
Genome-Scale Analysis of the Grapevine KCS Genes Reveals Its Potential Role in Male Sterility

Abstract
:1. Introduction
2. Results
2.1. Identification and Annotation of KCS Gene Family Members in Grapevine
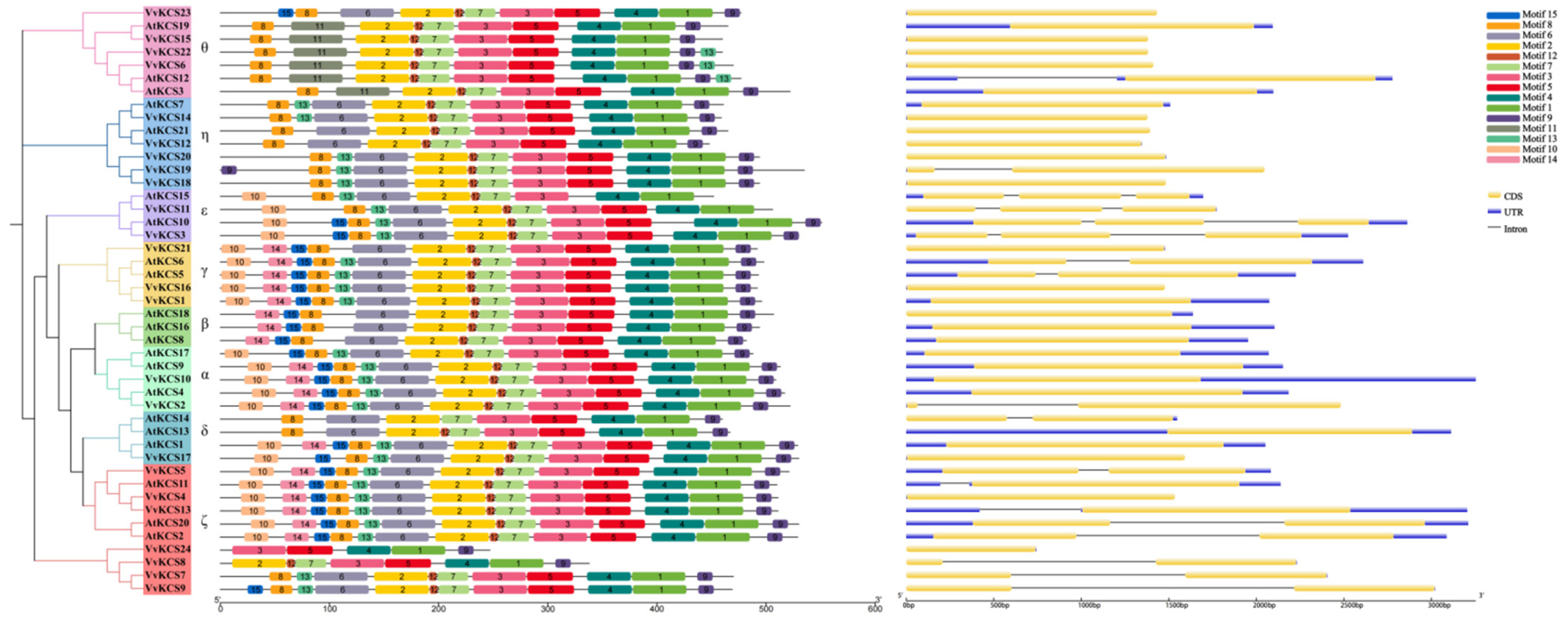
2.2. Phylogenetic Motif Composition and Structural Analysis of VvKCS Genes
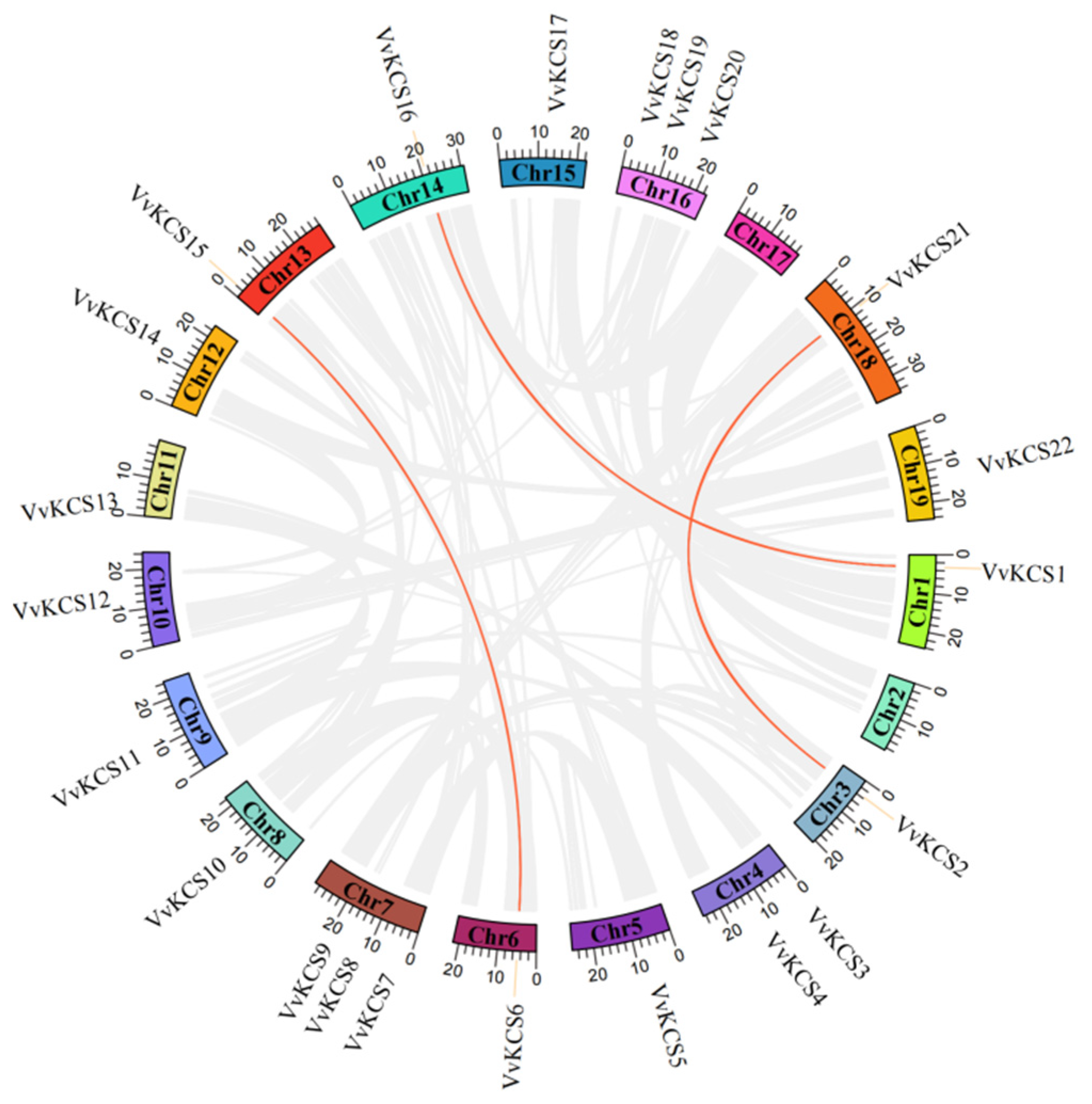
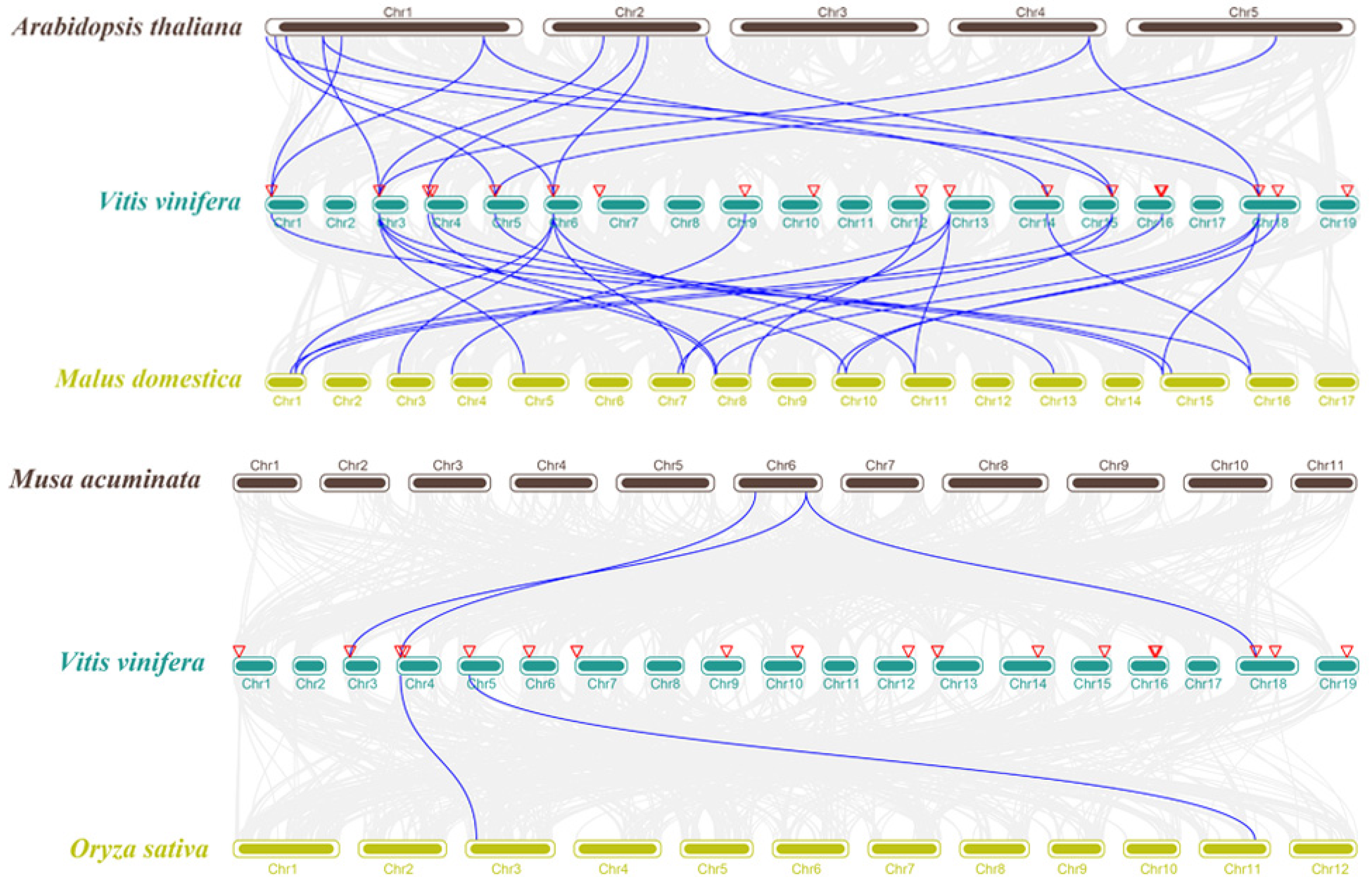
2.3. Synteny Analysis of VvKCS Genes
2.4. Expression of VvKCS Genes in Different Tissues of Grapevine
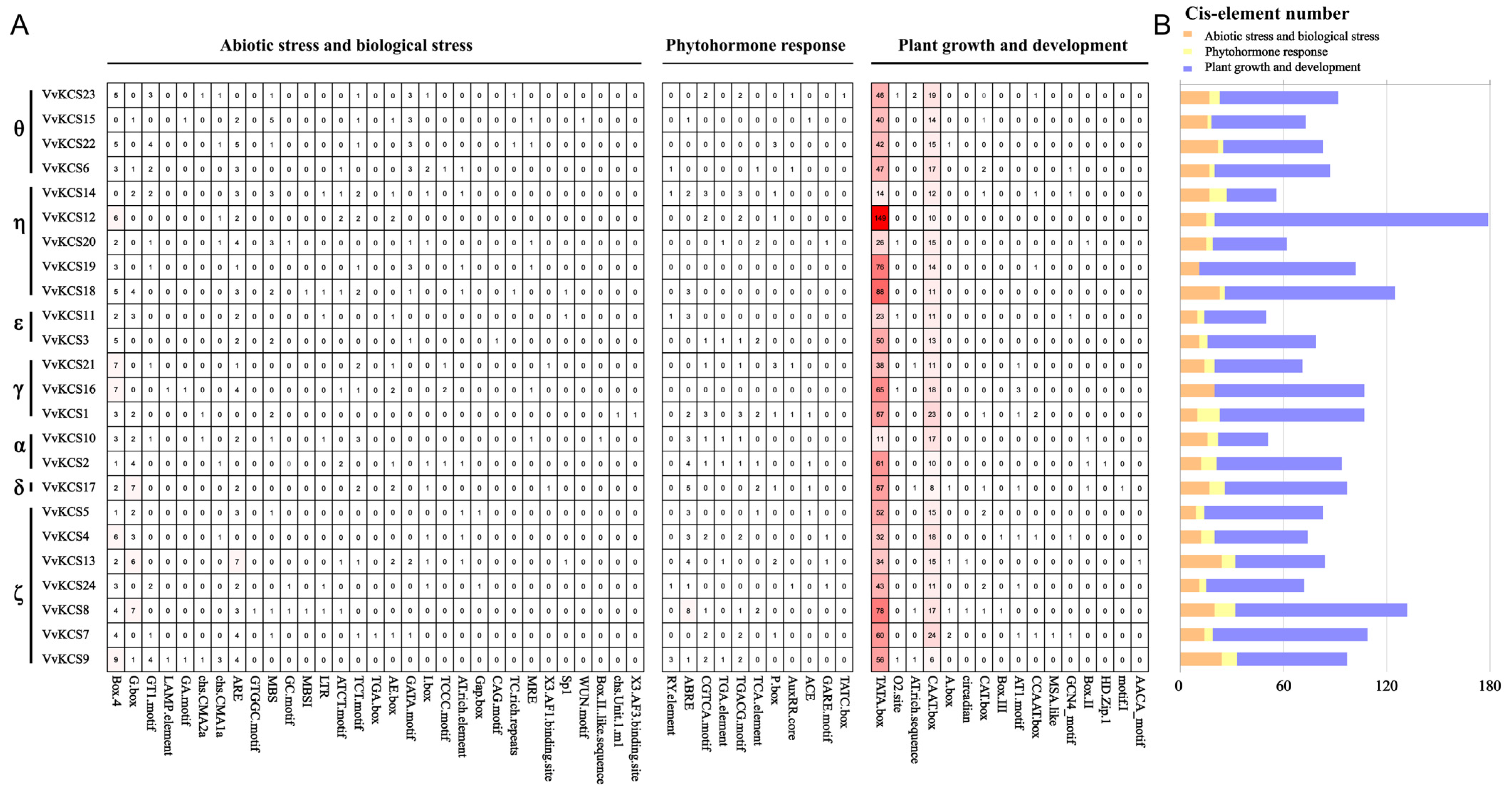
2.5. Analysis of Cis-Acting Elements in the Promoter Regions of VvKCS Genes
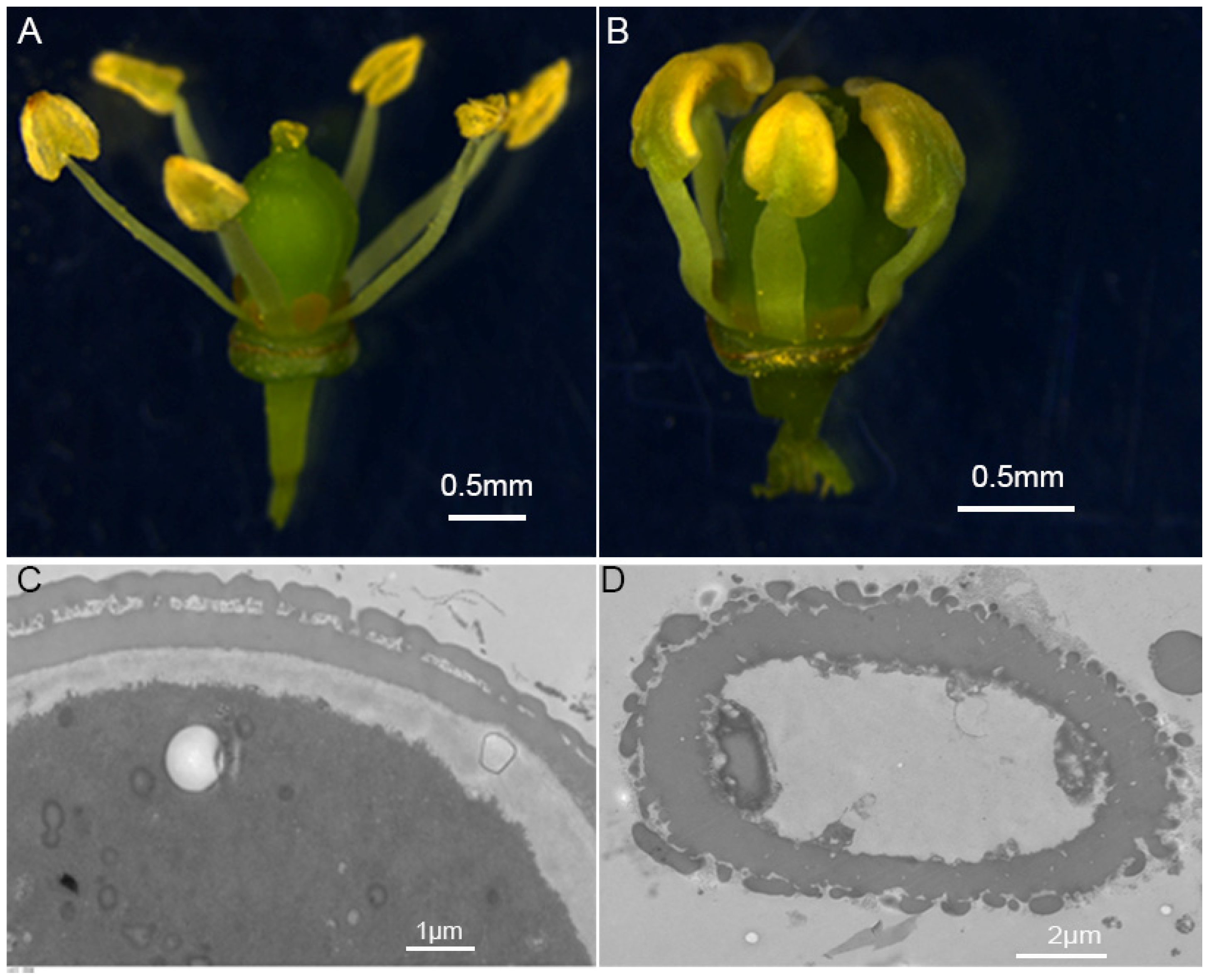
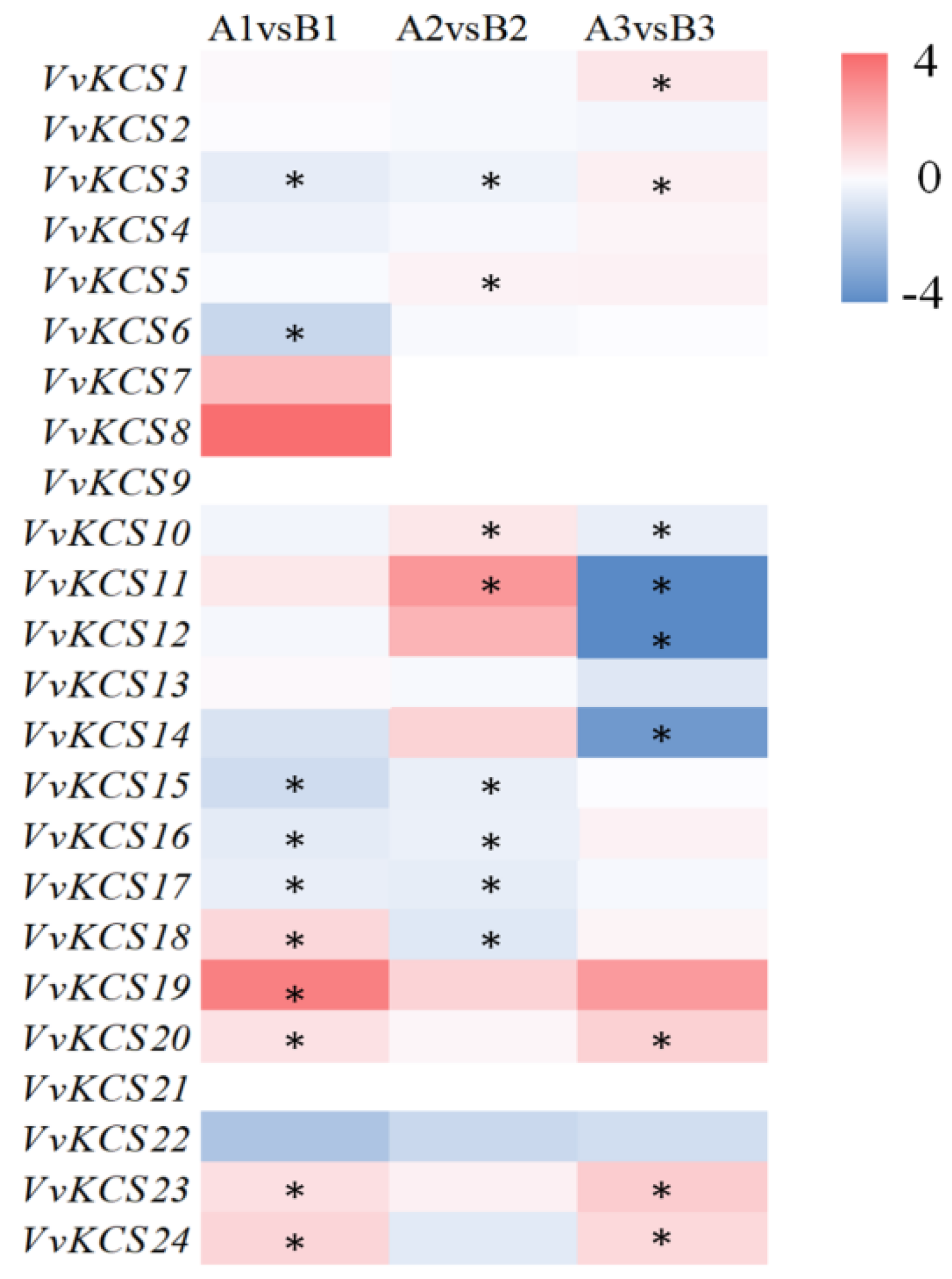
2.6. Expression Analysis of VvKCS Genes during the Critical Period of Pollen Abortion in Grapevine ‘Y−14’
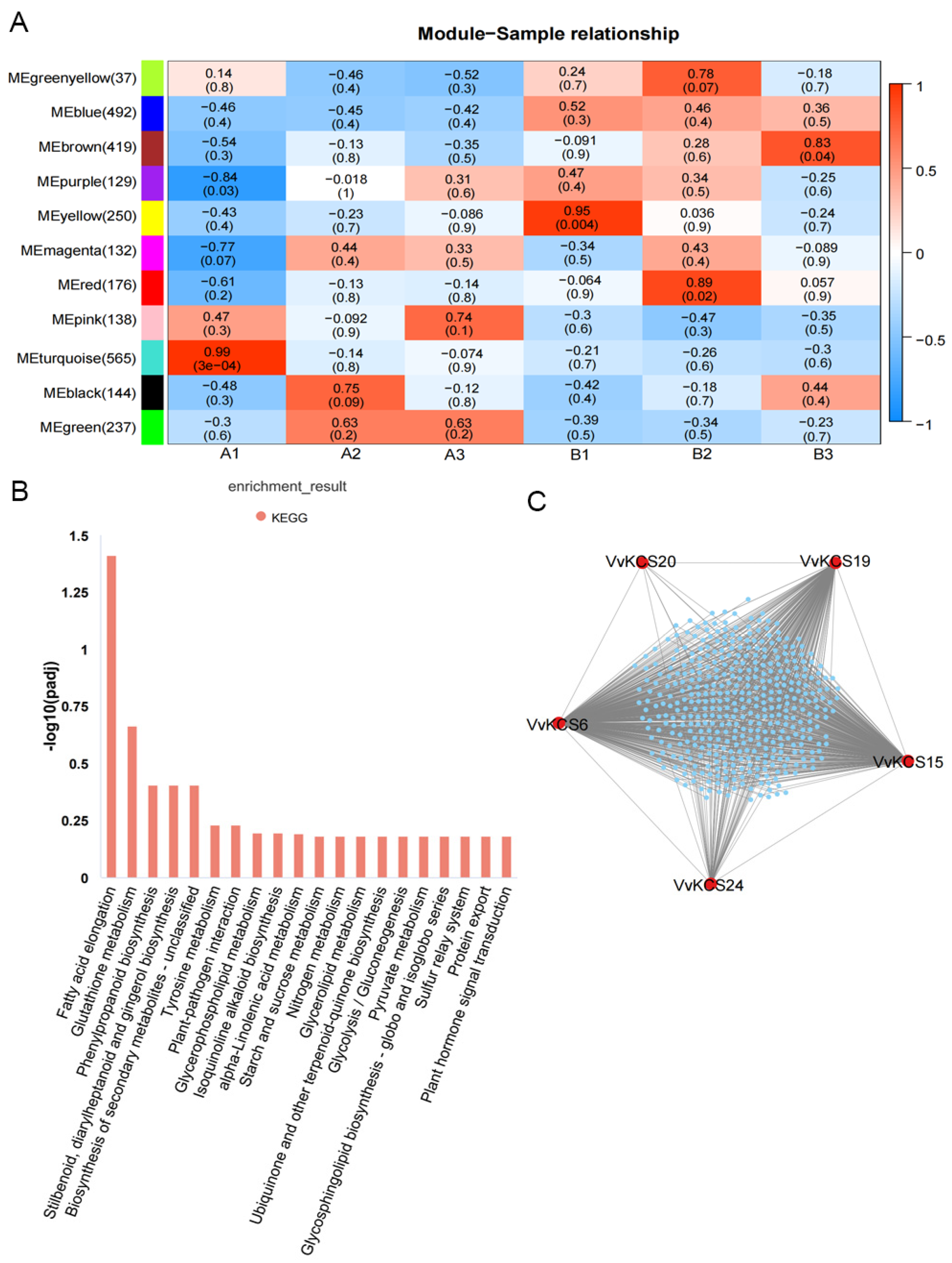
2.7. Screening for Key VvKCS Genes Regulating Pollen Development by WGCNA
3. Discussion
3.1. Systematic Analysis Lays the Foundation for Functional Characterization of the VvKCS Gene
3.2. VvKCS Genes May Be Involved in Pollen Cell Wall Formation
4. Materials and Methods
4.1. Genome-Wide Identification of VvKCS Family Genes in Grapevine
4.2. Phylogenetic, Motif and Gene Structure Analysis
4.3. Cis-Acting Elements Analysis of VvKCSs
4.4. Gene Duplication and Syntenic Analysis of VvKCSs
4.5. Expression Analysis of VvKCSs in Grapevine Tissues
4.6. Plant Materials and Phenotypic Analysis of ‘Y−14’
4.7. RNA Isolation and RNA-seq Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jianying, F.; Xia, W.; Zetian, F.; Weisong, M. Assessment of consumers’ perception and cognition toward table grape con-sumption in China. Br. Food J. 2014, 116, 611–628. [Google Scholar] [CrossRef]
- Zheng, H.; Yu, X.; Yuan, Y.; Zhang, Y.; Zhang, Z.; Zhang, J.; Zhang, M.; Ji, C.; Liu, Q.; Tao, J. The VviMYB80 gene is abnormally expressed in Vitis vinifera L. cv. ‘Zhong Shan Hong’and its expression in tobacco driven by the 35S promoter causes male sterility. Plant Cell Physiol. 2016, 57, 540–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewandowska, M.; Keyl, A.; Feussner, I. Wax biosynthesis in response to danger: Its regulation upon abiotic and biotic stress. New Phytol. 2020, 227, 698–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haslam, T.M.; Kunst, L. Extending the story of very-long-chain fatty acid elongation. Plant Sci. 2013, 210, 93–107. [Google Scholar] [CrossRef]
- Wan, X.; Wu, S.; Li, Z.; An, X.; Tian, Y. Lipid metabolism: Critical roles in male fertility and other aspects of reproductive development in plants. Mol. Plant 2020, 13, 955–983. [Google Scholar] [CrossRef]
- Ariizumi, T.; Toriyama, K. Genetic Regulation of Sporopollenin Synthesis and Pollen Exine Development. Annu. Rev. Plant Biol. 2011, 62, 437–460. [Google Scholar] [CrossRef]
- Shi, J.; Cui, M.; Yang, L.; Kim, Y.-J.; Zhang, D. Genetic and Biochemical Mechanisms of Pollen Wall Development. Trends Plant Sci. 2015, 20, 741–753. [Google Scholar] [CrossRef]
- Ishiguro, S.; Kawai-Oda, A.; Ueda, J.; Nishida, I.; Okada, K. The Defective in Anther Dehiscence1 gene encodes a novel phospholipase A1 catalyzing the initial step of jasmonic acid biosynthesis, which synchronizes pollen maturation, anther dehiscence, and flower opening in Arabidopsis. Plant Cell 2001, 13, 2191–2209. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Chen, Y.; Charnikhova, T.; Mulder, P.P.J.; Heijmans, J.; Hoogenboom, A.; Agalou, A.; Michel, C.; Morel, J.-B.; Dreni, L.; et al. OsJAR1 is required for JA-regulated floret opening and anther dehiscence in rice. Plant Mol. Biol. 2014, 86, 19–33. [Google Scholar] [CrossRef]
- Fiebig, A.; Mayfield, J.A.; Miley, N.L.; Chau, S.; Fischer, R.L.; Preuss, D. Alterations in CER6, a gene identical to CUT1, differentially affect long-chain lipid content on the surface of pollen and stems. Plant Cell 2000, 12, 2001–2008. [Google Scholar] [CrossRef] [Green Version]
- Zhan, H.; Xiong, H.; Wang, S.; Yang, Z.-N. Anther Endothecium-Derived Very-Long-Chain Fatty Acids Facilitate Pollen Hydration in Arabidopsis. Mol. Plant 2018, 11, 1101–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acosta, I.F.; Laparra, H.; Romero, S.P.; Schmelz, E.; Hamberg, M.; Mottinger, J.P.; Moreno, M.A.; Dellaporta, S.L. tasselseed1 Is a Lipoxygenase Affecting Jasmonic Acid Signaling in Sex Determination of Maize. Science 2009, 323, 262–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Lee, S.B.; Suh, M.C. Arabidopsis 3-ketoacyl-CoA synthase 4 is essential for root and pollen tube growth. J. Plant Biol. 2021, 64, 155–165. [Google Scholar] [CrossRef]
- Joubès, J.; Raffaele, S.; Bourdenx, B.; Garcia, C.; Laroche-Traineau, J.; Moreau, P.; Domergue, F.; Lessire, R. The VLCFA elongase gene family in Arabidopsis thaliana: Phylogenetic analysis, 3D modelling and expression profiling. Plant Mol. Biol. 2008, 67, 547–566. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.C.; Purugganan, M.D. The early stages of duplicate gene evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Wu, N.; Song, W.; Yin, G.; Qin, Y.; Yan, Y.; Hu, Y. Soybean (Glycine max) expansin gene superfamily origins: Segmental and tandem duplication events followed by divergent selection among subfamilies. BMC Plant Biol. 2014, 14, 93. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Meng, S.; Su, W.; Bao, Y.; Lu, Y.; Yin, W.; Liu, C.; Xia, X. Genome-Wide Analysis of Multiple Organellar RNA Editing Factor Family in Poplar Reveals Evolution and Roles in Drought Stress. Int. J. Mol. Sci. 2019, 20, 1425. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Wang, J.; Lin, W.; Li, S.; Li, H.; Zhou, J.; Ni, P.; Dong, W.; Hu, S.; Zeng, C.; et al. The Genomes of Oryza sativa: A History of Duplications. PLoS Biol. 2005, 3, e38. [Google Scholar] [CrossRef] [Green Version]
- Hong, B.; Tao, J.; Nong, H. Analysis of antioxidant enzymes activities and endogenous hormone contents in male sterile plant of ‘Shine Muscat’seeding during bud development. Acta Bot. Boreali-Occident. Sin. 2022, 42, 444–452. [Google Scholar]
- Guo, H.-S.; Zhang, Y.-M.; Sun, X.-Q.; Li, M.-M.; Hang, Y.-Y.; Xue, J.-Y. Evolution of the KCS gene family in plants: The history of gene duplication, sub/neofunctionalization and redundancy. Mol. Genet. Genom. 2015, 291, 739–752. [Google Scholar] [CrossRef]
- Heidari, P.; Puresmaeli, F.; Mora-Poblete, F. Genome-Wide Identification and Molecular Evolution of the Magnesium Transporter (MGT) Gene Family in Citrullus lanatus and Cucumis sativus. Agronomy 2022, 12, 2253. [Google Scholar] [CrossRef]
- Puresmaeli, F.; Heidari, P.; Lawson, S. Insights into the Sulfate Transporter Gene Family and Its Expression Patterns in Durum Wheat Seedlings under Salinity. Genes 2023, 14, 333. [Google Scholar] [CrossRef]
- Vinogradov, A.E. Noncoding DNA, isochores and gene expression: Nucleosome formation potential. Nucleic Acids Res. 2005, 33, 559–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J. Evolution by gene duplication: An update. Trends Ecol. Evol. 2003, 18, 292–298. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Zhang, Z.; Ni, E.; Lin, J.; Peng, G.; Huang, J.; Zhu, L.; Deng, L.; Yang, F.; Luo, Q. HMS1 interacts with HMS1I to regulate very-long-chain fatty acid biosynthesis and the humidity-sensitive genic male sterility in rice (Oryza sativa). New Phytol. 2020, 225, 2077–2093. [Google Scholar] [CrossRef]
- Morant, M.; Jørgensen, K.; Schaller, H.; Pinot, F.; Møller, B.L.; Werck-Reichhart, D.; Bak, S. CYP703 Is an Ancient Cytochrome P450 in Land Plants Catalyzing in-Chain Hydroxylation of Lauric Acid to Provide Building Blocks for Sporopollenin Synthesis in Pollen. Plant Cell 2007, 19, 1473–1487. [Google Scholar] [CrossRef] [Green Version]
- Mayfield, J.A.; Fiebig, A.; Johnstone, S.E.; Preuss, D. Gene Families from the Arabidopsis thaliana Pollen Coat Proteome. Science 2001, 292, 2482–2485. [Google Scholar] [CrossRef] [Green Version]
- Lavrentovich, M.O.; Horsley, E.M.; Radja, A.; Sweeney, A.M.; Kamien, R.D. First-order patterning transitions on a sphere as a route to cell morphology. Proc. Natl. Acad. Sci. USA 2016, 113, 5189–5194. [Google Scholar] [CrossRef] [Green Version]
- Gabarayeva, N.I.; Grigorjeva, V.V.; Lavrentovich, M.O. Artificial pollen walls simulated by the tandem processes of phase separation and self-assembly in vitro. New Phytol. 2019, 225, 1956–1973. [Google Scholar] [CrossRef] [PubMed]
- James, M.; Simmons, C.; Wise, C.J.; Jones, G.E.; Morris, G.E. Evidence for a utrophin-glycoprotein complex in cultured cell lines and a possible role in cell adhesion. Biochem. Soc. Trans. 1995, 23, 398S. [Google Scholar] [CrossRef] [Green Version]
- Millar, A.A.; Kunst, L. The natural genetic variation of the fatty-acyl composition of seed oils in different ecotypes of Arabidopsis thaliana. Phytochemistry 1999, 52, 1029–1033. [Google Scholar] [CrossRef]
- Franke, R.; Höfer, R.; Briesen, I.; Emsermann, M.; Efremova, N.; Yephremov, A.; Schreiber, L. The DAISY gene from Ara-bidopsis encodes a fatty acid elongase condensing enzyme involved in the biosynthesis of aliphatic suberin in roots and the chalazamicropyle region of seeds. Plant J. 2009, 57, 80–95. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhan, H.; Lu, J.; Xiong, S.; Yang, N.; Yuan, H.; Yang, Z.-N. Tapetal 3-Ketoacyl-Coenzyme A Synthases Are Involved in Pollen Coat Lipid Accumulation for Pollen-Stigma Interaction in Arabidopsis. Front. Plant Sci. 2021, 12, 770311. [Google Scholar] [CrossRef]
- Mitchell, A.L.; Attwood, T.K.; Babbitt, P.C.; Blum, M.; Bork, P.; Bridge, A.; Brown, S.D.; Chang, H.-Y.; El-Gebali, S.; Fraser, M.I.; et al. InterPro in 2019: Improving coverage, classification and access to protein sequence annotations. Nucleic Acids Res. 2019, 47, D351–D360. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Xie, T.; Chen, C.; Luan, A.; Long, J.; Li, C.; Ding, Y.; He, Y. Genome-wide organization and expression profiling of the R2R3-MYB transcription factor family in pineapple (Ananas comosus). BMC Genom. 2017, 18, 503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fasoli, M.; Dal Santo, S.; Zenoni, S.; Tornielli, G.B.; Farina, L.; Zamboni, A.; Porceddu, A.; Venturini, L.; Bicego, M.; Murino, V.; et al. The Grapevine Expression Atlas Reveals a Deep Transcriptome Shift Driving the Entire Plant into a Maturation Program. Plant Cell 2012, 24, 3489–3505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Chen, H.; He, Y.; Xia, R. TBtools, a toolkit for biologists integrating various biological data handling tools with a user-friendly interface. BioRxiv 2018, 2018, 289660. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2009, 26, 136–138. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Dong, Y.; Nong, H.; Huang, L.; Liu, J.; Yu, X.; Zhang, Y.; Yang, L.; Hong, B.; Wang, W.; et al. VvSUN may act in the auxin pathway to regulate fruit shape in grape. Hortic. Res. 2022, 9, 200. [Google Scholar] [CrossRef]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 29–34. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Total Number of Pollen Grains Tested | Total Number of Germinated Pollen Grains | Germination Rate (%) |
|---|---|---|---|
| ‘Y−14’ | 319 | 0 | 0 |
| ‘Shine Muscat’ | 351 | 254 | 72.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, H.; Liang, Y.; Hong, B.; Xu, Y.; Ren, M.; Wang, Y.; Huang, L.; Yang, L.; Tao, J. Genome-Scale Analysis of the Grapevine KCS Genes Reveals Its Potential Role in Male Sterility. Int. J. Mol. Sci. 2023, 24, 6510. https://doi.org/10.3390/ijms24076510
Zheng H, Liang Y, Hong B, Xu Y, Ren M, Wang Y, Huang L, Yang L, Tao J. Genome-Scale Analysis of the Grapevine KCS Genes Reveals Its Potential Role in Male Sterility. International Journal of Molecular Sciences. 2023; 24(7):6510. https://doi.org/10.3390/ijms24076510
Chicago/Turabian StyleZheng, Huan, Yueting Liang, Ben Hong, Yingyi Xu, Mengfan Ren, Yixu Wang, Liyuan Huang, Lina Yang, and Jianmin Tao. 2023. "Genome-Scale Analysis of the Grapevine KCS Genes Reveals Its Potential Role in Male Sterility" International Journal of Molecular Sciences 24, no. 7: 6510. https://doi.org/10.3390/ijms24076510