
Role of Muscarinic Acetylcholine Receptors in Intestinal Epithelial Homeostasis: Insights for the Treatment of Inflammatory Bowel Disease
 ,
,
Abstract
:1. Introduction
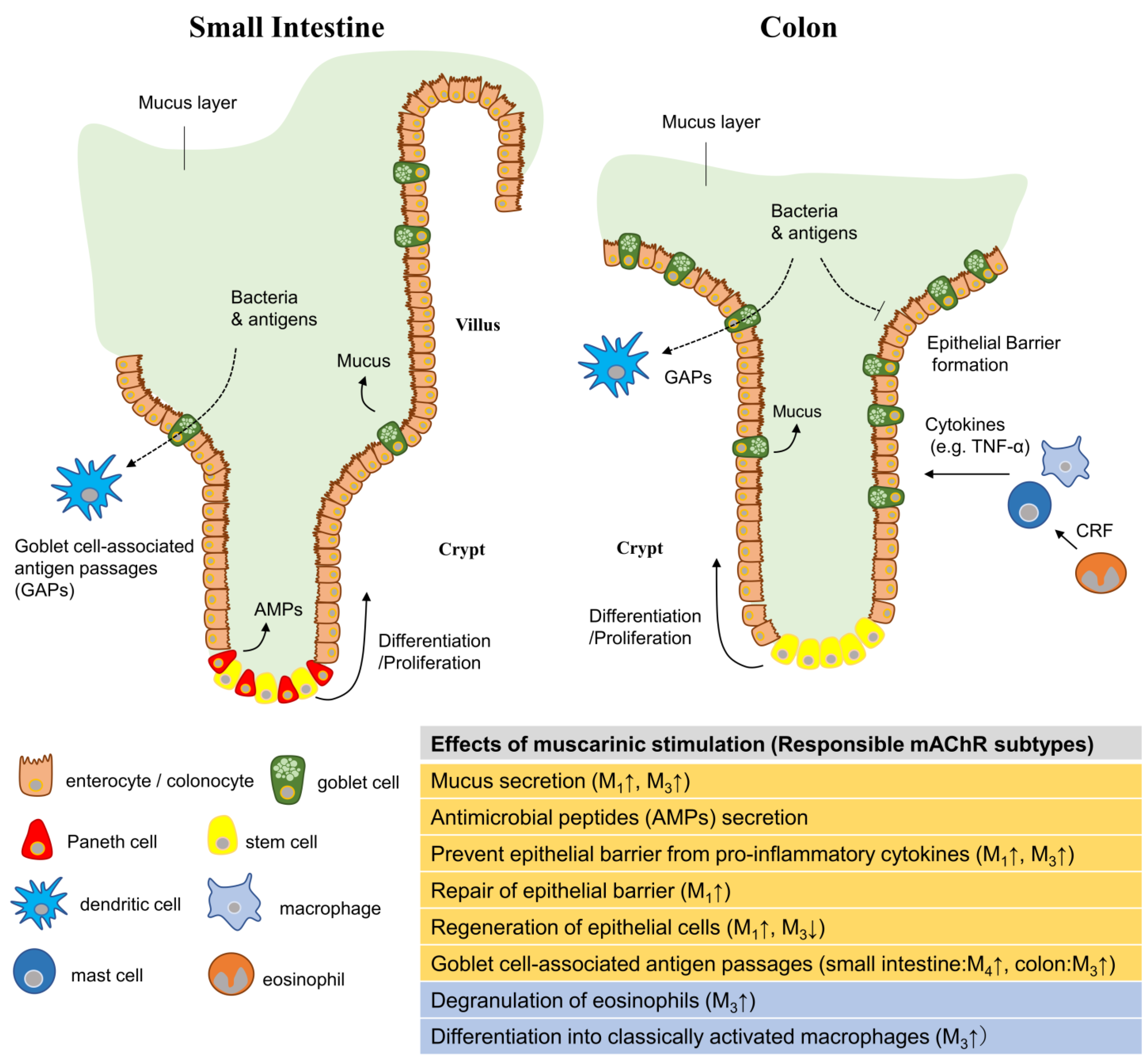
2. mAChRs in the Intestinal Epithelium
3. Role of mAChRs in Mucus Layer Maintenance
4. Role of mAChRs in the Epithelial Barrier against Inflammatory Cytokines
5. Role of mAChRs in Epithelial Barrier Repair
6. Role of mAChRs in Epithelial Cell Regeneration
7. Involvement of mAChRs in Epithelial Barrier Impairment
8. Non-Neuronal Acetylcholine
9. mAChRs as a Potential Therapeutic Target for IBD
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Irvine, E.J.; Marshall, J.K. Increased intestinal permeability precedes the onset of Crohn’s disease in a subject with familial risk. Gastroenterology 2000, 119, 1740–1744. [Google Scholar] [CrossRef] [PubMed]
- Michielan, A.; D’Inca, R. Intestinal Permeability in Inflammatory Bowel Disease: Pathogenesis, Clinical Evaluation, and Therapy of Leaky Gut. Mediat. Inflamm. 2015, 2015, 628157. [Google Scholar] [CrossRef] [PubMed]
- Tanahashi, Y.; Komori, S.; Matsuyama, H.; Kitazawa, T.; Unno, T. Functions of Muscarinic Receptor Subtypes in Gastrointestinal Smooth Muscle: A Review of Studies with Receptor-Knockout Mice. Int. J. Mol. Sci. 2021, 22, 926. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.R.; Islam, M.T.; Yazawa, T.; Hayashi, H.; Suzuki, Y.; Uwada, J.; Anisuzzaman, A.S.; Taniguchi, T. Muscarinic cholinoceptor-mediated activation of JNK negatively regulates intestinal secretion in mice. J. Pharmacol. Sci. 2015, 127, 150–153. [Google Scholar] [CrossRef]
- Keely, S.J.; Barrett, K.E. Regulation of chloride secretion. Novel pathways and messengers. Ann. N. Y. Acad. Sci. 2000, 915, 67–76. [Google Scholar] [CrossRef]
- Serafini, M.A.; Paz, A.H.; Nunes, N.S. Cholinergic immunomodulation in inflammatory bowel diseases. Brain Behav. Immun. Health 2022, 19, 100401. [Google Scholar] [CrossRef] [PubMed]
- Hirota, C.L.; McKay, D.M. Cholinergic regulation of epithelial ion transport in the mammalian intestine. Br. J. Pharmacol. 2006, 149, 463–479. [Google Scholar] [CrossRef]
- Davis, E.A.; Zhou, W.; Dailey, M.J. Evidence for a direct effect of the autonomic nervous system on intestinal epithelial stem cell proliferation. Physiol. Rep. 2018, 6, e13745. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Ohnishi, H.; Sugiura, Y.; Honda, K.; Suematsu, M.; Kawasaki, T.; Deguchi, T.; Fujii, T.; Orihashi, K.; Hippo, Y.; et al. Non-neuronal acetylcholine as an endogenous regulator of proliferation and differentiation of Lgr5-positive stem cells in mice. FEBS J. 2014, 281, 4672–4690. [Google Scholar] [CrossRef] [PubMed]
- Middelhoff, M.; Nienhuser, H.; Valenti, G.; Maurer, H.C.; Hayakawa, Y.; Takahashi, R.; Kim, W.; Jiang, Z.; Malagola, E.; Cuti, K.; et al. Prox1-positive cells monitor and sustain the murine intestinal epithelial cholinergic niche. Nat. Commun. 2020, 11, 111. [Google Scholar] [CrossRef]
- Muise, E.D.; Gandotra, N.; Tackett, J.J.; Bamdad, M.C.; Cowles, R.A. Distribution of muscarinic acetylcholine receptor subtypes in the murine small intestine. Life Sci. 2017, 169, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Knoop, K.A.; McDonald, K.G.; McCrate, S.; McDole, J.R.; Newberry, R.D. Microbial sensing by goblet cells controls immune surveillance of luminal antigens in the colon. Mucosal Immunol. 2015, 8, 198–210. [Google Scholar] [CrossRef] [PubMed]
- Greig, C.J.; Armenia, S.J.; Cowles, R.A. The M1 muscarinic acetylcholine receptor in the crypt stem cell compartment mediates intestinal mucosal growth. Exp. Biol. Med. 2020, 245, 1194–1199. [Google Scholar] [CrossRef]
- Khan, M.R.; Anisuzzaman, A.S.; Semba, S.; Ma, Y.; Uwada, J.; Hayashi, H.; Suzuki, Y.; Takano, T.; Ikeuchi, H.; Uchino, M.; et al. M1 is a major subtype of muscarinic acetylcholine receptors on mouse colonic epithelial cells. J. Gastroenterol. 2013, 48, 885–896. [Google Scholar] [CrossRef]
- Haberberger, R.; Schultheiss, G.; Diener, M. Epithelial muscarinic M1 receptors contribute to carbachol-induced ion secretion in mouse colon. Eur. J. Pharmacol. 2006, 530, 229–233. [Google Scholar] [CrossRef]
- Lundgren, O.; Jodal, M.; Jansson, M.; Ryberg, A.T.; Svensson, L. Intestinal epithelial stem/progenitor cells are controlled by mucosal afferent nerves. PLoS ONE 2011, 6, e16295. [Google Scholar] [CrossRef] [PubMed]
- Tsukimi, Y.; Pustovit, R.V.; Harrington, A.M.; Garcia-Caraballo, S.; Brierley, S.M.; Di Natale, M.; Molero, J.C.; Furness, J.B. Effects and sites of action of a M1 receptor positive allosteric modulator on colonic motility in rats and dogs compared with 5-HT(4) agonism and cholinesterase inhibition. Neurogastroenterol. Motil. 2020, 32, e13866. [Google Scholar] [CrossRef] [PubMed]
- Harrington, A.M.; Peck, C.J.; Liu, L.; Burcher, E.; Hutson, J.M.; Southwell, B.R. Localization of muscarinic receptors M1R, M2R and M3R in the human colon. Neurogastroenterol. Motil. 2010, 22, e262–e263. [Google Scholar] [CrossRef]
- Khan, M.R.; Uwada, J.; Yazawa, T.; Islam, M.T.; Krug, S.M.; Fromm, M.; Karaki, S.; Suzuki, Y.; Kuwahara, A.; Yoshiki, H.; et al. Activation of muscarinic cholinoceptor ameliorates tumor necrosis factor-alpha-induced barrier dysfunction in intestinal epithelial cells. FEBS Lett. 2015, 589, 3640–3647. [Google Scholar] [CrossRef]
- Khan, R.I.; Yazawa, T.; Anisuzzaman, A.S.; Semba, S.; Ma, Y.; Uwada, J.; Hayashi, H.; Suzuki, Y.; Ikeuchi, H.; Uchino, M.; et al. Activation of focal adhesion kinase via M1 muscarinic acetylcholine receptor is required in restitution of intestinal barrier function after epithelial injury. Biochim. Biophys. Acta 2014, 1842, 635–645. [Google Scholar] [CrossRef]
- Onali, P.; Adem, A.; Karlsson, E.; Olianas, M.C. The pharmacological action of MT-7. Life Sci. 2005, 76, 1547–1552. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, K.E.; Frizzell, R.A.; Sekar, M.C. Activation of T84 cell chloride channels by carbachol involves a phosphoinositide-coupled muscarinic M3 receptor. Eur. J. Pharmacol. 1992, 225, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Hirota, C.L.; McKay, D.M. M3 muscarinic receptor-deficient mice retain bethanechol-mediated intestinal ion transport and are more sensitive to colitis. Can. J. Physiol. Pharmacol. 2006, 84, 1153–1161. [Google Scholar] [CrossRef]
- Sun, J.; Shen, X.; Li, Y.; Guo, Z.; Zhu, W.; Zuo, L.; Zhao, J.; Gu, L.; Gong, J.; Li, J. Therapeutic Potential to Modify the Mucus Barrier in Inflammatory Bowel Disease. Nutrients 2016, 8, 44. [Google Scholar] [CrossRef] [PubMed]
- Specian, R.D.; Neutra, M.R. Mechanism of rapid mucus secretion in goblet cells stimulated by acetylcholine. J. Cell Biol. 1980, 85, 626–640. [Google Scholar] [CrossRef]
- Greenwood, B.; Mantle, M. Mucin and protein release in the rabbit jejunum: Effects of bethanechol and vagal nerve stimulation. Gastroenterology 1992, 103, 496–505. [Google Scholar] [CrossRef]
- Musayeva, A.; Jiang, S.; Ruan, Y.; Zadeh, J.K.; Chronopoulos, P.; Pfeiffer, N.; Muller, W.E.G.; Ackermann, M.; Xia, N.; Li, H.; et al. Aged Mice Devoid of the M(3) Muscarinic Acetylcholine Receptor Develop Mild Dry Eye Disease. Int. J. Mol. Sci. 2021, 22, 6133. [Google Scholar] [CrossRef]
- Gustafsson, J.K.; Davis, J.E.; Rappai, T.; McDonald, K.G.; Kulkarni, D.H.; Knoop, K.A.; Hogan, S.P.; Fitzpatrick, J.A.; Lencer, W.I.; Newberry, R.D. Intestinal goblet cells sample and deliver lumenal antigens by regulated endocytic uptake and transcytosis. Elife 2021, 10, e67292. [Google Scholar] [CrossRef]
- Haber, A.L.; Biton, M.; Rogel, N.; Herbst, R.H.; Shekhar, K.; Smillie, C.; Burgin, G.; Delorey, T.M.; Howitt, M.R.; Katz, Y.; et al. A single-cell survey of the small intestinal epithelium. Nature 2017, 551, 333–339. [Google Scholar] [CrossRef]
- Phillips, T.E.; Phillips, T.H.; Neutra, M.R. Regulation of intestinal goblet cell secretion. III. Isolated intestinal epithelium. Am. J. Physiol. 1984, 247, G674-81. [Google Scholar] [CrossRef]
- Epple, H.J.; Kreusel, K.M.; Hanski, C.; Schulzke, J.D.; Riecken, E.O.; Fromm, M. Differential stimulation of intestinal mucin secretion by cholera toxin and carbachol. Pflugers Arch. 1997, 433, 638–647. [Google Scholar] [CrossRef] [PubMed]
- McLean, L.P.; Smith, A.; Cheung, L.; Sun, R.; Grinchuk, V.; Vanuytsel, T.; Desai, N.; Urban, J.F., Jr.; Zhao, A.; Raufman, J.P.; et al. Type 3 Muscarinic Receptors Contribute to Clearance of Citrobacter rodentium. Inflamm. Bowel Dis. 2015, 21, 1860–1871. [Google Scholar] [CrossRef] [PubMed]
- Gubatan, J.; Holman, D.R.; Puntasecca, C.J.; Polevoi, D.; Rubin, S.J.; Rogalla, S. Antimicrobial peptides and the gut microbiome in inflammatory bowel disease. World J. Gastroenterol. 2021, 27, 7402–7422. [Google Scholar] [CrossRef] [PubMed]
- Satoh, Y. Atropine inhibits the degranulation of Paneth cells in ex-germ-free mice. Cell Tissue Res. 1988, 253, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.D.; Lloyd, K.C.; Walsh, J.H.; Lehrer, R.I. Secretion of type II phospholipase A2 and cryptdin by rat small intestinal Paneth cells. Infect. Immun. 1996, 64, 5161–5165. [Google Scholar] [CrossRef] [PubMed]
- Satoh, Y.; Ishikawa, K.; Oomori, Y.; Takeda, S.; Ono, K. Bethanechol and a G-protein activator, NaF/AlCl3, induce secretory response in Paneth cells of mouse intestine. Cell Tissue Res. 1992, 269, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Satoh, Y.; Habara, Y.; Ono, K.; Kanno, T. Carbamylcholine- and catecholamine-induced intracellular calcium dynamics of epithelial cells in mouse ileal crypts. Gastroenterology 1995, 108, 1345–1356. [Google Scholar] [CrossRef]
- Labed, S.A.; Wani, K.A.; Jagadeesan, S.; Hakkim, A.; Najibi, M.; Irazoqui, J.E. Intestinal Epithelial Wnt Signaling Mediates Acetylcholine-Triggered Host Defense against Infection. Immunity 2018, 48, 963–978.e3. [Google Scholar] [CrossRef]
- Dionne, S.; Hiscott, J.; D’Agata, I.; Duhaime, A.; Seidman, E.G. Quantitative PCR analysis of TNF-alpha and IL-1 beta mRNA levels in pediatric IBD mucosal biopsies. Dig. Dis. Sci. 1997, 42, 1557–1566. [Google Scholar] [CrossRef]
- Matsuda, R.; Koide, T.; Tokoro, C.; Yamamoto, T.; Godai, T.; Morohashi, T.; Fujita, Y.; Takahashi, D.; Kawana, I.; Suzuki, S.; et al. Quantitive cytokine mRNA expression profiles in the colonic mucosa of patients with steroid naive ulcerative colitis during active and quiescent disease. Inflamm. Bowel Dis. 2009, 15, 328–334. [Google Scholar] [CrossRef]
- Verma, R.; Verma, N.; Paul, J. Expression of inflammatory genes in the colon of ulcerative colitis patients varies with activity both at the mRNA and protein level. Eur. Cytokine Netw. 2013, 24, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Amasheh, M.; Grotjohann, I.; Amasheh, S.; Fromm, A.; Soderholm, J.D.; Zeitz, M.; Fromm, M.; Schulzke, J.D. Regulation of mucosal structure and barrier function in rat colon exposed to tumor necrosis factor alpha and interferon gamma in vitro: A novel model for studying the pathomechanisms of inflammatory bowel disease cytokines. Scand. J. Gastroenterol. 2009, 44, 1226–1235. [Google Scholar] [CrossRef] [PubMed]
- McKay, D.M.; Singh, P.K. Superantigen activation of immune cells evokes epithelial (T84) transport and barrier abnormalities via IFN-gamma and TNF alpha: Inhibition of increased permeability, but not diminished secretory responses by TGF-beta2. J. Immunol. 1997, 159, 2382–2390. [Google Scholar] [CrossRef]
- Billmeier, U.; Dieterich, W.; Neurath, M.F.; Atreya, R. Molecular mechanism of action of anti-tumor necrosis factor antibodies in inflammatory bowel diseases. World J. Gastroenterol. 2016, 22, 9300–9313. [Google Scholar] [CrossRef] [PubMed]
- Al-Sadi, R.; Guo, S.; Ye, D.; Rawat, M.; Ma, T.Y. TNF-alpha Modulation of Intestinal Tight Junction Permeability Is Mediated by NIK/IKK-alpha Axis Activation of the Canonical NF-kappaB Pathway. Am. J. Pathol. 2016, 186, 1151–1165. [Google Scholar] [CrossRef]
- Vereecke, L.; Sze, M.; Mc Guire, C.; Rogiers, B.; Chu, Y.; Schmidt-Supprian, M.; Pasparakis, M.; Beyaert, R.; van Loo, G. Enterocyte-specific A20 deficiency sensitizes to tumor necrosis factor-induced toxicity and experimental colitis. J. Exp. Med. 2010, 207, 1513–1523. [Google Scholar] [CrossRef]
- Vlantis, K.; Wullaert, A.; Sasaki, Y.; Schmidt-Supprian, M.; Rajewsky, K.; Roskams, T.; Pasparakis, M. Constitutive IKK2 activation in intestinal epithelial cells induces intestinal tumors in mice. J. Clin. Investig. 2011, 121, 2781–2793. [Google Scholar] [CrossRef] [PubMed]
- Krug, S.M.; Fromm, M.; Gunzel, D. Two-path impedance spectroscopy for measuring paracellular and transcellular epithelial resistance. Biophys. J. 2009, 97, 2202–2211. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Al-Lamki, R.S.; Zhang, H.; Kirkiles-Smith, N.; Gaeta, M.L.; Thiru, S.; Pober, J.S.; Bradley, J.R. Histamine antagonizes tumor necrosis factor (TNF) signaling by stimulating TNF receptor shedding from the cell surface and Golgi storage pool. J. Biol. Chem. 2003, 278, 21751–21760. [Google Scholar] [CrossRef]
- Van Zee, K.J.; Kohno, T.; Fischer, E.; Rock, C.S.; Moldawer, L.L.; Lowry, S.F. Tumor necrosis factor soluble receptors circulate during experimental and clinical inflammation and can protect against excessive tumor necrosis factor alpha in vitro and in vivo. Proc. Natl. Acad. Sci. USA 1992, 89, 4845–4849. [Google Scholar] [CrossRef]
- Peschon, J.J.; Slack, J.L.; Reddy, P.; Stocking, K.L.; Sunnarborg, S.W.; Lee, D.C.; Russell, W.E.; Castner, B.J.; Johnson, R.S.; Fitzner, J.N.; et al. An essential role for ectodomain shedding in mammalian development. Science 1998, 282, 1281–1284. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Derynck, R. Direct activation of TACE-mediated ectodomain shedding by p38 MAP kinase regulates EGF receptor-dependent cell proliferation. Mol. Cell 2010, 37, 551–566. [Google Scholar] [CrossRef] [PubMed]
- Uwada, J.; Yazawa, T.; Nakazawa, H.; Mikami, D.; Krug, S.M.; Fromm, M.; Sada, K.; Muramatsu, I.; Taniguchi, T. Store-operated calcium entry (SOCE) contributes to phosphorylation of p38 MAPK and suppression of TNF-alpha signalling in the intestinal epithelial cells. Cell. Signal. 2019, 63, 109358. [Google Scholar] [CrossRef]
- Uwada, J.; Yazawa, T.; Islam, M.T.; Khan, M.R.I.; Krug, S.M.; Fromm, M.; Karaki, S.I.; Suzuki, Y.; Kuwahara, A.; Yoshiki, H.; et al. Activation of muscarinic receptors prevents TNF-alpha-mediated intestinal epithelial barrier disruption through p38 MAPK. Cell. Signal. 2017, 35, 188–196. [Google Scholar] [CrossRef]
- McCole, D.F.; Keely, S.J.; Coffey, R.J.; Barrett, K.E. Transactivation of the epidermal growth factor receptor in colonic epithelial cells by carbachol requires extracellular release of transforming growth factor-alpha. J. Biol. Chem. 2002, 277, 42603–42612. [Google Scholar] [CrossRef] [PubMed]
- Chalaris, A.; Adam, N.; Sina, C.; Rosenstiel, P.; Lehmann-Koch, J.; Schirmacher, P.; Hartmann, D.; Cichy, J.; Gavrilova, O.; Schreiber, S.; et al. Critical role of the disintegrin metalloprotease ADAM17 for intestinal inflammation and regeneration in mice. J. Exp. Med. 2010, 207, 1617–1624. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, S.; Hiemstra, I.H.; Verseijden, C.; Hilbers, F.W.; Te Velde, A.A.; Willemsen, L.E.; Stap, J.; den Haan, J.M.; de Jonge, W.J. Cholinergic receptor activation on epithelia protects against cytokine-induced barrier dysfunction. Acta Physiol. 2015, 213, 846–859. [Google Scholar] [CrossRef]
- Ma, T.Y.; Hoa, N.T.; Tran, D.D.; Bui, V.; Pedram, A.; Mills, S.; Merryfield, M. Cytochalasin B modulation of Caco-2 tight junction barrier: Role of myosin light chain kinase. Am. J. Physiol. Gastrointest Liver Physiol. 2000, 279, G875–G885. [Google Scholar] [CrossRef]
- He, W.Q.; Wang, J.; Sheng, J.Y.; Zha, J.M.; Graham, W.V.; Turner, J.R. Contributions of Myosin Light Chain Kinase to Regulation of Epithelial Paracellular Permeability and Mucosal Homeostasis. Int. J. Mol. Sci. 2020, 21, 993. [Google Scholar] [CrossRef]
- Chassaing, B.; Aitken, J.D.; Malleshappa, M.; Vijay-Kumar, M. Dextran sulfate sodium (DSS)-induced colitis in mice. Curr. Protoc. Immunol. 2014, 104, 15.25.1–15.25.14. [Google Scholar] [CrossRef]
- Lenzen, H.; Qian, J.; Manns, M.P.; Seidler, U.; Jorns, A. Restoration of mucosal integrity and epithelial transport function by concomitant anti-TNFalpha treatment in chronic DSS-induced colitis. J. Mol. Med. 2018, 96, 831–843. [Google Scholar] [CrossRef]
- Magalhaes, D.A.; Batista, J.A.; Sousa, S.G.; Ferreira, J.D.S.; da Rocha Rodrigues, L.; Pereira, C.M.C.; do Nascimento Lima, J.V.; de Albuquerque, I.F.; Bezerra, N.; Monteiro, C.; et al. McN-A-343, a muscarinic agonist, reduces inflammation and oxidative stress in an experimental model of ulcerative colitis. Life Sci. 2021, 272, 119194. [Google Scholar] [CrossRef] [PubMed]
- Mitchelson, F.J. The pharmacology of McN-A-343. Pharmacol. Ther. 2012, 135, 216–245. [Google Scholar] [CrossRef] [PubMed]
- Hosic, S.; Lake, W.; Stas, E.; Koppes, R.; Breault, D.T.; Murthy, S.K.; Koppes, A.N. Cholinergic Activation of Primary Human Derived Intestinal Epithelium Does Not Ameliorate TNF-alpha Induced Injury. Cell Mol. Bioeng. 2020, 13, 487–505. [Google Scholar] [CrossRef] [PubMed]
- Lesko, S.; Wessler, I.; Gabel, G.; Petto, C.; Pfannkuche, H. Cholinergic modulation of epithelial integrity in the proximal colon of pigs. Cells Tissues Organs 2013, 197, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Semba, S.; Khan, R.I.; Bochimoto, H.; Watanabe, T.; Fujiya, M.; Kohgo, Y.; Liu, Y.; Taniguchi, T. Focal adhesion kinase regulates intestinal epithelial barrier function via redistribution of tight junction. Biochim. Biophys. Acta 2013, 1832, 151–159. [Google Scholar] [CrossRef]
- Howe, K.L.; Reardon, C.; Wang, A.; Nazli, A.; McKay, D.M. Transforming growth factor-beta regulation of epithelial tight junction proteins enhances barrier function and blocks enterohemorrhagic Escherichia coli O157:H7-induced increased permeability. Am. J. Pathol. 2005, 167, 1587–1597. [Google Scholar] [CrossRef] [PubMed]
- Ewaschuk, J.B.; Diaz, H.; Meddings, L.; Diederichs, B.; Dmytrash, A.; Backer, J.; Looijer-van Langen, M.; Madsen, K.L. Secreted bioactive factors from Bifidobacterium infantis enhance epithelial cell barrier function. Am. J. Physiol. Gastrointest Liver Physiol. 2008, 295, G1025–G1034. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Quach, A.; Das, S.; Barrett, K.E. Potentiation of calcium-activated chloride secretion and barrier dysfunction may underlie EGF receptor tyrosine kinase inhibitor-induced diarrhea. Physiol. Rep. 2020, 8, e14490. [Google Scholar] [CrossRef] [PubMed]
- McLean, L.P.; Smith, A.; Cheung, L.; Urban, J.F., Jr.; Sun, R.; Grinchuk, V.; Desai, N.; Zhao, A.; Raufman, J.P.; Shea-Donohue, T. Type 3 muscarinic receptors contribute to intestinal mucosal homeostasis and clearance of Nippostrongylus brasiliensis through induction of TH2 cytokines. Am. J. Physiol. Gastrointest Liver Physiol. 2016, 311, G130–G141. [Google Scholar] [CrossRef]
- Gross, E.R.; Gershon, M.D.; Margolis, K.G.; Gertsberg, Z.V.; Li, Z.; Cowles, R.A. Neuronal serotonin regulates growth of the intestinal mucosa in mice. Gastroenterology 2012, 143, 408–417.e2. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Shiraishi, A.; Murata, J.; Matsubara, S.; Nakaoka, S.; Kirimoto, S.; Osawa, M. Muscarinic receptor M3 contributes to intestinal stem cell maintenance via EphB/ephrin-B signaling. Life Sci. Alliance 2021, 4, e202000962. [Google Scholar] [CrossRef]
- Greig, C.J.; Cowles, R.A. Muscarinic acetylcholine receptors participate in small intestinal mucosal homeostasis. J. Pediatr. Surg. 2017, 52, 1031–1034. [Google Scholar] [CrossRef] [PubMed]
- Mashima, H.; Watanabe, N.; Sekine, M.; Matsumoto, S.; Asano, T.; Yuhashi, K.; Sagihara, N.; Urayoshi, S.; Uehara, T.; Fujiwara, J.; et al. The role of Galpha(q)/Galpha(11) signaling in intestinal epithelial cells. Biochem. Biophys. Rep. 2018, 13, 93–98. [Google Scholar] [PubMed]
- Watanabe, N.; Mashima, H.; Miura, K.; Goto, T.; Yoshida, M.; Goto, A.; Ohnishi, H. Requirement of Galpha(q)/Galpha(11) Signaling in the Preservation of Mouse Intestinal Epithelial Homeostasis. Cell Mol. Gastroenterol. Hepatol. 2016, 2, 767–782.e6. [Google Scholar] [PubMed]
- Bijlsma, P.B.; Kiliaan, A.J.; Scholten, G.; Heyman, M.; Groot, J.A.; Taminiau, J.A. Carbachol, but not forskolin, increases mucosal-to-serosal transport of intact protein in rat ileum in vitro. Am. J. Physiol. 1996, 271, G147-55. [Google Scholar] [CrossRef]
- Albanese, C.T.; Cardona, M.; Smith, S.D.; Watkins, S.; Kurkchubasche, A.G.; Ulman, I.; Simmons, R.L.; Rowe, M.I. Role of intestinal mucus in transepithelial passage of bacteria across the intact ileum in vitro. Surgery 1994, 116, 76–82. [Google Scholar]
- Cameron, H.L.; Perdue, M.H. Muscarinic acetylcholine receptor activation increases transcellular transport of macromolecules across mouse and human intestinal epithelium in vitro. Neurogastroenterol. Motil. 2007, 19, 47–56. [Google Scholar]
- Wallon, C.; Persborn, M.; Jonsson, M.; Wang, A.; Phan, V.; Lampinen, M.; Vicario, M.; Santos, J.; Sherman, P.M.; Carlson, M.; et al. Eosinophils express muscarinic receptors and corticotropin-releasing factor to disrupt the mucosal barrier in ulcerative colitis. Gastroenterology 2011, 140, 1597–1607. [Google Scholar]
- Keita, A.V.; Soderholm, J.D.; Ericson, A.C. Stress-induced barrier disruption of rat follicle-associated epithelium involves corticotropin-releasing hormone, acetylcholine, substance P. and mast cells. Neurogastroenterol. Motil. 2010, 22, 770–778, e221-2. [Google Scholar] [CrossRef]
- Gareau, M.G.; Jury, J.; Perdue, M.H. Neonatal maternal separation of rat pups results in abnormal cholinergic regulation of epithelial permeability. Am. J. Physiol. Gastrointest Liver Physiol. 2007, 293, G198-203. [Google Scholar] [CrossRef]
- Wang, H.; Yu, M.; Ochani, M.; Amella, C.A.; Tanovic, M.; Susarla, S.; Li, J.H.; Wang, H.; Yang, H.; Ulloa, L.; et al. Nicotinic acetylcholine receptor alpha7 subunit is an essential regulator of inflammation. Nature 2003, 421, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Beckmann, J.; Lips, K.S. The non-neuronal cholinergic system in health and disease. Pharmacology 2013, 92, 286–302. [Google Scholar] [CrossRef]
- Schutz, B.; Jurastow, I.; Bader, S.; Ringer, C.; von Engelhardt, J.; Chubanov, V.; Gudermann, T.; Diener, M.; Kummer, W.; Krasteva-Christ, G.; et al. Chemical coding and chemosensory properties of cholinergic brush cells in the mouse gastrointestinal and biliary tract. Front. Physiol. 2015, 6, 87. [Google Scholar]
- Bader, S.; Klein, J.; Diener, M. Choline acetyltransferase and organic cation transporters are responsible for synthesis and propionate-induced release of acetylcholine in colon epithelium. Eur. J. Pharmacol. 2014, 733, 23–33. [Google Scholar] [CrossRef]
- Peltekova, V.D.; Wintle, R.F.; Rubin, L.A.; Amos, C.I.; Huang, Q.; Gu, X.; Newman, B.; Van Oene, M.; Cescon, D.; Greenberg, G.; et al. Functional variants of OCTN cation transporter genes are associated with Crohn disease. Nat. Genet. 2004, 36, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Pochini, L.; Scalise, M.; Galluccio, M.; Pani, G.; Siminovitch, K.A.; Indiveri, C. The human OCTN1 (SLC22A4) reconstituted in liposomes catalyzes acetylcholine transport which is defective in the mutant L503F associated to the Crohn’s disease. Biochim. Biophys. Acta 2012, 1818, 559–565. [Google Scholar] [CrossRef]
- Song, P.; Rekow, S.S.; Singleton, C.A.; Sekhon, H.S.; Dissen, G.A.; Zhou, M.; Campling, B.; Lindstrom, J.; Spindel, E.R. Choline transporter-like protein 4 (CTL4) links to non-neuronal acetylcholine synthesis. J. Neurochem. 2013, 126, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Yajima, M.; Kimura, S.; Karaki, S.; Nio-Kobayashi, J.; Tsuruta, T.; Kuwahara, A.; Yajima, T.; Iwanaga, T. Non-neuronal, but atropine-sensitive ileal contractile responses to short-chain fatty acids: Age-dependent desensitization and restoration under inflammatory conditions in mice. Physiol. Rep. 2016, 4, e12759. [Google Scholar] [CrossRef]
- Ramirez, V.T.; Godinez, D.R.; Brust-Mascher, I.; Nonnecke, E.B.; Castillo, P.A.; Gardner, M.B.; Tu, D.; Sladek, J.A.; Miller, E.N.; Lebrilla, C.B.; et al. T-cell derived acetylcholine aids host defenses during enteric bacterial infection with Citrobacter rodentium. PLoS Pathog. 2019, 15, e1007719. [Google Scholar] [CrossRef]
- Dhawan, S.; De Palma, G.; Willemze, R.A.; Hilbers, F.W.; Verseijden, C.; Luyer, M.D.; Nuding, S.; Wehkamp, J.; Souwer, Y.; de Jong, E.C.; et al. Acetylcholine-producing T cells in the intestine regulate antimicrobial peptide expression and microbial diversity. Am. J. Physiol. Gastrointest Liver Physiol. 2016, 311, G920–G933. [Google Scholar] [CrossRef] [PubMed]
- Willemze, R.A.; Brinkman, D.J.; Welting, O.; van Hamersveld, P.H.P.; Verseijden, C.; Luyer, M.D.; Wildenberg, M.E.; Seppen, J.; de Jonge, W.J. Acetylcholine-producing T cells augment innate immune-driven colitis but are redundant in T cell-driven colitis. Am. J. Physiol. Gastrointest Liver Physiol. 2019, 317, G557–G568. [Google Scholar] [CrossRef]
- Belo, A.; Cheng, K.; Chahdi, A.; Shant, J.; Xie, G.; Khurana, S.; Raufman, J.P. Muscarinic receptor agonists stimulate human colon cancer cell migration and invasion. Am. J. Physiol. Gastrointest Liver Physiol. 2011, 300, G749–G760. [Google Scholar] [CrossRef]
- Cheng, K.; Shang, A.C.; Drachenberg, C.B.; Zhan, M.; Raufman, J.P. Differential expression of M3 muscarinic receptors in progressive colon neoplasia and metastasis. Oncotarget 2017, 8, 21106–21114. [Google Scholar] [CrossRef] [PubMed]
- Dwomoh, L.; Tejeda, G.S.; Tobin, A.B. Targeting the M1 muscarinic acetylcholine receptor in Alzheimer’s disease. Neuronal Signal. 2022, 6, NS20210004. [Google Scholar] [CrossRef]
- Nguyen, H.T.M.; van der Westhuizen, E.T.; Langmead, C.J.; Tobin, A.B.; Sexton, P.M.; Christopoulos, A.; Valant, C. Opportunities and challenges for the development of M(1) muscarinic receptor positive allosteric modulators in the treatment for neurocognitive deficits. Br. J. Pharmacol. 2022, in press. [Google Scholar] [CrossRef] [PubMed]
- Kurimoto, E.; Matsuda, S.; Shimizu, Y.; Sako, Y.; Mandai, T.; Sugimoto, T.; Sakamoto, H.; Kimura, H. An Approach to Discovering Novel Muscarinic M(1) Receptor Positive Allosteric Modulators with Potent Cognitive Improvement and Minimized Gastrointestinal Dysfunction. J. Pharmacol. Exp. Ther. 2018, 364, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Matsui, M.; Motomura, D.; Fujikawa, T.; Jiang, J.; Takahashi, S.; Manabe, T.; Taketo, M.M. Mice lacking M2 and M3 muscarinic acetylcholine receptors are devoid of cholinergic smooth muscle contractions but still viable. J. Neurosci. 2002, 22, 10627–10632. [Google Scholar] [CrossRef]
- Bader, S.; Diener, M. Novel aspects of cholinergic regulation of colonic ion transport. Pharmacol. Res. Perspect. 2015, 3, e00139. [Google Scholar] [CrossRef]
- Al-Barazie, R.M.; Bashir, G.H.; Qureshi, M.M.; Mohamed, Y.A.; Al-Sbiei, A.; Tariq, S.; Lammers, W.J.; Al-Ramadi, B.K.; Fernandez-Cabezudo, M.J. Cholinergic Activation Enhances Resistance to Oral Salmonella Infection by Modulating Innate Immune Defense Mechanisms at the Intestinal Barrier. Front. Immunol. 2018, 9, 551. [Google Scholar] [CrossRef]
- Singh, S.P.; Chand, H.S.; Banerjee, S.; Agarwal, H.; Raizada, V.; Roy, S.; Sopori, M. Acetylcholinesterase Inhibitor Pyridostigmine Bromide Attenuates Gut Pathology and Bacterial Dysbiosis in a Murine Model of Ulcerative Colitis. Dig. Dis. Sci. 2020, 65, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Okumura, T.; Nozu, T.; Ishioh, M.; Igarashi, S.; Kumei, S.; Ohhira, M. Brain orexin improves intestinal barrier function via the vagal cholinergic pathway. Neurosci. Lett. 2020, 714, 134592. [Google Scholar] [CrossRef] [PubMed]
- Yajima, T.; Inoue, R.; Yajima, M.; Tsuruta, T.; Karaki, S.; Hira, T.; Kuwahara, A. The G-protein on cholesterol-rich membrane microdomains mediates mucosal sensing of short- chain fatty acid and secretory response in rat colon. Acta Physiol. 2011, 203, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Yajima, T.; Inoue, R.; Matsumoto, M.; Yajima, M. Non-neuronal release of ACh plays a key role in secretory response to luminal propionate in rat colon. J. Physiol. 2011, 589, 953–962. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Tissues | Region/Cell Types | Subtypes | Methods | Ref. | |
|---|---|---|---|---|---|
| mouse | small intestine | villi | M1, M2, M3, M4 (M5-) | qPCR | [8] |
| crypts | M1, M3, M4 (M2-, M5-) | qPCR | [8] | ||
| epithelial cells | M1, M2, M3, M4, M5 | IHC | [9] | ||
| M1, M3 (M2-, M4-, M5-) | qPCR | [10] | |||
| M1, M2, M3, M4, M5 | qPCR, IHC | [11] | |||
| goblet cells | M3, M4, (M1-, M2-, M5-) | qPCR, microarray | [12] | ||
| Paneth cells | M2 | IHC | [11] | ||
| M3 | IHC | [10] | |||
| stem cells (crypt base) | M1 (M2-, M3-, M4-, M5-) | qPCR | [13] | ||
| M1, M3 | qPCR | [8] | |||
| M3 | IHC | [10] | |||
| endocrine cells | M3 | IHC | [10] | ||
| colon | crypts | M1 (80%), M3 (20%) | pharmacological | [14] | |
| M1 | qPCR | [15] | |||
| goblet cell | M3, M4, (M1-, M2-, M5-) | qPCR, microarray | [12] | ||
| rat | small intestine | stem cell | M3, M5 | IHC | [16] |
| human | colon | epithelial cells | M1 | ISH | [17] |
| M3 (M1-) | IHC | [18] | |||
| cell line (colon) | HT-29/B6 cells | M3 (M1-) | pharmacological, qPCR | [19] | |
| cell line (colon) | T84 cells | M3 (65%), M1 (35%) | pharmacological | [20] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uwada, J.; Nakazawa, H.; Muramatsu, I.; Masuoka, T.; Yazawa, T. Role of Muscarinic Acetylcholine Receptors in Intestinal Epithelial Homeostasis: Insights for the Treatment of Inflammatory Bowel Disease. Int. J. Mol. Sci. 2023, 24, 6508. https://doi.org/10.3390/ijms24076508
Uwada J, Nakazawa H, Muramatsu I, Masuoka T, Yazawa T. Role of Muscarinic Acetylcholine Receptors in Intestinal Epithelial Homeostasis: Insights for the Treatment of Inflammatory Bowel Disease. International Journal of Molecular Sciences. 2023; 24(7):6508. https://doi.org/10.3390/ijms24076508
Chicago/Turabian StyleUwada, Junsuke, Hitomi Nakazawa, Ikunobu Muramatsu, Takayoshi Masuoka, and Takashi Yazawa. 2023. "Role of Muscarinic Acetylcholine Receptors in Intestinal Epithelial Homeostasis: Insights for the Treatment of Inflammatory Bowel Disease" International Journal of Molecular Sciences 24, no. 7: 6508. https://doi.org/10.3390/ijms24076508