
Development of Highly Efficient Resistance to Beet Curly Top Iran Virus (Becurtovirus) in Sugar Beet (B. vulgaris) via CRISPR/Cas9 System
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. RCA Assay Indicated Lower Translocation of BCTIV from gRNA/Cas9 Treated Local Leaves to Newly Emerged Systemic Leaves
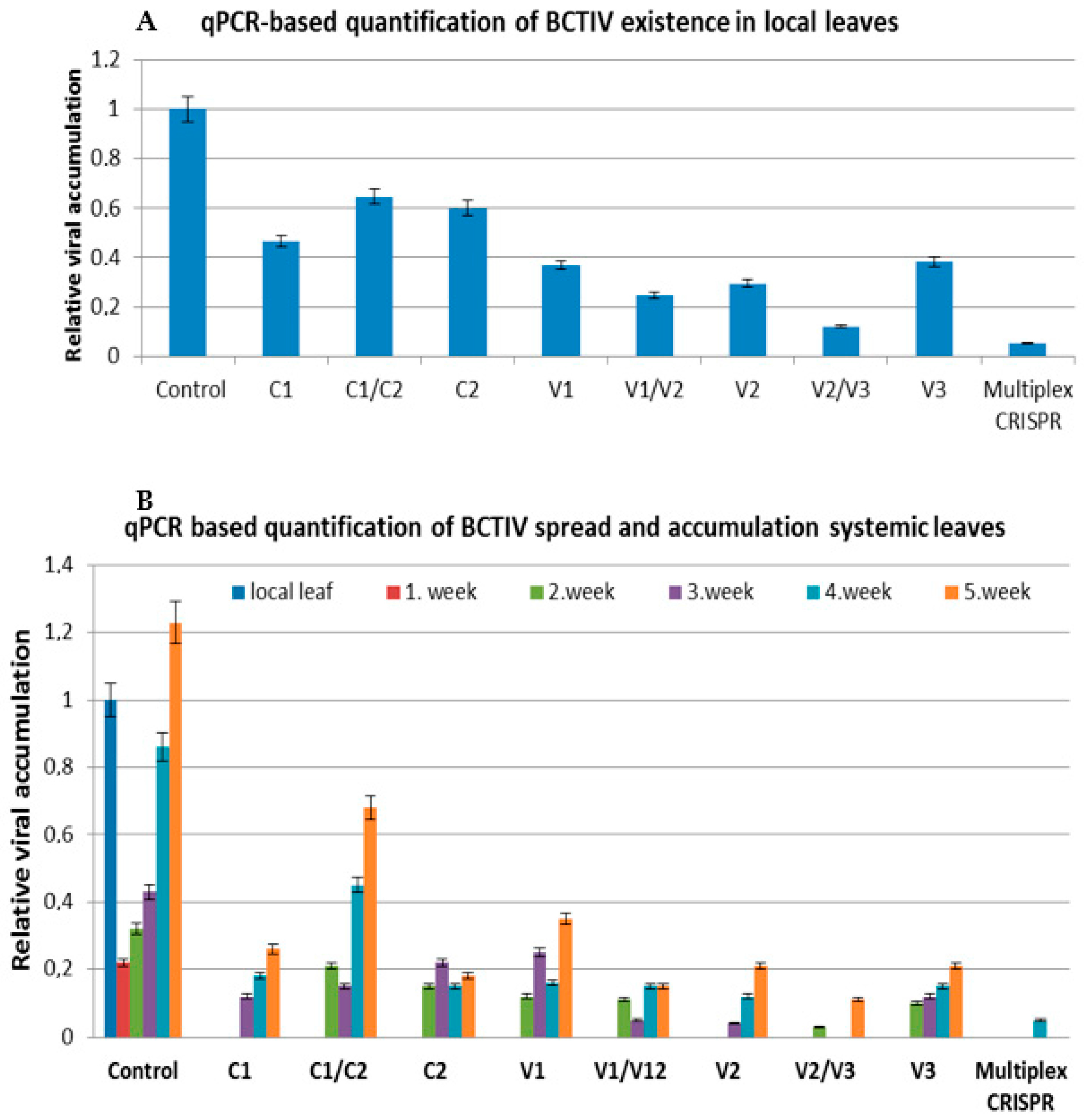
2.2. qPCR Assay Represented the Lower Accumulation of BCTIV in gRNA/Cas9-Treated Local and Systemic Leaves
2.3. PCR-RE Assay Revealed the High gRNA/Cas9 Efficiency on the Knockout of BCTIV Genome and No Mutant Escape from Local to Systemic Leaves
2.4. BCTD Symtoms and Biomass Reduction Decreased with gRNA/Cas9 Treatment in Sugar Beets
2.5. Multiplex CRISPR System Had Full Resistance to BCTIV in Sugar Beet
3. Discussion
4. Materials and Methods
4.1. Plant Material and Infectious Virus Construct
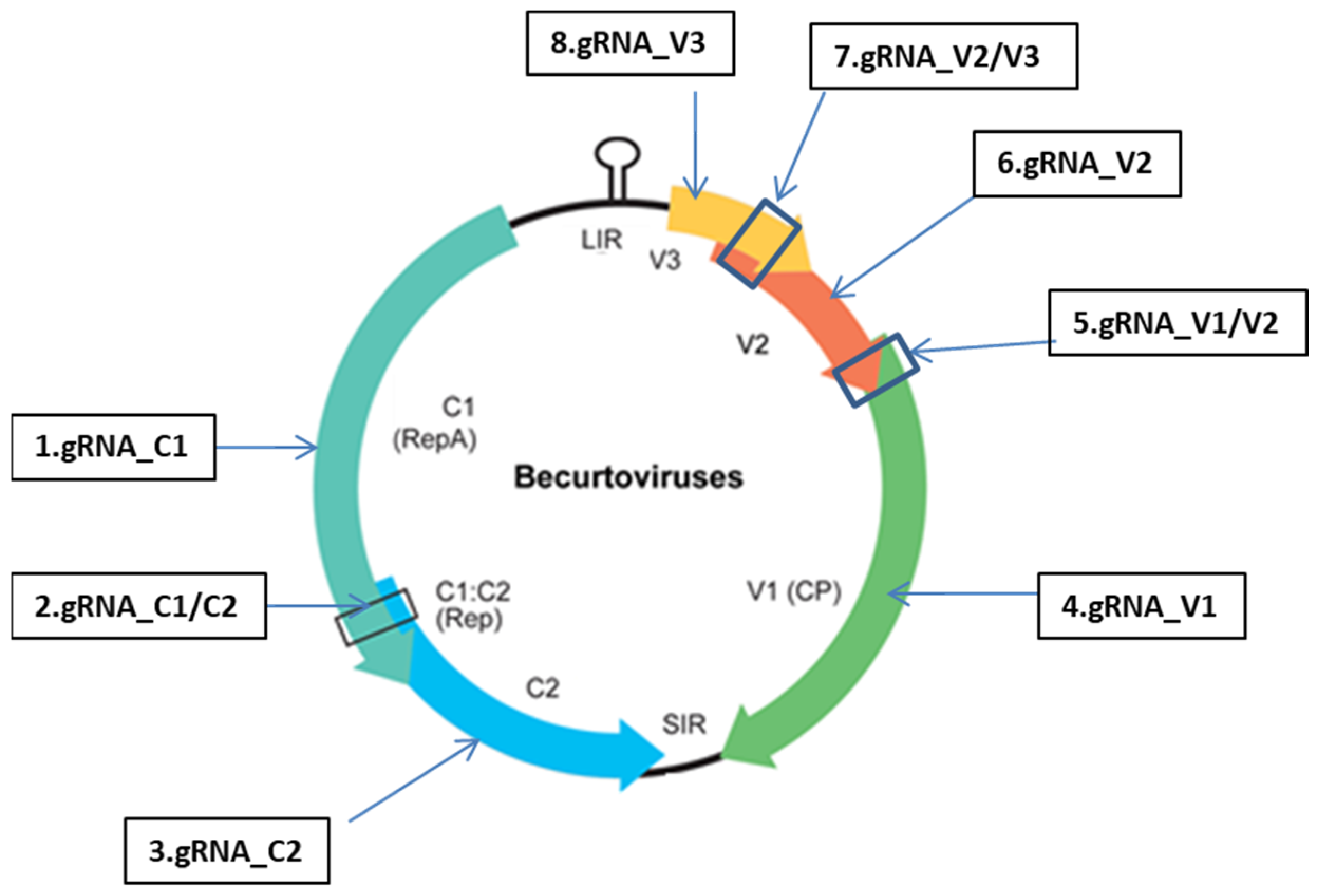
4.2. Designing the gRNAs Targeting the BCTIV Genome
4.3. Development of Multiplexed gRNA-Cas9 Modules for Efficient Knockout of BCTIV
4.4. Expression Analysis of Cas9 by Quantitative Reverse Transcriptase PCR (qRT-PCR)
4.5. Agroinoculation of gRNA/Cas9 Constructs and BCTIV on the Sugar Beet for Transient Expression Assay
4.6. Testing the Efficiency of gRNA/Cas9 on BCTIV Replication and Spread in Sugar Beet
- Test of gRNA/Cas9 efficiency on BCTIV spread in sugar beet via RCA assay
- qPCR-based detection of the gRNA/Cas9 efficiency on BCTIV replication and accumulation
- Mutation detection on BCTIV genome by loss of restriction site assay and Sanger sequencing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Varsani, A.; Martin, D.P.; Navas-Castillo, J.; Moriones, E.; Hernández-Zepeda, C.; Idris, A.; Zerbini, F.M.; Brown, J.K. Revisiting the classification of curtoviruses based on genome-wide pairwise identity. Arch. Virol. 2014, 159, 1873–1882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strausbaugh, C.A.; Eujayl, I.A. Beet curly top virus Strains Associated with Sugar Beet in Idaho, Oregon, and a Western U.S. Collection. Plant Dis. 2017, 101, 1373–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strausbaugh, C.A.; Wenninger, E.J.; Eujayl, I.A. Length of efficacy for control of curly top in sugar beet with seed and foliar insecticides. Plant Dis. 2016, 100, 1364–1370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heydarnejad, J.; Abhari, E.H.; Yazdi, H.R.B.; Massumi, H. Curly Top of Cultivated Plants and Weeds and Report of a Unique Curtovirus from Iran. J. Phytopathol. 2007, 155, 321–325. [Google Scholar] [CrossRef]
- Heydarnejad, J.; Keyvani, N.; Razavinejad, S.; Massumi, H.; Varsani, A. Fulfilling Koch’s postulates for beet curly top Iran virus and proposal for consideration of new genus in the family Geminiviridae. Arch. Virol. 2012, 158, 435–443. [Google Scholar] [CrossRef]
- Kamali, M.; Heydarnejad, J.; Pouramini, N.; Masumi, H.; Farkas, K.; Kraberger, S.; Varsani, A. Genome Sequences of Beet curly top Iran virus, Oat dwarf virus, Turnip curly top virus, and Wheat dwarf virus Identified in Leafhoppers. Genome Announc. 2017, 5, e01674-16. [Google Scholar] [CrossRef] [Green Version]
- Tahan, V.; Heydarnejad, J.; Jafarpour, B. Characterization of Beet curly top Iran virus infecting eggplant and pepper in north-eastern Iran. Indian Phytopathol. 2020, 73, 577–581. [Google Scholar] [CrossRef]
- Yıldırım, K.; Kavas, M.; Kaya, R.; Seçgin, Z.; Can, C.; Sevgen, I.; Saraç, G.; Tahan, V. Genome-based identification of beet curly top Iran virus infecting sugar beet in Turkey and investigation of its pathogenicity by agroinfection. J. Virol. Methods 2021, 300, 114380. [Google Scholar] [CrossRef] [PubMed]
- Varsani, A.; Navas-Castillo, J.; Moriones, E.; Hernández-Zepeda, C.; Idris, A.; Brown, J.K.; Zerbini, F.M.; Martin, D.P. Establishment of three new genera in the family Geminiviridae: Becurtovirus, Eragrovirus and Turncurtovirus. Arch. Virol. 2014, 159, 2193–2203. [Google Scholar] [CrossRef]
- Kaya, R. Şeker Pancarı Tepe Kıvırcıklığı (Beet Curly Top Virüs) Hastalığı. Anadolu Tarım Bilim. Derg. 2013, 28, 52–58. [Google Scholar] [CrossRef]
- Kaya, R.; Gürkan, Ş. Ankara Şeker Fabrikası şeker pancarı ekim alanlarında şeker pancarı tepe kıvırcıklığı hastalığının incelenmesi. Şeker Enstitüsü Rap. 2019, 23, 1–3. [Google Scholar]
- Ji, X.; Zhang, H.; Zhang, Y.; Wang, Y.; Gao, C. Establishing a CRISPR-Cas immune system conferring DNA virus resistance in plants. Nat. Plants 2015, 1, 15144. [Google Scholar] [CrossRef] [PubMed]
- Baltes, N.J.; Hummel, A.W.; Konecna, E.; Cegan, R.; Bruns, A.N.; Bisaro, D.M.; Voytas, D.F. Conferring resistance to geminiviruses with the CRISPR–Cas prokaryotic immune system. Nat. Plants 2015, 1, 15145. [Google Scholar] [CrossRef]
- Ali, Z.; Abulfaraj, A.; Idris, A.; Ali, S.; Tashkandi, M.; Mahfouz, M.M. CRISPR/Cas9-mediated viral interference in plants. Genome Biol. 2015, 16, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, K.; Han, T.; Xie, K.; Zhao, J.; Song, J.; Liu, Y. Engineer complete resistance to Cotton Leaf Curl Multan virus by the CRISPR/Cas9 system in Nicotiana benthamiana. Phytopathol. Res. 2019, 1, 9. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.; Zhai, Y.; Ortiz, J.; Neff, M.; Mandal, B.; Mukherjee, S.K.; Pappu, H.R. Multiplexed editing of a begomovirus genome restricts escape mutant formation and disease development. PLoS ONE 2019, 14, e0223765. [Google Scholar] [CrossRef] [Green Version]
- Aksoy, E.; Yildirim, K.; Kavas, M.; Kayihan, C.; Yerlikaya, B.A.; Calik, I.; Sevgen, I.; Demirel, U. General guidelines for CRISPR/Cas-based genome editing in plants. Mol. Biol. Rep. 2022, 49, 12151–12164. [Google Scholar] [CrossRef]
- Miladinovic, D.; Antunes, D.; Yildirim, K.; Bakhsh, A.; Cvejić, S.; Kondić-Špika, A.; Jeromela, A.M.; Opsahl-Sorteberg, H.-G.; Zambounis, A.; Hilioti, Z. Targeted plant improvement through genome editing: From laboratory to field. Plant Cell Rep. 2021, 40, 935–951. [Google Scholar] [CrossRef] [PubMed]
- Robertson, G.; Burger, J.; Campa, M. CRISPR/Cas-based tools for the targeted control of plant viruses. Mol. Plant Pathol. 2022, 23, 1701–1718. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.K.; Fauquet, C.M.; Briddon, R.W.; Zerbini, M.; Moriones, E.; Navas-Castillo, J. Geminivirdae. In Virus Taxonomy: Classification and Nomenclature of Viruses; Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adamas, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier Academic Press: San Diego, CA, USA, 2011; pp. 251–374. [Google Scholar]
- Wu, B.; Shang, X.; Schubert, J.; Habekuß, A.; Elena, S.F.; Wang, X. Global-scale computational analysis of genomic sequences reveals the recombination pattern and coevolution dynamics of cereal-infecting geminiviruses. Sci. Rep. 2015, 5, 8153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doudna, J.A.; Charpentier, E. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, S.S.E.A.; Mansoor, S.; Ali, Z.; Tashkandi, M.; Mahfouz, M.M. Engineering Plants for Geminivirus Resistance with CRISPR/Cas9 System. Trends Plant Sci. 2016, 21, 279–281. [Google Scholar] [CrossRef] [PubMed]
- Soleimani, R.; Matic, S.; Taheri, H.; Behjatnia, S.A.A.; Vecchiati, M.; Izadpanah, K.; Accotto, G.P. The unconventional geminivirus Beet curly top Iran virus: Satisfying Koch’s postulates and determining vector and host range. Ann. Appl. Biol. 2012, 162, 174–181. [Google Scholar] [CrossRef]
- Yazdi, H.R.B.; Heydarnejad, J.; Massumi, H. Genome characterization and genetic diversity of beet curly top Iran virus: A geminivirus with a novel nonanucleotide. Virus Genes 2008, 36, 539–545. [Google Scholar] [CrossRef]
- Ebrahimi, S.; Eini, O.; Koolivand, D.; Varrelmann, M. The Rep and C1 of Beet curly top Iran virus represent pathogenicity factors and induce hypersensitive response in Nicotiana benthamiana plants. Virus Genes 2022, 58, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Bahari, A.; Castillo, A.G.; Safaie, N.; Bejarano, E.R.; Luna, A.P.; Shams-Bakhsh, M. Functional Analysis of V2 Protein of Beet Curly Top Iran Virus. Plants 2022, 11, 3351. [Google Scholar] [CrossRef]
- Ebrahimi, S.; Alexandra Babler, A.; Eini, O.; Yildirim, Z.; Wassenegger, W.; Krczal, G.; Uslu, V.V. Beet curly top Iran virus Rep and V2 gene work as silencing suppressors through separate mechanisms. bioRxiv 2022. [Google Scholar] [CrossRef]
- Luna, A.P.; Romero-Rodríguez, B.; Rosas-Díaz, T.; Cerero, L.; Rodríguez-Negrete, E.A.; Castillo, A.G.; Bejarano, E.R. Characterization of Curtovirus V2 Protein, a Functional Homolog of Begomovirus V2. Front. Plant Sci. 2020, 11, 835. [Google Scholar] [CrossRef]
- Kis, A.; Hamar, É.; Tholt, G.; Bán, R.; Havelda, Z. Creating highly efficient resistance against wheat dwarf virus in barley by employing CRISPR/Cas9 system. Plant Biotechnol. J. 2019, 17, 1004–1006. [Google Scholar] [CrossRef]
- Liu, H.; Soyars, C.L.; Li, J.; Fei, Q.; He, G.; Peterson, B.A.; Meyers, B.C.; Nimchuk, Z.L.; Wang, X. CRISPR/Cas9-mediated resistance to cauliflower mosaic virus. Plant Direct 2018, 2, e00047. [Google Scholar] [CrossRef]
- Mehta, D.; Stürchler, A.; Anjanappa, R.B.; Zaidi, S.S.-e.-A.; Hirsch-Hoffmann, M.; Gruissem, W.; Vanderschuren, H. Linking CRISPR-Cas9 interference in cassava to the evolution of editing-resistant geminiviruses. Genome Biol. 2019, 20, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tashkandi, M.; Ali, Z.; Aljedaani, F.; Shami, A.; Mahfouz, M.M. Engineering resistance against Tomato yellow leaf curl virus via the CRISPR/Cas9 system in tomato. Plant Signal. Behav. 2018, 13, e1525996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yildirim, K.; Boylu, B.; Atici, E.; TKahraman Akkaya, M.S. In Turkish wheat cultivars the resistance allele of LR34 is ineffective against leaf rust. J. Plant Dis. Prot. 2012, 119, 135–141. [Google Scholar] [CrossRef]
- Shepherd, D.N.; Martin, D.P.; Lefeuvre, P.; Monjane, A.L.; Owor, B.E.; Rybicki, E.; Varsani, A. A protocol for the rapid isolation of full geminivirus genomes from dried plant tissue. J. Virol. Methods 2008, 149, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Secgin, Z.; Uluisik, S.; Yıldırım, K.; Abdulla, M.F.; Mostafa, K.; Kavas, M. Genome-Wide Identification of the Aconitase Gene Family in Tomato (Solanum lycopersicum) and CRISPR-Based Functional Characterization of SlACO2 on Male-Sterility. Int. J. Mol. Sci. 2022, 23, 13963. [Google Scholar] [CrossRef]
- Seçgin, Z.; Kavas, M.; Yildirim, K. Optimization of Agrobacterium-mediated transformation and regeneration for CRISPR/ Cas9 genome editing of commercial tomato cultivars. Turk. J. Agric. For. 2021, 45, 704–716. [Google Scholar] [CrossRef]
- Xing, H.-L.; Dong, L.; Wang, Z.-P.; Zhang, H.-Y.; Han, C.-Y.; Liu, B.; Wang, X.-C.; Chen, Q.-J. A CRISPR/Cas9 toolkit for multiplex genome editing in plants. BMC Plant Biol. 2014, 14, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Yıldırım, K.; Yağcı, A.; Sucu, S.; Tunç, S. Responses of grapevine rootstocks to drought through altered root system architecture and root transcriptomic regulations. Plant Physiol. Biochem. 2018, 127, 256–268. [Google Scholar] [CrossRef]
- Kavas, M.; Mostafa, K.; Seçgin, Z.; Yerlikaya, B.A.; Yıldırım, K.; Gökdemir, G. Genome-wide analysis of duf221 domain-containing gene family in common bean and identification of its role on abiotic and phytohormone stress response. Genet. Resour. Crop Evol. 2022, 70, 169–188. [Google Scholar] [CrossRef]
- Montazeri, R.; Shams-Bakhsh, M.; Mahmoudi, S.B.; Rajabi, A. Evaluation of sugar beet lines for resistance to beet curly top viruses. Euphytica 2016, 210, 31–40. [Google Scholar] [CrossRef]
- Gadiou, S.; Ripl, J.; Jaňourová, B.; Jarošová, J.; Kundu, J.K. Real-time PCR assay for the discrimination and quantification of wheat and barley strains of Wheat dwarf virus. Virus Genes 2011, 44, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Jarošová, J.; Kundu, J.K. Validation of reference genes as internal control for studying viral infections in cereals by quanti-587 tative real-time RT-PCR. BMC Plant Biol. 2010, 10, 146. [Google Scholar]
- Brinkman, E.K.; Chen, T.; Amendola, M.; Van Steensel, B. Easy quantitative assessment of genome editing by sequence trace decomposition. Nucleic Acids Res. 2014, 42, e168. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy Plants without Symptoms (%) | Plants with Mild Symptoms (%) | Plants with Moderate and Severe Symptoms (%) | Average Weight of Leaves(g) | Average Weight of Roots(g) | Rate of Reduction in Biomass (%) | |
|---|---|---|---|---|---|---|
| Mock vector agroinoculated controls | 100 | 0 | 0 | 124 ± 12 | 365 ± 24 | - |
| BCTIV agroinoculated positive controls | 5 | 15 | 80 | 36 ± 4 | 123 ± 12 | 68 |
| C1-REPA | 60 | 15 | 25 | 90 ± 5 | 256 ± 21 | 30 |
| C1/C2-REPA/REPB overlapping region | 45 | 30 | 35 | 76 ± 11 | 196 ± 9 | 45 |
| C2- REPB | 75 | 10 | 15 | 96 ± 6 | 266 ± 14 | 26 |
| V1- Capsid protein (CP) | 65 | 15 | 20 | 88 ± 3 | 286 ± 24 | 24 |
| V1/V2-CP/MP overlapping region | 85 | 15 | 0 | 106 ± 4 | 316 ± 16 | 14 |
| V2- Movement protein (MP) | 65 | 10 | 25 | 74 ± 5 | 275 ± 13 | 29 |
| V2/V3-MP/ssDNA Reg. overlapping region | 70 | 10 | 15 | 108 ± 12 | 321 ± 18 | 13 |
| V3 ssDNA regulator gene | 75 | 10 | 15 | 84 ± 2 | 295 ± 9 | 23 |
| Multiplex CRISPR (C1/C2/V1-V2/V2-V3) | 90 | 10 | 0 | 115 ± 9 | 322 ± 14 | 11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yıldırım, K.; Kavas, M.; Küçük, İ.S.; Seçgin, Z.; Saraç, Ç.G. Development of Highly Efficient Resistance to Beet Curly Top Iran Virus (Becurtovirus) in Sugar Beet (B. vulgaris) via CRISPR/Cas9 System. Int. J. Mol. Sci. 2023, 24, 6515. https://doi.org/10.3390/ijms24076515
Yıldırım K, Kavas M, Küçük İS, Seçgin Z, Saraç ÇG. Development of Highly Efficient Resistance to Beet Curly Top Iran Virus (Becurtovirus) in Sugar Beet (B. vulgaris) via CRISPR/Cas9 System. International Journal of Molecular Sciences. 2023; 24(7):6515. https://doi.org/10.3390/ijms24076515
Chicago/Turabian StyleYıldırım, Kubilay, Musa Kavas, İlkay Sevgen Küçük, Zafer Seçgin, and Çiğdem Gökcek Saraç. 2023. "Development of Highly Efficient Resistance to Beet Curly Top Iran Virus (Becurtovirus) in Sugar Beet (B. vulgaris) via CRISPR/Cas9 System" International Journal of Molecular Sciences 24, no. 7: 6515. https://doi.org/10.3390/ijms24076515