosha, P.; Neelamraju, S.; Ramanan, R. Cytokinin Delays Dark-Induced Senescence in Rice by Maintaining the Chlorophyll Cycle and Photosynthetic Complexes. J. Exp. Bot. 2016, 67, 1839–1851. [Google Scholar] [CrossRef] [Green Version]
- Janečková, H.; Husičková, A.; Lazár, D.; Ferretti, U.; Pospíšil, P.; Špundová, M. Exogenous Application of Cytokinin during Dark Senescence Eliminates the Acceleration of Photosystem II Impairment Caused by Chlorophyll b Deficiency in Barley. Plant Physiol. Biochem. 2019, 136, 43–51. [Google Scholar] [CrossRef]
- Wingler, A.; Marès, M.; Pourtau, N. Spatial Patterns and Metabolic Regulation of Photosynthetic Parameters during Leaf Senescence. New Phytol. 2004, 161, 781–789. [Google Scholar] [CrossRef]
- Fleta-Soriano, E.; Díaz, L.; Bonet, E.; Munné-Bosch, S. Melatonin May Exert a Protective Role against Drought Stress in Maize. J. Agron. Crop Sci. 2017, 203, 286–294. [Google Scholar] [CrossRef]
- Kusaba, M.; Ito, H.; Morita, R.; Iida, S.; Sato, Y.; Fujimoto, M.; Kawasaki, S.; Tanaka, R.; Hirochika, H.; Nishimura, M.; et al. Rice Non-Yellow Coloring1 Is Involved in Light-Harvesting Complex II and Grana Degradation during Leaf Senescence. Plant Cell 2007, 19, 1362–1375. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Cai, S.; Xing, Q.; Qi, Z.; Fotopoulos, V.; Yu, J.; Zhou, J. Melatonin Delays Dark-induced Leaf Senescence by Inducing MiR171b Expression in Tomato. J. Pineal Res. 2022, 72, e12792. [Google Scholar] [CrossRef]
- Bassi, R.; Dall’Osto, L. Dissipation of Light Energy Absorbed in Excess: The Molecular Mechanisms. Annu. Rev. Plant Biol. 2021, 72, 47–76. [Google Scholar] [CrossRef] [PubMed]
- Werner, C.; Ryel, R.J.; Correia, O.; Beyschlag, W. Effects of Photoinhibition on Whole-plant Carbon Gain Assessed with a Photosynthesis Model. Plant Cell Environ. 2001, 24, 27–40. [Google Scholar] [CrossRef]
- De Souza, A.P.; Burgess, S.J.; Doran, L.; Hansen, J.; Manukyan, L.; Maryn, N.; Gotarkar, D.; Leonelli, L.; Niyogi, K.K.; Long, S.P. Soybean Photosynthesis and Crop Yield Are Improved by Accelerating Recovery from Photoprotection. Science 2022, 377, 851–854. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.; Zhao, X.; Liu, S.; Sun, F.; Zhang, C.; Xi, Y. Beneficial Effects of Melatonin in Overcoming Drought Stress in Wheat Seedlings. Plant Physiol. Biochem. 2017, 118, 138–149. [Google Scholar] [CrossRef]
- Altaf, M.A.; Shahid, R.; Ren, M.-X.; Naz, S.; Altaf, M.M.; Khan, L.U.; Tiwari, R.K.; Lal, M.K.; Shahid, M.A.; Kumar, R.; et al. Melatonin Improves Drought Stress Tolerance of Tomato by Modulating Plant Growth, Root Architecture, Photosynthesis, and Antioxidant Defense System. Antioxidants 2022, 11, 309. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, L.; Zhou, K.; Zhang, Y.; Han, X.; Din, Y.; Ge, X.; Qin, W.; Wang, P.; Li, F.; et al. GhWRKY6 Acts as a Negative Regulator in Both Transgenic Arabidopsis and Cotton during Drought and Salt Stress. Front. Genet. 2019, 10, 392. [Google Scholar] [CrossRef]
- Murch, S.J.; KrishnaRaj, S.; Saxena, P.K. Tryptophan Is a Precursor for Melatonin and Serotonin Biosynthesis in in Vitro Regenerated St. John’s Wort (Hypericum perforatum L. Cv. Anthos) Plants. Plant Cell Rep. 2000, 19, 698–704. [Google Scholar] [CrossRef]
- Khattak, W.A.; He, J.; Abdalmegeed, D.; Hu, W.; Wang, Y.; Zhou, Z. Foliar Melatonin Stimulates Cotton Boll Distribution Characteristics by Modifying Leaf Sugar Metabolism and Antioxidant Activities during Drought Conditions. Physiol. Plant. 2022, 174, e13526. [Google Scholar] [CrossRef]
- Mao, L.; Zhang, L.; Zhao, X.; Liu, S.; van der Werf, W.; Zhang, S.; Spiertz, H.; Li, Z. Crop Growth, Light Utilization and Yield of Relay Intercropped Cotton as Affected by Plant Density and a Plant Growth Regulator. Field Crops Res. 2014, 155, 67–76. [Google Scholar] [CrossRef]
- Zhu, L.; Liu, L.; Sun, H.; Zhang, Y.; Liu, X.; Wang, N.; Chen, J.; Zhang, K.; Bai, Z.; Wang, G.; et al. The Responses of Lateral Roots and Root Hairs to Nitrogen Stress in Cotton Based on Daily Root Measurements. J. Agron. Crop Sci. 2022, 208, 89–105. [Google Scholar] [CrossRef]
- Ye, Z.P.; Yu, Q. A Coupled Model of Stomatal Conductance and Photosynthesis for Winter Wheat. Photosynthetica 2008, 46, 637–640. [Google Scholar] [CrossRef]
- Barrs, H.; Weatherley, P. A Re-Examination of the Relative Turgidity Technique for Estimating Water Deficits in Leaves. Aust. J. Biol. Sci. 1962, 15, 413. [Google Scholar] [CrossRef] [Green Version]
- Daud, M.K.; Quiling, H.; Lei, M.; Ali, B.; Zhu, S.J. Ultrastructural, Metabolic and Proteomic Changes in Leaves of Upland Cotton in Response to Cadmium Stress. Chemosphere 2015, 120, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Duan, W.; Lu, B.; Liu, L.; Meng, Y.; Ma, X.; Li, J.; Zhang, K.; Sun, H.; Zhang, Y.; Dong, H.; et al. Effects of Exogenous Melatonin on Root Physiology, Transcriptome and Metabolome of Cotton Seedlings under Salt Stress. Int. J. Mol. Sci. 2022, 23, 9456. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Pmax (μmol/m2 s−1) | LSP (μmol/m2 s−1) | LCP (μmol/m2 s−1) | Rd (μmol/m2 s−1) | AQE (μmol/mol) | R2 of Model Fitting |
|---|---|---|---|---|---|---|
| CK | 20.68 ± 3.94 a | 1761.44 ± 227.26 a | 50.64 ± 7.6 a | 3.18 ± 0.5 a | 0.0688 ± 0.0015 a | 0.9995 |
| DS | 12.84 ± 1.85 b | 1447.5 ± 179.85 a | 16.63 ± 3.06 c | 0.82 ± 0.05 c | 0.0526 ± 0.0123 a | 0.9984 |
| MT + DS | 17.43 ± 0.27 ab | 1637.41 ± 68.71 a | 33.21 ± 4.39 b | 2.08 ± 0.24 b | 0.0675 ± 0.0013 a | 0.9995 |
| Gene Name | Forward Primer (5′ to 3′) | Reverse Primer (5′ to 3′) |
|---|---|---|
| GhRBCL | TTCCGAGTAACTCCTCAACCC | CTTACAGATGCACCGCCCG |
| GhNYC1 | TGCTTGTGGTGGGCTGCTA | GTCAGCGAAAGTACGCACAA |
| GhWRKY27 | CCTACGGAACAGCCACAACAA | CTTCCAGCAGGTTTTCAGAGC |
| GhWRKY71 | ACTTGGGTGGATACGACTCTC | TCCTGCTCACCAATTCCATGT |
| GhNAC12 | CAAAAGAGTCCCAATGGCAA | CTGCTTTACGGTCTTCTATCGG |
| GhYUC5 | ACTCCTGCTTTTGTTCTTCTT | CTTGTGAAACCGTCTCGTT |
| GhNCED2 | TACGATGTTATCCAAAAGCC | GAAAATAGAGTCGGGAGGTG |
| GhGA3ox2 | AACTTAAAGCTGCAAGGTCTC | ACTCCTAATCTACTTCGTGGG |
| GhIPT1 | CAATCGCACGCCTAATCCCT | GCATTGGTGAATGATGACCCT |
| GhSNAT | CACAAATCGTTGAACCACC | AATTAGCCTCTTTTGCTCATT |
| ACTIN14 | ATCCTCCGTCTTGACCTTG | TGTCCGTCAGGCAACTCAT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, K.; Sun, H.; Liu, M.; Zhu, L.; Zhang, K.; Zhang, Y.; Li, A.; Zhang, H.; Zhu, J.; Liu, X.; et al. Morphological and Physiological Mechanisms of Melatonin on Delaying Drought-Induced Leaf Senescence in Cotton. Int. J. Mol. Sci. 2023, 24, 7269. https://doi.org/10.3390/ijms24087269
Yang K, Sun H, Liu M, Zhu L, Zhang K, Zhang Y, Li A, Zhang H, Zhu J, Liu X, et al. Morphological and Physiological Mechanisms of Melatonin on Delaying Drought-Induced Leaf Senescence in Cotton. International Journal of Molecular Sciences. 2023; 24(8):7269. https://doi.org/10.3390/ijms24087269
Chicago/Turabian StyleYang, Kai, Hongchun Sun, Mengxing Liu, Lingxiao Zhu, Ke Zhang, Yongjiang Zhang, Anchang Li, Haina Zhang, Jijie Zhu, Xiaoqing Liu, and et al. 2023. "Morphological and Physiological Mechanisms of Melatonin on Delaying Drought-Induced Leaf Senescence in Cotton" International Journal of Molecular Sciences 24, no. 8: 7269. https://doi.org/10.3390/ijms24087269