
Plasma Exosomal Non-Coding RNA Profile Associated with Renal Damage Reveals Potential Therapeutic Targets in Lupus Nephritis
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. Study Population
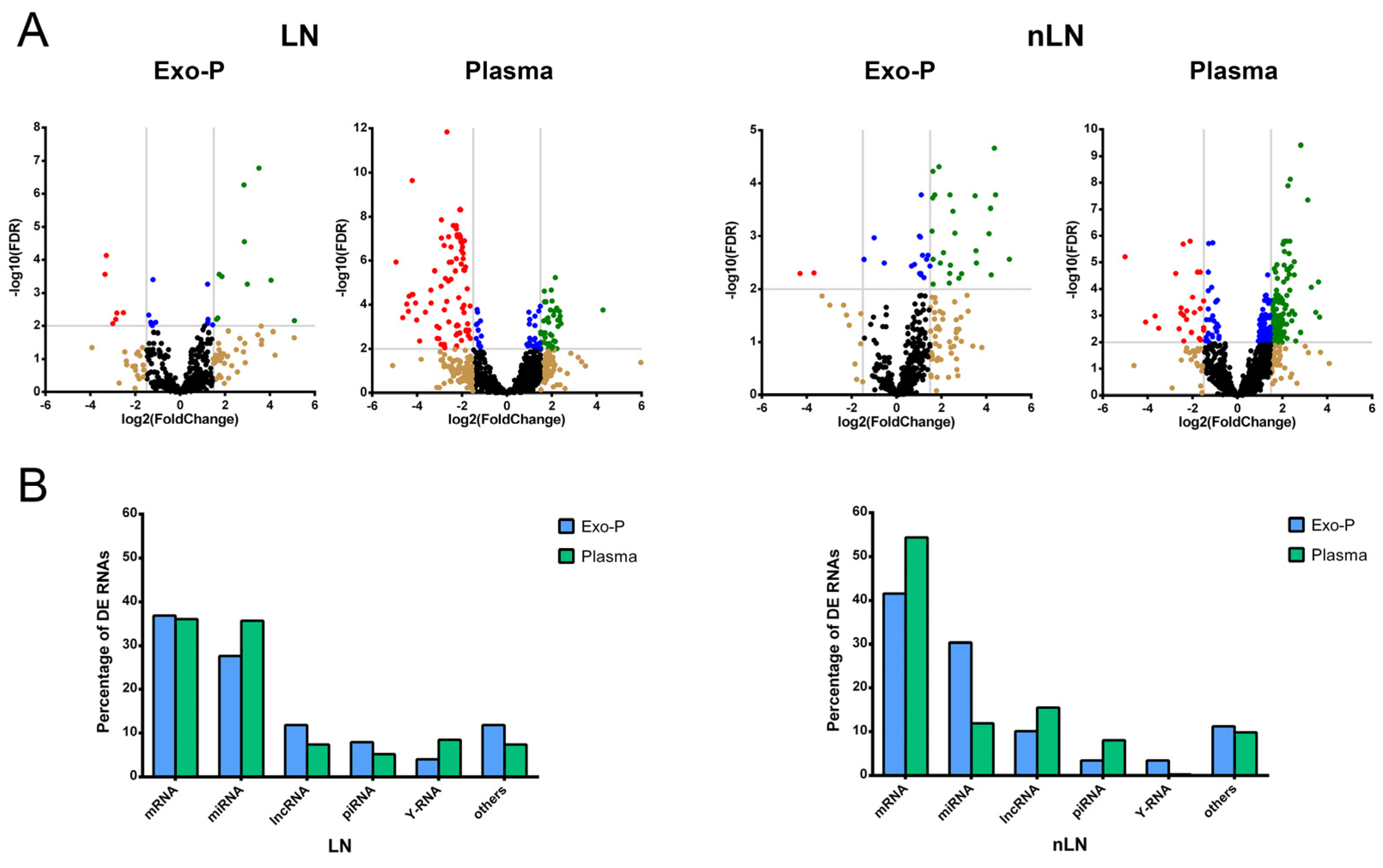
2.2. Proportions of Differentially Expressed RNA Types in Systemic Lupus Eryhtematosus with or without Lupus Nephritis According to Biofluid
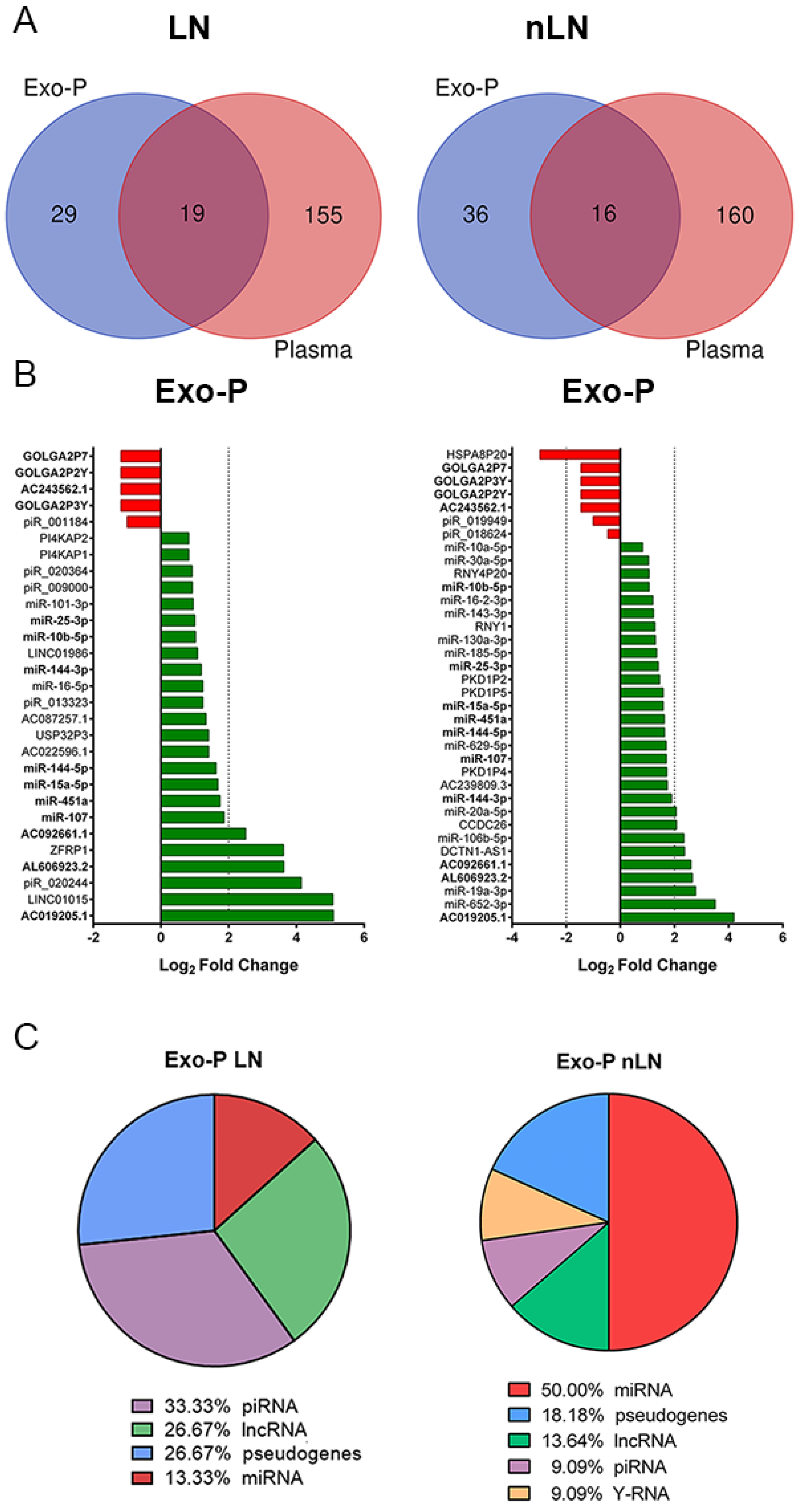
2.3. Plasma Exosomal ncRNA Signature Sssociated with Renal Damage
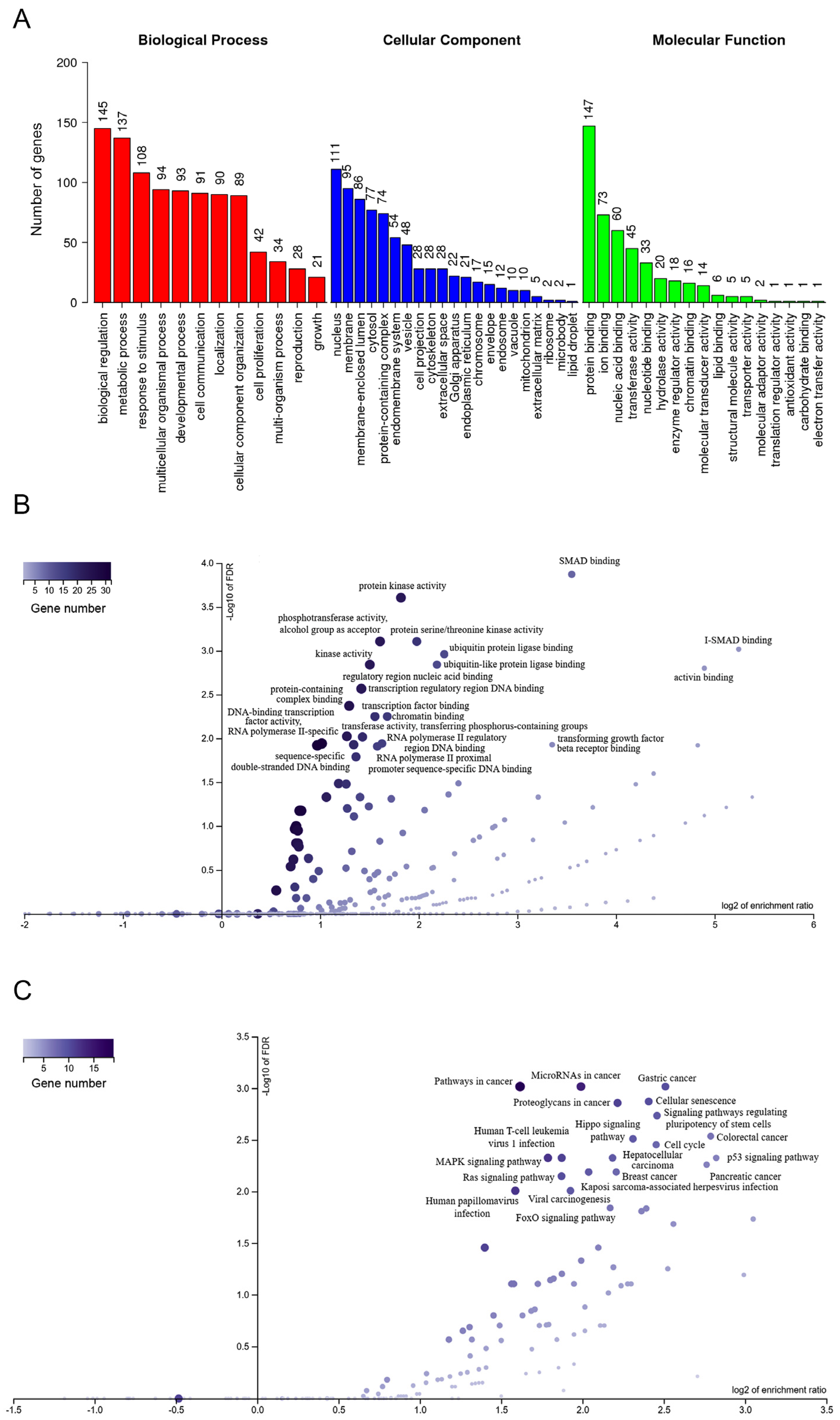
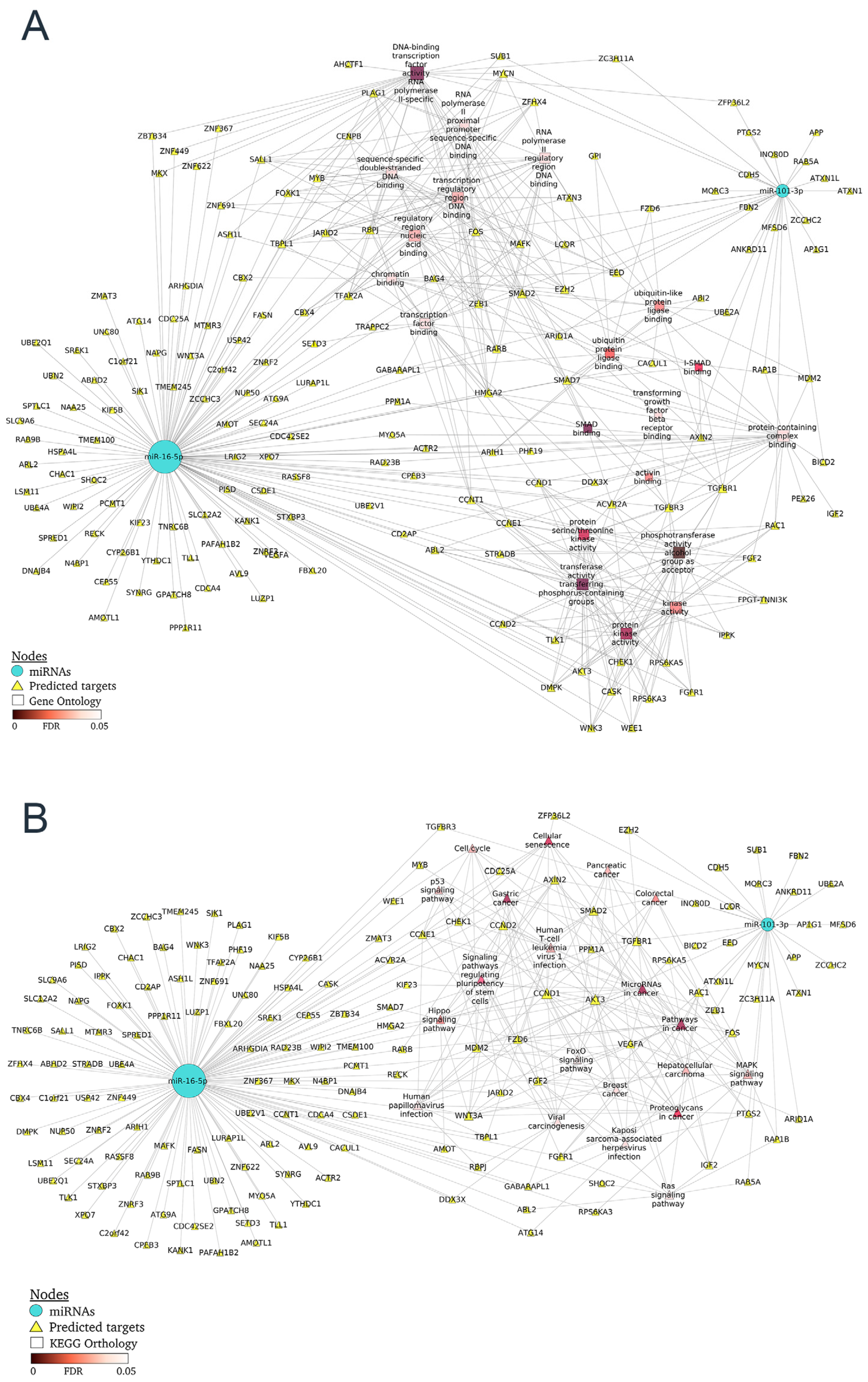
2.4. Differentially Expressed miRNA-mRNA Network from Patients with Nephritis
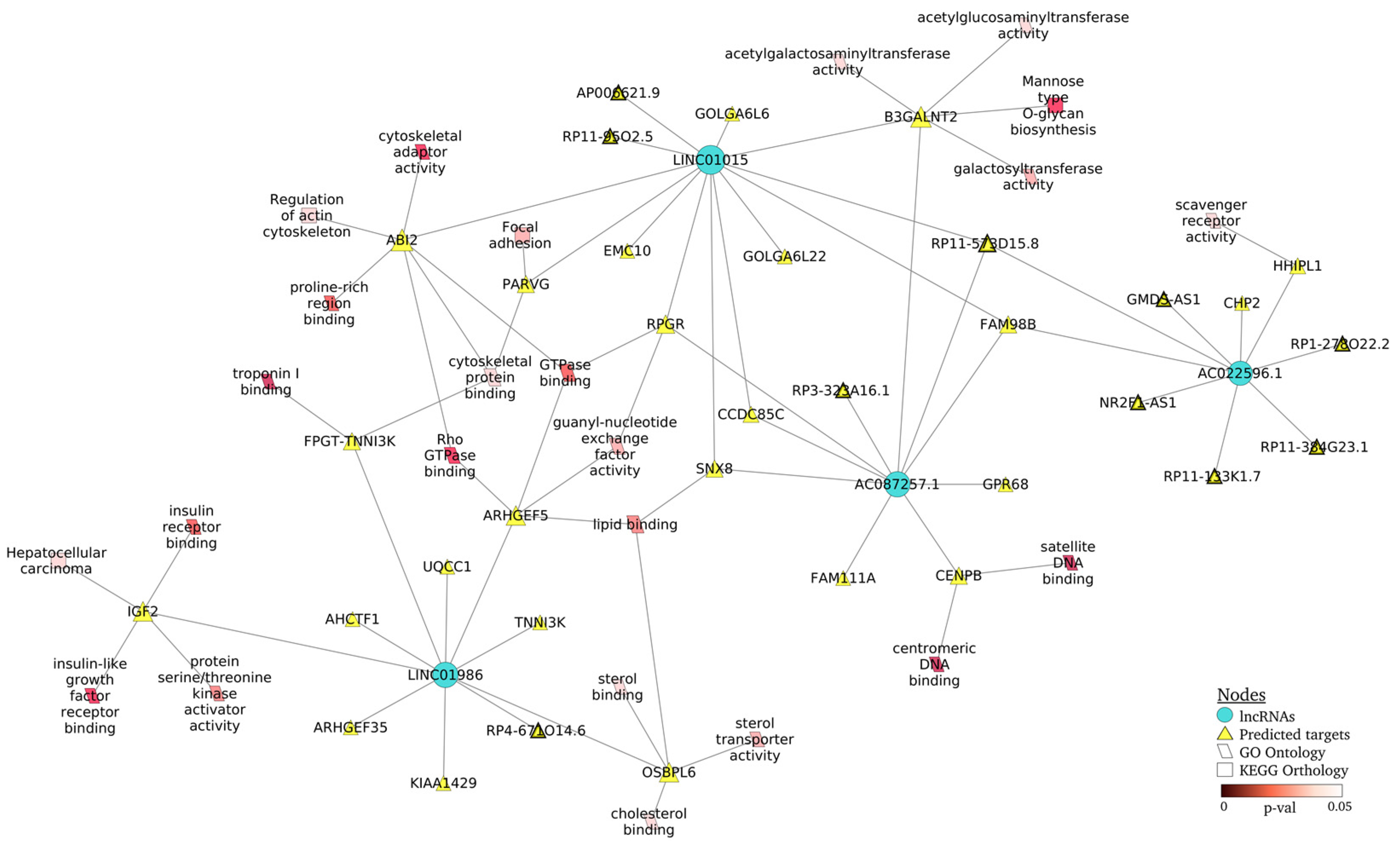
2.5. Regulation Networks of lncRNA–mRNA from Patients with Nephritis
3. Discussion
4. Materials and Methods
4.1. Subjects and Samples
4.2. Samples
4.3. RNA Extraction, Small RNA Library Preparation, and Next-Generation Sequencing
4.4. Statistical Analysis
4.5. Non-Coding RNA Target Predictions and Functional Enrichment Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Beermann, J.; Piccoli, M.T.; Viereck, J.; Thum, T. Non-coding RNAs in Development and Disease: Background, Mechanisms, and Therapeutic Approaches. Physiol. Rev. 2016, 96, 1297–1325. [Google Scholar] [CrossRef] [Green Version]
- Perez-Hernandez, J.; Forner, M.J.; Pinto, C.; Chaves, F.J.; Cortes, R.; Redon, J. Increased Urinary Exosomal MicroRNAs in Patients with Systemic Lupus Erythematosus. PLoS ONE 2015, 10, e0138618. [Google Scholar] [CrossRef] [Green Version]
- Cortes, R.; Forner, M.J. Circular RNAS: Novel biomarkers of disease activity in systemic lupus erythematosus? Clin. Sci. 2019, 133, 1049–1052. [Google Scholar] [CrossRef]
- Perez-Hernandez, J.; Martinez-Arroyo, O.; Ortega, A.; Galera, M.; Solis-Salguero, M.A.; Chaves, F.J.; Redon, J.; Forner, M.J.; Cortes, R. Urinary exosomal miR-146a as a marker of albuminuria, activity changes and disease flares in lupus nephritis. J. Nephrol. 2021, 34, 1157–1167. [Google Scholar] [CrossRef]
- Song, W.; Qiu, J.; Yin, L.; Hong, X.; Dai, W.; Tang, D.; Liu, D.; Dai, Y. Integrated analysis of competing endogenous RNA networks in peripheral blood mononuclear cells of systemic lupus erythematosus. J. Transl. Med. 2021, 19, 362. [Google Scholar] [CrossRef]
- Ye, H.; Wang, X.; Wang, L.; Chu, X.; Hu, X.; Sun, L.; Jiang, M.; Wang, H.; Wang, Z.; Zhao, H.; et al. Full high-throughput sequencing analysis of differences in expression profiles of long noncoding RNAs and their mechanisms of action in systemic lupus erythematosus. Arthritis Res. Ther. 2019, 21, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, X.; Chen, S.; Shen, R.; Liu, S.; You, Y. HOXA11-OS participates in lupus nephritis by targeting miR-124-3p mediating Cyr61 to regulate podocyte autophagy. Mol. Med. 2022, 28, 138. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, R.F.; Shaker, O.G.; Abdelghany, H.M.; Helmy Abdallah, N.; Elsayed, S.H.; Kamel, B.A. Role of micro-RNA132 and its long non coding SOX2 in diagnosis of lupus nephritis. Lupus 2022, 31, 89–96. [Google Scholar] [CrossRef]
- Driedonks, T.A.P.; Nolte-’t Hoen, E.N.M. Circulating Y-RNAs in Extracellular Vesicles and Ribonucleoprotein Complexes; Implications for the Immune System. Front. Immunol. 2018, 9, 3164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boccitto, M.; Wolin, S.L. Ro60 and Y RNAs: Structure, functions, and roles in autoimmunity. Crit. Rev. Biochem. Mol. Biol. 2019, 54, 133–152. [Google Scholar] [CrossRef]
- Yang, P.; Zhang, X.; Chen, S.; Tao, Y.; Ning, M.; Zhu, Y.; Liang, J.; Kong, W.; Shi, B.; Li, Z.; et al. A Novel Serum tsRNA for Diagnosis and Prediction of Nephritis in SLE. Front. Immunol. 2021, 12, 735105. [Google Scholar] [CrossRef]
- Martinez-Arroyo, O.; Ortega, A.; Redon, J.; Cortes, R. Therapeutic Potential of Extracellular Vesicles in Hypertension-Associated Kidney Disease. Hypertension 2021, 77, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Goh, T.X.; Tan, S.L.; Roebuck, M.M.; Teo, S.H.; Kamarul, T. A Systematic Review of Extracellular Vesicle-Derived Piwi-Interacting RNA in Human Body Fluid and Its Role in Disease Progression. Tissue Eng. Part. C Methods 2022, 28, 511–528. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.Y.; Hsieh, S.C.; Lu, C.S.; Wu, T.H.; Liao, H.T.; Wu, C.H.; Li, K.J.; Kuo, Y.M.; Lee, H.T.; Shen, C.Y.; et al. Cross-Talk between Mitochondrial Dysfunction-Provoked Oxidative Stress and Aberrant Noncoding RNA Expression in the Pathogenesis and Pathophysiology of SLE. Int. J. Mol. Sci. 2019, 20, 5183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Arroyo, O.; Ortega, A.; Forner, M.J.; Cortes, R. Mesenchymal Stem Cell-Derived Extracellular Vesicles as Non-Coding RNA Therapeutic Vehicles in Autoimmune Diseases. Pharmaceutics 2022, 14, 733. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Vives, E.; Sole, C.; Moline, T.; Vidal, M.; Agraz, I.; Ordi-Ros, J.; Cortes-Hernandez, J. The Urinary Exosomal miRNA Expression Profile is Predictive of Clinical Response in Lupus Nephritis. Int. J. Mol. Sci. 2020, 21, 1372. [Google Scholar] [CrossRef] [Green Version]
- Tan, L.; Zhao, M.; Wu, H.; Zhang, Y.; Tong, X.; Gao, L.; Zhou, L.; Lu, Q.; Zeng, J. Downregulated Serum Exosomal miR-451a Expression Correlates with Renal Damage and Its Intercellular Communication Role in Systemic Lupus Erythematosus. Front. Immunol. 2021, 12, 630112. [Google Scholar] [CrossRef]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Villarroya-Beltri, C.; Gutierrez-Vazquez, C.; Sanchez-Cabo, F.; Perez-Hernandez, D.; Vazquez, J.; Martin-Cofreces, N.; Martinez-Herrera, D.J.; Pascual-Montano, A.; Mittelbrunn, M.; Sanchez-Madrid, F. Sumoylated hnRNPA2B1 controls the sorting of miRNAs into exosomes through binding to specific motifs. Nat. Commun. 2013, 4, 2980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Hernandez, J.; Riffo-Campos, A.L.; Ortega, A.; Martinez-Arroyo, O.; Perez-Gil, D.; Olivares, D.; Solaz, E.; Martinez, F.; Martinez-Hervas, S.; Chaves, F.J.; et al. Urinary- and Plasma-Derived Exosomes Reveal a Distinct MicroRNA Signature Associated with Albuminuria in Hypertension. Hypertension 2021, 77, 960–971. [Google Scholar] [CrossRef]
- Zhao, J.Y.; Wang, X.L.; Yang, Y.C.; Zhang, B.; Wu, Y.B. Upregulated miR-101 inhibits acute kidney injury-chronic kidney disease transition by regulating epithelial-mesenchymal transition. Hum. Exp. Toxicol. 2020, 39, 1628–1638. [Google Scholar] [CrossRef]
- Wang, Q.; Tao, Y.; Xie, H.; Liu, C.; Liu, P. MicroRNA-101 inhibits renal tubular epithelial-to-mesenchymal transition by targeting TGF-beta1 type I receptor. Int. J. Mol. Med. 2021, 47, 119. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Li, S.; Wang, Z.; Bai, N.; Feng, Y. miR-101-3p negatively regulates inflammation in systemic lupus erythematosus via MAPK1 targeting and inhibition of the NF-kappaB pathway. Mol. Med. Rep. 2021, 23, 359. [Google Scholar] [CrossRef] [PubMed]
- Connor, K.L.; Teenan, O.; Cairns, C.; Banwell, V.; Thomas, R.A.; Rodor, J.; Finnie, S.; Pius, R.; Tannahill, G.M.; Sahni, V.; et al. Identifying cell-enriched miRNAs in kidney injury and repair. JCI Insight 2020, 5, e140399. [Google Scholar] [CrossRef]
- Li, G.; Zhang, J.; Liu, D.; Wei, Q.; Wang, H.; Lv, Y.; Ye, Z.; Liu, G.; Li, L. Identification of Hub Genes and Potential ceRNA Networks of Diabetic Nephropathy by Weighted Gene Co-Expression Network Analysis. Front. Genet. 2021, 12, 767654. [Google Scholar] [CrossRef]
- Tamayo, E.; Alvarez, P.; Merino, R. TGFbeta Superfamily Members as Regulators of B Cell Development and Function-Implications for Autoimmunity. Int. J. Mol. Sci. 2018, 19, 3928. [Google Scholar] [CrossRef] [Green Version]
- Morianos, I.; Papadopoulou, G.; Semitekolou, M.; Xanthou, G. Activin-A in the regulation of immunity in health and disease. J. Autoimmun. 2019, 104, 102314. [Google Scholar] [CrossRef] [PubMed]
- El-Gendi, S.S.; Moniem, A.E.; Tawfik, N.M.; Ashmawy, M.M.; Mohammed, O.A.; Mostafa, A.K.; Zakhari, M.M.; Herdan, O.M. Value of serum and synovial fluid activin A and inhibin A in some rheumatic diseases. Int. J. Rheum. Dis. 2010, 13, 273–279. [Google Scholar] [CrossRef]
- Torricelli, M.; Bellisai, F.; Novembri, R.; Galeazzi, L.R.; Iuliano, A.; Voltolini, C.; Spreafico, A.; Galeazzi, M.; Petraglia, F. High levels of maternal serum IL-17 and activin A in pregnant women affected by systemic lupus erythematosus. Am. J. Reprod. Immunol. 2011, 66, 84–89. [Google Scholar] [CrossRef]
- Kadiombo, A.T.; Maeshima, A.; Kayakabe, K.; Ikeuchi, H.; Sakairi, T.; Kaneko, Y.; Hiromura, K.; Nojima, Y. Involvement of infiltrating macrophage-derived activin A in the progression of renal damage in MRL-lpr mice. Am. J. Physiol. Renal Physiol. 2017, 312, F297–F304. [Google Scholar] [CrossRef] [Green Version]
- Hill, G.S.; Delahousse, M.; Nochy, D.; Mandet, C.; Bariety, J. Proteinuria and tubulointerstitial lesions in lupus nephritis. Kidney Int. 2001, 60, 1893–1903. [Google Scholar] [CrossRef] [Green Version]
- Pamfil, C.; Makowska, Z.; De Groof, A.; Tilman, G.; Babaei, S.; Galant, C.; Montigny, P.; Demoulin, N.; Jadoul, M.; Aydin, S.; et al. Intrarenal activation of adaptive immune effectors is associated with tubular damage and impaired renal function in lupus nephritis. Ann. Rheum. Dis. 2018, 77, 1782–1789. [Google Scholar] [CrossRef]
- Pal, M.; Bhattacharya, S.; Kalyan, G.; Hazra, S. Cadherin profiling for therapeutic interventions in Epithelial Mesenchymal Transition (EMT) and tumorigenesis. Exp. Cell Res. 2018, 368, 137–146. [Google Scholar] [CrossRef]
- Liu, Q.; Du, Y.; Li, K.; Zhang, W.; Feng, X.; Hao, J.; Li, H.; Liu, S. Anti-OSM Antibody Inhibits Tubulointerstitial Lesion in a Murine Model of Lupus Nephritis. Mediat. Inflamm. 2017, 2017, 3038514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, D.; Senouthai, S.; Wang, J.; You, Y. FKN Facilitates HK-2 Cell EMT and Tubulointerstitial Lesions via the Wnt/beta-Catenin Pathway in a Murine Model of Lupus Nephritis. Front. Immunol. 2019, 10, 784. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Liu, J.; Zhang, X.; Chen, Q.; Bai, X.; Hong, X.; Zhou, L.; Liu, Y. Role of miRNA-671-5p in Mediating Wnt/beta-Catenin-Triggered Podocyte Injury. Front. Pharmacol. 2021, 12, 784489. [Google Scholar] [CrossRef] [PubMed]
- Lennon, R.; Randles, M.J.; Humphries, M.J. The importance of podocyte adhesion for a healthy glomerulus. Front. Endocrinol. 2014, 5, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asano-Matsuda, K.; Ibrahim, S.; Takano, T.; Matsuda, J. Role of Rho GTPase Interacting Proteins in Subcellular Compartments of Podocytes. Int. J. Mol. Sci. 2021, 22, 3656. [Google Scholar] [CrossRef] [PubMed]
- Komiya, Y.; Onodera, Y.; Kuroiwa, M.; Nomimura, S.; Kubo, Y.; Nam, J.M.; Kajiwara, K.; Nada, S.; Oneyama, C.; Sabe, H.; et al. The Rho guanine nucleotide exchange factor ARHGEF5 promotes tumor malignancy via epithelial-mesenchymal transition. Oncogenesis 2016, 5, e258. [Google Scholar] [CrossRef] [Green Version]
- Rogg, M.; Maier, J.I.; Van Wymersch, C.; Helmstadter, M.; Sammarco, A.; Lindenmeyer, M.; Zareba, P.; Montanez, E.; Walz, G.; Werner, M.; et al. alpha-Parvin Defines a Specific Integrin Adhesome to Maintain the Glomerular Filtration Barrier. J. Am. Soc. Nephrol. 2022, 33, 786–808. [Google Scholar] [CrossRef]
- Takenawa, T.; Suetsugu, S. The WASP-WAVE protein network: Connecting the membrane to the cytoskeleton. Nat. Rev. Mol. Cell Biol. 2007, 8, 37–48. [Google Scholar] [CrossRef]
- Iwasaki, Y.W.; Siomi, M.C.; Siomi, H. PIWI-Interacting RNA: Its Biogenesis and Functions. Annu. Rev. Biochem. 2015, 84, 405–433. [Google Scholar] [CrossRef] [PubMed]
- Floege, J.; Barbour, S.J.; Cattran, D.C.; Hogan, J.J.; Nachman, P.H.; Tang, S.C.W.; Wetzels, J.F.M.; Cheung, M.; Wheeler, D.C.; Winkelmayer, W.C.; et al. Management and treatment of glomerular diseases (part 1): Conclusions from a Kidney Disease: Improving Global Outcomes (KDIGO) Controversies Conference. Kidney Int. 2019, 95, 268–280. [Google Scholar] [CrossRef] [Green Version]
- Rovin, B.H.; Caster, D.J.; Cattran, D.C.; Gibson, K.L.; Hogan, J.J.; Moeller, M.J.; Roccatello, D.; Cheung, M.; Wheeler, D.C.; Winkelmayer, W.C.; et al. Management and treatment of glomerular diseases (part 2): Conclusions from a Kidney Disease: Improving Global Outcomes (KDIGO) Controversies Conference. Kidney Int. 2019, 95, 281–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Fukunaga, T.; Iwakiri, J.; Ono, Y.; Hamada, M. LncRRIsearch: A Web Server for lncRNA-RNA Interaction Prediction Integrated With Tissue-Specific Expression and Subcellular Localization Data. Front. Genet. 2019, 10, 462. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Wang, J.; Jaehnig, E.J.; Shi, Z.; Zhang, B. WebGestalt 2019: Gene set analysis toolkit with revamped UIs and APIs. Nucleic Acids Res. 2019, 47, W199–W205. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Non LN (n = 73) | LN (n = 23) | CNT (n = 25) |
|---|---|---|---|
| Age, years | 51.9 ± 11.8 | 48.7 ± 9.1 | 40.2 ± 12.0 |
| Female sex. (%) | 82.6 | 87.2 | 60.7 |
| Serum creatinine (mg/dL) | 1.6 ± 7.3 | 0.8 ± 0.3 | 0.71 ± 0.11 |
| dsDNA (U/mL) | 69.0 ± 92.6 | 200.7 ± 289.2 ** | - |
| ANA | 8.6 ± 4.7 | 8.2 ± 5.3 | - |
| C3 (mg/dL) | 100.1 ± 26.3 | 87.6 ± 29.6 | - |
| C4 (mg/dL) | 20.8 ± 12.4 | 16.2 ± 10.5 | - |
| SLEDAI index | 5.3 ± 5.3 | 14.3 ± 6.6 ** | - |
| eGFR (CKD-EPI) | 98.2 ± 14.4 | 84.5 ± 30.1 ** | 105.9 ± 12.5 |
| Log urinary albumin/urinary creatinine | 0.62 (0.39–0.89) | 2.12 (1.88–2.48) ** | 0.58 (0.44–0.72) |
| Basal Treatment (%) | |||
| Hydroxychloroquine | 68.5 | 56.5 | |
| Azathioprine | 19.2 | 17.4 | |
| Prednisone | 38.4 | 47.8 | |
| Methotrexate | 6.8 | 4.3 | |
| Deflazacort | 12.3 | 9.3 | |
| Immunosuppressive therapy | 3.7 | 9.0 | |
| Monoclonal antibodies | 4.1 | 13.0 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flores-Chova, A.; Martinez-Arroyo, O.; Riffo-Campos, A.L.; Ortega, A.; Forner, M.J.; Cortes, R. Plasma Exosomal Non-Coding RNA Profile Associated with Renal Damage Reveals Potential Therapeutic Targets in Lupus Nephritis. Int. J. Mol. Sci. 2023, 24, 7088. https://doi.org/10.3390/ijms24087088
Flores-Chova A, Martinez-Arroyo O, Riffo-Campos AL, Ortega A, Forner MJ, Cortes R. Plasma Exosomal Non-Coding RNA Profile Associated with Renal Damage Reveals Potential Therapeutic Targets in Lupus Nephritis. International Journal of Molecular Sciences. 2023; 24(8):7088. https://doi.org/10.3390/ijms24087088
Chicago/Turabian StyleFlores-Chova, Ana, Olga Martinez-Arroyo, Angela L. Riffo-Campos, Ana Ortega, Maria J. Forner, and Raquel Cortes. 2023. "Plasma Exosomal Non-Coding RNA Profile Associated with Renal Damage Reveals Potential Therapeutic Targets in Lupus Nephritis" International Journal of Molecular Sciences 24, no. 8: 7088. https://doi.org/10.3390/ijms24087088