
Lung Epithelial Cells from Obese Patients Have Impaired Control of SARS-CoV-2 Infection
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
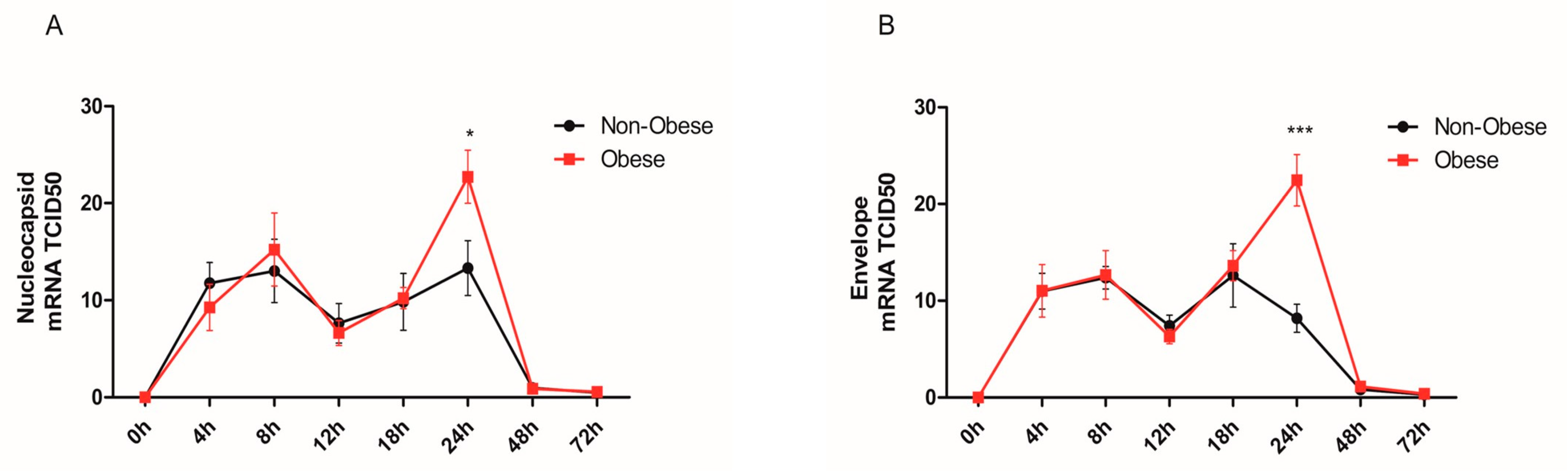
2.1. Increased Viral Detection and Replicates in Bronchial Epithelial Cells from Obese Subjects
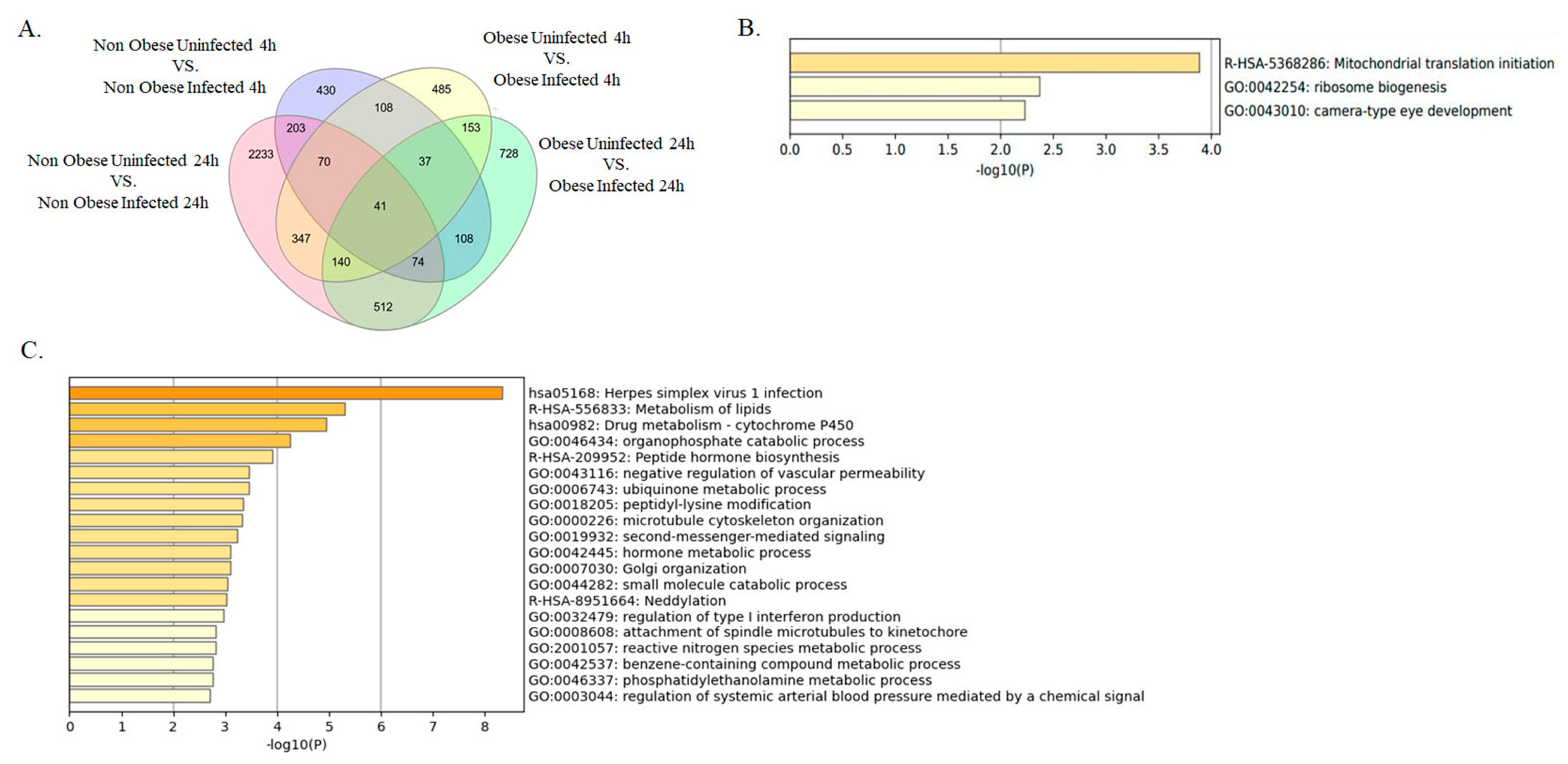
2.2. Enrichment of Lipid Metabolism-Related Pathways in Infected Bronchial Epithelial Cells from Obese Subjects
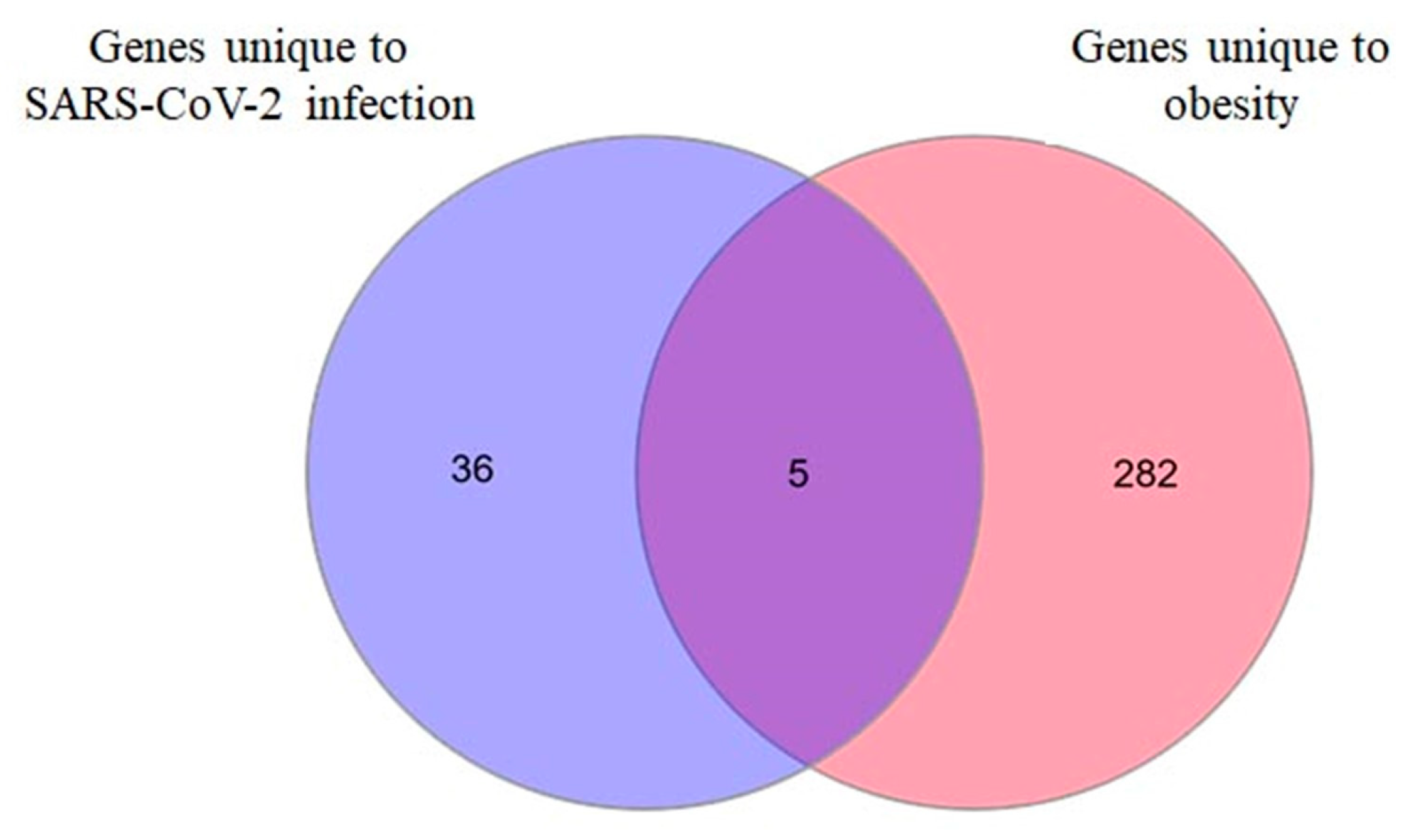
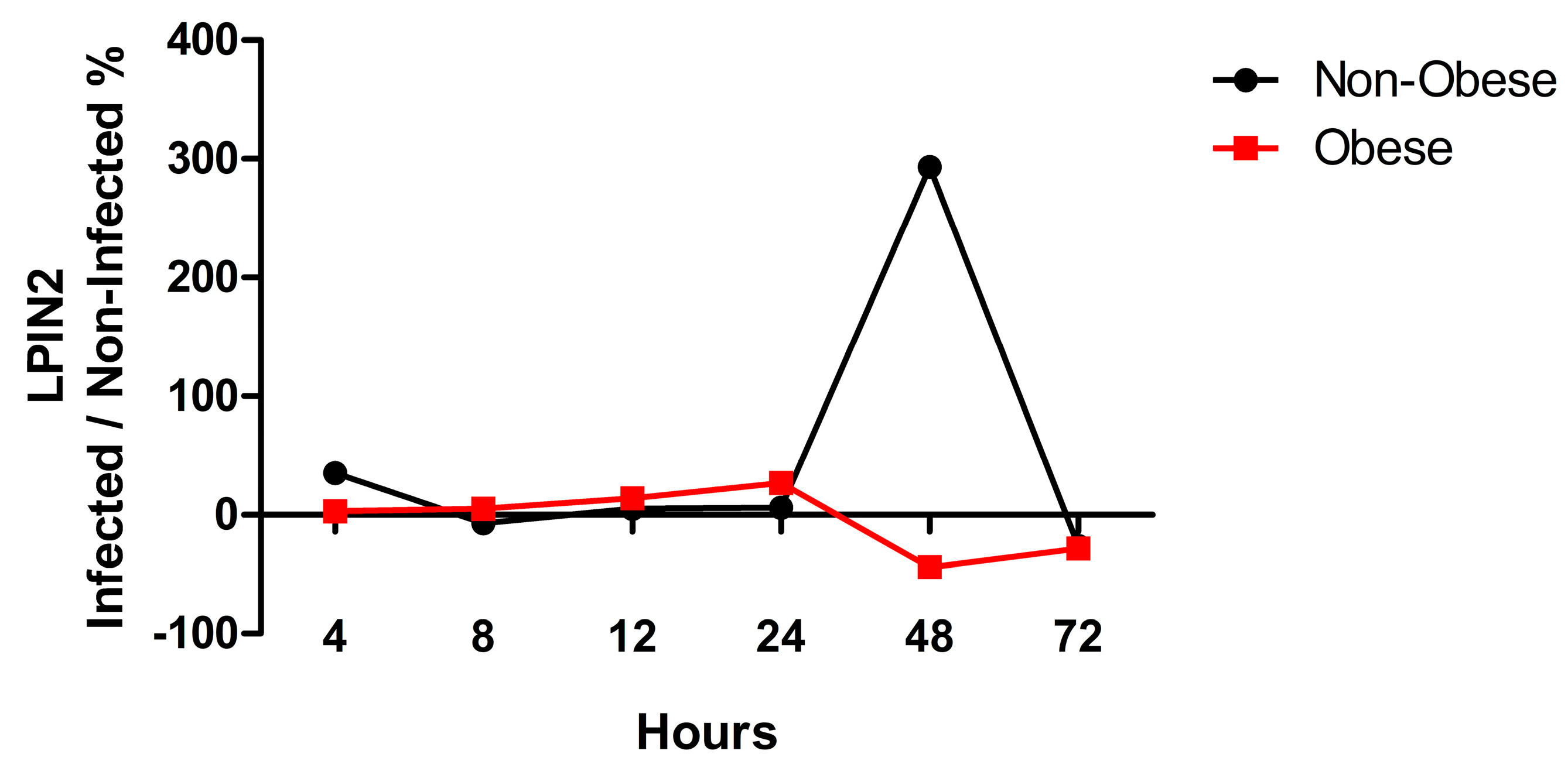
2.3. LPIN2 as a Unique DEG in Infected Bronchial Epithelial Cells from Obese Subjects
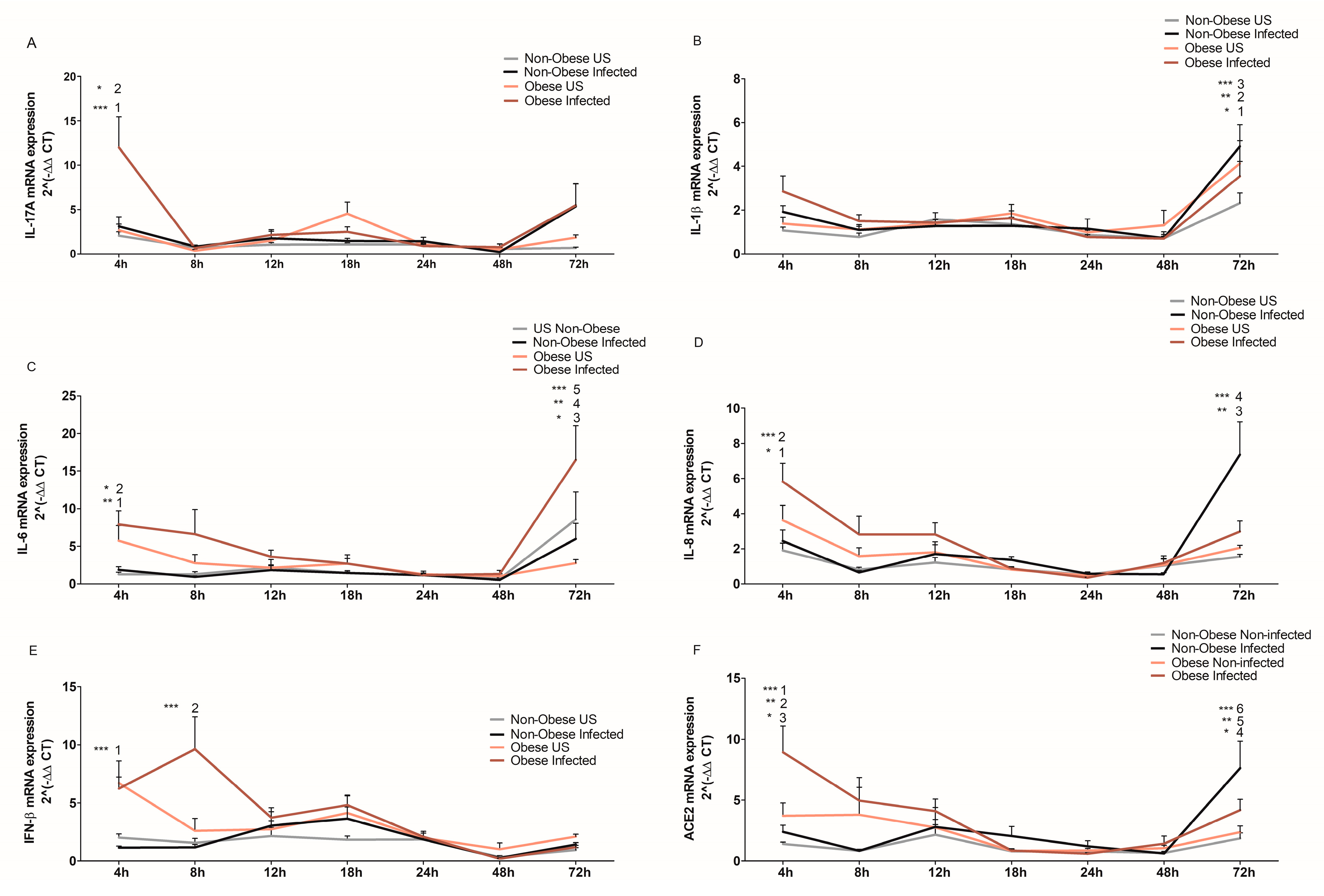
2.4. Bronchial Epithelial Cells from Obese Subjects Mount a Stronger Cytokine Response upon SARS-CoV-2 Infection at Early Stages of Infection
2.5. Upregulated ACE2 Expression in Bronchial Epithelial Cells from Obese Subjects
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. SARS-CoV-2 Infection of NHBE Cells
4.3. RNA Extraction
4.4. RNA Sequencing
4.5. Identification of Differentially Expressed Genes (DEGs)
4.6. Inflammatory Marker Quantification Using qRT-PCR
4.7. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, Y.; Lu, Y.; Huang, Y.-M.; Wang, M.; Ling, W.; Sui, Y.; Zhao, H.-L.J.M. Obesity in patients with COVID-19: A systematic review and meta-analysis. Metabolism 2020, 113, 154378. [Google Scholar] [CrossRef]
- Division of Nutrition, Physical Activity, and Obesity, National Center for Chronic Disease Prevention and Health Promotion. Adult Obesity Facts. 2022. Available online: https://www.cdc.gov/obesity/data/adult.html (accessed on 24 May 2022).
- National Institute of Diabetes and Digestive and Kidney Diseases. Overweight & Obesity Statistics. 2021. Available online: https://www.niddk.nih.gov/health-information/health-statistics/overweight-obesity (accessed on 24 May 2022).
- Simonnet, A.; Chetboun, M.; Poissy, J.; Raverdy, V.; Noulette, J.; Duhamel, A.; Labreuche, J.; Mathieu, D.; Pattou, F.; Jourdain, M.J.O. High prevalence of obesity in severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) requiring invasive mechanical ventilation. Obesity 2020, 28, 1195–1199. [Google Scholar] [CrossRef]
- Zheng, K.I.; Gao, F.; Wang, X.-B.; Sun, Q.-F.; Pan, K.-H.; Wang, T.-Y.; Ma, H.-L.; Chen, Y.-P.; Liu, W.-Y.; George, J.J.M. Obesity as a risk factor for greater severity of COVID-19 in patients with metabolic associated fatty liver disease. Metabolism 2020, 108, 154244. [Google Scholar] [CrossRef]
- Moustafa, J.S.E.-S.; Jackson, A.U.; Brotman, S.M.; Guan, L.; Villicaña, S.; Roberts, A.L.; Zito, A.; Bonnycastle, L.; Erdos, M.R.; Narisu, N.J.m. ACE2 expression in adipose tissue is associated with COVID-19 cardio-metabolic risk factors and cell type composition. medRxiv 2020. [Google Scholar] [CrossRef]
- Al Heialy, S.; Hachim, M.Y.; Senok, A.; Gaudet, M.; Abou Tayoun, A.; Hamoudi, R.; Alsheikh-Ali, A.; Hamid, Q.J. Regulation of angiotensin-converting enzyme 2 in obesity: Implications for COVID-19. J. Front. Physiol. 2020, 11, 555039. [Google Scholar] [CrossRef]
- Jalaleddine, N.; Bouzid, A.; Hachim, M.; Sharif-Askari, N.S.; Mahboub, B.; Senok, A.; Halwani, R.; Hamoudi, R.A.; Al Heialy, S. ACE2 polymorphisms impact COVID-19 severity in obese patients. Sci. Rep. 2022, 12, 21491. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, Y.; Zhang, F.; Wang, Q.; Li, T.; Liu, Z.; Wang, J.; Qin, Y.; Zhang, X.; Yan, X.J. The use of anti-inflammatory drugs in the treatment of people with severe coronavirus disease 2019 (COVID-19): The Perspectives of clinical immunologists from China. Clin. Immunol. 2020, 214, 108393. [Google Scholar] [CrossRef]
- Hamer, M.; Kivimäki, M.; Gale, C.R.; Batty, G.D. Lifestyle risk factors, inflammatory mechanisms, and COVID-19 hospitalization: A community-based cohort study of 387,109 adults in UK. Brain Behav. Immun. 2020, 87, 184–187. [Google Scholar] [CrossRef]
- McNeill, J.N.; Lau, E.S.; Paniagua, S.M.; Liu, E.E.; Wang, J.K.; Bassett, I.V.; Selvaggi, C.A.; Lubitz, S.A.; Foulkes, A.S.; Ho, J.E. The role of obesity in inflammatory markers in COVID-19 patients. Obes. Res. Clin. Pract. 2021, 15, 96–99. [Google Scholar] [CrossRef]
- Lordén, G.; Sanjuán-García, I.; de Pablo, N.; Meana, C.; Alvarez-Miguel, I.; Pérez-García, M.T.; Pelegrín, P.; Balsinde, J.; Balboa, M.A. Lipin-2 regulates NLRP3 inflammasome by affecting P2X7 receptor activation. J. Exp. Med. 2017, 214, 511–528. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Hu, X.-Y.; Zhao, Z.-P.; Guo, R.-L.; Guo, J.; Li, W.; Hao, C.-J.; Xu, B.-P. Compound heterozygous LPIN2 pathogenic variants in a patient with Majeed syndrome with recurrent fever and severe neutropenia: Case report. BMC Med. Genet. 2019, 20, 182. [Google Scholar] [CrossRef] [Green Version]
- Jiao, X.-L.; Jing, J.-J.; Qiao, L.-Y.; Liu, J.-H.; Li, L.-A.; Zhang, J.; Jia, X.-L.; Liu, W.-Z. Ontogenetic expression of Lpin2 and Lpin3 genes and their associations with traits in two breeds of Chinese fat-tailed sheep. Asian Australas. J. Anim. Sci. 2016, 29, 333. [Google Scholar] [CrossRef] [Green Version]
- Pelosi, M.; Testet, E.; Le Lay, S.; Dugail, I.; Tang, X.; Mabilleau, G.; Hamel, Y.; Madrange, M.; Blanc, T.; Odent, T.J. Normal human adipose tissue functions and differentiation in patients with biallelic LPIN1 inactivating mutations. J. Lipid Res. 2017, 58, 2348–2364. [Google Scholar] [CrossRef] [Green Version]
- Stoppelenburg, A.J.; Salimi, V.; Hennus, M.; Plantinga, M.; Veld, R.H.; Walk, J.; Meerding, J.; Coenjaerts, F.; Bont, L.; Boes, M. Local IL-17A potentiates early neutrophil recruitment to the respiratory tract during severe RSV infection. PLoS ONE 2013, 8, e78461. [Google Scholar] [CrossRef]
- Cakebread, J.A.; Xu, Y.; Grainge, C.; Kehagia, V.; Howarth, P.H.; Holgate, S.T.; Davies, D.E. Exogenous IFN-β has antiviral and anti-inflammatory properties in primary bronchial epithelial cells from asthmatic subjects exposed to rhinovirus. J. Allergy Clin. Immunol. 2011, 127, 1148–1154.e9. [Google Scholar] [CrossRef]
- Nakhlband, A.; Fakhari, A.; Azizi, H.J. Interferon-beta offers promising avenues to COVID-19 treatment: A systematic review and meta-analysis of clinical trial studies. Naunyn Schmiedeberg’s Arch. Pharmacol. 2021, 394, 829–838. [Google Scholar] [CrossRef]
- Zhang, P.; Shi, L.; Xu, J.; Wang, Y.; Yang, H. Elevated interleukin-6 and adverse outcomes in COVID-19 patients: A meta-analysis based on adjusted effect estimates. Immunogenetics 2020, 72, 431–437. [Google Scholar] [CrossRef]
- Liu, K.; Yang, T.; Peng, X.F.; Lv, S.M.; Ye, X.L.; Zhao, T.S.; Li, J.C.; Shao, Z.J.; Lu, Q.B.; Li, J.Y.; et al. A systematic meta-analysis of immune signatures in patients with COVID-19. Rev. Med. Virol. 2021, 31, e2195. [Google Scholar] [CrossRef]
- Ritter, A.; Kreis, N.N.; Louwen, F.; Yuan, J. Obesity and COVID-19: Molecular Mechanisms Linking Both Pandemics. Int. J. Mol. Sci. 2020, 21, 5793. [Google Scholar] [CrossRef]
- Jalaleddine, N.; Hachim, M.; Al-Hroub, H.; Saheb Sharif-Askari, N.; Senok, A.; Elmoselhi, A.; Mahboub, B.; Samuel Kurien, N.M.; Kandasamy, R.K.; Semreen, M.H.; et al. N6-Acetyl-L-Lysine and p-Cresol as Key Metabolites in the Pathogenesis of COVID-19 in Obese Patients. Front. Immunol. 2022, 13, 827603. [Google Scholar] [CrossRef]
- Higham, A.; Singh, D.J.O. Increased ACE2 expression in bronchial epithelium of COPD patients who are overweight. Obesity 2020, 28, 1586–1589. [Google Scholar] [CrossRef]
- Hulme, K.D.; Noye, E.C.; Short, K.R.; Labzin, L.I. Dysregulated Inflammation during Obesity: Driving Disease Severity in Influenza Virus and SARS-CoV-2 Infections. Front. Immunol. 2021, 12, 4356. [Google Scholar] [CrossRef]
- Caterino, M.; Gelzo, M.; Sol, S.; Fedele, R.; Annunziata, A.; Calabrese, C.; Fiorentino, G.; D’Abbraccio, M.; Dell’Isola, C.; Fusco, F.M. Dysregulation of lipid metabolism and pathological inflammation in patients with COVID-19. Sci. Rep. 2021, 11, 2941. [Google Scholar] [CrossRef]
- Yan, B.; Yuan, S.; Cao, J.; Fung, K.; Lai, P.-M.; Yin, F.; Sze, K.-H.; Qin, Z.; Xie, Y.; Ye, Z.-W. Phosphatidic acid phosphatase 1 impairs SARS-CoV-2 replication by affecting the glycerophospholipid metabolism pathway. Int. J. Biol. Sci. 2022, 18, 4744. [Google Scholar] [CrossRef]
- Xu, K.; Nagy, P.D. RNA virus replication depends on enrichment of phosphatidylethanolamine at replication sites in subcellular membranes. Proc. Natl. Acad. Sci. USA 2015, 112, E1782–E1791. [Google Scholar] [CrossRef] [Green Version]
- Yan, T.; Xiao, R.; Wang, N.; Shang, R.; Lin, G.J.T. Obesity and severe coronavirus disease 2019: Molecular mechanisms, paths forward, and therapeutic opportunities. J. Theranostics 2021, 11, 8234. [Google Scholar] [CrossRef]
- Honce, R.; Schultz-Cherry, S.J. Impact of obesity on influenza A virus pathogenesis, immune response, and evolution. J. Front. Immunol. 2019, 10, 1071. [Google Scholar] [CrossRef] [Green Version]
- Terán-Cabanillas, E.; Hernández, J.J. Role of leptin and SOCS3 in inhibiting the type I interferon response during obesity. J. Inflamm. 2017, 40, 58–67. [Google Scholar] [CrossRef]
- Kanneganti, T.-D.; Dixit, V.D. Immunological complications of obesity. J. Nat. Immunol. 2012, 13, 707–712. [Google Scholar] [CrossRef]
- Channappanavar, R.; Fehr, A.R.; Vijay, R.; Mack, M.; Zhao, J.; Meyerholz, D.K.; Perlman, S. Dysregulated Type I Interferon and Inflammatory Monocyte-Macrophage Responses Cause Lethal Pneumonia in SARS-CoV-Infected Mice. Cell Host Microbe 2016, 19, 181–193. [Google Scholar] [CrossRef] [Green Version]
- Terán-Cabanillas, E.; Montalvo-Corral, M.; Silva-Campa, E.; Caire-Juvera, G.; Moya-Camarena, S.Y.; Hernández, J.J. Production of interferon α and β, pro-inflammatory cytokines and the expression of suppressor of cytokine signaling (SOCS) in obese subjects infected with influenza A/H1N1. J. Clin. Nutr. 2014, 33, 922–926. [Google Scholar] [CrossRef]
- Smith, A.G.; Sheridan, P.A.; Harp, J.B.; Beck, M.A. Diet-induced obese mice have increased mortality and altered immune responses when infected with influenza virus. J. Nutr. 2007, 137, 1236–1243. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Xu, Y.; Zhang, X.; Wang, S.; Peng, Z.; Guo, J.; Jiang, H.; Liu, J.; Xie, Y.; Wang, J.J. Leptin correlates with monocytes activation and severe condition in COVID-19 patients. J. Leukoc. Biol. 2021, 110, 9–20. [Google Scholar] [CrossRef]
- Tsatsanis, C.; Margioris, A.; Kontoyiannis, D.J. Association between H1N1 infection severity and obesity—Adiponectin as a potential etiologic factor. J. Infect. Dis. 2010, 202, 459–460. [Google Scholar] [CrossRef] [Green Version]
- Richter, F.C.; Alrubayyi, A.; Teijeira Crespo, A.; Consortium, O.-C.C.-L.; Hulin-Curtis, S.J. Impact of obesity and SARS-CoV-2 infection: Implications for host defence—A living review. Oxf. Open Immunol. 2021, 2, iqab001. [Google Scholar] [CrossRef]
- Ni, W.; Yang, X.; Yang, D.; Bao, J.; Li, R.; Xiao, Y.; Hou, C.; Wang, H.; Liu, J.; Yang, D.J. Role of angiotensin-converting enzyme 2 (ACE2) in COVID-19. J. Crit. Care 2020, 24, 422. [Google Scholar] [CrossRef]
- Brandolini, M.; Taddei, F.; Marino, M.M.; Grumiro, L.; Scalcione, A.; Turba, M.E.; Gentilini, F.; Fantini, M.; Zannoli, S.; Dirani, G.; et al. Correlating qRT-PCR, dPCR and Viral Titration for the Identification and Quantification of SARS-CoV-2: A New Approach for Infection Management. Viruses 2021, 13, 1022. [Google Scholar] [CrossRef]
- Emig, D.; Salomonis, N.; Baumbach, J.; Lengauer, T.; Conklin, B.R.; Albrecht, M. AltAnalyze and DomainGraph: Analyzing and visualizing exon expression data. Nucleic Acids Res. 2010, 38, W755–W762. [Google Scholar] [CrossRef] [Green Version]
- Takele Assefa, A.; Vandesompele, J.; Thas, O. On the utility of RNA sample pooling to optimize cost and statistical power in RNA sequencing experiments. BMC Genom. 2020, 21, 312. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Zaim, S.R.; Aberasturi, D.; Berghout, J.; Li, H.; Vitali, F.; Kenost, C.; Zhang, H.H.; Lussier, Y.A. Interpretation of ′Omics dynamics in a single subject using local estimates of dispersion between two transcriptomes. AMIA Annu. Symp. Proc. 2020, 2019, 582–591. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-Obese | Obese | |
|---|---|---|
| N | 4 | 4 |
| Age, y | 42.3 ± 7.5 | 32 ± 6.6 |
| BMI, kg/m2 | 23.6 ± 5.3 | 37.1 ± 2.8 |
| Sex (Male/female) | 2/2 | 3/1 |
| Primer Name | Oligo Sequence (5′ to 3′) |
|---|---|
| SARS-CoV2 N-F | AAGCTGGACTTCCCTATGGTG |
| SARS-CoV2 N-R | CGATTGCAGCATTGTTAGCAGG |
| SARS2-UpE-F | ATTGTTGATGAGCCTGAAG |
| SARS2-UpE-R | TTCGTACTCATCAGCTTG |
| ACE2-F | TACTGTGACCCCGCATCTCT |
| ACE2-R | TCCAACAGTTTCTGTCCAGC |
| IL-1β-F | TACATCAGCACCTCTCAAGCA |
| IL-1β-R | CCACATTCAGCACAGGACTCT |
| GAPDH-F | GAAGGTGAAGGTCGGAGT |
| GAPDH-R | GAAGATGGTGATGGGATTTC |
| IL-17A-F | GAGGACAAGAACTTCCCCCG |
| IL-17A-R | CATTGCCGTGGAGATTCCAAG |
| IFN-β-F | CTTGGATTCCTACAAAGAAGCAGC |
| IFN-β-R | TCCTCCTTCTGGAACTGCTGCA |
| IL-8-F | TCTGCAGCTCTGTGTGAAGGTG |
| IL-8-R | AATTTCTGTGTTGGCGCAGTG |
| IL-6-F | ACCTTCCAAAGATGGCTGAAA |
| IL-6-R | GCTCTGGCTTGTTCCCTCACTAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaudet, M.; Kaufmann, E.; Jalaleddine, N.; Mogas, A.; Hachim, M.; Senok, A.; Divangahi, M.; Hamid, Q.; Al Heialy, S. Lung Epithelial Cells from Obese Patients Have Impaired Control of SARS-CoV-2 Infection. Int. J. Mol. Sci. 2023, 24, 6729. https://doi.org/10.3390/ijms24076729
Gaudet M, Kaufmann E, Jalaleddine N, Mogas A, Hachim M, Senok A, Divangahi M, Hamid Q, Al Heialy S. Lung Epithelial Cells from Obese Patients Have Impaired Control of SARS-CoV-2 Infection. International Journal of Molecular Sciences. 2023; 24(7):6729. https://doi.org/10.3390/ijms24076729
Chicago/Turabian StyleGaudet, Mellissa, Eva Kaufmann, Nour Jalaleddine, Andrea Mogas, Mahmood Hachim, Abiola Senok, Maziar Divangahi, Qutayba Hamid, and Saba Al Heialy. 2023. "Lung Epithelial Cells from Obese Patients Have Impaired Control of SARS-CoV-2 Infection" International Journal of Molecular Sciences 24, no. 7: 6729. https://doi.org/10.3390/ijms24076729