
EjFAD8 Enhances the Low-Temperature Tolerance of Loquat by Desaturation of Sulfoquinovosyl Diacylglycerol (SQDG)

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
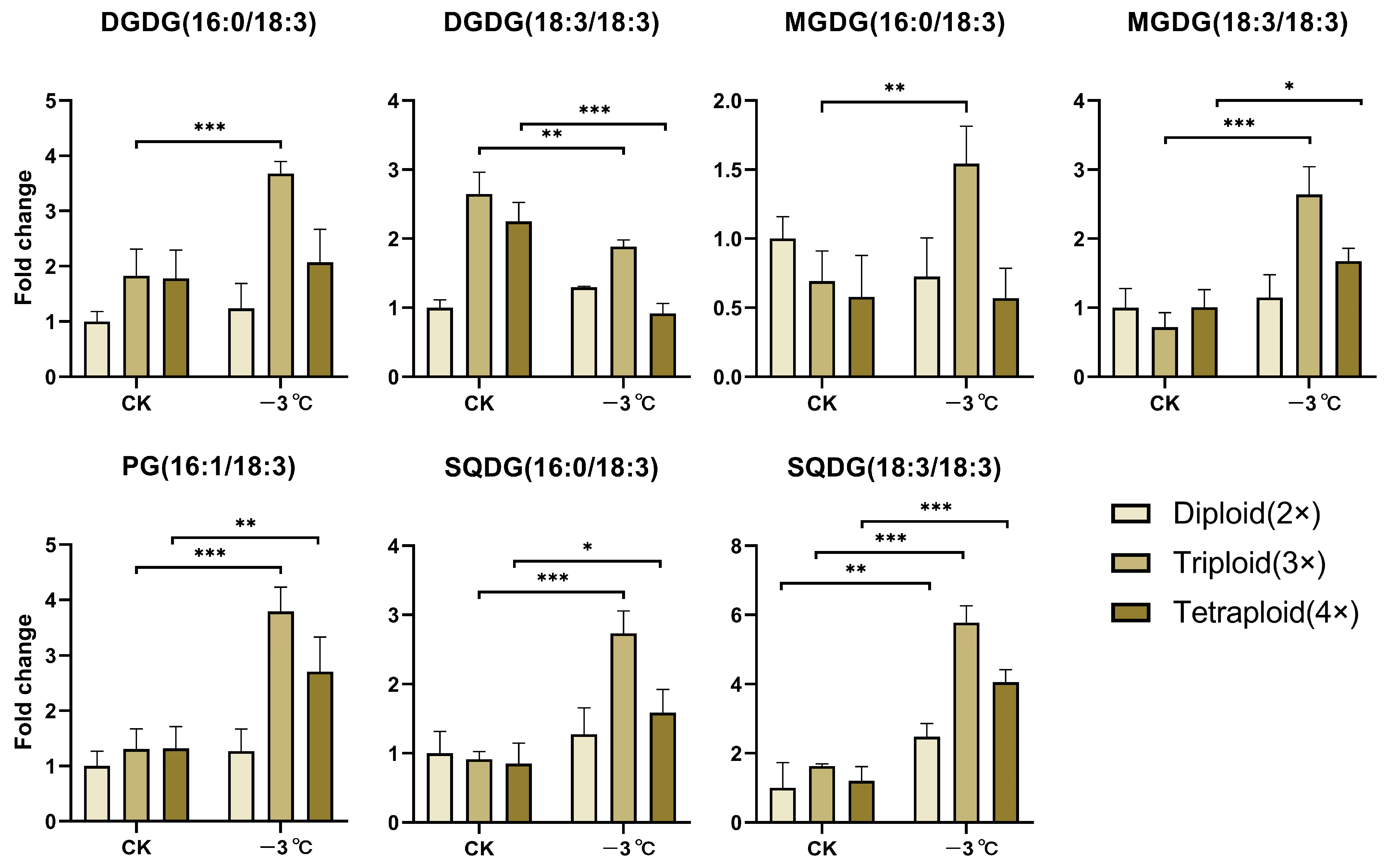
2.1. EjFAD8 Plays an Important Role in Lipid Metabolism in the Triploid Loquat at Low Temperatures
2.2. Characterization of EjFAD8 from Loquat
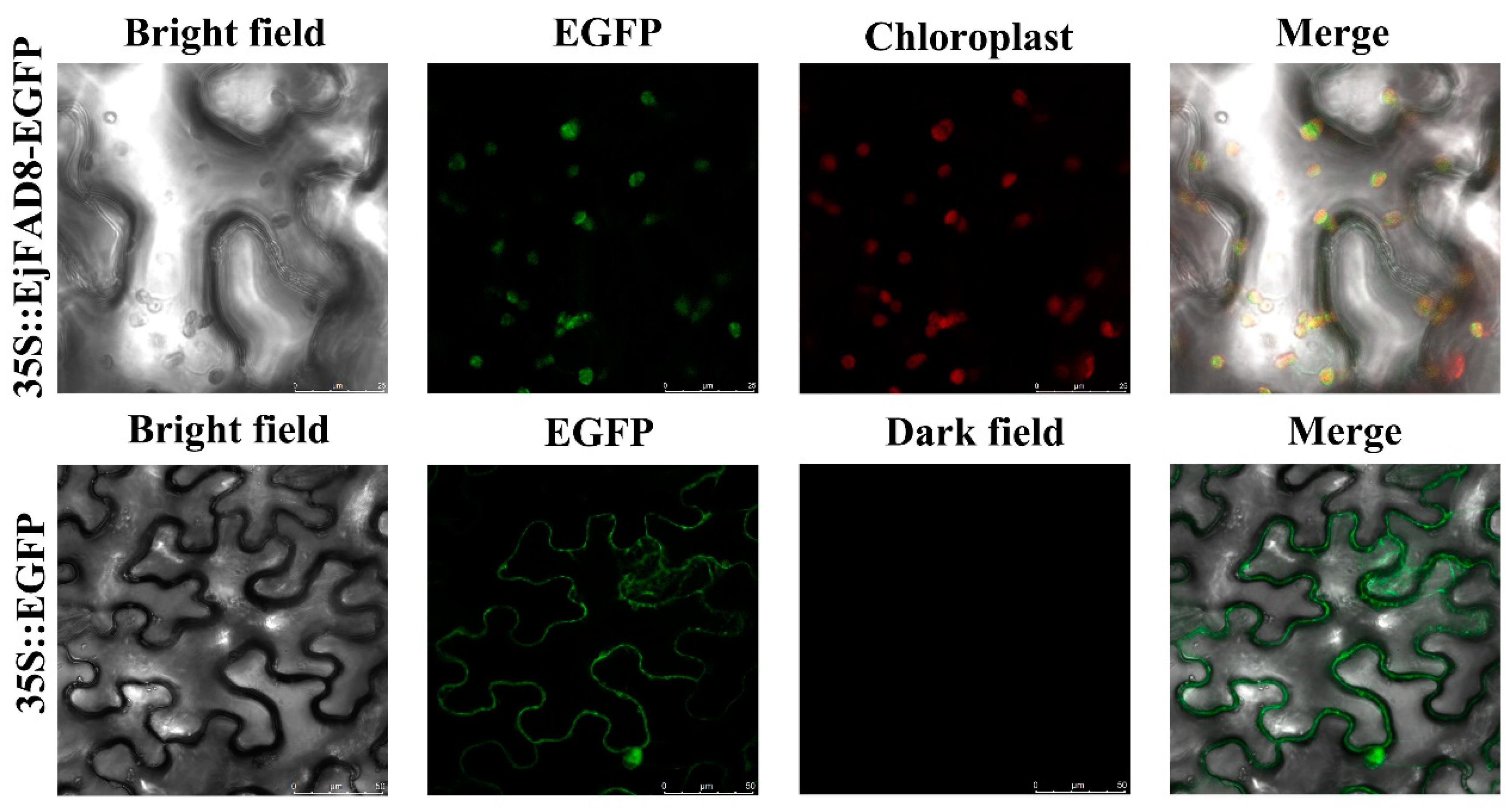
2.3. Subcellular Localization of EjFAD8
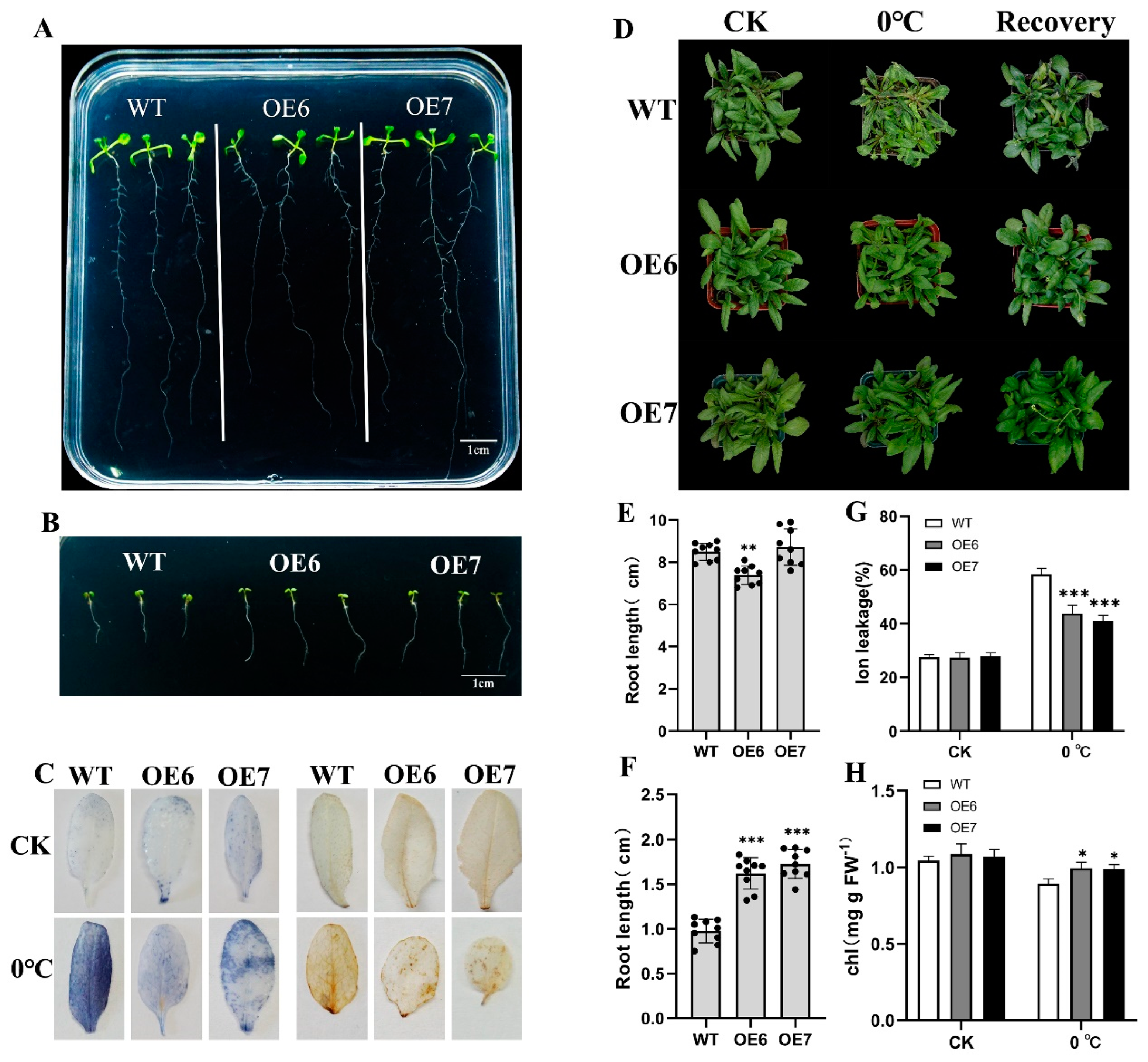
2.4. Overexpression of EjFAD8 Enhances Low-Temperature Tolerance in Arabidopsis
2.5. Overexpression of EjFAD8 under Low-Temperature Stress Increases Transcription of Low-Temperature-Tolerant Genes in Transgenic Arabidopsis
2.6. FAD8-Related Lipids Are Increased in Transgenic Arabidopsis under Low-Temperature Stress
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Cloning and Sequence Analysis of EjFAD8
4.3. Arabidopsis Transformation
4.4. Subcellular Localization of EjFAD8
4.5. Total RNA Extraction and Quantitative Real-Time PCR Analysis
4.6. Low-Temperature Stress Treatments
4.7. Measurement of the Content of ROS and Chlorophyll
4.8. RNA-Seq Analysis
4.9. Untargeted Relative Quantitative Lipidomics Analysis
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gong, R.G.; Lai, J.; Yang, W.; Liao, M.A.; Wang, Z.H.; Liang, G.L. Analysis of Alterations to the Transcriptome of Loquat (Eriobotrya japonica Lindl.) under Low Temperature Stress via de Novo Sequencing. Genet. Mol. Res. 2015, 14, 9423–9436. [Google Scholar] [CrossRef]
- Adiletta, G.; Pasquariello, M.; Zampella, L.; Mastrobuoni, F.; Scortichini, M.; Petriccione, M. Chitosan Coating: A Postharvest Treatment to Delay Oxidative Stress in Loquat Fruits during Cold Storage. Agronomy 2018, 8, 54. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, S.; Nagano, Y.; Matsuguma, K.; Ishimoto, K.; Hiehata, N.; Nagano, A.J.; Tezuka, A.; Yamamoto, T. Construction of a High-Density Linkage Map for Bronze Loquat Using RAD-Seq. Sci. Hortic. 2019, 251, 59–64. [Google Scholar] [CrossRef]
- Zhang, J.; An, H.; Zhang, X.; Xu, F.; Zhou, B. Transcriptomic Analysis Reveals Potential Gene Regulatory Networks Under Cold Stress of Loquat (Eriobotrya Japonica Lindl.). Front. Plant Sci. 2022, 13, 944269. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.; Wang, Y.; Tao, L.; Zhang, H.; Deng, Q.; Yang, Z.; Chi, Z.; Yang, Y. Single-Molecule Real-Time Sequencing of the Full-Length Transcriptome of Loquat under Low-Temperature Stress. PLoS ONE 2020, 15, e0238942. [Google Scholar] [CrossRef]
- Ma, X.; Chen, C.; Yang, M.; Dong, X.; Lv, W.; Meng, Q. Cold-Regulated Protein (SlCOR413IM1) Confers Chilling Stress Tolerance in Tomato Plants. Plant Physiol. Biochem. 2018, 124, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Mironov, K.S.; Sidorov, R.A.; Trofimova, M.S.; Bedbenov, V.S.; Tsydendambaev, V.D.; Allakhverdiev, S.I.; Los, D.A. Light-Dependent Cold-Induced Fatty Acid Unsaturation, Changes in Membrane Fluidity, and Alterations in Gene Expression in Synechocystis. Biochim. Et Biophys. Acta (BBA)-Bioenerg. 2012, 1817, 1352–1359. [Google Scholar] [CrossRef] [Green Version]
- Dhaliwal, L.K.; Angeles-Shim, R.B. Cell Membrane Features as Potential Breeding Targets to Improve Cold Germination Ability of Seeds. Plants 2022, 11, 3400. [Google Scholar] [CrossRef]
- Kenchanmane Raju, S.K.; Barnes, A.C.; Schnable, J.C.; Roston, R.L. Low-Temperature Tolerance in Land Plants: Are Transcript and Membrane Responses Conserved? Plant Sci. 2018, 276, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Valitova, J.; Renkova, A.; Mukhitova, F.; Dmitrieva, S.; Beckett, R.P.; Minibayeva, F.V. Membrane Sterols and Genes of Sterol Biosynthesis Are Involved in the Response of Triticum Aestivum Seedlings to Cold Stress. Plant Physiol. Biochem. 2019, 142, 452–459. [Google Scholar] [CrossRef]
- de Dios Barajas-Lopez, J.; Tiwari, A.; Zarza, X.; Shaw, M.W.; Pascual, J.; Punkkinen, M.; Bakowska, J.C.; Munnik, T.; Fujii, H. EARLY RESPONSE TO DEHYDRATION 7 Remodels Cell Membrane Lipid Composition during Cold Stress in Arabidopsis. Plant Cell Physiol. 2021, 62, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Halim, N.F.A.A.; Ali, M.S.M.; Leow, A.T.C.; Rahman, R.N.Z.R.A. Membrane Fatty Acid Desaturase: Biosynthesis, Mechanism, and Architecture. Appl. Microbiol. Biotechnol. 2022, 106, 5957–5972. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Liu, F.; Sun, X.; Wang, B.; Liu, J.; Ni, X.; Hu, C.; Deng, G.; Tong, Z.; Zhang, Y.; et al. Genome-Wide Identification of FAD Gene Family and Their Contributions to the Temperature Stresses and Mutualistic and Parasitic Fungi Colonization Responses in Banana. Int. J. Biol. Macromol. 2022, 204, 661–676. [Google Scholar] [CrossRef]
- Dyer, J.M.; Mullen, R.T. Immunocytological Localization of Two Plant Fatty Acid Desaturases in the Endoplasmic Reticulum. FEBS Lett. 2001, 494, 44–47. [Google Scholar] [CrossRef]
- Luján, M.A.; Soria-García, Á.; Claver, A.; Lorente, P.; Rubio, M.C.; Picorel, R.; Alfonso, M. Different Cis-Regulatory Elements Control the Tissue-Specific Contribution of Plastid ω-3 Desaturases to Wounding and Hormone Responses. Front. Plant Sci. 2021, 12, 727292. [Google Scholar] [CrossRef]
- Soria-García, Á.; Rubio, M.C.; Lagunas, B.; López-Gomollón, S.; de los Luján, M.; Díaz-Guerra, R.; Picorel, R.; Alfonso, M. Tissue Distribution and Specific Contribution of Arabidopsis FAD7 and FAD8 Plastid Desaturases to the JA- and ABA-Mediated Cold Stress or Defense Responses. Plant Cell Physiol. 2019, 60, 1025–1040. [Google Scholar] [CrossRef] [Green Version]
- Gibson, S.; Arondel, V.; Iba, K.; Somerville, C. Cloning of a Temperature-Regulated Gene Encoding a Chloroplast ω-3 Desaturase from Arabidopsis thaliana. Plant Physiol. 1994, 106, 7. [Google Scholar] [CrossRef] [Green Version]
- Román, Á.; Hernández, M.L.; Soria-García, Á.; López-Gomollón, S.; Lagunas, B.; Picorel, R.; Martínez-Rivas, J.M.; Alfonso, M. Non-Redundant Contribution of the Plastidial FAD8 ω-3 Desaturase to Glycerolipid Unsaturation at Different Temperatures in Arabidopsis. Mol. Plant 2015, 8, 1599–1611. [Google Scholar] [CrossRef] [Green Version]
- Gopalakrishnan Nair, P.M.; Kang, I.-S.; Moon, B.-Y.; Lee, C.-H. Effects of Low Temperature Stress on Rice (Oryza Sativa L.) Plastid ω-3 Desaturase Gene, OsFAD8 and Its Functional Analysis Using T-DNA Mutants. Plant Cell Tissue Organ Cult. 2009, 98, 87–96. [Google Scholar] [CrossRef]
- Zhao, X.; Wei, J.; He, L.; Zhang, Y.; Zhao, Y.; Xu, X.; Wei, Y.; Ge, S.; Ding, D.; Liu, M.; et al. Identification of Fatty Acid Desaturases in Maize and Their Differential Responses to Low and High Temperature. Genes 2019, 10, 445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Ding, Y.; Yang, S. Molecular Regulation of CBF Signaling in Cold Acclimation. Trends Plant Sci. 2018, 23, 623–637. [Google Scholar] [CrossRef]
- Vyse, K.; Pagter, M.; Zuther, E.; Hincha, D.K. Deacclimation after Cold Acclimation—A Crucial, but Widely Neglected Part of Plant Winter Survival. J. Exp. Bot. 2019, 70, 4595–4604. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Jiang, L.; Wang, F.; Yu, D. Jasmonate Regulates the INDUCER OF CBF EXPRESSION–C-REPEAT BINDING FACTOR/DRE BINDING FACTOR1 Cascade and Freezing Tolerance in Arabidopsis. Plant Cell 2013, 25, 2907–2924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopeć, P.; Rapacz, M.; Arora, R. Post-Translational Activation of CBF for Inducing Freezing Tolerance. Trends Plant Sci. 2022, 27, 415–417. [Google Scholar] [CrossRef]
- Yang, J.; Guo, X.; Mei, Q.; Qiu, L.; Chen, P.; Li, W.; Mao, K.; Ma, F. MdbHLH4 Negatively Regulates Apple Cold Tolerance by Inhibiting MdCBF1/3 Expression and Promoting MdCAX3L-2 Expression. Plant Physiol. 2022, 191, kiac512. [Google Scholar] [CrossRef]
- Gray, G.R.; Chauvin, L.P.; Sarhan, F.; Huner, N.P.A. Cold Acclimation and Freezing Tolerance (A Complex Interaction of Light and Temperature). Plant Physiol. 1997, 114, 467–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huner, N.P.A.; Öquist, G.; Sarhan, F. Energy Balance and Acclimation to Light and Cold. Trends Plant Sci. 1998, 3, 224–230. [Google Scholar] [CrossRef]
- Li, C.; Sun, Y.; Li, J.; Zhang, T.; Zhou, F.; Song, Q.; Liu, Y.; Brestic, M.; Chen, T.H.H.; Yang, X. ScCBF1 Plays a Stronger Role in Cold, Salt and Drought Tolerance than StCBF1 in Potato (Solanum tuberosum). J. Plant Physiol. 2022, 278, 153806. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wang, H.; Zhang, Y.; Zhang, B.; Liu, S.; Li, Y.; Dang, J.; He, Q.; Liang, G.; Guo, Q. Comparative Analysis of the Photosystem Activity of Triploid Loquats and Their Tetraploid, Diploid Parents. Acta Hortic. Sin. 2021, 48, 37–48. [Google Scholar] [CrossRef]
- Li, Q.; Zheng, Q.; Shen, W.; Cram, D.; Fowler, D.B.; Wei, Y.; Zou, J. Understanding the Biochemical Basis of Temperature-Induced Lipid Pathway Adjustments in Plants. Plant Cell 2015, 27, 86–103. [Google Scholar] [CrossRef] [Green Version]
- Narayanan, S.; Prasad, P.V.V.; Welti, R. Wheat Leaf Lipids during Heat Stress: II. Lipids Experiencing Coordinated Metabolism Are Detected by Analysis of Lipid Co-occurrence. Plant Cell Environ. 2016, 39, 608–617. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Kong, X.; Huang, H.; Wu, W.; Park, J.; Yun, D.-J.; Lee, B.; Shi, H.; Zhu, J.-K. STCH4/REIL2 Confers Cold Stress Tolerance in Arabidopsis by Promoting RRNA Processing and CBF Protein Translation. Cell Rep. 2020, 30, 229–242.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimojima, M. Biosynthesis and Functions of the Plant Sulfolipid. Prog. Lipid Res. 2011, 50, 234–239. [Google Scholar] [CrossRef]
- Gan, P.; Liu, F.; Li, R.; Wang, S.; Luo, J. Chloroplasts— Beyond Energy Capture and Carbon Fixation: Tuning of Photosynthesis in Response to Chilling Stress. IJMS 2019, 20, 5046. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Benning, C. Anionic Lipids Are Required for Chloroplast Structure and Function in Arabidopsis. Plant J. 2003, 36, 762–770. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Wada, H. Role of Lipids in Chloroplast Biogenesis. In Lipids in Plant and Algae Development; Nakamura, Y., Li-Beisson, Y., Eds.; Subcellular Biochemistry; Springer International Publishing: Cham, Switzerland, 2016; Volume 86, pp. 103–125. ISBN 978-3-319-25977-2. [Google Scholar]
- Kurepin, L.; Dahal, K.; Savitch, L.; Singh, J.; Bode, R.; Ivanov, A.; Hurry, V.; Hüner, N. Role of CBFs as Integrators of Chloroplast Redox, Phytochrome and Plant Hormone Signaling during Cold Acclimation. IJMS 2013, 14, 12729–12763. [Google Scholar] [CrossRef]
- Kim, Y.; Park, S.-U.; Shin, D.-M.; Pham, G.; Jeong, Y.S.; Kim, S.-H. Corrigendum: ATBS1-INTERACTING FACTOR 2 Negatively Regulates Dark- and Brassinosteroid-Induced Leaf Senescence through Interactions with INDUCER OF CBF EXPRESSION 1. J. Exp. Bot. 2020, 71, 2210. [Google Scholar] [CrossRef] [Green Version]
- Fowler, S.G.; Cook, D.; Thomashow, M.F. Low Temperature Induction of Arabidopsis CBF1, 2, and 3 Is Gated by the Circadian Clock. Plant Physiol. 2005, 137, 961–968. [Google Scholar] [CrossRef] [Green Version]
- Sandve, S.R.; Kosmala, A.; Rudi, H.; Fjellheim, S.; Rapacz, M.; Yamada, T.; Rognli, O.A. Molecular Mechanisms Underlying Frost Tolerance in Perennial Grasses Adapted to Cold Climates. Plant Sci. 2011, 180, 69–77. [Google Scholar] [CrossRef]
- Suzuki, N.; Mittler, R. Reactive Oxygen Species and Temperature Stresses: A Delicate Balance between Signaling and Destruction. Physiol. Plant. 2006, 126, 45–51. [Google Scholar] [CrossRef]
- Lee, C.-J.; Kim, S.-E.; Park, S.-U.; Lim, Y.-H.; Ji, C.Y.; Jo, H.; Lee, J.-D.; Yoon, U.-H.; Kim, H.S.; Kwak, S.-S. Overexpression of IbFAD8 Enhances the Low-Temperature Storage Ability and Alpha-Linolenic Acid Content of Sweetpotato Tuberous Roots. Front. Plant Sci. 2021, 12, 764100. [Google Scholar] [CrossRef] [PubMed]
- Mihailova, G.; Kocheva, K.; Goltsev, V.; Kalaji, H.M.; Georgieva, K. Application of a Diffusion Model to Measure Ion Leakage of Resurrection Plant Leaves Undergoing Desiccation. Plant Physiol. Biochem. 2018, 125, 185–192. [Google Scholar] [CrossRef]
- Jing, D.; Liu, X.; He, Q.; Dang, J.; Hu, R.; Xia, Y.; Wu, D.; Wang, S.; Zhang, Y.; Xia, Q.; et al. Genome Assembly of Wild Loquat (Eriobotrya japonica) and Resequencing Provide New Insights into the Genomic Evolution and Fruit Domestication in Loquat. Hortic. Res. 2023, 10, uhac265. [Google Scholar] [CrossRef]
- Jiang, S.; An, H.; Xu, F.; Zhang, X. Chromosome-Level Genome Assembly and Annotation of the Loquat (Eriobotrya japonica) Genome. GigaScience 2020, 9, giaa015. [Google Scholar] [CrossRef] [Green Version]
- Oguz, B. Genetic Characterization of Toxocara Vitilorum in Turkey by Mitochondrial Gene Markers (Cox1). Acta Sci. Vet. 2018, 46, 6. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral Dip: A Simplified Method ForAgrobacterium-Mediated Transformation Of Arabidopsis Thaliana: Floral Dip Transformation of Arabidopsis. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Sparkes, I.A.; Runions, J.; Kearns, A.; Hawes, C. Rapid, Transient Expression of Fluorescent Fusion Proteins in Tobacco Plants and Generation of Stably Transformed Plants. Nat. Protoc. 2006, 1, 2019–2025. [Google Scholar] [CrossRef] [PubMed]
- Aragón, C.; Carvalho, L.; González, J.; Escalona, M.; Amâncio, S. Ex Vitro Acclimatization of Plantain Plantlets Micropropagated in Temporary Immersion Bioreactor. Biol. Plant. 2010, 54, 237–244. [Google Scholar] [CrossRef] [Green Version]
- Romero-Puertas, M.C.; Rodriguez-Serrano, M.; Corpas, F.J.; Gomez, M.; Del Rio, L.A.; Sandalio, L.M. Cadmium-Induced Subcellular Accumulation of O2.- and H2O2 in Pea Leaves. Plant Cell Env. 2004, 27, 1122–1134. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of Total Carotenoids and Chlorophylls a and b of Leaf Extracts in Different Solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; Yang, H.; Suo, X.; Liu, M.; Jing, D.; Zhang, Y.; Dang, J.; Wu, D.; He, Q.; Xia, Y.; et al. EjFAD8 Enhances the Low-Temperature Tolerance of Loquat by Desaturation of Sulfoquinovosyl Diacylglycerol (SQDG). Int. J. Mol. Sci. 2023, 24, 6946. https://doi.org/10.3390/ijms24086946
Xu X, Yang H, Suo X, Liu M, Jing D, Zhang Y, Dang J, Wu D, He Q, Xia Y, et al. EjFAD8 Enhances the Low-Temperature Tolerance of Loquat by Desaturation of Sulfoquinovosyl Diacylglycerol (SQDG). International Journal of Molecular Sciences. 2023; 24(8):6946. https://doi.org/10.3390/ijms24086946
Chicago/Turabian StyleXu, Xun, Hao Yang, Xiaodong Suo, Mingxiu Liu, Danlong Jing, Yin Zhang, Jiangbo Dang, Di Wu, Qiao He, Yan Xia, and et al. 2023. "EjFAD8 Enhances the Low-Temperature Tolerance of Loquat by Desaturation of Sulfoquinovosyl Diacylglycerol (SQDG)" International Journal of Molecular Sciences 24, no. 8: 6946. https://doi.org/10.3390/ijms24086946