
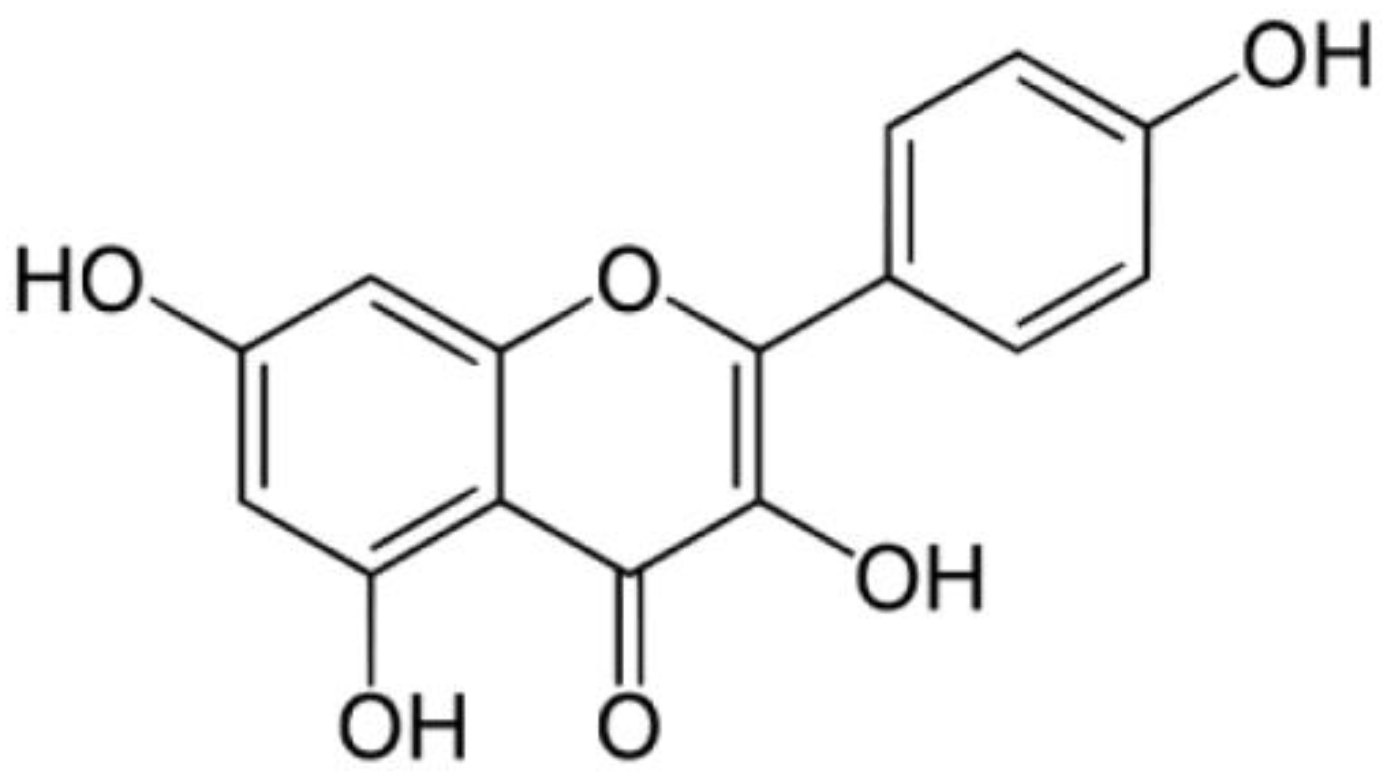
Kaempferol Suppresses Carbon Tetrachloride-Induced Liver Damage in Rats via the MAPKs/NF-κB and AMPK/Nrf2 Signaling Pathways


Abstract
:1. Introduction
2. Results
2.1. Antioxidative Activities of Kaempferol against FeSO4/H2O2-Induced Lipid Peroxidation and DPPH Radicals
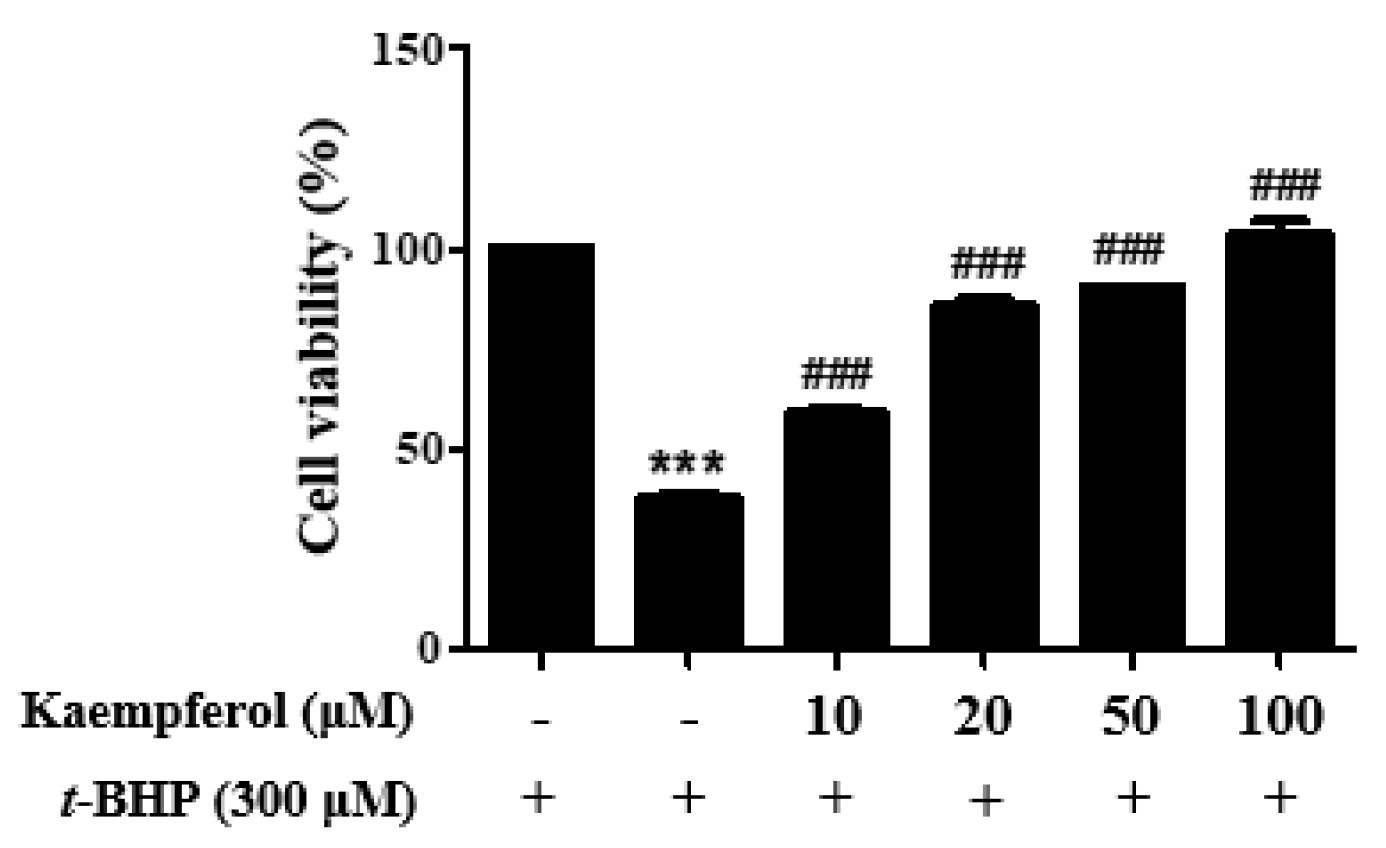
2.2. Protective Effect of Kaempferol on t-Butyl Hydroperoxide (t-BHP)-Induced HepG2 Cell Damage
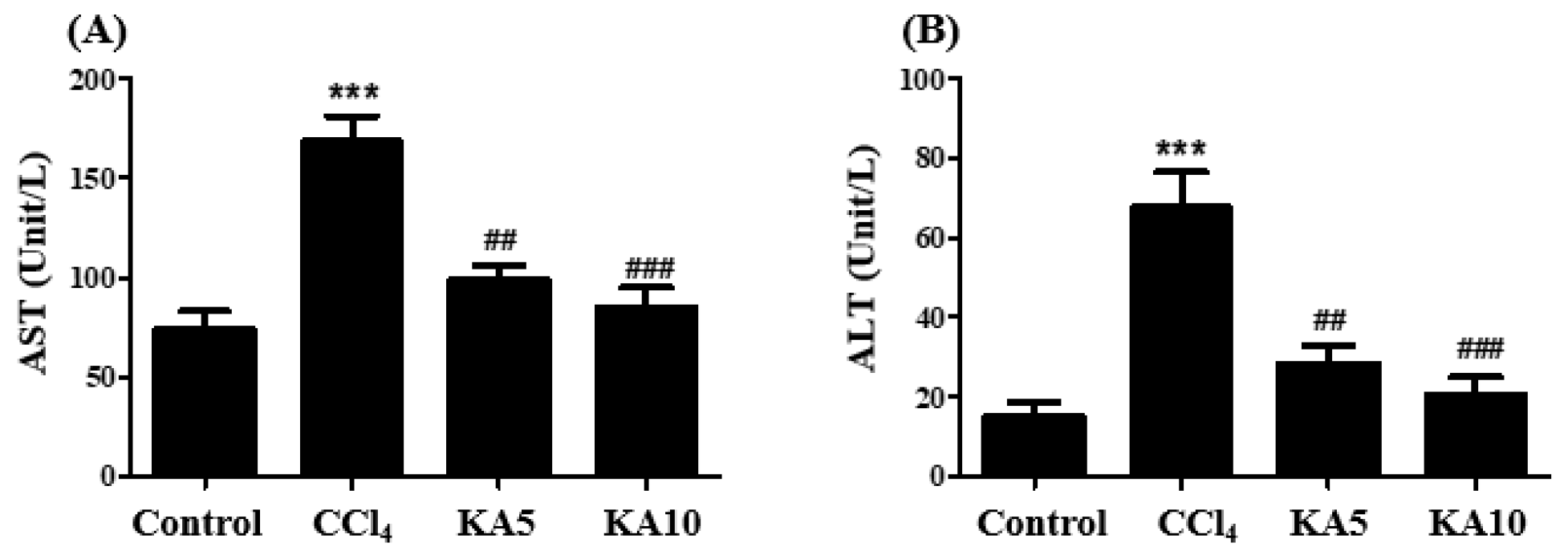
2.3. Changes in Body and Liver Weights and Serum Parameters in CCl4-Intoxicated Rats
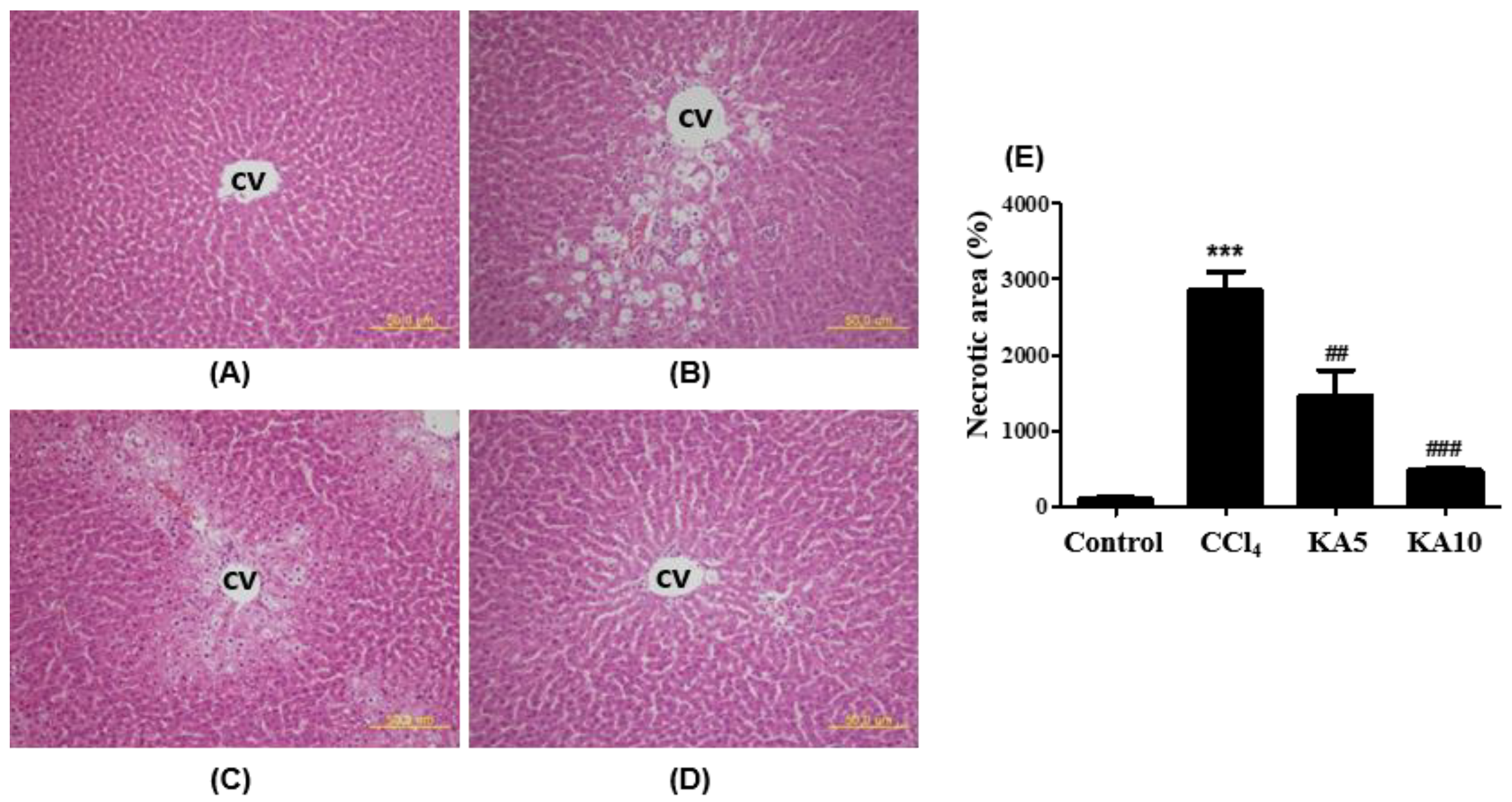
2.4. Liver Histopathology
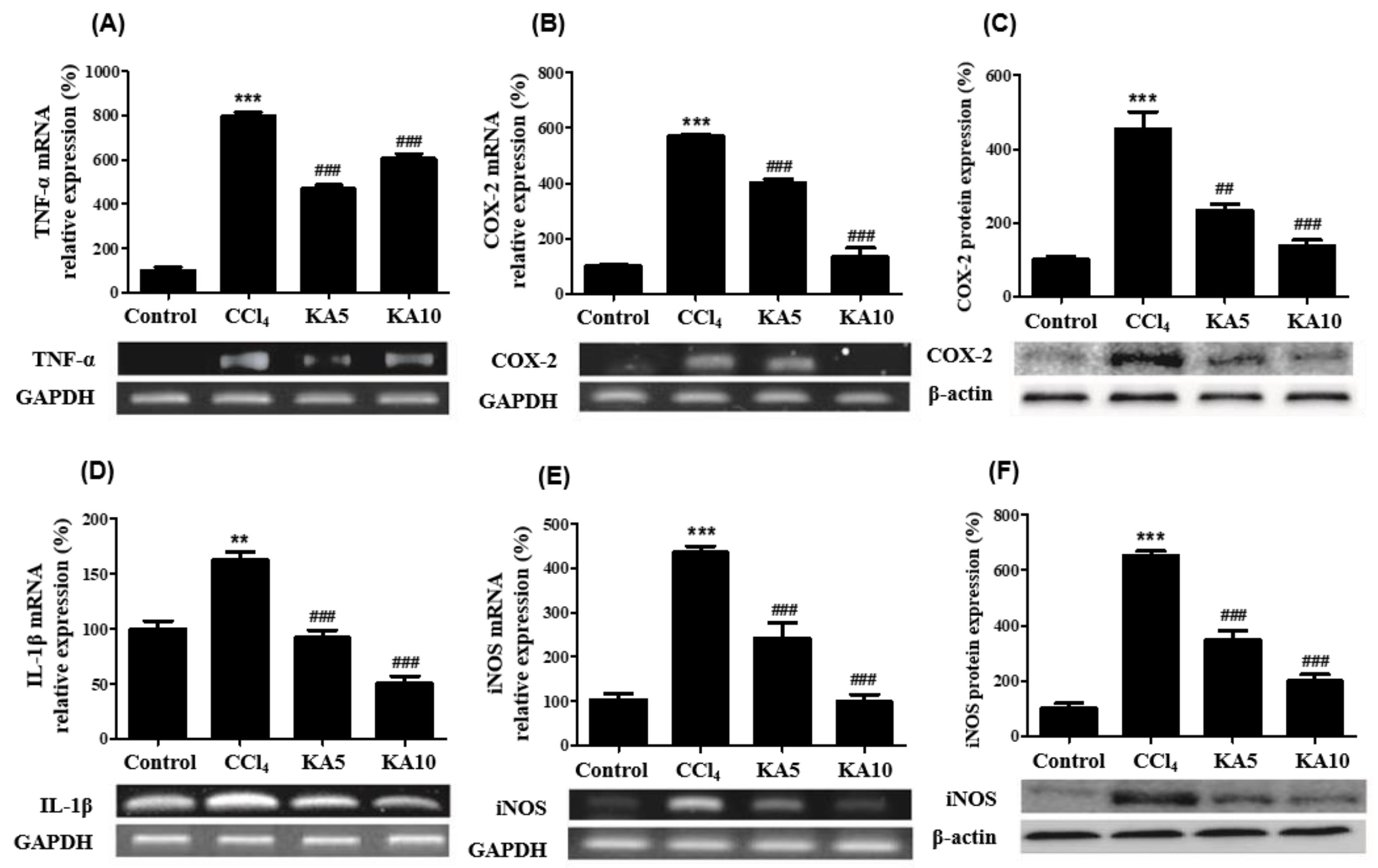
2.5. Effects of Kaempferol on CCl4-Intoxicated Liver Inflammatory Mediators
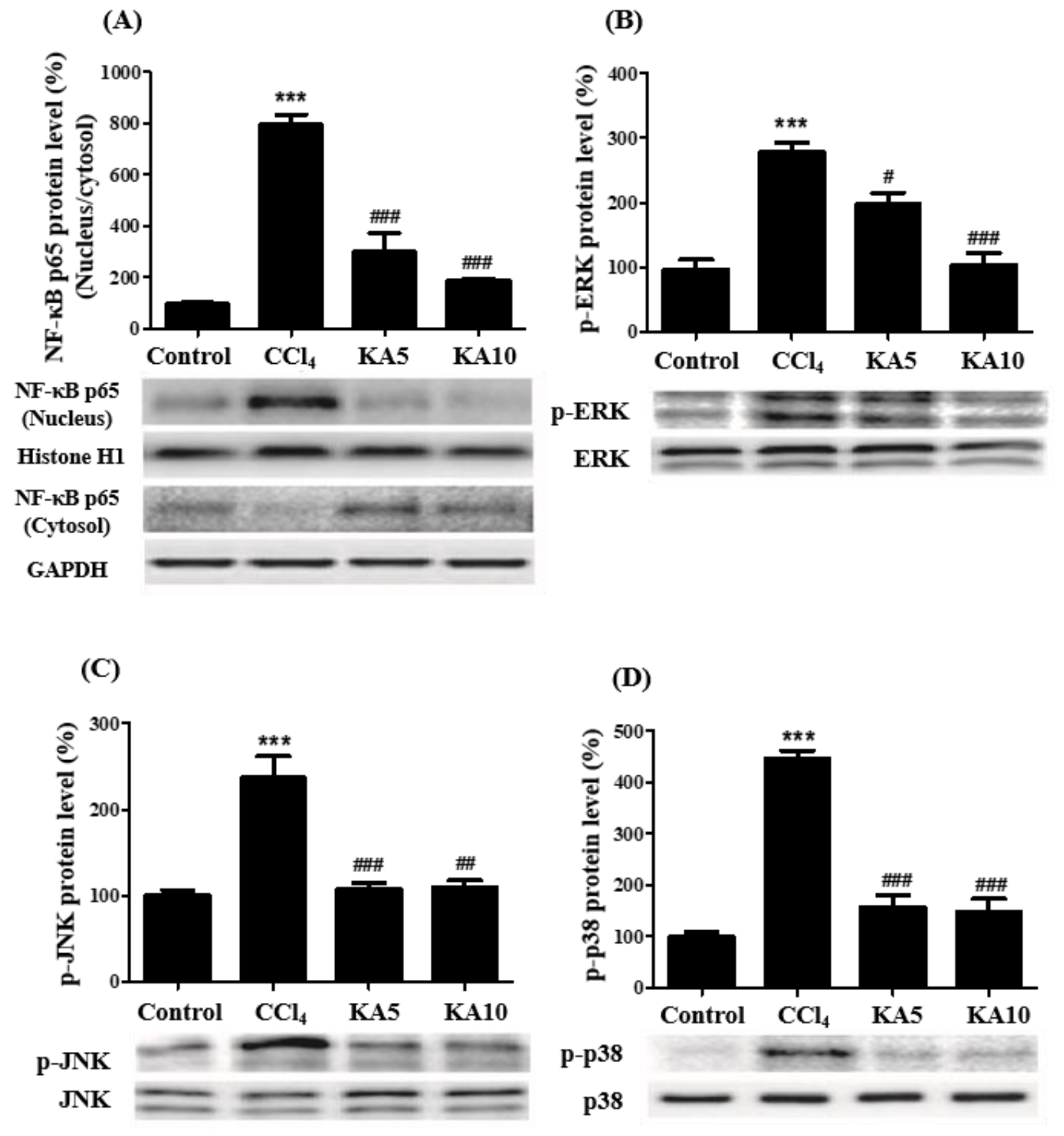
2.6. Effects of Kaempferol on MAPK/NF-κB Signaling Pathway in CCl4-Induced Liver Damages
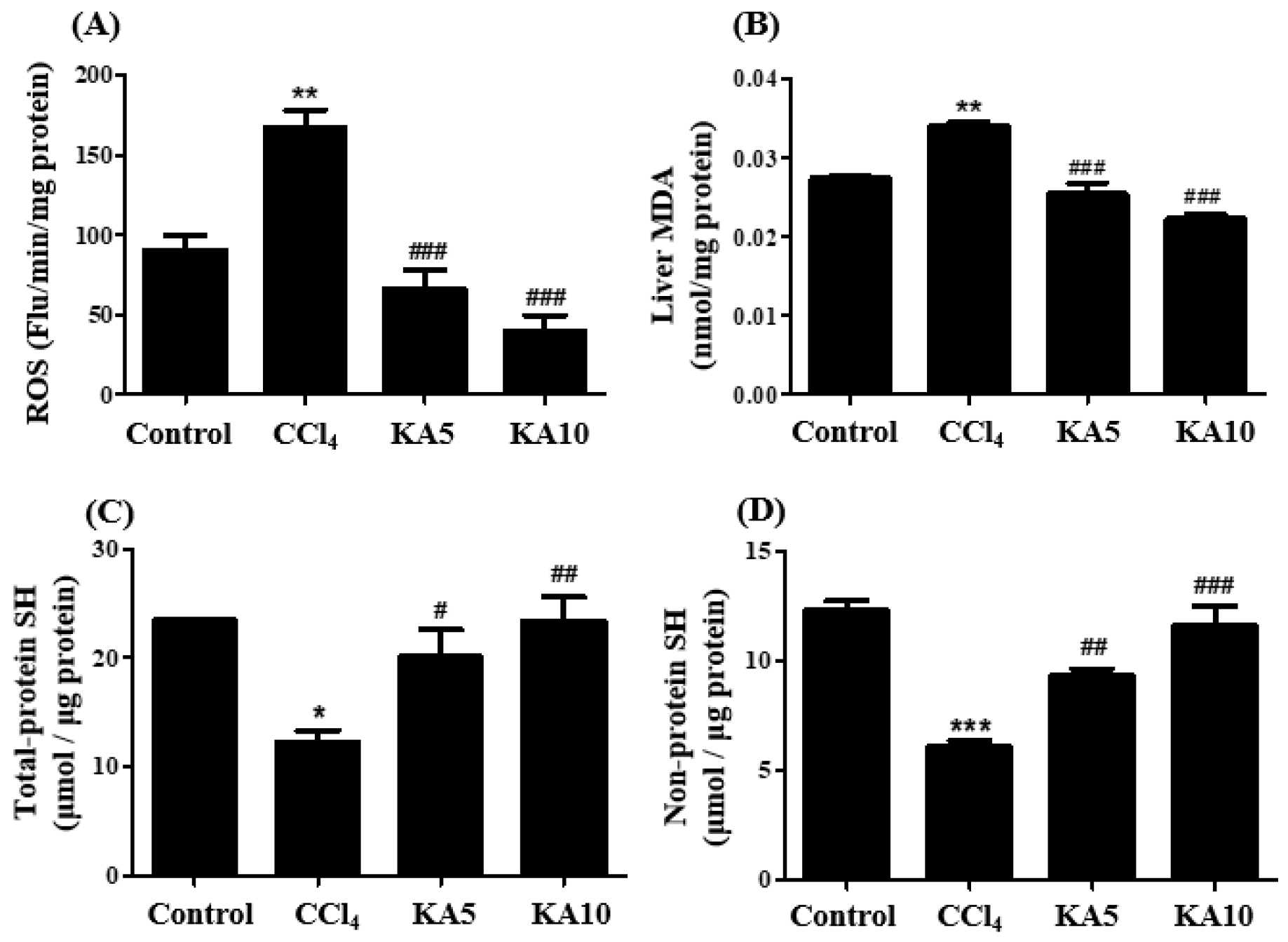
2.7. Effect of Kaempferol on Oxidative Status in CCl4-Intoxicated Rat’s Liver
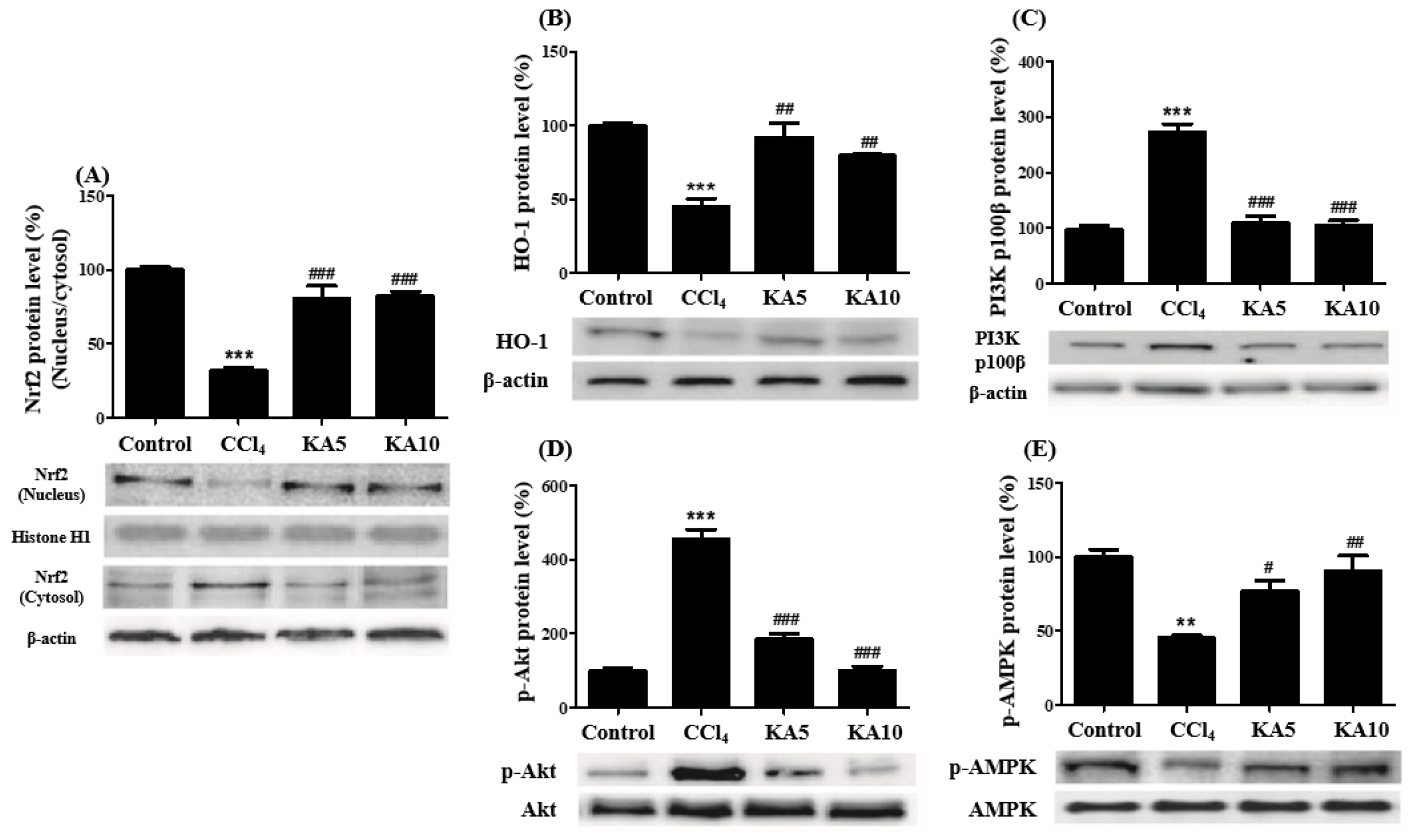
2.8. Effects of Kaempferol on Nrf2 Activation and the PI3K/Akt and AMPK Signaling Pathways
3. Discussion
4. Materials and Methods
4.1. DPPH Assay and Antioxidative Activity against FeSO4/H2O2-Induced Lipid Peroxidation
4.2. HepG2 Cell Damage Induced by t-BHP
4.3. Animals and Induction of Acute Liver Injury with CCl4
4.4. Liver Histology and Biochemical Analysis of Serum Parameters
4.5. MDA and ROS Level in Liver Tissues
4.6. Total SH and Non-Protein SH Contents in Liver Tissues
4.7. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
4.8. Western Blot Analysis
4.9. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vona, R.; Pallotta, L.; Cappelletti, M.; Severi, C.; Matarrese, P. The impact of oxidative stress in human pathology: Focus on gastrointestinal disorders. Antioxidants 2021, 10, 201. [Google Scholar] [CrossRef] [PubMed]
- Arias, I.M.; Alter, H.J.; Boyer, J.L.; Cohen, D.E.; Shafritz, D.A.; Thorgeirsson, S.S.; Wolkoff, A.W. The Liver: Biology and Pathobiology; John Wiley & Sons: Hoboken, NJ, USA, 2020; pp. 714–727. [Google Scholar]
- Ramos-Tovar, E.; Muriel, P. Molecular mechanisms that link oxidative stress, inflammation, and fibrosis in the liver. Antioxidants 2020, 9, 1279. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Tan, H.Y.; Wang, N.; Zhang, Z.J.; Lao, L.; Wong, C.W.; Feng, Y. The role of oxidative stress and antioxidants in liver diseases. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, J.; Casado-Andres, M.; Goikoetxea-Usandizaga, N.; Serrano-Macia, M.; Martinez-Chantar, M.L. Nutraceutical properties of polyphenols against liver diseases. Nutrients 2020, 12, 3517. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.H. Health-promoting components of fruits and vegetables in the diet. Adv. Nutr. 2013, 4, 384S–392S. [Google Scholar] [CrossRef] [Green Version]
- Dwivedi, S.; Kushalan, S.; Paithankar, J.G.; D’Souza, L.C.; Hegde, S.; Sharma, A. Environmental toxicants, oxidative stress and health adversities: Interventions of phytochemicals. J. Pharm. Pharm. 2022, 74, 516–536. [Google Scholar] [CrossRef]
- Alam, W.; Khan, H.; Shah, M.A.; Cauli, O.; Saso, L. Kaempferol as a dietary anti-inflammatory agent: Current therapeutic standing. Molecules 2020, 25, 4073. [Google Scholar] [CrossRef]
- Calderon-Montano, J.M.; Burgos-Moron, E.; Perez-Guerrero, C.; Lopez-Lazaro, M. A review on the dietary flavonoid kaempferol. Mini. Rev. Med. Chem. 2011, 11, 298–344. [Google Scholar] [CrossRef]
- Devi, K.P.; Malar, D.S.; Nabavi, S.F.; Sureda, A.; Xiao, J.; Nabavi, S.M.; Daglia, M. Kaempferol and inflammation: From chemistry to medicine. Pharm. Res. 2015, 99, 1–10. [Google Scholar] [CrossRef]
- Rajendran, P.; Rengarajan, T.; Nandakumar, N.; Palaniswami, R.; Nishigaki, Y.; Nishigaki, I. Kaempferol, a potential cytostatic and cure for inflammatory disorders. Eur. J. Med. Chem. 2014, 86, 103–112. [Google Scholar] [CrossRef]
- Silva Dos Santos, J.; Goncalves Cirino, J.P.; de Oliveira Carvalho, P.; Ortega, M.M. The pharmacological action of kaempferol in central nervous system diseases: A review. Front. Pharm. 2020, 11, 565700. [Google Scholar] [CrossRef]
- Correa, J.G.S.; Bianchin, M.; Lopes, A.P.; Silva, E.; Ames, F.Q.; Pomini, A.M.; Carpes, S.T.; de Carvalho Rinaldi, J.; Cabral Melo, R.; Kioshima, E.S. Chemical profile, antioxidant and anti-inflammatory properties of Miconia albicans (Sw.) Triana (Melastomataceae) fruits extract. J. Ethnopharmacol. 2021, 273, 113979. [Google Scholar] [CrossRef]
- Kumar, A.; Kaur, V.; Pandit, K.; Tuli, H.S.; Sak, K.; Jain, S.K.; Kaur, S. Antioxidant phytoconstituents from Onosma bracteata Wall. (Boraginaceae) ameliorate the CCl4 induced hepatic damage: In vivo study in male wistar rats. Front. Pharm. 2020, 11, 1301. [Google Scholar] [CrossRef]
- Al-Olayan, E.M.; El-Khadragy, M.F.; Aref, A.M.; Othman, M.S.; Kassab, R.B.; Abdel Moneim, A.E. The potential protective effect of Physalis peruviana L. against carbon tetrachloride-induced hepatotoxicity in rats is mediated by suppression of oxidative stress and downregulation of MMP-9 expression. Oxid. Med. Cell. Longev. 2014, 2014, 381413. [Google Scholar] [CrossRef] [Green Version]
- Hawas, U.W.; Soliman, G.M.; Abou El-Kassem, L.T.; Farrag, A.R.; Mahmoud, K.; Leon, F. A new flavonoid C-glycoside from Solanum elaeagnifolium with hepatoprotective and curative activities against paracetamol-induced liver injury in mice. Z. Nat. C. J. Biosci. 2013, 68, 19–28. [Google Scholar] [CrossRef]
- BinMowyna, M.N.; AlFaris, N.A. Kaempferol suppresses acetaminophen-induced liver damage by upregulation/activation of SIRT1. Pharm. Biol. 2021, 59, 146–156. [Google Scholar] [CrossRef]
- Wang, M.; Sun, J.; Jiang, Z.; Xie, W.; Zhang, X. Hepatoprotective effect of kaempferol against alcoholic liver injury in mice. Am. J. Chin. Med. 2015, 43, 241–254. [Google Scholar] [CrossRef] [Green Version]
- Shih, T.Y.; Young, T.H.; Lee, H.S.; Hsieh, C.B.; Hu, O.Y. Protective effects of kaempferol on isoniazid- and rifampicin-induced hepatotoxicity. AAPS J. 2013, 15, 753–762. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.S.; Kim, K.S.; Ko, W.; Li, B.; Jeong, G.S.; Jang, J.H.; Oh, H.; Kim, Y.C. The cytoprotective effect of sulfuretin against tert-butyl hydroperoxide-induced hepatotoxicity through Nrf2/ARE and JNK/ERK MAPK-mediated heme oxygenase-1 expression. Int. J. Mol. Sci. 2014, 15, 8863–8877. [Google Scholar] [CrossRef] [Green Version]
- Krithika, R.; Mohankumar, R.; Verma, R.J.; Shrivastav, P.S.; Mohamad, I.L.; Gunasekaran, P.; Narasimhan, S. Isolation, characterization and antioxidative effect of phyllanthin against CCl4-induced toxicity in HepG2 cell line. Chem. Biol. Interact. 2009, 181, 351–358. [Google Scholar] [CrossRef]
- McCay, P.B.; Lai, E.K.; Poyer, J.L.; DuBose, C.M.; Janzen, E.G. Oxygen- and carbon-centered free radical formation during carbon tetrachloride metabolism. Observation of lipid radicals In vivo and In vitro. J. Biol. Chem. 1984, 259, 2135–2143. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Hwang, S.; Ahmed, Y.A.; Feng, D.; Li, N.; Ribeiro, M.; Lafdil, F.; Kisseleva, T.; Szabo, G.; Gao, B. Immunopathobiology and therapeutic targets related to cytokines in liver diseases. Cell Mol. Immunol. 2021, 18, 18–37. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; Choi, E.J. Compromised MAPK signaling in human diseases: An update. Arch. Toxicol. 2015, 89, 867–882. [Google Scholar] [CrossRef] [PubMed]
- Sova, M.; Saso, L. Design and development of Nrf2 modulators for cancer chemoprevention and therapy: A review. Drug Des. Devel. 2018, 12, 3181–3197. [Google Scholar] [CrossRef] [Green Version]
- Iranshahy, M.; Iranshahi, M.; Abtahi, S.R.; Karimi, G. The role of nuclear factor erythroid 2-related factor 2 in hepatoprotective activity of natural products: A review. Food Chem. Toxicol. 2018, 120, 261–276. [Google Scholar] [CrossRef]
- Rajendran, P.; Ammar, R.B.; Al-Saeedi, F.J.; Mohamed, M.E.; ElNaggar, M.A.; Al-Ramadan, S.Y.; Bekhet, G.M.; Soliman, A.M. Kaempferol inhibits zearalenone-induced oxidative stress and apoptosis via the PI3K/Akt-mediated Nrf2 signaling pathway: In vitro and in vivo studies. Int. J. Mol. Sci. 2020, 22, 217. [Google Scholar] [CrossRef]
- Zhang, L.; Guo, Z.; Wang, Y.; Geng, J.; Han, S. The protective effect of kaempferol on heart via the regulation of Nrf2, NF-kappabeta, and PI3K/Akt/GSK-3beta signaling pathways in isoproterenol-induced heart failure in diabetic rats. Drug Dev. Res. 2019, 80, 294–309. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, X.; Li, D.; Hao, W.; Meng, F.; Wang, B.; Han, J.; Zheng, Q. Kaempferide protects against myocardial ischemia/reperfusion injury through activation of the PI3K/Akt/GSK-3β pathway. Mediat. Inflamm. 2017, 2017, 5278218. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.L.; Liu, J.X.; Dong, W.; Li, P.; Li, L.; Hou, J.C.; Zheng, Y.Q.; Lin, C.R.; Ren, J.G. Protective effect of kaempferol on LPS plus ATP-induced inflammatory response in cardiac fibroblasts. Inflammation 2015, 38, 94–101. [Google Scholar] [CrossRef]
- Jiang, K.; Guo, S.; Yang, C.; Yang, J.; Chen, Y.; Shaukat, A.; Zhao, G.; Wu, H.; Deng, G. Barbaloin protects against lipopolysaccharide (LPS)-induced acute lung injury by inhibiting the ROS-mediated PI3K/AKT/NF-kappaB pathway. Int. Immunopharmacol. 2018, 64, 140–150. [Google Scholar] [CrossRef]
- Zhu, M.; Zhou, X.; Zhao, J. Quercetin prevents alcohol-induced liver injury through targeting of PI3K/Akt/nuclear factor-κB and STAT3 signaling pathway. Exp. Ther. Med. 2017, 14, 6169–6175. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Wang, Q.; Wang, W.; Wei, S.; Zeng, Q.; Zhang, A. Semaphorin 4A antibody alleviates arsenic-induced hepatotoxicity in mice via inhibition of AKT2/NF-kappaB inflammatory signaling. Toxicol. Appl. Pharm. 2021, 410, 115364. [Google Scholar] [CrossRef]
- Steinberg, G.R.; Kemp, B.E. AMPK in Health and Disease. Physiol. Rev. 2009, 89, 1025–1078. [Google Scholar] [CrossRef]
- Ding, X.; Jian, T.; Li, J.; Lv, H.; Tong, B.; Li, J.; Meng, X.; Ren, B.; Chen, J. Chicoric acid ameliorates nonalcoholic fatty liver disease via the AMPK/Nrf2/NFkappaB signaling pathway and restores gut microbiota in high-fat-diet-fed mice. Oxid. Med. Cell Longev. 2020, 2020, 9734560. [Google Scholar] [CrossRef]
- Xu, W.; Zhao, T.; Xiao, H. The implication of oxidative stress and AMPK-Nrf2 antioxidative signaling in pneumonia pathogenesis. Front. Endocrinol. Lausanne 2020, 11, 400. [Google Scholar] [CrossRef]
- Velagapudi, R.; Jamshaid, F.; Lepiarz, I.; Katola, F.O.; Hemming, K.; Olajide, O.A. The tiliroside derivative, 3-O-[(E)-(2-oxo-4-(p-tolyl) but-3-en-1-yl] kaempferol produced inhibition of neuroinflammation and activation of AMPK and Nrf2/HO-1 pathways in BV-2 microglia. Int. Immunopharmacol. 2019, 77, 105951. [Google Scholar] [CrossRef]
- Du, Y.; Han, J.; Zhang, H.; Xu, J.; Jiang, L.; Ge, W. Kaempferol prevents against Ang II-induced cardiac remodeling through attenuating Ang II-induced inflammation and oxidative stress. J. Cardiovasc. Pharm. 2019, 74, 326–335. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.-E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Bak, J.; Je, N.K.; Chung, H.Y.; Yokozawa, T.; Yoon, S.; Moon, J.O. Oligonol ameliorates CCl4-induced liver injury in rats via the NF-Kappa B and MAPK signaling pathways. Oxid. Med. Cell. Longev. 2016, 2016, 3935841. [Google Scholar] [CrossRef] [Green Version]
- Cheong, K.O.; Shin, D.S.; Bak, J.; Lee, C.; Kim, K.W.; Je, N.K.; Chung, H.Y.; Yoon, S.; Moon, J.O. Hepatoprotective effects of zingerone on carbon tetrachloride- and dimethylnitrosamine-induced liver injuries in rats. Arch. Pharm. Res. 2016, 39, 279–291. [Google Scholar] [CrossRef]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Concentration (μM) | Inhibition (%) | IC50 (μM) |
|---|---|---|---|
| Kaempferol | 2 | 0.34 ± 0.01 | 9.69 |
| 5 | 17.94 ± 0.01 | ||
| 10 | 54.99 ± 0.00 | ||
| 20 | 75.80 ± 0.00 | ||
| BHT | 2 | 15.39 ± 0.00 | 8.66 |
| 5 | 27.33 ± 0.00 | ||
| 10 | 60.57 ± 0.02 | ||
| 20 | 70.05 ± 0.00 |
| Compound | Concentration (μM) | Inhibition (%) | IC50 (μM) |
|---|---|---|---|
| Kaempferol | 2 | 8.74 ± 0.01 | 21.87 |
| 5 | 16.02 ± 0.39 | ||
| 10 | 23.29 ± 0.01 | ||
| 25 | 53.61 ± 0.00 | ||
| 50 | 73.31 ± 0.00 | ||
| BHT | 5 | 15.00 ± 0.00 | 23.71 |
| 10 | 20.56 ± 0.00 | ||
| 20 | 44.51 ± 0.00 | ||
| 50 | 83.36 ± 0.10 | ||
| 100 | 92.49 ± 0.07 |
| Compound | Bodyweight (g) | Liver Weight (g) | Ratio (%) a |
|---|---|---|---|
| Control | 154.02 ± 7.40 | 5.96 ± 1.25 | 3.85 |
| CCl4 | 158.01 ± 7.29 | 7.69 ± 0.66 ** | 4.87 |
| KA5 | 158.16 ± 4.62 | 6.98 ± 0.34 # | 4.42 |
| KA10 | 156.75 ± 10.05 | 6.71 ± 0.29 # | 4.29 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, C.; Yoon, S.; Moon, J.-O. Kaempferol Suppresses Carbon Tetrachloride-Induced Liver Damage in Rats via the MAPKs/NF-κB and AMPK/Nrf2 Signaling Pathways. Int. J. Mol. Sci. 2023, 24, 6900. https://doi.org/10.3390/ijms24086900
Lee C, Yoon S, Moon J-O. Kaempferol Suppresses Carbon Tetrachloride-Induced Liver Damage in Rats via the MAPKs/NF-κB and AMPK/Nrf2 Signaling Pathways. International Journal of Molecular Sciences. 2023; 24(8):6900. https://doi.org/10.3390/ijms24086900
Chicago/Turabian StyleLee, Changyong, Sik Yoon, and Jeon-Ok Moon. 2023. "Kaempferol Suppresses Carbon Tetrachloride-Induced Liver Damage in Rats via the MAPKs/NF-κB and AMPK/Nrf2 Signaling Pathways" International Journal of Molecular Sciences 24, no. 8: 6900. https://doi.org/10.3390/ijms24086900