
Multi-Region Microdialysis Imaging Platform Revealed Dorsal Raphe Nucleus Calcium Signaling and Serotonin Dynamics during Nociceptive Pain

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Formalin-Induced Pain Increased Calcium Fluorescence Intensity in the DRN
2.2. Formalin-Induced Pain Increased Serotonin Levels in the CeA and the ACC
2.3. Formalin-Induced Pain Generated Lateral Responses in the DRN, CeA, and ACC
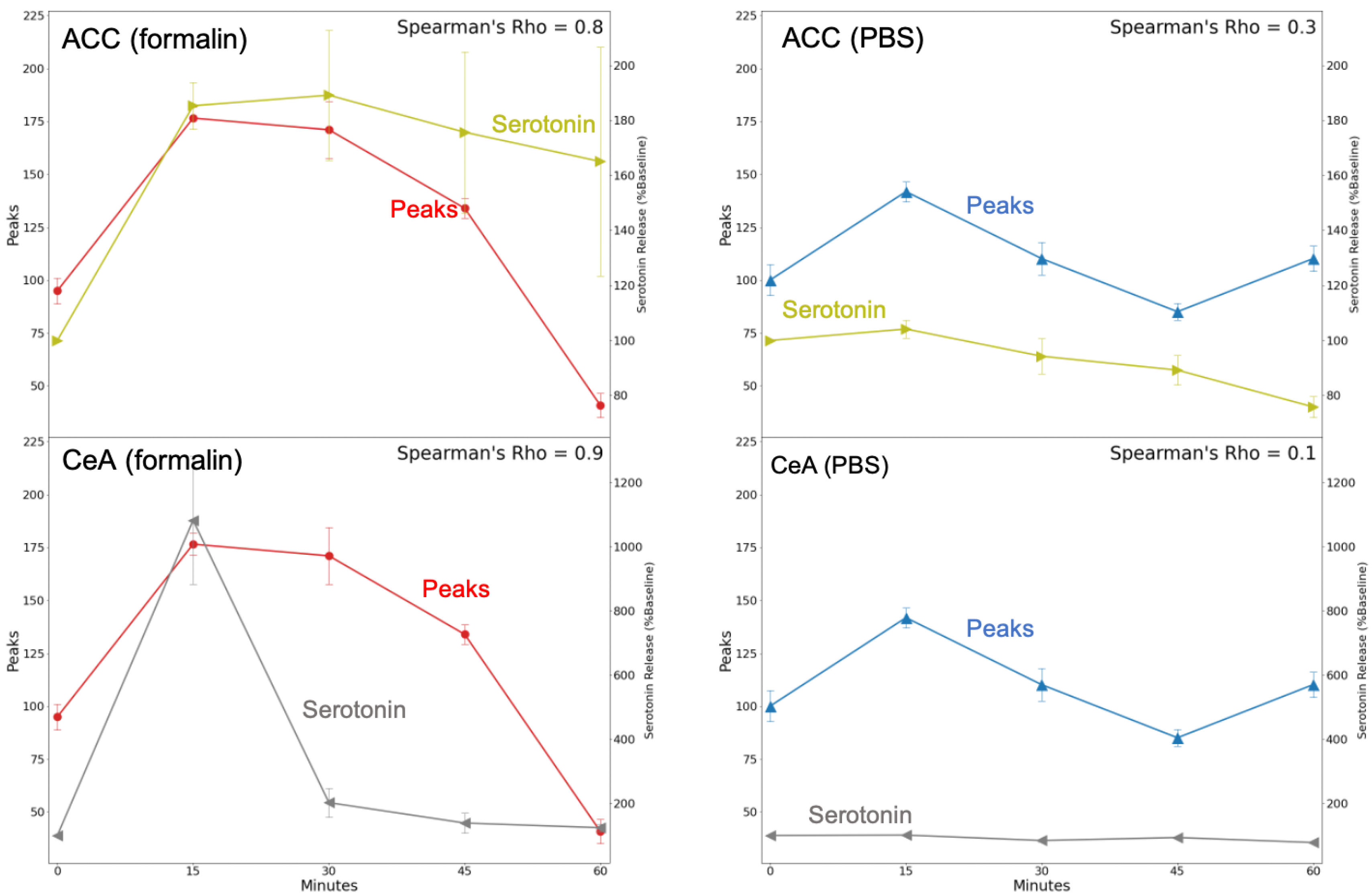
2.4. Calcium Fluorescence Intensity and Serotonin Release Were Correlated after Formalin Injection
2.5. Calcium Fluorescence Intensity and Licking Behavior Were Correlated after Formalin Injection
3. Discussion
3.1. CMOS Implantable Device for Probing Neural Activity
3.2. Biphasic Response in the DRN Imaging during Nociception
3.3. Microdialysis Imaging Platform for Investigating Pain in the Central Nervous System
3.4. Laterality in Nociceptive Pain Processing
4. Materials and Methods
4.1. Imaging Device Fabrication
4.2. Animal Surgery and Device Implantation
4.3. Imaging and Microdialysis Experiment
4.4. Pain Stimulation
4.5. Brain Slicing and Immunostaining
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gilam, G.; Gross, J.J.; Wager, T.D.; Keefe, F.J.; Mackey, S.C. What Is the Relationship between Pain and Emotion? Bridging Constructs and Communities. Neuron 2020, 107, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Voscopoulos, C.; Lema, M. When Does Acute Pain Become Chronic? Br. J. Anaesth. 2010, 105, i69–i85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, S.P.; Vase, L.; Hooten, W.M. Chronic Pain: An Update on Burden, Best Practices, and New Advances. Lancet 2021, 397, 2082–2097. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Larrea, L.; Peyron, R. Pain Matrices and Neuropathic Pain Matrices: A Review. Pain 2013, 154 (Suppl. S1), S29–S43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legrain, V.; Iannetti, G.D.; Plaghki, L.; Mouraux, A. The Pain Matrix Reloaded: A Salience Detection System for the Body. Prog. Neurobiol. 2011, 93, 111–124. [Google Scholar] [CrossRef] [Green Version]
- Talbot, K.; Madden, V.J.; Jones, S.L.; Moseley, G.L. The Sensory and Affective Components of Pain: Are They Differentially Modifiable Dimensions or Inseparable Aspects of a Unitary Experience? A Systematic Review. Br. J. Anaesth. 2019, 123, e263–e272. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, Z.; Zamponi, G.W. Pain: Integration of Sensory and Affective Aspects of Pain. Curr. Biol. 2020, 30, R393–R395. [Google Scholar] [CrossRef]
- Chen, T.W.; Wardill, T.J.; Sun, Y.; Pulver, S.R.; Renninger, S.L.; Baohan, A.; Schreiter, E.R.; Kerr, R.A.; Orger, M.B.; Jayaraman, V.; et al. Ultra-Sensitive Fluorescent Proteins for Imaging Neuronal Activity. Nature 2013, 499, 295. [Google Scholar] [CrossRef] [Green Version]
- Aharoni, D.; Hoogland, T.M. Circuit Investigations with Open-Source Miniaturized Microscopes: Past, Present and Future. Front. Cell. Neurosci. 2019, 13, 141. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, K.K.; Burns, L.D.; Cocker, E.D.; Nimmerjahn, A.; Ziv, Y.; Gamal, A.E.; Schnitzer, M.J. Miniaturized Integration of a Fluorescence Microscope. Nat. Methods 2011, 8, 871–878. [Google Scholar] [CrossRef] [Green Version]
- Klioutchnikov, A.; Wallace, D.J.; Frosz, M.H.; Zeltner, R.; Sawinski, J.; Pawlak, V.; Voit, K.M.; Russell, P.S.J.; Kerr, J.N.D. Three-Photon Head-Mounted Microscope for Imaging Deep Cortical Layers in Freely Moving Rats. Nat. Methods 2020, 17, 509–513. [Google Scholar] [CrossRef]
- Zong, W.; Wu, R.; Chen, S.; Wu, J.; Wang, H.; Zhao, Z.; Chen, G.; Tu, R.; Wu, D.; Hu, Y.; et al. Miniature Two-Photon Microscopy for Enlarged Field-of-View, Multi-Plane and Long-Term Brain Imaging. Nat. Methods 2021, 18, 46–49. [Google Scholar] [CrossRef]
- Ohta, J.; Ohta, Y.; Takehara, H.; Noda, T.; Sasagawa, K.; Tokuda, T.; Haruta, M.; Kobayashi, T.; Akay, Y.M.; Akay, M. Implantable Microimaging Device for Observing Brain Activities of Rodents. Proc. IEEE 2017, 105, 158–166. [Google Scholar] [CrossRef]
- Rebusi, R.; Olorocisimo, J.P.; Briones, J.; Ohta, Y.; Haruta, M.; Takehara, H.; Tashiro, H.; Sasagawa, K.; Ohta, J. Simultaneous CMOS-Based Imaging of Calcium Signaling of the Central Amygdala and the Dorsal Raphe Nucleus during Nociception in Freely Moving Mice. Front. Neurosci. 2021, 15, 582. [Google Scholar] [CrossRef]
- Sunaga, Y.; Ohta, Y.; Akay, Y.M.; Ohta, J.; Akay, M. Monitoring Neural Activities in the VTA in Response to Nicotine Intake Using a Novel Implantable Microimaging Device. IEEE Access 2020, 8, 68013–68020. [Google Scholar] [CrossRef]
- Paredes, S.; Cantillo, S.; Candido, K.D.; Knezevic, N.N. An Association of Serotonin with Pain Disorders and Its Modulation by Estrogens. Int. J. Mol. Sci. 2019, 20, 5729. [Google Scholar] [CrossRef] [Green Version]
- Butkevich, I.P.; Mikhailenko, V.A.; Leont’eva, M.N. Sequelae of Prenatal Serotonin Depletion and Stress on Pain Sensitivity in Rats. Neurosci. Behav. Physiol. 2005, 35, 925–930. [Google Scholar] [CrossRef]
- Huang, K.W.; Ochandarena, N.E.; Philson, A.C.; Hyun, M.; Birnbaum, J.E.; Cicconet, M.; Sabatini, B.L. Molecular and Anatomical Organization of the Dorsal Raphe Nucleus. Elife 2019, 8, e46464. [Google Scholar] [CrossRef]
- Šimi, G.; Tkalči, M.; Vuki, V.; Mulc, D.; Špani, E.; Šagud, M.; Olucha-Bordonau, F.E.; Vukši, M.; Hof, P.R.; Uversky, N. Understanding Emotions: Origins and Roles of the Amygdala. Biomolecules 2021, 11, 823. [Google Scholar] [CrossRef]
- Neugebauer, V.; Li, W.; Bird, G.C.; Han, J.S. The Amygdala and Persistent Pain. Neuroscientist 2004, 10, 221–234. [Google Scholar] [CrossRef]
- Ressler, K.J. Amygdala Activity, Fear, and Anxiety: Modulation by Stress. Biol. Psychiatry 2010, 67, 1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, J.P.; Siemer, M.; Gotlib, I.H. Amygdala Volume in Major Depressive Disorder: A Meta-Analysis of Magnetic Resonance Imaging Studies. Mol. Psychiatry 2008, 13, 993–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, P.N.; Peng, Y.B.; Boyette-Davis, J.A.; Uhelski, M.L. The Anterior Cingulate Cortex and Pain Processing. Front. Integr. Neurosci. 2014, 8, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shackman, A.J.; Salomons, T.V.; Slagter, H.A.; Fox, A.S.; Winter, J.J.; Davidson, R.J. The Integration of Negative Affect, Pain and Cognitive Control in the Cingulate Cortex. Nat. Rev. Neurosci. 2011, 12, 154–167. [Google Scholar] [CrossRef]
- Hunskaar, S.; Hole, K. The Formalin Test in Mice: Dissociation between Inflammatory and Non-Inflammatory Pain. Pain 1987, 30, 103–114. [Google Scholar] [CrossRef]
- Murray, C.W.; Porreca, F.; Cowan, A. Methodological Refinements to the Mouse Paw Formalin Test. An Animal Model of Tonic Pain. J. Pharmacol. Methods 1988, 20, 175–186. [Google Scholar] [CrossRef]
- Ohkura, M.; Sasaki, T.; Sadakari, J.; Gengyo-Ando, K.; Kagawa-Nagamura, Y.; Kobayashi, C.; Ikegaya, Y.; Nakai, J. Genetically Encoded Green Fluorescent Ca2+ Indicators with Improved Detectability for Neuronal Ca2+ Signals. PLoS ONE 2012, 7, e51286. [Google Scholar] [CrossRef] [Green Version]
- Hartung, J.E.; Gold, M.S. GCaMP as an Indirect Measure of Electrical Activity in Rat Trigeminal Ganglion Neurons. Cell Calcium 2020, 89, 102225. [Google Scholar] [CrossRef]
- Takehara, H.; Ohta, Y.; Motoyama, M.; Haruta, M.; Nagasaki, M.; Takehara, H.; Noda, T.; Sasagawa, K.; Tokuda, T.; Ohta, J. Intravital Fluorescence Imaging of Mouse Brain Using Implantable Semiconductor Devices and Epi-Illumination of Biological Tissue. Biomed. Opt. Express 2015, 6, 1553. [Google Scholar] [CrossRef] [Green Version]
- Kulmala, N.; Sasagawa, K.; Treepetchkul, T.; Takehara, H.; Haruta, M.; Tashiro, H.; Ohta, J. Lensless Dual-Color Fluorescence Imaging Device Using Hybrid Filter. Jpn. J. Appl. Phys. 2022, 61, SC1020. [Google Scholar] [CrossRef]
- Sasagawa, K.; Kimura, A.; Haruta, M.; Noda, T.; Tokuda, T.; Ohta, J. Highly Sensitive Lens-Free Fluorescence Imaging Device Enabled by a Complementary Combination of Interference and Absorption Filters. Biomed. Opt. Express 2018, 9, 4329–4344. [Google Scholar] [CrossRef]
- Azmer, M.I.; Sasagawa, K.; Rustami, E.; Sugie, K.; Ohta, Y.; Haruta, M.; Takehara, H.; Tashiro, H.; Ohta, J. Miniaturized LED Light Source with an Excitation Filter for Fluorescent Imaging. Jpn. J. Appl. Phys. 2021, 60, SBBG07. [Google Scholar] [CrossRef]
- Rustami, E.; Sasagawa, K.; Sugie, K.; Ohta, Y.; Haruta, M.; Noda, T.; Tokuda, T.; Ohta, J. Needle-Type Imager Sensor with Band-Pass Composite Emission Filter and Parallel Fiber-Coupled Laser Excitation. IEEE Trans. Circuits Syst. I Regul. Pap. 2020, 67, 1082–1091. [Google Scholar] [CrossRef]
- Olorocisimo, J.P.; Briones, J.; Sasagawa, K.; Haruta, M.; Takehara, H.; Tashiro, H.; Ishida-Kitagawa, N.; Bessho, Y.; Ohta, J. Ultrasmall Compact CMOS Imaging System for Bioluminescence Reporter-Based Live Gene Expression Analysis. J. Biomed. Opt. 2021, 26, 116002. [Google Scholar] [CrossRef]
- Sugie, K.; Sasagawa, K.; Guinto, M.C.; Haruta, M.; Tokuda, T.; Ohta, J. Implantable CMOS Image Sensor with Incident-Angle-Selective Pixels. Electron. Lett. 2019, 55, 729–731. [Google Scholar] [CrossRef]
- Zong, W.; Obenhaus, H.A.; Skytøen, E.R.; Eneqvist, H.; de Jong, N.L.; Vale, R.; Jorge, M.R.; Moser, M.B.; Moser, E.I. Large-Scale Two-Photon Calcium Imaging in Freely Moving Mice. Cell 2022, 185, 1240–1256.e30. [Google Scholar] [CrossRef]
- Li, Y.; Zhong, W.; Wang, D.; Feng, Q.; Liu, Z.; Zhou, J.; Jia, C.; Hu, F.; Zeng, J.; Guo, Q.; et al. Serotonin Neurons in the Dorsal Raphe Nucleus Encode Reward Signals. Nat. Commun. 2016, 7, 10503. [Google Scholar] [CrossRef] [Green Version]
- Matthews, G.A.; Nieh, E.H.; Vander Weele, C.M.; Halbert, S.A.; Pradhan, R.V.; Yosafat, A.S.; Glober, G.F.; Izadmehr, E.M.; Thomas, R.E.; Lacy, G.D.; et al. Dorsal Raphe Dopamine Neurons Represent the Experience of Social Isolation. Cell 2016, 164, 617–631. [Google Scholar] [CrossRef] [Green Version]
- Seo, C.; Guru, A.; Jin, M.; Ito, B.; Sleezer, B.J.; Ho, Y.Y.; Wang, E.; Boada, C.; Krupa, N.A.; Kullakanda, D.S.; et al. Intense Threat Switches Dorsal Raphe Serotonin Neurons to a Paradoxical Operational Mode. Science 2019, 363, 539–542. [Google Scholar] [CrossRef]
- Paquelet, G.E.; Carrion, K.; Lacefield, C.O.; Zhou, P.; Hen, R.; Miller, B.R. Single-Cell Activity and Network Properties of Dorsal Raphe Nucleus Serotonin Neurons during Emotionally Salient Behaviors. Neuron 2022, 110, 2664–2679.e8. [Google Scholar] [CrossRef]
- Qing-Ping, W.; Nakai, Y. The Dorsal Raphe: An Important Nucleus in Pain Modulation. Brain Res. Bull. 1994, 34, 575–585. [Google Scholar] [CrossRef] [PubMed]
- López-Cano, M.; Fernández-Dueñas, V.; Llebaria, A.; Ciruela, F. Formalin Murine Model of Pain. Bio-Protocol 2017, 7, e2628. [Google Scholar] [CrossRef] [PubMed]
- Dubuisson, D.; Dennis, S.G. The Formalin Test: A Quantitative Study of the Analgesic Effects of Morphine, Meperidine, and Brain Stem Stimulation in Rats and Cats. Pain 1977, 4, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Zouikr, I.; Tadros, M.A.; Clifton, V.L.; Beagley, K.W.; Hodgson, D.M. Low Formalin Concentrations Induce Fine-Tuned Responses That Are Sex and Age-Dependent: A Developmental Study. PLoS ONE 2013, 8, e53384. [Google Scholar] [CrossRef] [Green Version]
- Hwang, H.J.; Kim, P.; Kim, C.J.; Lee, H.J.; Shim, I.; Yin, C.S.; Yang, Y.; Hahm, D.H. Antinociceptive Effect of Amygdalin Isolated from Prunus Armeniaca on Formalin-Induced Pain in Rats. Biol. Pharm. Bull. 2008, 31, 1559–1564. [Google Scholar] [CrossRef] [Green Version]
- Mcnamara, C.R.; Mandel-Brehm, J.; Bautista, D.M.; Siemens, J.; Deranian, K.L.; Zhao, M.; Hayward, N.J.; Chong, J.A.; Julius, D.; Moran, M.M.; et al. TRPA1 Mediates Formalin-Induced Pain. Proc. Natl. Acad. Sci. USA 2007, 14, 13525–13530. [Google Scholar] [CrossRef] [Green Version]
- Mohammad-Zadeh, M.; Azhdari-Zarmehri, H.; Mosavi, F.; Haghdoost-Yazdi, H.; Nazeri, M.; Shabani, M. Modulation of Different Phases of Formalin Test by Force Swim Stress. Basic Clin. Neurosci. 2014, 5, 303. [Google Scholar]
- Henry, J.L.; Yashpal, K.; Pitcher, G.M.; Chabot, J.; Coderre, T.J. Evidence for Tonic Activation of NK-1 Receptors during the Second Phase of the Formalin Test in the Rat. J. Neurosci. 1999, 19, 6588–6598. [Google Scholar] [CrossRef] [Green Version]
- Rosland, J.H.; Tjølsen, A.; Mæhle, B.; Hole, K. The Formalin Test in Mice: Effect of Formalin Concentration. Pain 1990, 42, 235–242. [Google Scholar] [CrossRef]
- Tjølsen, A.; Berge, O.G.; Hunskaar, S.; Rosland, J.H.; Hole, K. The Formalin Test: An Evaluation of the Method. Pain 1992, 51, 5–17. [Google Scholar] [CrossRef]
- Abbott, F.V.; Franklin, K.B.J.; Westbrook, R.F. The Formalin Test: Scoring Properties of the First and Second Phases of the Pain Response in Rats. Pain 1995, 60, 91–102. [Google Scholar] [CrossRef]
- Taylor, B.K.; Peterson, M.A.; Basbaumi, A.I. Persistent Cardiovascular and Behavioral Nociceptive Responses to Subcutaneous Formalin Require Peripheral Nerve Input. J. Neurosci. 1995, 15, 7575–7584. [Google Scholar] [CrossRef]
- Shields, S.D.; Cavanaugh, D.J.; Lee, H.; Anderson, D.J.; Basbaum, A.I. Pain Behavior in the Formalin Test Persists after Ablation of the Great Majority of C-Fiber Nociceptors. Pain 2010, 151, 422–429. [Google Scholar] [CrossRef] [Green Version]
- Chisholm, K.I.; Khovanov, N.; Lopes, D.M.; la Russa, F.; McMahon, S.B. Large Scale In Vivo Recording of Sensory Neuron Activity with GCaMP6. eNeuro 2018, 5, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Marinelli, S.; Schnell, S.A.; Hack, S.P.; Christie, M.J.; Wessendorf, M.W.; Vaughan, C.W. Serotonergic and Nonserotonergic Dorsal Raphe Neurons Are Pharmacologically and Electrophysiologically Heterogeneous. J. Neurophysiol. 2004, 92, 3532–3537. [Google Scholar] [CrossRef] [Green Version]
- Heijmans, L.; Mons, M.R.; Joosten, E.A. A Systematic Review on Descending Serotonergic Projections and Modulation of Spinal Nociception in Chronic Neuropathic Pain and after Spinal Cord Stimulation. Mol. Pain 2021, 17, 17448069211043965. [Google Scholar] [CrossRef]
- Ishimura, K.; Takeuchi, Y.; Fujiwara, K.; Tominaga, M.; Yoshioka, H.; Sawada, T. Quantitative Analysis of the Distribution of Serotonin-Immunoreactive Cell Bodies in the Mouse Brain. Neurosci. Lett. 1988, 91, 265–270. [Google Scholar] [CrossRef]
- Garland, E.L. Pain Processing in the Human Nervous System: A Selective Review of Nociceptive and Biobehavioral Pathways. Prim. Care 2012, 39, 561. [Google Scholar] [CrossRef] [Green Version]
- Telner, J.; Lepore, F.; Guillemot, J.P. Effects of Serotonin Content on Pain Sensitivity in the Rat. Pharmacol. Biochem. Behav. 1979, 10, 657–661. [Google Scholar] [CrossRef]
- Marks, D.; Shah, M.; Patkar, A.; Masand, P.; Park, G.-Y.; Pae, C.-U. Serotonin-Norepinephrine Reuptake Inhibitors for Pain Control: Premise and Promise. Curr. Neuropharmacol. 2009, 7, 331–336. [Google Scholar] [CrossRef] [Green Version]
- Bravo, L.; Llorca-Torralba, M.; Berrocoso, E.; Micó, J.A. Monoamines as Drug Targets in Chronic Pain: Focusing on Neuropathic Pain. Front. Neurosci. 2019, 13, 1268. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.Y.; Morrow, K.; Bonacquisti, E.; Zhang, W.Y.; Shah, D.K. Antibody Pharmacokinetics in Rat Brain Determined Using Microdialysis. MAbs 2018, 10, 843–853. [Google Scholar] [CrossRef]
- Forster, G.L.; Feng, N.; Watt, M.J.; Korzan, W.J.; Mouw, N.J.; Summers, C.H.; Renner, K.J. Corticotropin-Releasing Factor in the Dorsal Raphe Elicits Temporally Distinct Serotonergic Responses in the Limbic System in Relation to Fear Behavior. Neuroscience 2006, 141, 1047–1055. [Google Scholar] [CrossRef] [PubMed]
- Mo, B.; Feng, N.; Renner, K.; Forster, G. Restraint Stress Increases Serotonin Release in the Central Nucleus of the Amygdala via Activation of Corticotropin-Releasing Factor Receptors. Brain Res. Bull. 2008, 76, 493–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Scholl, J.L.; Tu, W.; Hassell, J.E.; Watt, M.J.; Forster, G.L.; Renner, K.J. Serotonergic Responses to Stress Are Enhanced in the Central Amygdala and Inhibited in the Ventral Hippocampus during Amphetamine Withdrawal. Eur. J. Neurosci. 2014, 40, 3684–3692. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, M.; Suzuki, E.; Sato, T.; Maruta, S.; Watanabe, S.; Miyaoka, H. Amygdalic Levels of Dopamine and Serotonin Rise upon Exposure to Conditioned Fear Stress without Elevation of Glutamate. Neurosci. Lett. 2005, 379, 37–41. [Google Scholar] [CrossRef]
- Tokunaga, R.; Shimoju, R.; Takagi, N.; Shibata, H.; Kurosawa, M. Serotonin Release in the Central Nucleus of the Amygdala in Response to Noxious and Innocuous Cutaneous Stimulation in Anesthetized Rats. J. Physiol. Sci. 2016, 66, 307–314. [Google Scholar] [CrossRef]
- Neugebauer, V. CHAPTER 15. Amygdala Pain Mechanisms. Handb. Exp. Pharmacol. 2015, 227, 261–284. [Google Scholar]
- Bocchio, M.; Fucsina, G.; Oikonomidis, L.; McHugh, S.B.; Bannerman, D.M.; Sharp, T.; Capogna, M. Increased Serotonin Transporter Expression Reduces Fear and Recruitment of Parvalbumin Interneurons of the Amygdala. Neuropsychopharmacology 2015, 40, 3015–3026. [Google Scholar] [CrossRef] [Green Version]
- Gauriau, C.; Bernard, J.F. Pain Pathways and Parabrachial Circuits in the Rat. Exp. Physiol. 2002, 87, 251–258. [Google Scholar] [CrossRef]
- Bushnell, M.C.; Čeko, M.; Low, L.A. Cognitive and Emotional Control of Pain and Its Disruption in Chronic Pain. Nat. Rev. Neurosci. 2013, 14, 502–511. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.S.; Meng, F.C.; Cui, Y.; Xiong, Y.L.; Li, X.Y.; Meng, F.B.; Niu, Z.X.; Zheng, J.X.; Quan, Y.Q.; Wu, S.X.; et al. Regular Aerobic Exercise Attenuates Pain and Anxiety in Mice by Restoring Serotonin-Modulated Synaptic Plasticity in the Anterior Cingulate Cortex. Med. Sci. Sports Exerc. 2022, 54, 566–581. [Google Scholar] [CrossRef]
- Li, L.; Zhang, L.Z.; He, Z.X.; Ma, H.; Zhang, Y.T.; Xun, Y.F.; Yuan, W.; Hou, W.J.; Li, Y.T.; Lv, Z.J.; et al. Dorsal Raphe Nucleus to Anterior Cingulate Cortex 5-Htergic Neural Circuit Modulates Consolation and Sociability. Elife 2021, 10, e67638. [Google Scholar] [CrossRef]
- Hasegawa, E.; Maejima, T.; Yoshida, T.; Masseck, O.A.; Herlitze, S.; Yoshioka, M.; Sakurai, T.; Mieda, M. Serotonin Neurons in the Dorsal Raphe Mediate the Anticataplectic Action of Orexin Neurons by Reducing Amygdala Activity. Proc. Natl. Acad. Sci. USA 2017, 114, E3526–E3535. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.D.; Zhu, Y.; Sun, Q.X.; Deng, F.; Wan, J.; Zheng, D.; Gong, W.; Xie, S.Z.; Shen, C.J.; Fu, J.Y.; et al. Distinct Serotonergic Pathways to the Amygdala Underlie Separate Behavioral Features of Anxiety. Nat. Neurosci. 2022, 25, 1651–1663. [Google Scholar] [CrossRef]
- Hall, W.; Hayward, L.; Chapman, C.R. On “The Lateralization of Pain”. Pain 1981, 10, 337–351. [Google Scholar] [CrossRef]
- Spernal, J.; Krieg, J.C.; Lautenbacher, S. Pain Thresholds as a Putative Functional Test for Cerebral Laterality in Major Depressive Disorder and Panic Disorder. Neuropsychobiology 2003, 48, 146–151. [Google Scholar] [CrossRef]
- Symonds, L.L.; Gordon, N.S.; Bixby, J.C.; Mande, M.M. Right-Lateralized Pain Processing in the Human Cortex: An FMRI Study. J. Neurophysiol. 2006, 95, 3823–3830. [Google Scholar] [CrossRef] [Green Version]
- Vrtička, P.; Sander, D.; Vuilleumier, P. Lateralized Interactive Social Content and Valence Processing within the Human Amygdala. Front. Hum. Neurosci. 2013, 6, 2–31. [Google Scholar] [CrossRef] [Green Version]
- Ross, E.D. Differential Hemispheric Lateralization of Emotions and Related Display Behaviors: Emotion-Type Hypothesis. Brain Sci. 2021, 11, 1034. [Google Scholar] [CrossRef]
- Sorkin, L.S.; Eddinger, K.A.; Woller, S.A.; Yaksh, T.L. Origins of Antidromic Activity in Sensory Afferent Fibers and Neurogenic Inflammation. Semin. Immunopathol. 2018, 40, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Harding, E.K.; Fung, S.W.; Bonin, R.P. Insights Into Spinal Dorsal Horn Circuit Function and Dysfunction Using Optical Approaches. Front. Neural Circuits 2020, 14, 31. [Google Scholar] [CrossRef] [PubMed]
- Spike, R.C.; Puskár, Z.; Andrew, D.; Todd, A.J. A Quantitative and Morphological Study of Projection Neurons in Lamina I of the Rat Lumbar Spinal Cord. Eur. J. Neurosci. 2003, 18, 2433–2448. [Google Scholar] [CrossRef]
- Vulliemoz, S.; Raineteau, O.; Jabaudon, D. Reaching beyond the Midline: Why Are Human Brains Cross Wired? Lancet Neurol. 2005, 4, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Innocenti, G.M. Dynamic Interactions between the Cerebral Hemispheres. Exp. Brain Res. 2009, 192, 417–423. [Google Scholar] [CrossRef]
- Zhao, R.; Zhou, H.; Huang, L.; Xie, Z.; Wang, J.; Gan, W.B.; Yang, G. Neuropathic Pain Causes Pyramidal Neuronal Hyperactivity in the Anterior Cingulate Cortex. Front. Cell. Neurosci. 2018, 12, 107. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.W.; Zhang, Q.; Xia, S.H.; Zhao, W.N.; Li, Q.Z.; Yang, J.X.; An, S.; Ding, H.L.; Zhang, H.; Cao, J.L. Contralateral Projection of Anterior Cingulate Cortex Contributes to Mirror-Image Pain. J. Neurosci. 2021, 41, 9988–100083. [Google Scholar] [CrossRef]
- Kawai, S.; Takagi, Y.; Kaneko, S.; Kurosawa, T. Effect of Three Types of Mixed Anesthetic Agents Alternate to Ketamine in Mice. Exp. Anim. 2011, 60, 481–487. [Google Scholar] [CrossRef] [Green Version]
- Mogil, J.S. Animal Models of Pain: Progress and Challenges. Nat. Rev. Neurosci. 2009, 10, 283–294. [Google Scholar] [CrossRef]
- van der Walt, S.; Schönberger, J.L.; Nunez-Iglesias, J.; Boulogne, F.; Warner, J.D.; Yager, N.; Gouillart, E.; Yu, T. Scikit-Image: Image Processing in Python. PeerJ 2014, 2014, e453. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akbar, L.; Castillo, V.C.G.; Olorocisimo, J.P.; Ohta, Y.; Kawahara, M.; Takehara, H.; Haruta, M.; Tashiro, H.; Sasagawa, K.; Ohsawa, M.; et al. Multi-Region Microdialysis Imaging Platform Revealed Dorsal Raphe Nucleus Calcium Signaling and Serotonin Dynamics during Nociceptive Pain. Int. J. Mol. Sci. 2023, 24, 6654. https://doi.org/10.3390/ijms24076654
Akbar L, Castillo VCG, Olorocisimo JP, Ohta Y, Kawahara M, Takehara H, Haruta M, Tashiro H, Sasagawa K, Ohsawa M, et al. Multi-Region Microdialysis Imaging Platform Revealed Dorsal Raphe Nucleus Calcium Signaling and Serotonin Dynamics during Nociceptive Pain. International Journal of Molecular Sciences. 2023; 24(7):6654. https://doi.org/10.3390/ijms24076654
Chicago/Turabian StyleAkbar, Latiful, Virgil Christian Garcia Castillo, Joshua Philippe Olorocisimo, Yasumi Ohta, Mamiko Kawahara, Hironari Takehara, Makito Haruta, Hiroyuki Tashiro, Kiyotaka Sasagawa, Masahiro Ohsawa, and et al. 2023. "Multi-Region Microdialysis Imaging Platform Revealed Dorsal Raphe Nucleus Calcium Signaling and Serotonin Dynamics during Nociceptive Pain" International Journal of Molecular Sciences 24, no. 7: 6654. https://doi.org/10.3390/ijms24076654