
Nisin S, a Novel Nisin Variant Produced by Ligilactobacillus salivarius P1CEA3
 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Direct and Extracellular Antimicrobial Activity of L. salivarius P1CEA3 and Stability to pH, Heat, and Proteases
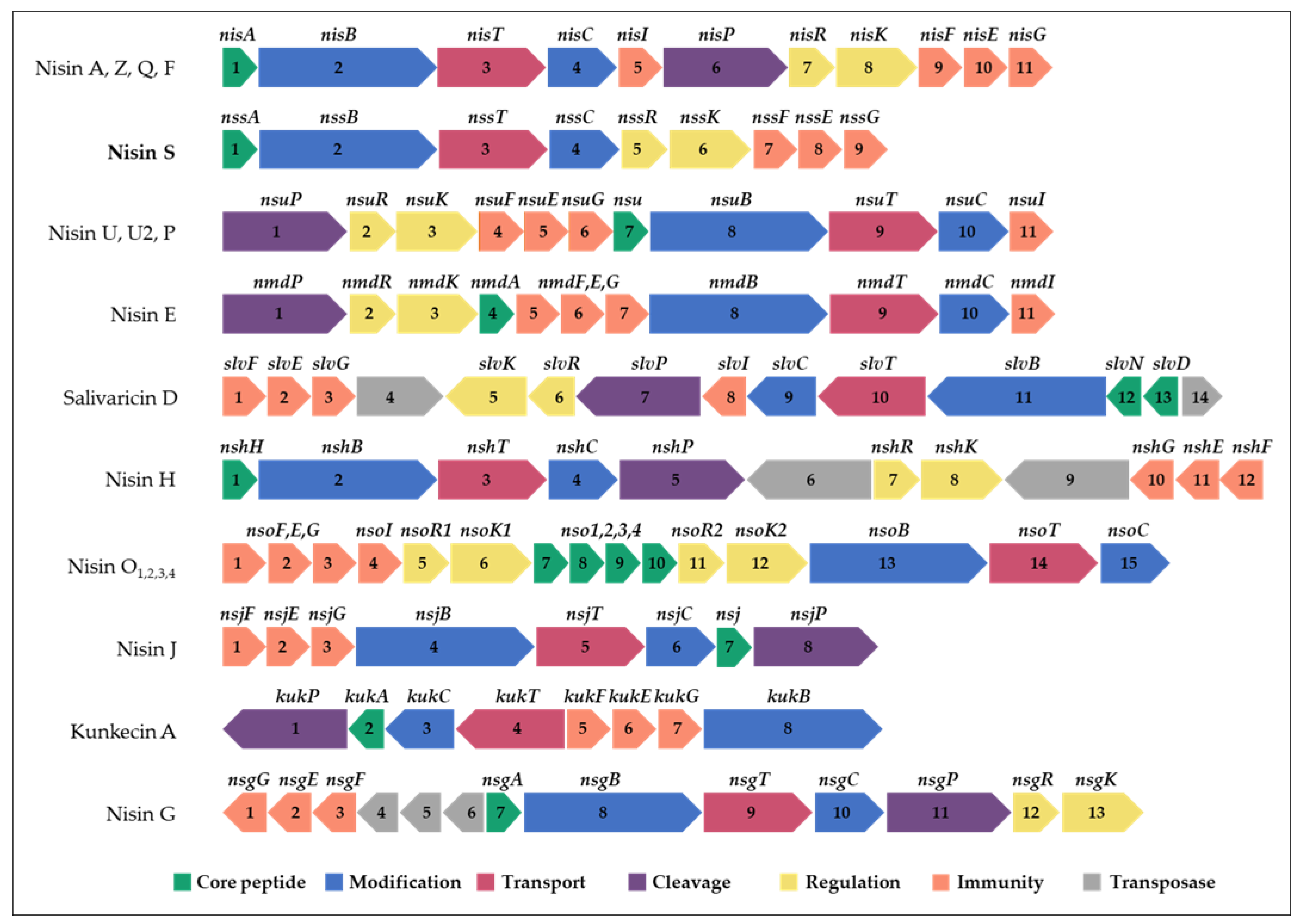
2.2. Identification of Two Bacteriocin Gene Clusters in the L. salivarius P1CEA3 Genome
2.3. In Vitro Cell-Free Protein Synthesis (IV-CFPS) and Evaluation of the Antimicrobial Activity of Abp118 (Abp118α + Abp118β), Salivaricin B (SalB), and Nisin S
2.4. Colony MALDI-TOF MS for Identification of the Nisin S Produced by L. salivarius P1CEA3
2.5. Purification of Bacteriocins from Supernatants (CFS) of L. salivarius P1ACE3, and MALDI-TOF MS Analysis of the Purified Fractions

2.6. Massive Peptide Analysis of Trypsinized RP-FPLC-Purified Fractions from Supernatants of L. salivarius P1ACE3, and Predicted Structure of the Nisin S
2.7. Antimicrobial Activity of Nisin S
3. Discussion
4. Materials and Methods
4.1. Isolation and Identification of Ligilactobacillus salivarius P1CEA3
4.2. Bacterial Strains, Media and Culture Conditions
4.3. Direct Antimicrobial Activity of L. salivarius P1CEA3
4.4. Isolation of DNA, Whole Genome Sequencing (WGS), and Bioinformatic Analysis
4.5. In Vitro Cell-Free Protein Synthesis (IV-CFPS) of Bacteriocins and Peptides of Interest, and Evaluation of Their Antimicrobial Activity
4.6. Culture Media, Growth Conditions, and Antimicrobial Activity in Supernatants of L. salivarius P1CEA3
4.7. Colony Matrix-Assisted Laser Desorption/Ionization Time of Flight Mass Spectrometry (MALDI-TOF MS) of L. salivarius P1CEA3
4.8. Effect of Heat, pH, and Proteases on Cell-Free Supernatants (CFS) of L. salivarius P1CEA3
4.9. Purification of the Bacteriocins Produced by L. salivarius P1CEA3
4.10. Targeted Proteomics Identification Combined with Massive Peptide Analysis of the Eluted Purified Fractions with Higher Antimicrobial Activity, Derived from L. salivarius P1ACE3
4.11. Antimicrobial Activity of the Purified Nisin S against Different Indicator Strains
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ben Lagha, A.; Haas, B.; Gottschalk, M.; Grenier, D. Antimicrobial potential of bacteriocins in poultry and swine production. Vet. Res. 2017, 48, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, B.M.; Levy, S.B. Food animals and antimicrobials: Impacts on human health. Clin. Microbiol. Rev. 2011, 24, 718–733. [Google Scholar] [CrossRef] [Green Version]
- EUR-Lex—Regulation (EU) 2019/6 of the European Parliament and of the Council of 11 December 2018 on Veterinary Medicinal Products and Repealing Directive 2001/82/EC. PE/45/2018/REV/1. Available online: https://eur-lex.europa.eu/eli/reg/2019/6/oj (accessed on 26 October 2022).
- Hibbing, M.E.; Fuqua, C.; Parsek, M.R.; Peterson, S.B. Bacterial competition: Surviving and thriving in the microbial jungle. Nat. Rev. Microbiol. 2010, 8, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A viable alternative to antibiotics? Nat. Rev. Microbiol. 2012, 11, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Ma, L.; Nie, Y.; Chen, J.; Zheng, W.; Wang, X.; Xie, C.; Zheng, Z.; Wang, Z.; Yang, T.; et al. A microbiota-derived bacteriocin targets the host to confer diarrhea resistance in early-weaned piglets. Cell. Host. Microbe 2018, 24, 817–832.e8. [Google Scholar] [CrossRef] [Green Version]
- Gharsallaoui, A.; Oulahal, N.; Joly, C.; Degraeve, P. Nisin as a food preservative: Part 1: Physicochemical properties, antimicrobial activity, and main uses. Crit. Rev. Food. Sci. Nutr. 2016, 56, 1262–1274. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, J.N.; O’Connor, P.M.; Rea, M.C.; O’Sullivan, O.; Walsh, C.J.; Healy, B.; Mathur, H.; Field, D.; Hill, C.; Paul Ross, R. Nisin J, a novel natural nisin variant, is produced by Staphylococcus capitis sourced from the human skin microbiota. J. Bacteriol. Res. 2020, 202, e00639-19. [Google Scholar] [CrossRef] [Green Version]
- Draper, L.A.; Cotter, P.D.; Hill, C.; Ross, R.P. Lantibiotic resistance. Microbiol. Mol. Biol. Rev. 2015, 79, 171–191. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Fang, J.; Tian, Y.; Lu, X.Y. Mechanisms of nisin resistance in Gram-positive bacteria. Ann. Microbiol. 2014, 64, 413–420. [Google Scholar] [CrossRef]
- Umu, Ö.C.O.; Rudi, K.; Diep, D.B. Modulation of the gut microbiota by prebiotic fibres and bacteriocins. Microb. Ecol. Health Dis. 2017, 28, 1348886. [Google Scholar] [CrossRef] [Green Version]
- de Kwaadsteniet, M.; Ten Doeschate, K.; Dicks, L.M. T Characterization of the structural gene encoding nisin F, a new lantibiotic produced by a Lactococcus lactis subsp. lactis isolate from freshwater catfish (Clarias gariepinus). Appl. Environ. Microbiol. 2008, 74, 547–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soltani, S.; Hammami, R.; Cotter, P.D.; Rebuffat, S.; ben Said, L.; Gaudreau, H.; Bédard, F.; Biron, E.; Drider, D.; Fliss, I. Bacteriocins as a new generation of antimicrobials: Toxicity aspects and regulations. FEMS Microbiol. Rev. 2021, 45, 1–24. [Google Scholar] [CrossRef]
- Hansen, J.N. Nisin as a model food preservative. Crit. Rev. Food. Sci. Nutr. 1994, 34, 69–93. [Google Scholar] [CrossRef]
- Rogers, L.A. The inhibiting effect of Streptococcus. lactis on Lactobacillus bulgaricus. J. Bacteriol. 1928, 16, 321–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulders, J.W.; Boerrigter, I.J.; Rollema, H.S.; Siezen, R.J.; de Vos, W.M. Identification and characterization of the lantibiotic nisin Z, a natural nisin variant. Eur. J. Biochem. 1991, 201, 581–584. [Google Scholar] [CrossRef]
- Zendo, T.; Fukao, M.; Ueda, K.; Higuchi, T.; Nakayama, J.; Sonomoto, K. Identification of the lantibiotic nisin Q, a new natural nisin variant produced by Lactococcus lactis 61-14 isolated from a river in Japan. Biosci. Biotechnol. Biochem. 2003, 67, 1616–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirawan, R.E.; Klesse, N.A.; Jack, R.W.; Tagg, J.R. Molecular and genetic characterization of a novel nisin variant produced by Streptococcus uberis. Appl. Environ. Microbiol. 2006, 72, 1148–1156. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, P.M.; O’Shea, E.F.; Guinane, C.M.; O’Sullivan, O.; Cotter, P.D.; Ross, R.P.; Hill, C. Nisin H is a new nisin variant produced by the gut-derived strain Streptococcus hyointestinalis DPC6484. Appl. Environ. Microbiol. 2015, 81, 3953–3960. [Google Scholar] [CrossRef] [Green Version]
- Hatziioanou, D.; Gherghisan-Filip, C.; Saalbach, G.; Horn, N.; Wegmann, U.; Duncan, S.H.; Flint, H.J.; Mayer, M.J.; Narbad, A. Discovery of a novel lantibiotic nisin O from Blautia. obeum A2-162, isolated from the human gastrointestinal tract. Microbiology 2017, 163, 1292–1305. [Google Scholar] [CrossRef]
- Garcia-Gutierrez, E.; O’Connor, P.M.; Saalbach, G.; Walsh, C.J.; Hegarty, J.W.; Guinane, C.M.; Mayer, M.J.; Narbad, A.; Cotter, P.D. First evidence of production of the lantibiotic nisin P. Sci. Rep. 2020, 10, 3738. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, G.W.; Garcia-Gutierrez, E.; Walsh, C.J.; O’connor, P.M.; Begley, M.; Cotter, P.D.; Guinane, C.M. Nisin G is a novel nisin variant produced by a gut-derived Streptococcus salivarius. bioRxiv 2022. [Google Scholar] [CrossRef]
- Christophers, M.; Heng, L.; Heng, N. Nisin E, a new nisin variant produced by Streptococcus equinus MDC1. Appl. Sci. 2023, 13, 1186. [Google Scholar] [CrossRef]
- Birri, D.J.; Brede, D.A.; Nes, I.F. Salivaricin D, a novel intrinsically trypsin-resistant lantibiotic from Streptococcus. salivarius 5M6c isolated from a healthy infant. Appl. Environ. Microbiol. 2012, 78, 402–410. [Google Scholar] [CrossRef] [Green Version]
- Zendo, T.; Ohashi, C.; Maeno, S.; Piao, X.; Salminen, S.; Sonomoto, K.; Endo, A. Kunkecin A, a new nisin variant bacteriocin produced by the fructophilic lactic acid bacterium, Apilactobacillus kunkeei FF30-6 isolated from honey bees. Front. Microbiol. 2020, 11, 571903. [Google Scholar] [CrossRef]
- Panina, I.; Krylov, N.; Nolde, D.; Efremov, R.; Chugunov, A. Environmental and dynamic effects explain how nisin captures membrane-bound lipid II. Sci. Rep. 2020, 10, 8821. [Google Scholar] [CrossRef]
- Zheng, Y.; Du, Y.; Qiu, Z.; Liu, Z.; Qiao, J.; Li, Y.; Caiyin, Q. Nisin variants generated by protein engineering and their properties. Bioengineering 2022, 9, 251. [Google Scholar] [CrossRef]
- Yeluri Jonnala, B.R.; Feehily, C.; O’Connor, P.M.; Field, D.; Hill, C.; Ross, R.P.; McSweeney, P.L.H.; Sheehan, J.J.; Cotter, P.D. Assessing the ability of nisin A and derivatives thereof to inhibit gram-negative bacteria from the genus Thermus. J. Dairy Sci. 2021, 104, 2632–2640. [Google Scholar] [CrossRef]
- Field, D.; Considine, K.; O’connor, P.M.; Ross, R.P.; Hill, C.; Cotter, P.D. Bio-engineered nisin with increased anti-Staphylococcus and selectively reduced anti-Lactococcus activity for treatment of bovine mastitis. Int. J. Mol. Sci. 2021, 22, 3480. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, M.; Field, D.; Grainger, A.; O’Connor, P.M.; Draper, L.; Ross, R.P.; Hill, C. Nisin M: A bioengineered nisin A variant that retains full induction capacity but has significantly reduced antimicrobial activity. Appl. Environ. Microbiol. 2020, 86, e00984-20. [Google Scholar] [CrossRef] [PubMed]
- EFSA BIOHAZ Panel (EFSA Panel on Biological Hazards); Koutsoumanis, K.; Allende, A.; Álvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; De Cesare, A.; Hilbert, F.; Lindqvist, R.; et al. Scientific Opinion on the update of the list of qualified presumption of safety (QPS) recommended microorganisms intentionally added to food or feed as notified to EFSA. EFSA J. 2023, 21, e07747-23. [Google Scholar] [CrossRef]
- Guerrero Sanchez, M.; Passot, S.; Campoy, S.; Olivares, M.; Fonseca, F. Ligilactobacillus salivarius functionalities, applications, and manufacturing challenges. Appl. Microbiol. Biotechnol. 2021, 106, 57–80. [Google Scholar] [CrossRef]
- Flynn, S.; van Sinderen, D.; Thornton, G.M.; Holo, H.; Nes, I.F.; Collins, J.K. Characterization of the genetic locus responsible for the production of ABP-118, a novel bacteriocin produced by the probiotic bacterium Lactobacillus salivarius subsp. salivarius UCC118. Microbiology 2002, 148 Pt 4, 973–984. [Google Scholar] [CrossRef] [Green Version]
- Çataloluk, O. Molecular characterization of the gene encoding for the Salivaricin B activity and its flanking sequences. Turk. J. Biol. 2001, 25, 379–386. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Talebi Bezmin Abadi, A.; Rizvanov, A.A.; Haertlé, T.; Blatt, N.L. World Health Organization Report: Current crisis of antibiotic resistance. BioNanoSci 2019, 9, 778–788. [Google Scholar] [CrossRef]
- Su, W.; Gong, T.; Jiang, Z.; Lu, Z.; Wang, Y. The role of probiotics in alleviating postweaning diarrhea in piglets from the perspective of intestinal barriers. Front. Cell. Infect. Microbiol. 2022, 12, 883107. [Google Scholar] [CrossRef]
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. In Review on Antimicrobial Resistance; 2016; Available online: https://amr-review.org/sites/default/files/160525_Final%20paper_with%20cover.pdf (accessed on 26 October 2022).
- Jiménez, E.; Martín, R.; Maldonado, A.; Martín, V.; Gómez De Segura, A.; Fernández, L.; Rodríguez, J.M. Complete genome sequence of Lactobacillus salivarius CECT 5713, a probiotic strain isolated from human milk and infant feces. J. Bacteriol. 2010, 192, 5266–5267. [Google Scholar] [CrossRef] [Green Version]
- Gabant, P.; Borrero, J. PARAGEN 1.0: A standardized synthetic gene library for fast cell-free bacteriocin synthesis. Front. Bioeng. Biotechnol. 2019, 7, 213. [Google Scholar] [CrossRef] [PubMed]
- Montalbán-López, M.; Scott, T.A.; Ramesh, S.; Rahman, I.R.; van Heel, A.J.; Viel, J.H.; Bandarian, V.; Dittmann, E.; Genilloud, O.; Goto, Y.; et al. New developments in RiPP discovery, enzymology and engineering. Nat. Prod. Rep. 2021, 38, 130–239. [Google Scholar] [CrossRef]
- Ongpipattanakul, C.; Desormeaux, E.K.; Dicaprio, A.; van der Donk, W.A.; Mitchell, D.A.; Nair, S.K. Mechanism of action of ribosomally synthesized and post-translationally modified peptides. Chem. Rev. 2022, 122, 14722–14814. [Google Scholar] [CrossRef]
- Claesson, M.J.; Li, Y.; Leahy, S.; Canchaya, C.; van Pijkeren, J.P.; Cerdeño-Tárraga, A.M.; Parkhill, J.; Flynn, S.; O’Sullivan, G.C.; Collins, J.K.; et al. Multireplicon genome architecture of Lactobacillus salivarius. Proc. Natl. Acad. Sci. USA 2006, 103, 6718–6723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, E.; Hayes, M.; O’Connor, P.; Gardiner, G.; Fitzgerald, G.F.; Stanton, C.; Ross, R.P.; Hill, C. Salivaricin P, one of a family of two-component antilisterial bacteriocins produced by intestinal isolates of Lactobacillus salivarius. Appl. Environ. Microbiol. 2007, 73, 3719–3723. [Google Scholar] [CrossRef] [Green Version]
- O’Shea, E.F.; O’Connor, P.M.; Raftis, E.J.; O’Toole, P.W.; Stanton, C.; Cotter, P.D.; Ross, R.P.; Hill, C. Production of multiple bacteriocins from a single locus by gastrointestinal strains of Lactobacillus salivarius. J. Bacteriol 2011, 193, 6973–6982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havarstein, L.S.; Diep, D.B.; Nes, I.F. A family of bacteriocin ABC transporters carry out proteolytic processing of their substrates concomitant with export. Mol. Microbiol. 1995, 16, 229–240. [Google Scholar] [CrossRef]
- Kuipers, O.P.; Beerthuyzen, M.M.; Siezen, R.J.; De Vos, W.M. Characterization of the nisin gene cluster nisABTCIPR of Lactococcus. lactis. Requirement of expression of the nisA and nisI genes for development of immunity. Eur. J. Biochem. 1993, 216, 281–291. [Google Scholar] [CrossRef]
- Montalbán-López, M.; Deng, J.; van Heel, A.J.; Kuipers, O.P. Specificity and application of the lantibiotic protease NisP. Front. Microbiol. 2018, 9, 160. [Google Scholar] [CrossRef] [Green Version]
- Corvey, C.; Stein, T.; Düsterhus, S.; Karas, M.; Entian, K.D. Activation of subtilin precursors by Bacillus subtilis extracellular serine proteases subtilisin (AprE), WprA, and Vpr. Biochem. Biophys. Res. Commun. 2003, 304, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Qi, F.; Chen, P.; Caufield, P.W. The group I strain of Streptococcus mutans, UA140, produces both the lantibiotic mutacin I and a nonlantibiotic bacteriocin, mutacin IV. Appl. Environ. Microbiol. 2001, 67, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Stein, T.; Heinzmann, S.; Solovieva, I.; Entian, K.D. Function of Lactococcus. lactis nisin immunity genes nisI and nisFEG after coordinated expression in the surrogate host Bacillus subtilis. J. Biol. Chem. 2003, 278, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Lohans, C.T.; Vederas, J.C. Structural characterization of thioether-bridged bacteriocins. J. Antibiot. 2014, 67, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Field, D.; Begley, M.; O’Connor, P.M.; Daly, K.M.; Hugenholtz, F.; Cotter, P.D.; Hill, C.; Ross, R.P. Bioengineered nisin A derivatives with enhanced activity against both Gram positive and Gram negative pathogens. PLoS ONE 2012, 7, e46884. [Google Scholar] [CrossRef] [Green Version]
- Healy, B.; Field, D.; O’Connor, P.M.; Hill, C.; Cotter, P.D.; Ross, R.P. Intensive mutagenesis of the nisin hinge leads to the rational design of enhanced derivatives. PLoS ONE 2013, 8, e79563. [Google Scholar] [CrossRef] [Green Version]
- Field, D.; Connor, P.M.O.; Cotter, P.D.; Hill, C.; Ross, R.P. The generation of nisin variants with enhanced activity against specific gram-positive pathogens. Mol. Microbiol. 2008, 69, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Sugrue, I.; Hill, D.; O’Connor, P.M.; Day, L.; Stanton, C.; Hill, C.; Ross, R.P. Nisin E is a novel nisin variant produced by multiple Streptococcus equinus strains. Microorganisms 2023, 11, 427. [Google Scholar] [CrossRef]
- Sun, Z.; Zhong, J.; Liang, X.; Liu, J.; Chen, X.; Huan, L. Novel mechanism for nisin resistance via proteolytic degradation of nisin by the nisin resistance protein NSR. Antimicrob. Agents. Chemother. 2009, 53, 1964–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Field, D.; Blake, T.; Mathur, H.; O’ Connor, P.M.; Cotter, P.D.; Paul Ross, R.; Hill, C. Bioengineering nisin to overcome the nisin resistance protein. Mol. Microbiol. 2019, 111, 717–731. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.; Cosentino, S.; Lukjancenko, O.; Saputra, D.; Rasmussen, S.; Hasman, H.; Sicheritz-Pontén, T.; Aarestrup, F.M.; Ussery, D.W.; Lund, O. Benchmarking of methods for genomic taxonomy. J. Clin. Microbiol. 2014, 52, 1529–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joensen, K.G.; Scheutz, F.; Lund, O.; Hasman, H.; Kaas, R.S.; Nielsen, E.M.; Aarestrup, F.M. Real-time whole-genome sequencing for routine typing, surveillance, and outbreak detection of verotoxigenic Escherichia coli. J. Clin. Microbiol. 2014, 5, 1501–1510. [Google Scholar] [CrossRef] [Green Version]
- Hasman, H.; Saputra, D.; Sicheritz-Ponten, T.; Lund, O.; Svendsen, C.A.; Frimodt-Moller, N.; Aarestrup, F.M. Rapid whole-genome sequencing for detection and characterization of microorganisms directly from clinical samples. J. Clin. Microbiol. 2014, 52, 139–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, D206–D214. [Google Scholar] [CrossRef]
- van Heel, A.J.; de Jong, A.; Song, C.; Viel, J.H.; Kok, J.; Kuipers, O.P. BAGEL4: A user-friendly web server to thoroughly mine RiPPs and bacteriocins. Nucleic Acids Res. 2018, 46, W278–W281. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- UniProt Consortium. UniProt: The Universal Protein knowledgebase in 2023. Nucleic Acids Res. 2022, 51, D523–D531. [Google Scholar] [CrossRef]
- Cintas, L.M.; Rodriguez, J.M.; Fernandez, M.F.; Sletten, K.; Nes, I.F.; Hernandez, P.E.; Holo, H. Isolation and characterization of pediocin L50, a new bacteriocin from Pediococcus acidilactici with a broad inhibitory spectrum. Appl. Environ. Microbiol. 1995, 61, 2643–2648. [Google Scholar] [CrossRef] [Green Version]
- Borrero, J.; Jiménez, J.J.; Gútiez, L.; Herranz, C.; Cintas, L.M.; Hernández, P.E. Protein expression vector and secretion signal peptide optimization to drive the production, secretion, and functional expression of the bacteriocin enterocin A in lactic acid bacteria. J. Biotechnol. 2011, 156, 76–86. [Google Scholar] [CrossRef]
- Ma, B.; Zhang, K.; Hendrie, C.; Liang, C.; Li, M.; Doherty-Kirby, A.; Lajoie, G. PEAKS: Powerful software for peptide de novo sequencing by tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2003, 17, 2337–2342. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indicator Strain | Activity a |
|---|---|
| Ligilactobacillus salivarius P1CEA3 | - |
| Lactococcus lactis BB24 | - |
| Lactococcus lactis WA2–67 | - |
| Pediococcus damnosus CECT 4797 | +++ |
| Pediococcus pentosaceus FBB61 | +++ |
| Listeria seeligeri CECT 917 | ++ |
| Listeria monocytogenes CECT 4032 | + |
| Listeria innocua CECT 910 | + |
| Staphylococcus aureus ZTA11/00117ST | + |
| Staphylococcus aureus ZTA11/00310ST | + |
| Streptococcus suis CECT 958 | ++ |
| Streptococcus suis C2969/03 | ++ |
| Escherichia coli DH5α | + |
| Peptide Sequence a | Amino Acid Position | Theoretical MH+ (Da) | Found in Sample | Detected m/z |
|---|---|---|---|---|
| ITSYSLCTPGCK | [1–12] | 1385.63 | High | 693.82 |
| TGALMGCTMK | [13–22] | 1069.48 | High | 535.24 |
| TGALMGCTMK | [13–22] | 1085.47 | High | 543.24 |
| TGALMGCTMK | [13–22] | 1101.47 | High | 551.24 |
| TASCGCDVHISK | [23–34] | 1355.61 | High | 339.30 |
| Indicator Strain | Activity a | |
|---|---|---|
| Nisin S | Nisin A | |
| Ligilactobacillus salivarius P1CEA3 | ++ | + |
| Ligilactobacillus salivarius PG21 | +++ | ++ |
| Lactococcus lactis subsp. lactis BB24 | + | - |
| Pediococcus damnosus CECT 4797 | +++ | +++ |
| Pediococcus pentosaceus FBB61 | +++ | +++ |
| Enterococcus faecium ER46 | +++ | +++ |
| Enterococcus faecalis 12Ep11 | +++ | +++ |
| Listeria seeligeri CECT 917 | +++ | ++ |
| Listeria monocytogenes CECT 4032 | ++ | + |
| Listeria innocua CECT 910 | ++ | + |
| Staphylococcus aureus ZTA11/00117ST | ++ | - |
| Staphylococcus aureus DICM10/00243 | ++ | + |
| Staphylococcus epidermidis DICM15/00071 | ++ | + |
| Staphylococcus pseudintermedius ICM21/02217 | ++ | ++ |
| Streptococcus suis CECT 958 | ++ | ++ |
| Streptococcus suis serotype 1 DICM10/01182–1C | ++ | ++ |
| Streptococcus iniae ATCC 29178 | ++ | + |
| Streptococcus agalactiae DICM11/00863 | +++ | ++ |
| Streptococcus dysgalactiae VSE16/01903A | +++ | ++ |
| Streptococcus equi zooepidermicus VSE16/00697 | ++ | + |
| Corynebacterium ulcerans ICM19/00922–1B | ++ | + |
| Corynebacterium pseudotuberculosis Cam2 | ++ | - |
| Corynebacterium mastitidis INDA2 | ++ | - |
| Corynebacterium bovis AM3 | - | - |
| Bacillus cereus ICM17/00252 | - | - |
| Trueperella pyogenes ICM17/02091–1 | ++ | - |
| Erysipelothrix rhusiopathiae ICM21/01900 | +++ | + |
| Clostridium perfringens DICM15/00067–5A | +++ | ++ |
| Escherichia coli DH5α | + | - |
| Salmonella paratyphi CECT 554 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sevillano, E.; Peña, N.; Lafuente, I.; Cintas, L.M.; Muñoz-Atienza, E.; Hernández, P.E.; Borrero, J. Nisin S, a Novel Nisin Variant Produced by Ligilactobacillus salivarius P1CEA3. Int. J. Mol. Sci. 2023, 24, 6813. https://doi.org/10.3390/ijms24076813
Sevillano E, Peña N, Lafuente I, Cintas LM, Muñoz-Atienza E, Hernández PE, Borrero J. Nisin S, a Novel Nisin Variant Produced by Ligilactobacillus salivarius P1CEA3. International Journal of Molecular Sciences. 2023; 24(7):6813. https://doi.org/10.3390/ijms24076813
Chicago/Turabian StyleSevillano, Ester, Nuria Peña, Irene Lafuente, Luis M. Cintas, Estefanía Muñoz-Atienza, Pablo E. Hernández, and Juan Borrero. 2023. "Nisin S, a Novel Nisin Variant Produced by Ligilactobacillus salivarius P1CEA3" International Journal of Molecular Sciences 24, no. 7: 6813. https://doi.org/10.3390/ijms24076813