
Cell-Free Supernatant (CFS) from Bacillus subtilis EB2004S and Lactobacillus helveticus EL2006H Cultured at a Range of pH Levels Modulates Potato Plant Growth under Greenhouse Conditions


Abstract
:1. Introduction
2. Results
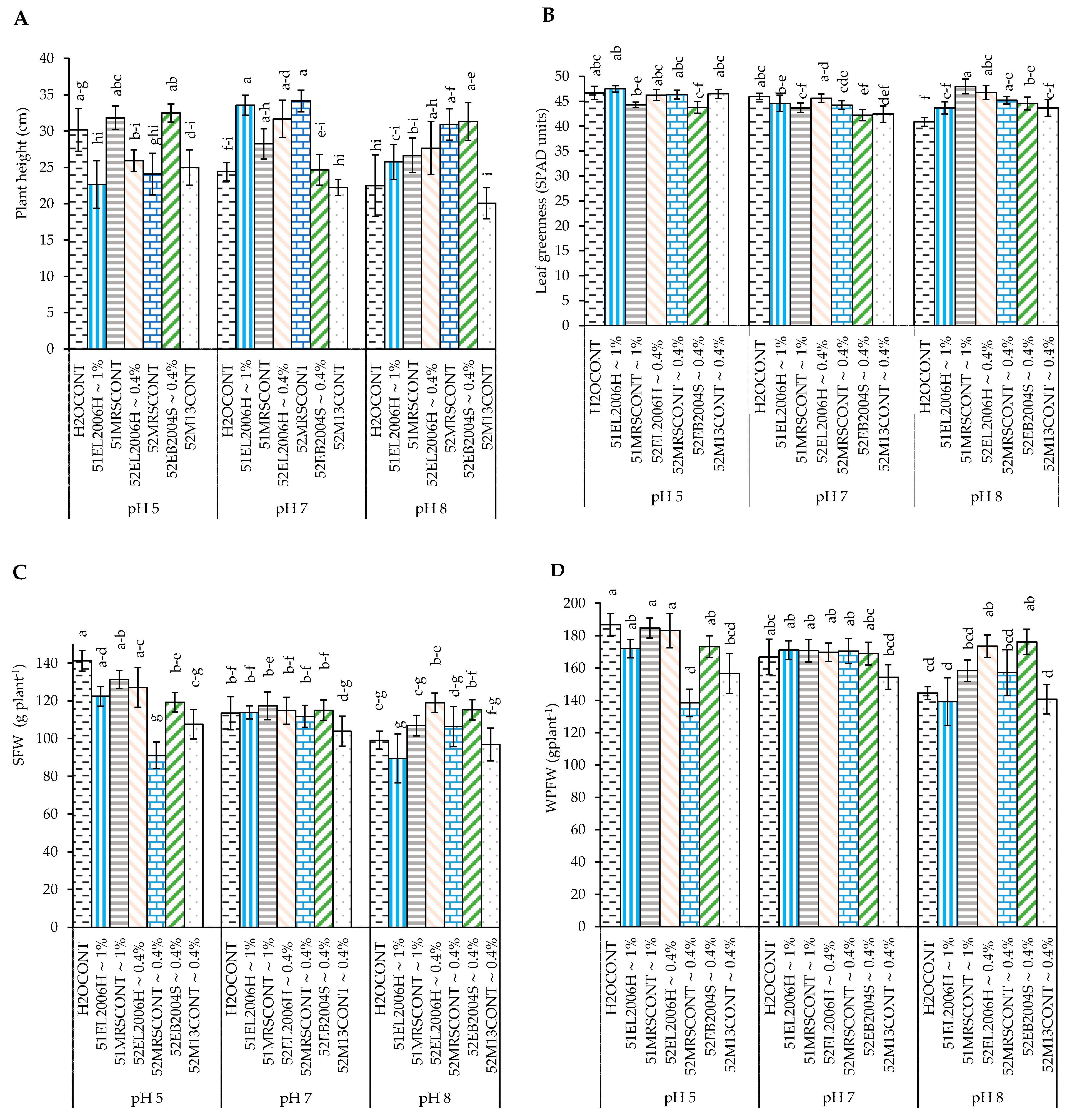
2.1. The Effect of pH and CFS Treatments on Potato Growth and Physiological Variables
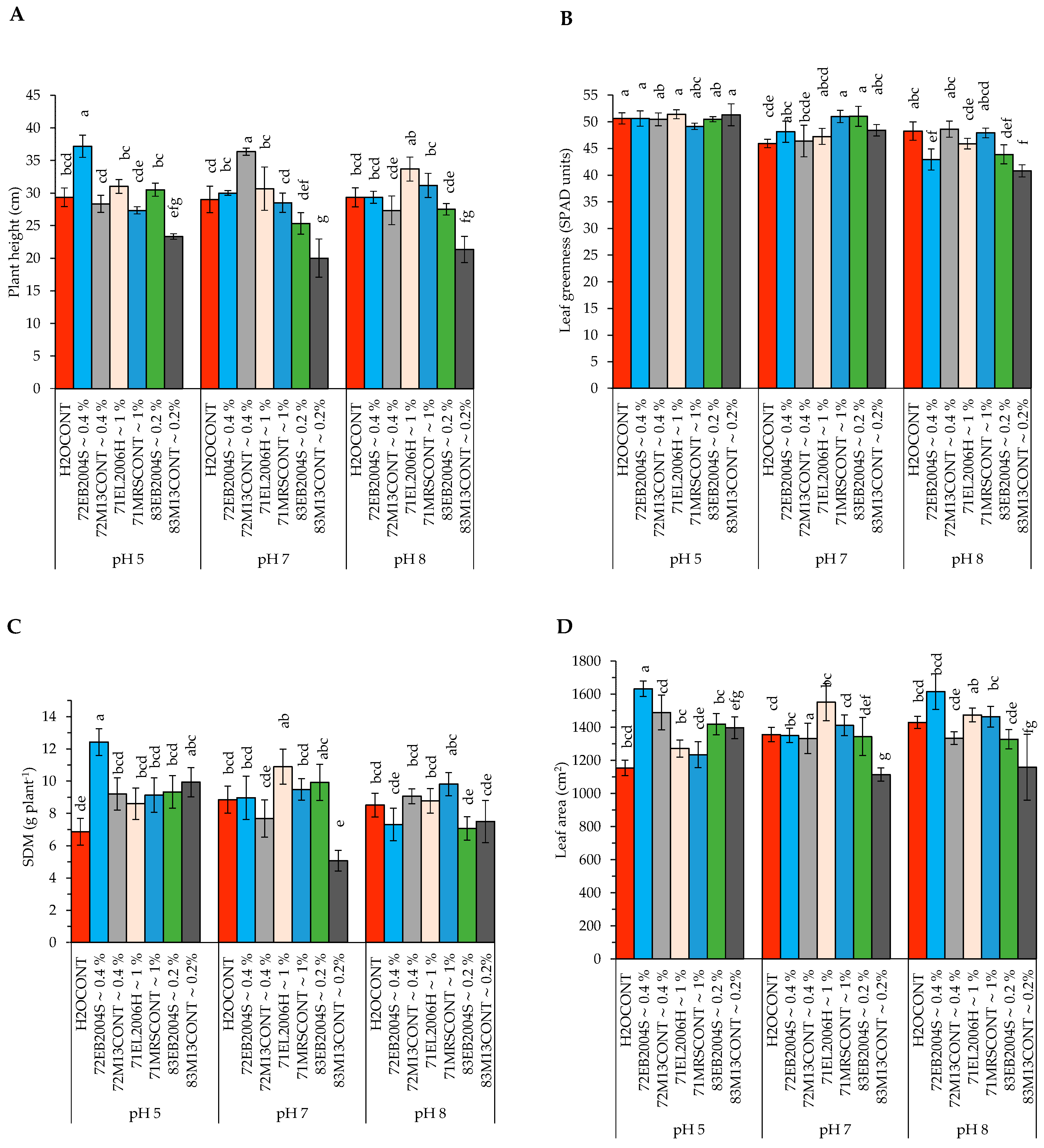
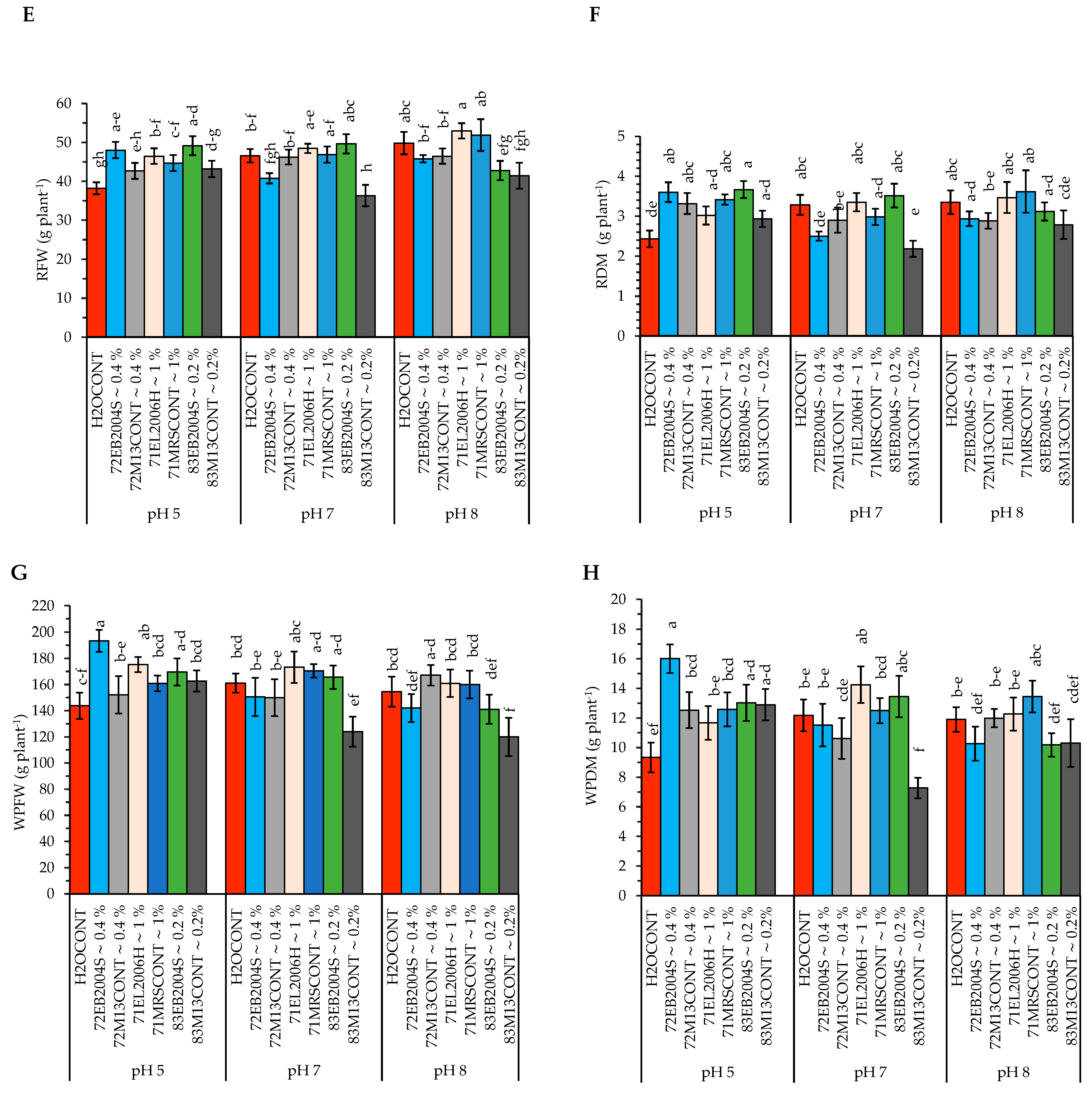
2.2. The Interactive Effect of pH and CFS Treatments on Potato Growth and Physiological Variables
3. Discussion
3.1. Effect of CFS Treatment and pH on Leaf Greenness (LG), Leaf Area (LA), Photosynthetic Rate, and Stomatal Conductance
3.2. Effect of CFS Treatment and pH on Potato Plants’ Stem Diameter and Height
3.3. Effect of CFS Treatment and pH on Potato Shoot Fresh Weight (SFW), Shoot Dry Matter (SDM), Root Fresh Weight (RFW), and Root Dry Matter (RDM)
3.4. Effect of CFS Treatment and pH on Potato Plant WPFW and WPDM
4. Materials and Methods
4.1. Preparation of CFS Treatments
4.2. Experimental Design and Layout
4.3. Treatment Application and Experiment Management
4.4. Data Collection
4.5. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Crespo-Mendes, N.; Laurent, A.; Bruun, H.H.; Hauschild, M.Z. Relationships between plant species richness and soil pH at the level of biome and ecoregion in Brazil. Ecol. Indic. 2019, 98, 266–275. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.-Y.; Wu, W.; Liu, H. Factors affecting variations of soil pH in different horizons in hilly regions. PLoS ONE 2019, 14, e0218563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Yi, D.; Ye, Y.; Guo, X.; Liu, S. Response of spatiotemporal variability in soil pH and associated influencing factors to land use change in a red soil hilly region in southern China. Catena 2022, 212, 106074. [Google Scholar] [CrossRef]
- Neina, D. The role of soil pH in plant nutrition and soil remediation. Appl. Environ. Soil Sci. 2019, 2019, 5794869. [Google Scholar] [CrossRef] [Green Version]
- Olajuyigbe, F.M.; Fatokun, C.O. Biochemical characterization of an extremely stable pH-versatile laccase from Sporothrix carnis CPF-05. Int. J. Biol. Macromol. 2017, 94, 535–543. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Grinyer, J.; Reich, P.B.; Singh, B.K. Relative importance of soil properties and microbial community for soil functionality: Insights from a microbial swap experiment. Funct. Ecol. 2016, 30, 1862–1873. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Martínez, V.; Pérez-Guzmán, L.; Johnson, J.M. Simultaneous determination of β-glucosidase, β-glucosaminidase, acid phosphomonoesterase, and arylsulfatase activities in a soil sample for a biogeochemical cycling index. Appl. Soil Ecol. 2019, 142, 72–80. [Google Scholar] [CrossRef]
- Bradl, H.B. Adsorption of heavy metal ions on soils and soils constituents. J. Colloid Interface Sci. 2004, 277, 1–18. [Google Scholar] [CrossRef]
- Kumaragamage, D.; Warren, J.; Spiers, G. Soil Chemistry. Digging Into Can. Soils 2021. Available online: https://openpress.usask.ca/soilscience/chapter/soil-chemistry/ (accessed on 29 March 2023).
- Meng, C.; Tian, D.; Zeng, H.; Li, Z.; Yi, C.; Niu, S. Global soil acidification impacts on belowground processes. Environ. Res. Lett. 2019, 14, 074003. [Google Scholar] [CrossRef]
- Bui, E.N. Causes of soil salinization, sodification, and alkalinization. In Oxford Research Encyclopedia of Environmental Science; Oxford University Press: New York, NY, USA, 2017. [Google Scholar]
- Msimbira, L.A.; Smith, D.L. The roles of plant growth promoting microbes in enhancing plant tolerance to acidity and alkalinity stresses. Front. Sustain. Food Syst. 2020, 4, 106. [Google Scholar] [CrossRef]
- Goulding, K. Soil acidification and the importance of liming agricultural soils with particular reference to the United Kingdom. Soil Use Manag. 2016, 32, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.; Nazari, M.; Antar, M.; Msimbira, L.A.; Naamala, J.; Lyu, D.; Rabileh, M.; Zajonc, J.; Smith, D.L. PGPR in agriculture: A sustainable approach to increasing climate change resilience. Front. Sustain. Food Syst. 2021, 5, 211. [Google Scholar] [CrossRef]
- Stark, J.C.; Thornton, M.; Nolte, P. Potato Production Systems; Springer Nature: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Reyniers, S.; Ooms, N.; Gomand, S.V.; Delcour, J.A. What makes starch from potato (Solanum tuberosum L.) tubers unique: A review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2588–2612. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Dulanto, P.N.; Thiry, A.A.; Glorio-Paulet, P.; Vögler, O.; Carvalho, F.P. Increasing the impact of science and technology to provide more people with healthier and safer food. Food Energy Secur. 2021, 10, e259. [Google Scholar] [CrossRef]
- Pellerin, A.; Parent, L.-É.; Fortin, J.; Tremblay, C.; Khiari, L.; Giroux, M. Environmental Mehlich-III soil phosphorus saturation indices for Quebec acid to near neutral mineral soils varying in texture and genesis. Can. J. Soil Sci. 2006, 86, 711–723. [Google Scholar] [CrossRef]
- Parent, S.É.; Dossou-Yovo, W.; Ziadi, N.; Leblanc, M.; Tremblay, G.; Pellerin, A.; Parent, L.E. Corn response to banded phosphorus fertilizers with or without manure application in Eastern Canada. Agron. J. 2020, 112, 2176–2187. [Google Scholar] [CrossRef]
- Mikkelsen, R. Phosphorus management for potatoes. Better Crops Plant Food 2015, 99, 10–11. [Google Scholar]
- Hedley, M.; McLaughlin, M. Reactions of phosphate fertilizers and by-products in soils. Phosphorus Agric. Environ. 2005, 46, 181–252. [Google Scholar]
- Naqqash, T.; Hameed, S.; Imran, A.; Hanif, M.K.; Majeed, A.; van Elsas, J.D. Differential response of potato toward inoculation with taxonomically diverse plant growth promoting rhizobacteria. Front. Plant Sci. 2016, 7, 144. [Google Scholar] [CrossRef] [Green Version]
- Morcillo, R.J.; Manzanera, M. The effects of plant-associated bacterial exopolysaccharides on plant abiotic stress tolerance. Metabolites 2021, 11, 337. [Google Scholar] [CrossRef] [PubMed]
- Naamala, J.; Smith, D.L. Microbial derived compounds, a step toward enhancing microbial inoculants technology for sustainable agriculture. Front. Microbiol. 2021, 12, 634807. [Google Scholar] [CrossRef] [PubMed]
- Jasim, A.; Sharma, L.K.; Zaeen, A.; Bali, S.K.; Buzza, A.; Alyokhin, A. Potato phosphorus response in soils with high value of phosphorus. Agriculture 2020, 10, 264. [Google Scholar] [CrossRef]
- Bai, S.H.; Omidvar, N.; Gallart, M.; Kämper, W.; Tahmasbian, I.; Farrar, M.B.; Singh, K.; Zhou, G.; Muqadass, B.; Xu, C.-Y. Combined effects of biochar and fertilizer applications on yield: A review and meta-analysis. Sci. Total Environ. 2022, 808, 152073. [Google Scholar] [CrossRef] [PubMed]
- Francis, I.; Holsters, M.; Vereecke, D. The Gram-positive side of plant–microbe interactions. Environ. Microbiol. 2010, 12, 1–12. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant growth-promoting rhizobacteria: Context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef] [Green Version]
- Antar, M.; Gopal, P.; Msimbira, L.A.; Naamala, J.; Nazari, M.; Overbeek, W.; Backer, R.; Smith, D.L. Inter-organismal signaling in the rhizosphere. In Rhizosphere Biology: Interactions between Microbes and Plants; Springer: Singapore, 2021; pp. 255–293. [Google Scholar]
- Emmanuel, O.C.; Babalola, O.O. Productivity and quality of horticultural crops through co-inoculation of arbuscular mycorrhizal fungi and plant growth promoting bacteria. Microbiol. Res. 2020, 239, 126569. [Google Scholar] [CrossRef]
- De Bruijn, F.J.; Hungria, M. Biological nitrogen fixation. In Good Microbes in Medicine, Food Production, Biotechnology, Bioremediation, and Agriculture; John Wiley & Sons: Hoboken, NJ, USA, 2022; pp. 466–475. [Google Scholar]
- Khan, N.; Bano, A. Exopolysaccharide producing rhizobacteria and their impact on growth and drought tolerance of wheat grown under rainfed conditions. PLoS ONE 2019, 14, e0222302. [Google Scholar] [CrossRef] [Green Version]
- Kour, D.; Yadav, A.N. Bacterial mitigation of drought stress in plants: Current perspectives and future challenges. Curr. Microbiol. 2022, 79, 248. [Google Scholar] [CrossRef]
- Subramanian, S.; Ricci, E.; Souleimanov, A.; Smith, D.L. A proteomic approach to lipo-chitooligosaccharide and thuricin 17 effects on soybean germinationunstressed and salt stress. PLoS ONE 2016, 11, e0160660. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.L.; Waqas, M.; Asaf, S.; Kamran, M.; Shahzad, R.; Bilal, S.; Khan, M.A.; Kang, S.-M.; Kim, Y.-H.; Yun, B.-W. Plant growth-promoting endophyte Sphingomonas sp. LK11 alleviates salinity stress in Solanum pimpinellifolium. Environ. Exp. Bot. 2017, 133, 58–69. [Google Scholar] [CrossRef]
- Naamala, J.; Msimbira, L.A.; Antar, M.; Subramanian, S.; Smith, D.L. Cell-Free Supernatant Obtained From a Salt Tolerant Bacillus amyloliquefaciens Strain Enhances Germination and Radicle Length Under NaCl Stressed and Optimal Conditions. Front. Sustain. Food Syst. 2022, 6, 788939. [Google Scholar] [CrossRef]
- Gondal, A.H.; Farooq, Q.; Sohail, S.; Kumar, S.S.; Toor, M.D.; Zafar, A.; Rehman, B. Adaptability of soil pH through innovative microbial approach. Curr. Res. Agric. Sci. 2021, 8, 71–79. [Google Scholar]
- Msimbira, L.A.; Naamala, J.; Antar, M.; Subramanian, S.; Smith, D.L. Effect of Microbial Cell-Free Supernatants Extracted From a Range of pH Levels on Corn (Zea mays L.) and Tomato (Solanum lycopersicum L.) Seed Germination and Seedling Growth. Front. Sustain. Food Syst. 2022, 6, 9. [Google Scholar] [CrossRef]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.; Reddy, M.; El Enshasy, H. Plant growth promoting rhizobacteria (PGPR) as green bioinoculants: Recent developments, constraints, and prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Arnon, D.; Hoagland, D. Crop production in artificial culture solutions and in soils with special reference to factors influencing yields and absorption of inorganic nutrients. Soil Sci. 1940, 50, 463–485. [Google Scholar]
- Pathak, D.; Lone, R.; Koul, K. Arbuscular Mycorrhizal fungi (AMF) and plant growth-promoting Rhizobacteria (PGPR) Association in Potato (Solanum tuberosum L.): A brief Review. In Probiotics Plant Health; Springer: Singapore, 2017; pp. 401–420. [Google Scholar]
- Mickelbart, M.V.; Gosney, M.J.; Camberato, J.; Stanton, K.M. Soil pH effects on growth and foliar nutrient concentrations of Spiraea alba Du Roi and Spiraea tomentosa L. HortScience 2012, 47, 902–906. [Google Scholar] [CrossRef] [Green Version]
- Stanton, K.M.; Mickelbart, M.V. Growth and foliar nutrition of Spiraea alba Du Roi and Spiraea tomentosa L. in response to root zone pH. Sci. Hortic. 2014, 165, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.Y.; Oh, S.; Oh, M.M.; Son, J.E. Estimation of individual leaf area, fresh weight, and dry weight of hydroponically grown cucumbers (Cucumis sativus L.) using leaf length, width, and SPAD value. Sci. Hortic. 2007, 111, 330–334. [Google Scholar] [CrossRef]
- Ramírez, D.; Yactayo, W.; Gutiérrez, R.; Mares, V.; De Mendiburu, F.; Posadas, A.; Quiroz, R. Chlorophyll concentration in leaves is an indicator of potato tuber yield in water-shortage conditions. Sci. Hortic. 2014, 168, 202–209. [Google Scholar] [CrossRef]
- Zarzecka, K.; Gugała, M.; Mystkowska, I.; Sikorska, A. The Leaf Greenness Index SPAD and Selected Features of Potato Following an Application of Herbicides and Biostimulants. J. Ecol. Eng. 2021, 22, 54–63. [Google Scholar] [CrossRef]
- Lee, S.; Behringer, G.; Hung, R.; Bennett, J. Effects of fungal volatile organic compounds on Arabidopsis thaliana growth and gene expression. Fungal Ecol. 2019, 37, 1–9. [Google Scholar] [CrossRef]
- Khan, A.R.; Ullah, I.; Waqas, M.; Shahzad, R.; Hong, S.-J.; Park, G.-S.; Jung, B.K.; Lee, I.-J.; Shin, J.-H. Plant growth-promoting potential of endophytic fungi isolated from Solanum nigrum leaves. World J. Microbiol. Biotechnol. 2015, 31, 1461–1466. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Vaziriyeganeh, M.; Zwiazek, J.J. Effects of pH and mineral nutrition on growth and physiological responses of trembling aspen (Populus tremuloides), jack pine (Pinus banksiana), and white spruce (Picea glauca) seedlings in sand culture. Plants 2020, 9, 682. [Google Scholar] [CrossRef]
- Epstein, E.; Bloom, A. Mineral Nutrition of Plants: Principles and Perspectives, 2nd ed.; Sinauer Associates Inc.: Sunderland, UK, 2005; pp. 292–305. [Google Scholar]
- Baset, M.M.; Shamsuddin, Z.; Wahab, Z.; Marziah, M. Effect of plant growth promoting rhizobacterial (PGPR) inoculation on growth and nitrogen incorporation of tissue-cultured’musa’plantlets under nitrogen-free hydroponics condition. Aust. J. Crop Sci. 2010, 4, 85–90. [Google Scholar]
- Felle, H.H.; Hanstein, S. The apoplastic pH of the substomatal cavity of Vicia faba leaves and its regulation responding to different stress factors. J. Exp. Bot. 2002, 53, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Blanco, F.F.; Folegatti, M.V. Estimation of leaf area for greenhouse cucumber by linear measurements under salinity and grafting. Sci. Agric. 2005, 62, 305–309. [Google Scholar] [CrossRef] [Green Version]
- Williams, L.E. Growth of ‘Thompson Seedless’ grapevines: I. Leaf area development and dry weight distribution. J. Am. Soc. Hortic. Sci. 1987, 112, 325–330. [Google Scholar] [CrossRef]
- Pandey, S.; Singh, H. A simple, cost-effective method for leaf area estimation. J. Bot. 2011, 2011, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Elgaml, N.M.; Salama, A.B.; Shehata, H.S.; Abdelhamid, M.T. Effective Microorganisms Improve Growth, Nutrients Uptake, Normalized Difference Vegetation Index, Photosystem II, and Essential Oil While Reducing Canopy Temperature in Water-Stressed Salvia sclarea Plants. Int. J. Agron. 2022, 2022, 1767347. [Google Scholar] [CrossRef]
- Reyes-Díaz, M.; Ulloa-Inostroza, E.M.; González-Villagra, J.; Ivanov, A.G.; Kurepin, L.V. Phytohormonal responses to soil acidity in plants. In Plant Hormones under Challenging Environmental Factors; Springer: Dordrecht, The Netherlands, 2016; pp. 133–155. [Google Scholar]
- Invers, O.; Romero, J.; Pérez, M. Effects of pH on seagrass photosynthesis: A laboratory and field assessment. Aquat. Bot. 1997, 59, 185–194. [Google Scholar] [CrossRef]
- Hwang, J.-S.; You, Y.-H.; Bae, J.-J.; Khan, S.A.; Kim, J.-G.; Choo, Y.-S. Effects of endophytic fungal secondary metabolites on the growth and physiological response of Carex kobomugi Ohwi. J. Coast. Res. 2011, 27, 544–548. [Google Scholar] [CrossRef]
- Liu, F.; Andersen, M.N.; Jensen, C.R. Capability of the ‘Ball-Berry’model for predicting stomatal conductance and water use efficiency of potato leaves under different irrigation regimes. Sci. Hortic. 2009, 122, 346–354. [Google Scholar] [CrossRef]
- Ishartati, E.; Sufianto, S.; Zakia, A.; Mejaya, M.J.; Madianto, R. Determination of Agronomic Characteristics as Selection Criteria in Potato Crossing Lines. Planta Trop. J. Agro Sci. 2022, 10, 34–44. [Google Scholar] [CrossRef]
- Sadik, E.; Hussien, M.; Tewodros, A. Effects of seed tuber size on growth and yield performance of potato (Solanum tuberosum L.) varieties under field conditions. Afr. J. Agric. Res. 2018, 13, 2077–2086. [Google Scholar] [CrossRef] [Green Version]
- Buensateai, N.; Sompong, M.; Thamnu, K.; Athinuwat, D.; Brauman, A.; Plassard, C. The plant growth promoting bacterium Bacillus sp. CaSUT007 produces phytohormone and extracellular proteins for enhanced growth of cassava. Afr. J. Microbiol. Res. 2013, 7, 4949–4954. [Google Scholar]
- Posada, L.F.; Ramírez, M.; Ochoa-Gómez, N.; Cuellar-Gaviria, T.Z.; Argel-Roldan, L.E.; Ramírez, C.A.; Villegas-Escobar, V. Bioprospecting of aerobic endospore-forming bacteria with biotechnological potential for growth promotion of banana plants. Sci. Hortic. 2016, 212, 81–90. [Google Scholar] [CrossRef]
- Msimbira, L.A.; Subramanian, S.; Naamala, J.; Antar, M.; Smith, D.L. Secretome Analysis of the Plant Biostimulant Bacteria Strains Bacillus subtilis (EB2004S) and Lactobacillus helveticus (EL2006H) in Response to pH Changes. Int. J. Mol. Sci. 2022, 23, 15144. [Google Scholar] [CrossRef]
- Bell, I.M.; Fisher, M.L.; Wu, Z.P.; Hilvert, D. Kinetic studies on the peroxidase activity of selenosubtilisin. Biochemistry 1993, 32, 3754–3762. [Google Scholar] [CrossRef]
- Baroja-Fernández, E.; Almagro, G.; Sánchez-López, Á.M.; Bahaji, A.; Gámez-Arcas, S.; De Diego, N.; Dolezal, K.; Muñoz, F.J.; Climent Sanz, E.; Pozueta-Romero, J. Enhanced yield of pepper plants promoted by soil application of volatiles from cell-free fungal culture filtrates is associated with activation of the beneficial soil microbiota. Front. Plant Sci. 2021, 12, 752653. [Google Scholar] [CrossRef]
- Cao, W.; Tibbitts, T.W. Responses of potatoes to solution pH levels with different forms of nitrogen. J. Plant Nutr. 1994, 17, 109–126. [Google Scholar] [CrossRef]
- Nookaraju, A.; Kappachery, S.; Yu, J.W.; Park, S.W. Rhizobacteria influence potato tuberization through enhancing lipoxygenase activity. Am. J. Potato Res. 2011, 88, 441–449. [Google Scholar] [CrossRef]
- Khan, M.; Gemenet, D.C.; Villordon, A. Root system architecture and abiotic stress tolerance: Current knowledge in root and tuber crops. Front. Plant Sci. 2016, 7, 1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kratzke, M.G.; Palta, J.P. Evidence for the existence of functional roots on potato tubers and stolons: Significance in water transport to the tuber. Am. Potato J. 1985, 62, 227–236. [Google Scholar] [CrossRef]
- Cao, W.; Tibbitts, T.W. Phasic temperature change patterns affect growth and tuberization in potatoes. J. Am. Soc. Hortic. Sci. 1994, 119, 775–778. [Google Scholar] [CrossRef] [Green Version]
- El-Shanshoury, A. Growth promotion of wheat seedlings by Streptomyces atroolivaceus. J. Agron. Crop Sci. 1989, 163, 109–114. [Google Scholar] [CrossRef]
- Buensanteai, N.; Yuen, G.; Prathuangwong, S. The biocontrol bacterium Bacillus amyloliquefaciens KPS46 produces auxin, surfactin and extracellular proteins for enhanced growth of soybean plant. Thai J. Agric. Sci. 2008, 41, 101–116. [Google Scholar]
- Ofori, P.A.; Mizuno, A.; Suzuki, M.; Martinoia, E.; Reuscher, S.; Aoki, K.; Shibata, D.; Otagaki, S.; Matsumoto, S.; Shiratake, K. Genome-wide analysis of ATP binding cassette (ABC) transporters in tomato. PLoS ONE 2018, 13, e0200854. [Google Scholar] [CrossRef] [Green Version]
- Hamayun, M.; Khan, S.A.; Khan, A.L.; Rehman, G.; Kim, Y.-H.; Iqbal, I.; Hussain, J.; Sohn, E.-Y.; Lee, I.-J. Gibberellin production and plant growth promotion from pure cultures of Cladosporium sp. MH-6 isolated from cucumber (Cucumis sativus L.). Mycologia 2010, 102, 989–995. [Google Scholar] [CrossRef]
- Hegazi, M.; Metwaly, M.; Belal, E. Influence of plant growth promoting bacteria (pgpb) on coriander (Coriandrum sativum L.) and dill (Anethum graveolens L.) plants. J. Plant Prod. 2015, 6, 205–218. [Google Scholar] [CrossRef]
- Gray, E.; Lee, K.; Souleimanov, A.; Di Falco, M.; Zhou, X.; Ly, A.; Charles, T.; Driscoll, B.; Smith, D. A novel bacteriocin, thuricin 17, produced by plant growth promoting rhizobacteria strain Bacillus thuringiensis NEB17: Isolation and classification. J. Appl. Microbiol. 2006, 100, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, S.; Souleimanov, A.; Smith, D.L. Proteomic studies on the effects of lipo-chitooligosaccharide and thuricin 17 under unstressed and salt stressed conditions in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kesaulya, H.; Zakaria, B.; Syaiful, S.A. Isolation and physiological characterization of PGPR from potato plant rhizosphere in medium land of Buru Island. Procedia Food Sci. 2015, 3, 190–199. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Factors | Stem Diameter | Plant Height | Chlorophyll | Photosynthesis | Stomatal Conductance | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | CFS Treatments | (mm) | (cm) | SPAD Units | (μmol m−2 sec−1) | molm−2s−1 | |||||
| 30 DAP | 45 DAP | 30 DAP | 45 DAP | 30 DAP | 45 DAP | 30 DAP | 45 DAP | 30 DAP | 45 DAP | ||
| H2OCONT | 7.15 ± 0.30 | 8.39 ± 0.368 | 25.7 ± 1.6 a | 47.9 ± 2.0 | 44.49 ± 0.8 | 43.32 ± 0.7 | 8.08 ± 0.2 b | 10.66 ± 0.61 bc | 0.06 ± 0.004 | 0.06 ± 0.003 bc | |
| 51EL2006H∼1% | 7.30 ± 0.42 | 8.37 ± 0.456 | 27.0 ± 1.5 a | 51.5 ± 1.9 | 45.23 ± 0.7 | 44.07 ± 0.76 | 8.41 ± 0.4 ab | 11.23 ± 0.63 ab | 0.06 ± 0.006 | 0.07 ± 0.006 b | |
| 51MRSCONT∼ | 7.67 ± 0.24 | 8.33 ± 0.361 | 28.9 ± 1.2 a | 51.4 ± 2.2 | 45.31 ± 0.7 | 44.45 ± 0.47 | 7.68 ± 0.3 b | 9.88 ± 0.53 bc | 0.05 ± 0.004 | 0.04 ± 0.003 c | |
| 52EL2006H∼0.4% | 7.64 ± 0.25 | 8.63 ± 0.364 | 28.4 ± 1.2 a | 49.7 ± 2.1 | 46.18 ± 0.6 | 45.93 ± 0.35 | 8.49 ± 0.4 ab | 10.78 ± 0.56 abc | 0.06 ± 0.009 | 0.06 ± 0.004 bc | |
| 52MRSCONT∼Positive control | 7.39 ± 0.30 | 8.34 ± 0.365 | 29.7 ± 1.5 a | 52.1 ± 2.6 | 45.25 ± 0.5 | 43.76 ± 0.62 | 7.97 ± 0.3 b | 9.42 ± 0.33 c | 0.06 ± 0.007 | 0.05 ± 0.003 c | |
| 52EB2004S∼0.4% | 7.45 ± 0.29 | 8.55 ± 0.188 | 29.5 ± 1.4 a | 50.6 ± 1.9 | 43.51 ± 0.6 | 44.42 ± 0.66 | 9.27 ± 0.4 a | 12.33 ± 0.7 a | 0.06 ± 0.006 | 0.1 ± 0.011 a | |
| 52M13CONT∼Positive control | 8.02 ± 0.36 | 8.60 ± 0.344 | 22.4 ± 1.1 b | 45.2 ± 2.4 | 44.18 ± 0.8 | 44.35 ± 0.55 | 8.12 ± 0.2 b | 9.69 ± 0.45 bc | 0.06 ± 0.004 | 0.05 ± 0.003 bc | |
| pH | |||||||||||
| 5 | 7.81 ± 0.20 | 8.74 ± 0.158 | 28.4 ± 0.9 | 50.2 ± 1.2 | 45.89 ± 0.3 a | 44.44 ± 0.34 | 9.03 ± 0.2 a | 10.57 ± 0.37 | 0.07 ± 0.004 a | 0.07 ± 0.005 a | |
| 7 | 7.37 ± 0.18 | 8.43 ± 0.246 | 27.5 ± 0.9 | 50.1 ± 1.5 | 44.09 ± 0.5 b | 44.05 ± 0.45 | 8.08 ± 0.2 b | 10.32 ± 0.38 | 0.05 ± 0.003 b | 0.06 ± 0.003 ab | |
| 8 | 7.37 ± 0.22 | 8.20 ± 0.266 | 26.4 ± 1.0 | 49.1 ± 1.5 | 44.65 ± 0.4 b | 44.49 ± 0.41 | 7.75 ± 0.2 b | 10.83 ± 0.4 | 0.05 ± 0.003 b | 0.05 ± 0.003 b | |
| p-values | |||||||||||
| pH | 0.2868 | 0.2866 | 0.297 | 0.8354 | 0.0136 | 0.6955 | 0.0004 | 0.6297 | 0.0046 | 0.0121 | |
| CFS Treatments | 0.6784 | 0.9931 | 0.0033 | 0.33 | 0.1168 | 0.1006 | 0.0537 | 0.0066 | 0.9453 | <0.0001 | |
| Interaction | 0.3251 | 0.942 | 0.0009 | 0.7959 | 0.0047 | 0.3081 | 0.5901 | 0.6361 | 0.8756 | 0.5838 | |
| 2 | H2OCONT∼Negative control | 9.67 ± 0.22 | 9.3788 ± 0.353 | 29.22 ± 0.9 bcd | 55.7 ± 1.0 | 48.27 ± 0.82 | 46.77 ± 0.53 a | 12.3 ± 0.60 | 12.8 ± 0.66 | 0.09 ± 0.009 | 0.09 ± 0.01 |
| 72EB2004S∼0.4 % | 10.17 ± 0.23 | 10.13 ± 0.284 | 32.16 ± 1.05 a | 55.8 ± 1.6 | 47.22 ± 1.25 | 45.52 ± 0.48 abc | 12.2 ± 0.68 | 13.19 ± 0.66 | 0.09 ± 0.012 | 0.09 ± 0.009 | |
| 72M13CONT∼Positive control | 9.8 ± 0.29 | 9.8077 ± 0.317 | 30.66 ± 1.27 abc | 54.4 ± 1.5 | 48.48 ± 1.17 | 44.59 ± 0.45 c | 12.42 ± 0.59 | 13.91 ± 0.70 | 0.09 ± 0.010 | 0.09 ± 0.008 | |
| 71EL2006H∼1 % | 10.03 ± 0.30 | 9.7177 ± 0.396 | 31.77 ± 1.27 ab | 56.9 ± 1.9 | 48.18 ± 0.84 | 46.63 ± 0.38 a | 12.94 ± 0.68 | 13.73 ± 0.61 | 0.08 ± 0.008 | 0.09 ± 0.016 | |
| 71MRSCONT∼Positive control | 9.82 ± 0.20 | 9.5727 ± 0.303 | 29 ± 0.85 cd | 56.0 ± 1.4 | 49.35 ± 0.58 | 46.22 ± 0.74 ab | 11.64 ± 0.63 | 12.85 ± 0.60 | 0.08 ± 0.008 | 0.09 ± 0.01 | |
| 83EB2004S∼0.2 % | 10.18 ± 0.24 | 10.052 ± 0.314 | 27.77 ± 0.83 d | 55.2 ± 1.1 | 48.46 ± 1.14 | 46.36 ± 0.42 ab | 11.99 ± 0.68 | 14.54 ± 0.62 | 0.09 ± 0.012 | 0.12 ± 0.01 | |
| 83M13CONT∼Positive control | 9.21 ± 0.17 | 9.26 ± 0.216 | 21.55 ± 1.16 e | 50.7 ± 1.6 | 46.83 ± 1.34 | 44.93 ± 0.61 bc | 12.08 ± 0.54 | 14.03 ± 0.70 | 0.09 ± 0.01 | 0.11 ± 0.012 | |
| pH | |||||||||||
| 5 | 9.53 ± 0.12 b | 10.2 ± 0.198 a | 29.57 ± 0.72 | 54.7 ± 1.0 | 50.57 ± 0.43 a | 46.94 ± 0.33 a | 12.45 ± 0.39 | 12.87 ± 0.47 | 0.11 ± 0.008 a | 0.08 ± 0.006 b | |
| 7 | 9.79 ± 0.13 ab | 9.42 ± 0.184 b | 28.54 ± 1.01 | 55.2 ± 0.9 | 48.3 ± 0.68 b | 45.52 ± 0.33 b | 12.42 ± 0.33 | 13.57 ± 0.41 | 0.08 ± 0.005 b | 0.09 ± 0.006 b | |
| 8 | 10.2 ± 0.2 a | 9.48 ± 0.221 b | 28.52 ± 0.8 | 55.0 ± 1.1 | 45.47 ± 0.67 c | 45.12 ± 0.35 b | 11.8 ± 0.49 | 14.3 ± 0.36 | 0.08 ± 0.007 b | 0.11 ± 0.008 a | |
| p-values | |||||||||||
| pH | 0.0095 | 0.0166 | 0.4053 | 0.9415 | <0.0001 | 0.0006 | 0.4956 | 0.0555 | 0.0162 | 0.0025 | |
| CFS Treatments | 0.063 | 0.4053 | <0.0001 | 0.1222 | 0.4699 | 0.0162 | 0.9004 | 0.394 | 0.6465 | 0.3426 | |
| Interaction | 0.4793 | 0.9914 | 0.0003 | 0.598 | 0.0043 | 0.8664 | 0.9991 | 0.2057 | 0.9451 | 0.079 | |
| Experiment | Factors | Leaf Area | SFW | SDM | RFW | RDM | WPFW | WPDM |
|---|---|---|---|---|---|---|---|---|
| CFS Treatments | (cm2) | (g Plant−1) | (g Plant−1) | (g Plant−1) | (g Plant−1) | (g Plant−1) | (g Plant−1) | |
| 1 | H2OCONT∼Negative control | 1548.23 ± 114.11 | 117.89 ± 5.53 a | 12.15 ± 0.51 | 48.17 ± 1.83 b | 7.77 ± 1.08 | 166.07 ± 5.98 abc | 19.93 ± 1.12 |
| 51EL2006H∼1% | 1657.10 ± 76.03 | 108.63 ± 5.63 ab | 12.53 ± 0.96 | 52.1 ± 1.93 ab | 7.89 ± 0.93 | 160.74 ± 6.44 bcd | 20.42 ± 0.66 | |
| 51MRSCONT∼Positive control | 1539.28 ± 98.13 | 118.55 ± 4.01 a | 13.00 ± 0.57 | 52.76 ± 1.28 ab | 8.55 ± 1.31 | 171.31 ± 4.44 ab | 21.56 ± 1.09 | |
| 52EL2006H∼0.4% | 1614.25 ± 86.47 | 120.32 ± 4.45 a | 13.55 ± 0.71 | 55.12 ± 1.43 a | 8.8 ± 1.27 | 175.45 ± 4.57 a | 22.36 ± 1.26 | |
| 52MRSCONT∼Positive control | 1456.31 ± 110.13 | 103.13 ± 4.88 b | 11.69 ± 0.87 | 52.27 ± 2.18 ab | 8.3 ± 1.21 | 155.40 ± 6.57 cd | 19.99 ± 1.00 | |
| 52EB2004S∼0.4% | 1612.54 ± 78.82 | 116.52 ± 2.91 a | 12.58 ± 0.55 | 56.25 ± 2.29 a | 8.97 ± 1.25 | 172.78 ± 4.00 ab | 21.56 ± 1.17 | |
| 52M13CONT∼Positive control | 1380.04 ± 114.75 | 102.82 ± 4.55 b | 12.12 ± 0.77 | 47.76 ± 2.22 b | 7.8 ± 1.04 | 150.58 ± 5.62 d | 19.92 ± 0.74 | |
| pH | ||||||||
| 5 | 1642.81 ± 60.26 | 120.03 ± 3.35 a | 12.86 ± 0.47 | 50.67 ± 1.35 b | 9.09 ± 0.8 | 170.70 ± 3.89 a | 21.95 ± 0.72 a | |
| 7 | 1509.37 ± 72.86 | 112.91 ± 2.43 a | 13.00 ± 0.45 | 54.57 ± 1.04 a | 8.38 ± 0.74 | 167.48 ± 2.79 a | 21.39 ± 0.64 a | |
| 8 | 1479.70 ± 57.16 | 104.72 ± 3.22 b | 11.70 ± 0.47 | 50.95 ± 1.43 b | 7.42 ± 0.68 | 155.67 ± 4.07 b | 19.12 ± 0.58 b | |
| p-values | ||||||||
| pH | 0.1844 | 0.0007 | 0.1033 | 0.0469 | 0.3325 | 0.0041 | 0.0069 | |
| CFS Treatment | 0.4787 | 0.0062 | 0.6183 | 0.0117 | 0.9855 | 0.0034 | 0.4452 | |
| Interaction | 0.9912 | 0.0203 | 0.4197 | 0.3315 | 0.9972 | 0.0265 | 0.5673 | |
| 2 | H2OCONT∼Negative control | 1312.98 ± 36.47 bc | 108.28 ± 4.86 | 8.07 ± 0.48 | 44.85 ± 1.65 b | 3.02 ± 0.17 ab | 153.15 ± 5.57 a | 11.13 ± 0.61 ab |
| 72EB2004S∼0.4 % | 1532.2 ± 49.84 a | 117.1 ± 7.66 | 9.56 ± 0.77 | 44.84 ± 1.1 b | 3.01 ± 0.15 ab | 161.96 ± 8.29 a | 12.60 ± 0.88 a | |
| 72M13CONT∼Positive control | 1384.71 ± 48.74 b | 111.27 ± 6.42 | 8.65 ± 0.52 | 45.1 ± 1.15 b | 3.03 ± 0.14 ab | 156.4 ± 7.02 a | 11.71 ± 0.63 ab | |
| 71EL2006H∼1 % | 1431.94 ± 49.87 ab | 120.53 ± 5.61 | 9.42 ± 0.57 | 49.29 ± 1.15 a | 3.27 ± 0.16 a | 169.83 ± 5.53 a | 12.72 ± 0.68 a | |
| 71MRSCONT∼Positive control | 1369.52 ± 44.06 b | 115.98 ± 3.92 | 9.47 ± 0.45 | 47.8 ± 1.72 ab | 3.33 ± 0.19 a | 163.81 ± 4.3 a | 12.84 ± 0.56 a | |
| 83EB2004S∼0.2 % | 1362.88 ± 46.11 b | 111.56 ± 5.50 | 8.77 ± 0.6 | 47.18 ± 1.54 ab | 3.43 ± 0.14 a | 158.76 ± 6.27 a | 12.21 ± 0.72 a | |
| 83M13CONT∼Positive control | 1222.79 ± 73.1 c | 95.17 ± 7.24 | 7.5 ± 0.72 | 40.3 ± 1.66 c | 2.63 ± 0.16 b | 135.51 ± 7.87 b | 10.15 ± 0.85 b | |
| pH | ||||||||
| 5 | 1370.59 ± 33.57 | 120.76 ± 3.54 a | 9.35 ± 0.40 | 44.62 ± 0.89 | 3.19 ± 0.09 | 165.4 ± 4.02 a | 12.57 ± 0.48 | |
| 7 | 1351.12 ± 33.30 | 111.43 ± 3.91 ab | 8.69 ± 0.44 | 44.97 ± 0.97 | 2.95 ± 0.10 | 156.42 ± 4.55 ab | 11.68 ± 0.52 | |
| 8 | 1399.87 ± 39.07 | 102.05 ± 4.14 b | 8.29 ± 0.33 | 47.29 ± 1.13 | 3.16 ± 0.12 | 149.37 ± 4.54 b | 11.47 ± 0.41 | |
| p-values | ||||||||
| pH | 0.5307 | 0.0021 | 0.1084 | 0.0614 | 0.1991 | 0.0199 | 0.1551 | |
| CFS Treatment | 0.0008 | 0.0505 | 0.0616 | 0.0002 | 0.0083 | 0.0061 | 0.0393 | |
| Interaction | 0.00258 | 0.0946 | 0.0008 | 0.0014 | 0.0203 | 0.0277 | 0.0009 |
| Experiment 1 Treatments | Experiment 2 Treatments |
|---|---|
| H2OCONT∼Negative control | H2OCONT∼Negative control |
| 51EL2006H∼1% | 72EB2004S∼0.4% |
| 51MRSCONT∼1% (Positive control) | 72M13CONT∼0.4% (Positive control) |
| 52EL2006H∼0.4% | 71EL2006H∼1% |
| 52MRSCONT∼0.4% (Positive control) | 71MRSCONT∼1% (Positive control) |
| 52EB2004S∼0.4% | 83EB2004S∼0.2% |
| 52M13CONT∼0.4% (Positive control) | 83M13CONT∼0.2% (Positive control) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Msimbira, L.A.; Naamala, J.; Subramanian, S.; Smith, D.L. Cell-Free Supernatant (CFS) from Bacillus subtilis EB2004S and Lactobacillus helveticus EL2006H Cultured at a Range of pH Levels Modulates Potato Plant Growth under Greenhouse Conditions. Int. J. Mol. Sci. 2023, 24, 6620. https://doi.org/10.3390/ijms24076620
Msimbira LA, Naamala J, Subramanian S, Smith DL. Cell-Free Supernatant (CFS) from Bacillus subtilis EB2004S and Lactobacillus helveticus EL2006H Cultured at a Range of pH Levels Modulates Potato Plant Growth under Greenhouse Conditions. International Journal of Molecular Sciences. 2023; 24(7):6620. https://doi.org/10.3390/ijms24076620
Chicago/Turabian StyleMsimbira, Levini A., Judith Naamala, Sowmyalakshmi Subramanian, and Donald L. Smith. 2023. "Cell-Free Supernatant (CFS) from Bacillus subtilis EB2004S and Lactobacillus helveticus EL2006H Cultured at a Range of pH Levels Modulates Potato Plant Growth under Greenhouse Conditions" International Journal of Molecular Sciences 24, no. 7: 6620. https://doi.org/10.3390/ijms24076620