
Progressive Motor and Non-Motor Symptoms in Park7 Knockout Zebrafish
 ,
,
Abstract
:1. Introduction
2. Results and Discussion
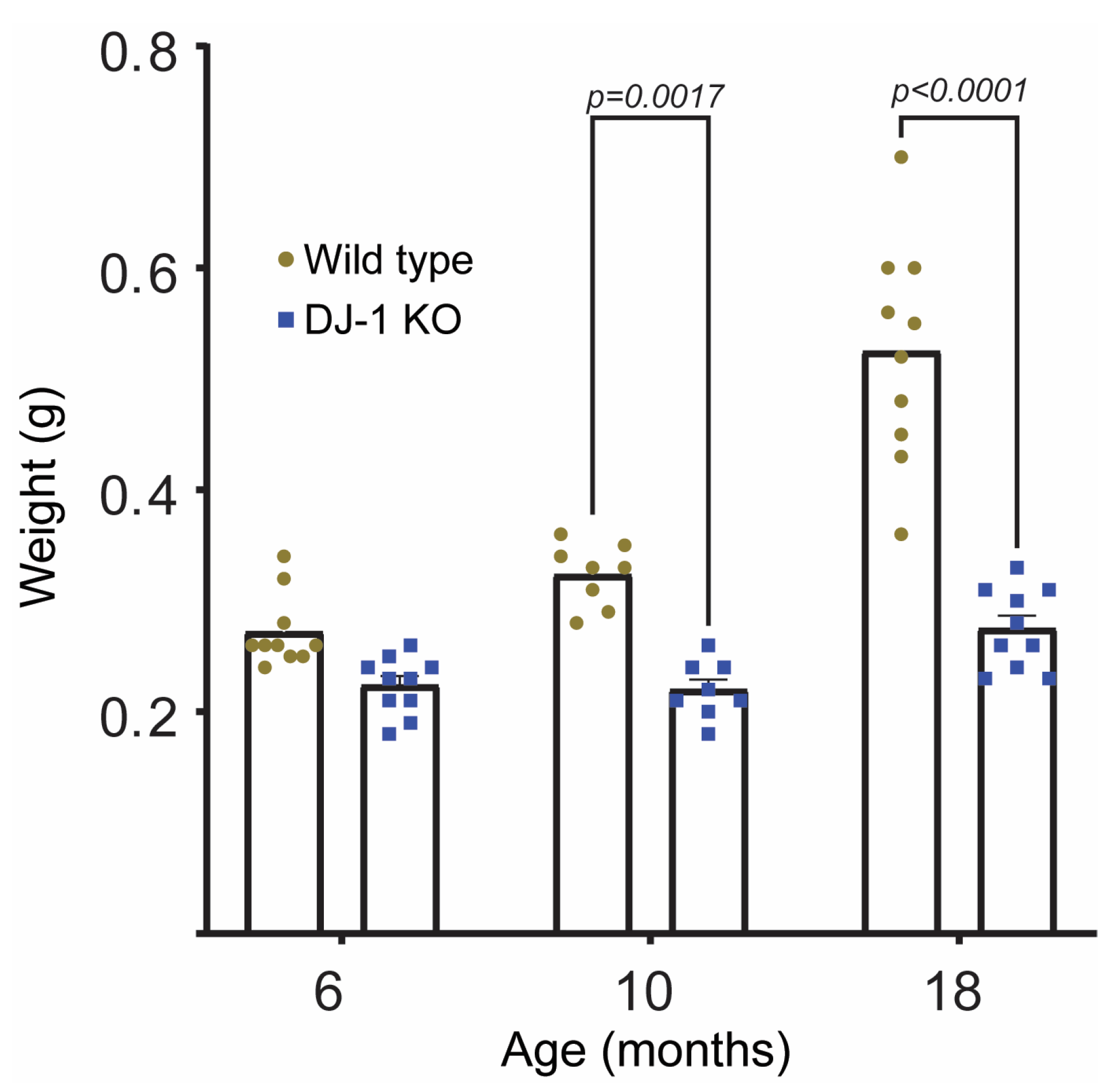
2.1. DJ-1 Knockouts Do Not Gain Weight after 6 Months of Age
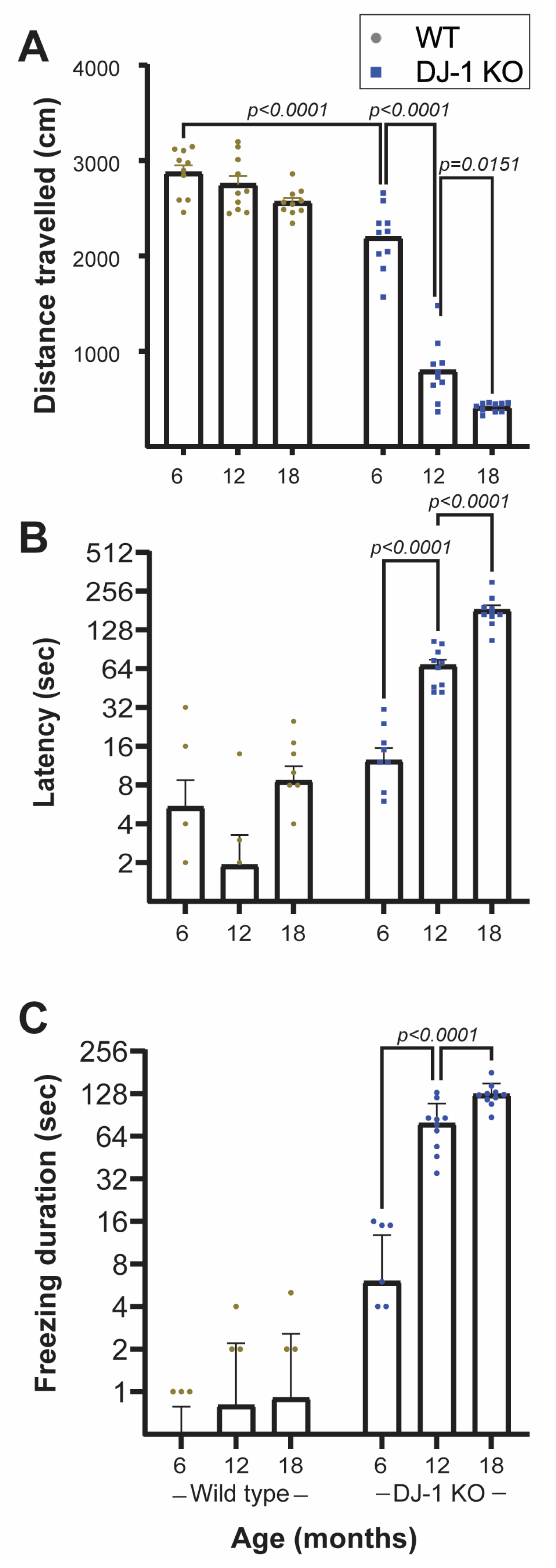
2.2. Loss of DJ-1 Induces Age-Dependent Progressive Motor and Anxiety Associated Behavior Deficits
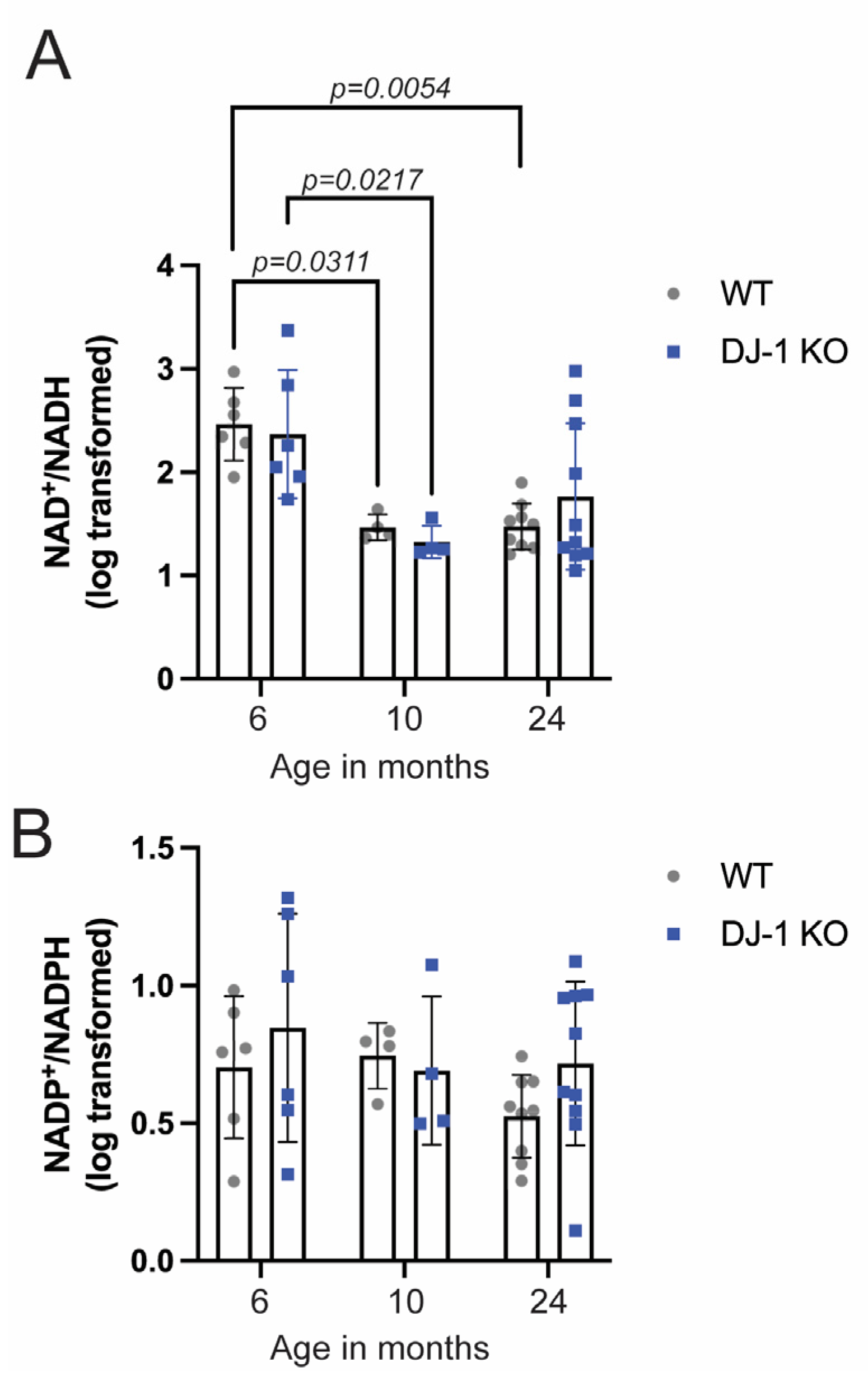
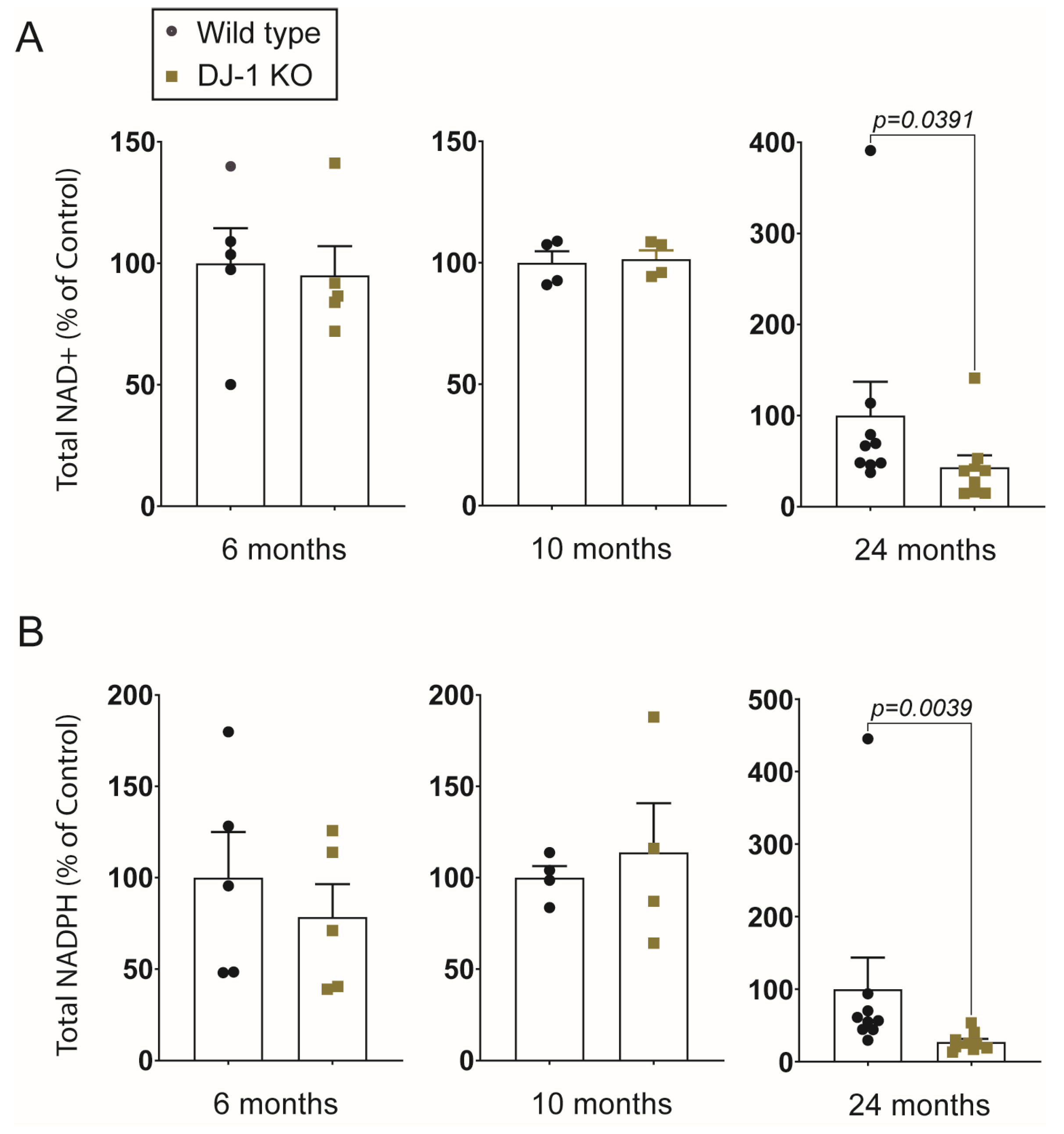
2.3. NAD+/NADH Ratio Declines in Aging Zebrafish, but DJ-1 Knockout Only Affects NAD+ Levels at the Late Adult Stage
2.4. Altered Behavior Preceeds Down-Regulation in Tyrosine Hydroxylase, but Co-Incides with Changes in the Mitochondrial Proteome
2.5. DJ-1 Deficient Zebrafish as a Model of Parkinson’s Disease
3. Material and Methods
3.1. Animal Maintenance
3.2. Zebrafish Lines
3.3. Weight Measurement
3.4. Novel Tank Test
3.5. Zebrafish Behavior Analysis
3.6. Sampling of Adult Zebrafish Brains
3.7. Sample Preparation for Metabolomics Directed Mass Spectrometry and Protein Measurement
3.8. Targeted Metabolomic Analysis
3.9. Western Blotting
3.10. Sample Preparation of Mitochondrial Lysates
3.11. Label Free Proteomics Analysis of Brain Mitochondria Fractions
3.12. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- GBD 2016 Parkinson’s Disease Collaborators. Global, regional, and national burden of Parkinson’s disease, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2018, 17, 939–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tansey, M.G.; Wallings, R.L.; Houser, M.C.; Herrick, M.K.; Keating, C.E.; Joers, V. Inflammation and immune dysfunction in Parkinson disease. Nat. Rev. Immunol. 2022, 22, 657–673. [Google Scholar] [CrossRef] [PubMed]
- Bonifati, V.; Rizzu, P.; van Baren, M.J.; Schaap, O.; Breedveld, G.J.; Krieger, E.; Dekker, M.C.; Squitieri, F.; Ibanez, P.; Joosse, M.; et al. Mutations in the DJ-1 gene associated with autosomal recessive early-onset parkinsonism. Science 2003, 299, 256–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Im, J.Y.; Lee, K.W.; Junn, E.; Mouradian, M.M. DJ-1 protects against oxidative damage by regulating the thioredoxin/ASK1 complex. Neurosci. Res. 2010, 67, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Inden, M.; Kitamura, Y.; Takahashi, K.; Takata, K.; Ito, N.; Niwa, R.; Funayama, R.; Nishimura, K.; Taniguchi, T.; Honda, T.; et al. Protection against dopaminergic neurodegeneration in Parkinson’s disease-model animals by a modulator of the oxidized form of DJ-1, a wild-type of familial Parkinson’s disease-linked PARK7. J. Pharmacol. Sci. 2011, 117, 189–203. [Google Scholar] [CrossRef] [Green Version]
- Moore, D.J.; Zhang, L.; Troncoso, J.; Lee, M.K.; Hattori, N.; Mizuno, Y.; Dawson, T.M.; Dawson, V.L. Association of DJ-1 and parkin mediated by pathogenic DJ-1 mutations and oxidative stress. Hum. Mol. Genet. 2005, 14, 71–84. [Google Scholar] [CrossRef] [Green Version]
- Saito, Y.; Miyasaka, T.; Hatsuta, H.; Takahashi-Niki, K.; Hayashi, K.; Mita, Y.; Kusano-Arai, O.; Iwanari, H.; Ariga, H.; Hamakubo, T.; et al. Immunostaining of oxidized DJ-1 in human and mouse brains. J. Neuropathol. Exp. Neurol. 2014, 73, 714–728. [Google Scholar] [CrossRef]
- Duan, X.; Kelsen, S.G.; Merali, S. Proteomic analysis of oxidative stress-responsive proteins in human pneumocytes: Insight into the regulation of DJ-1 expression. J. Proteome Res. 2008, 7, 4955–4961. [Google Scholar] [CrossRef]
- Chandran, J.S.; Lin, X.; Zapata, A.; Hoke, A.; Shimoji, M.; Moore, S.O.; Galloway, M.P.; Laird, F.M.; Wong, P.C.; Price, D.L.; et al. Progressive behavioral deficits in DJ-1-deficient mice are associated with normal nigrostriatal function. Neurobiol. Dis. 2008, 29, 505–514. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Cagniard, B.; Mathews, T.; Jones, S.; Koh, H.C.; Ding, Y.; Carvey, P.M.; Ling, Z.; Kang, U.J.; Zhuang, X. Age-dependent motor deficits and dopaminergic dysfunction in DJ-1 null mice. J. Biol. Chem. 2005, 280, 21418–21426. [Google Scholar] [CrossRef] [Green Version]
- Giangrasso, D.M.; Furlong, T.M.; Keefe, K.A. Characterization of striatum-mediated behavior and neurochemistry in the DJ-1 knock-out rat model of Parkinson’s disease. Neurobiol. Dis. 2020, 134, 104673. [Google Scholar] [CrossRef]
- Sanchez-Catasus, C.A.; Bohnen, N.I.; D’Cruz, N.; Muller, M. Striatal Acetylcholine-Dopamine Imbalance in Parkinson Disease: In Vivo Neuroimaging Study with Dual-Tracer PET and Dopaminergic PET-Informed Correlational Tractography. J. Nucl. Med. 2022, 63, 438–445. [Google Scholar] [CrossRef]
- Edson, A.J.; Hushagen, H.A.; Froyset, A.K.; Elda, I.; Khan, E.A.; Di Stefano, A.; Fladmark, K.E. Dysregulation in the Brain Protein Profile of Zebrafish Lacking the Parkinson’s Disease-Related Protein DJ-1. Mol. Neurobiol. 2019, 56, 8306–8322. [Google Scholar] [CrossRef]
- Hughes, G.L.; Lones, M.A.; Bedder, M.; Currie, P.D.; Smith, S.L.; Pownall, M.E. Machine learning discriminates a movement disorder in a zebrafish model of Parkinson’s disease. Dis. Model. Mech. 2020, 13, 045815. [Google Scholar] [CrossRef]
- De Miranda, B.R.; Rocha, E.M.; Bai, Q.; El Ayadi, A.; Hinkle, D.; Burton, E.A.; Timothy Greenamyre, J. Astrocyte-specific DJ-1 overexpression protects against rotenone-induced neurotoxicity in a rat model of Parkinson’s disease. Neurobiol. Dis. 2018, 115, 101–114. [Google Scholar] [CrossRef]
- Froyset, A.K.; Edson, A.J.; Gharbi, N.; Khan, E.A.; Dondorp, D.; Bai, Q.; Tiraboschi, E.; Suster, M.L.; Connolly, J.B.; Burton, E.A.; et al. Astroglial DJ-1 over-expression up-regulates proteins involved in redox regulation and is neuroprotective in vivo. Redox. Biol. 2018, 16, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Krebiehl, G.; Ruckerbauer, S.; Burbulla, L.F.; Kieper, N.; Maurer, B.; Waak, J.; Wolburg, H.; Gizatullina, Z.; Gellerich, F.N.; Woitalla, D.; et al. Reduced basal autophagy and impaired mitochondrial dynamics due to loss of Parkinson’s disease-associated protein DJ-1. PLoS ONE 2010, 5, e9367. [Google Scholar] [CrossRef] [Green Version]
- Abou-Sleiman, P.M.; Muqit, M.M.; Wood, N.W. Expanding insights of mitochondrial dysfunction in Parkinson’s disease. Nat. Rev. Neurosci. 2006, 7, 207–219. [Google Scholar] [CrossRef]
- Henchcliffe, C.; Beal, M.F. Mitochondrial biology and oxidative stress in Parkinson disease pathogenesis. Nat. Clin. Pract. Neurol. 2008, 4, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Ma, Y.; Kong, X.; Ding, X.; Gu, H.; Chu, T.; Ying, W. NAD(+) administration decreases doxorubicin-induced liver damage of mice by enhancing antioxidation capacity and decreasing DNA damage. Chem. Biol. Interact. 2014, 212, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Lautrup, S.; Sinclair, D.A.; Mattson, M.P.; Fang, E.F. NAD(+) in Brain Aging and Neurodegenerative Disorders. Cell Metab. 2019, 30, 630–655. [Google Scholar] [CrossRef]
- Schondorf, D.C.; Ivanyuk, D.; Baden, P.; Sanchez-Martinez, A.; De Cicco, S.; Yu, C.; Giunta, I.; Schwarz, L.K.; Di Napoli, G.; Panagiotakopoulou, V.; et al. The NAD+ Precursor Nicotinamide Riboside Rescues Mitochondrial Defects and Neuronal Loss in iPSC and Fly Models of Parkinson’s Disease. Cell Rep. 2018, 23, 2976–2988. [Google Scholar] [CrossRef]
- Schwab, A.J.; Sison, S.L.; Meade, M.R.; Broniowska, K.A.; Corbett, J.A.; Ebert, A.D. Decreased Sirtuin Deacetylase Activity in LRRK2 G2019S iPSC-Derived Dopaminergic Neurons. Stem Cell Rep. 2017, 9, 1839–1852. [Google Scholar] [CrossRef] [Green Version]
- Cummings, J.; Lee, G.; Mortsdorf, T.; Ritter, A.; Zhong, K. Alzheimer’s disease drug development pipeline: 2017. Alzheimers Dement. 2017, 3, 367–384. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Xiong, N.; Shen, Y.; Han, C.; Liu, L.; Zhang, G.; Wang, L.; Guo, S.; Guo, X.; Xia, Y.; et al. Weight Loss and Malnutrition in Patients with Parkinson’s Disease: Current Knowledge and Future Prospects. Front. Aging Neurosci. 2018, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Liu, X.M.; Sun, J.G.; Chen, H.; Ma, J.; Dong, M.; Peng, S.; Wang, J.Q.; Ding, J.Q.; Li, D.H.; et al. DJ-1 maintains energy and glucose homeostasis by regulating the function of brown adipose tissue. Cell Discov. 2017, 3, 16054. [Google Scholar] [CrossRef] [Green Version]
- Zang, L.; Maddison, L.A.; Chen, W. Zebrafish as a Model for Obesity and Diabetes. Front. Cell Dev. Biol. 2018, 6, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berardelli, A.; Rothwell, J.C.; Thompson, P.D.; Hallett, M. Pathophysiology of bradykinesia in Parkinson’s disease. Brain 2001, 124, 2131–2146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landau, S.; Harris, V.; Burn, D.J.; Hindle, J.V.; Hurt, C.S.; Samuel, M.; Wilson, K.C.; Brown, R.G. Anxiety and anxious-depression in Parkinson’s disease over a 4-year period: A latent transition analysis. Psychol. Med. 2016, 46, 657–667. [Google Scholar] [CrossRef] [Green Version]
- Lemke, M.R.; Brecht, H.M.; Koester, J.; Kraus, P.H.; Reichmann, H. Anhedonia, depression, and motor functioning in Parkinson’s disease during treatment with pramipexole. J. Neuropsychiatry Clin. Neurosci. 2005, 17, 214–220. [Google Scholar] [CrossRef]
- Wong, K.; Elegante, M.; Bartels, B.; Elkhayat, S.; Tien, D.; Roy, S.; Goodspeed, J.; Suciu, C.; Tan, J.; Grimes, C.; et al. Analyzing habituation responses to novelty in zebrafish (Danio rerio). Behav. Brain Res. 2010, 208, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Cachat, J.; Stewart, A.; Grossman, L.; Gaikwad, S.; Kadri, F.; Chung, K.M.; Wu, N.; Wong, K.; Roy, S.; Suciu, C.; et al. Measuring behavioral and endocrine responses to novelty stress in adult zebrafish. Nat. Protoc. 2010, 5, 1786–1799. [Google Scholar] [CrossRef] [PubMed]
- Campos, F.L.; Carvalho, M.M.; Cristovao, A.C.; Je, G.; Baltazar, G.; Salgado, A.J.; Kim, Y.S.; Sousa, N. Rodent models of Parkinson’s disease: Beyond the motor symptomatology. Front. Behav. Neurosci. 2013, 7, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Covarrubias, A.J.; Perrone, R.; Grozio, A.; Verdin, E. NAD(+) metabolism and its roles in cellular processes during ageing. Nat. Rev. Mol. Cell Biol. 2021, 22, 119–141. [Google Scholar] [CrossRef] [PubMed]
- Massudi, H.; Grant, R.; Braidy, N.; Guest, J.; Farnsworth, B.; Guillemin, G.J. Age-associated changes in oxidative stress and NAD+ metabolism in human tissue. PLoS ONE 2012, 7, e42357. [Google Scholar] [CrossRef] [PubMed]
- Caito, S.W.; Aschner, M. NAD+ Supplementation Attenuates Methylmercury Dopaminergic and Mitochondrial Toxicity in Caenorhabditis Elegans. Toxicol. Sci. 2016, 151, 139–149. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Li, X.; Gao, H.; Feng, Z.; Li, X.; Zhao, L.; Jia, X.; Zhang, H.; Liu, J. High doses of nicotinamide prevent oxidative mitochondrial dysfunction in a cellular model and improve motor deficit in a Drosophila model of Parkinson’s disease. J. Neurosci. Res. 2008, 86, 2083–2090. [Google Scholar] [CrossRef]
- Lehmann, S.; Loh, S.H.; Martins, L.M. Enhancing NAD(+) salvage metabolism is neuroprotective in a PINK1 model of Parkinson’s disease. Biol. Open 2017, 6, 141–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brakedal, B.; Dolle, C.; Riemer, F.; Ma, Y.; Nido, G.S.; Skeie, G.O.; Craven, A.R.; Schwarzlmuller, T.; Brekke, N.; Diab, J.; et al. The NADPARK study: A randomized phase I trial of nicotinamide riboside supplementation in Parkinson’s disease. Cell Metab. 2022, 34, 396–407.e6. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.E.; Paek, S.H. Mitochondrial Dysfunction in Parkinson’s Disease. Exp. Neurobiol. 2015, 24, 103–116. [Google Scholar] [CrossRef] [Green Version]
- Sai, Y.; Zou, Z.; Peng, K.; Dong, Z. The Parkinson’s disease-related genes act in mitochondrial homeostasis. Neurosci. Biobehav. Rev. 2012, 36, 2034–2043. [Google Scholar] [CrossRef] [PubMed]
- Blacher, E.; Dadali, T.; Bespalko, A.; Haupenthal, V.J.; Grimm, M.O.; Hartmann, T.; Lund, F.E.; Stein, R.; Levy, A. Alzheimer’s disease pathology is attenuated in a CD38-deficient mouse model. Ann. Neurol. 2015, 78, 88–103. [Google Scholar] [CrossRef] [PubMed]
- Stein, L.R.; Imai, S. Specific ablation of Nampt in adult neural stem cells recapitulates their functional defects during aging. EMBO J. 2014, 33, 1321–1340. [Google Scholar] [CrossRef] [Green Version]
- Bradshaw, P.C. Cytoplasmic and Mitochondrial NADPH-Coupled Redox Systems in the Regulation of Aging. Nutrients 2019, 11, 504. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Bartolome, A.; Penas, A.; Illescas, M.; Bermejo, V.; Lopez-Calcerrada, S.; Perez-Perez, R.; Marin-Buera, L.; Dominguez-Gonzalez, C.; Arenas, J.; Martin, M.A.; et al. Altered Expression Ratio of Actin-Binding Gelsolin Isoforms Is a Novel Hallmark of Mitochondrial OXPHOS Dysfunction. Cells 2020, 9, 1922. [Google Scholar] [CrossRef] [PubMed]
- Kotiadis, V.N.; Leadsham, J.E.; Bastow, E.L.; Gheeraert, A.; Whybrew, J.M.; Bard, M.; Lappalainen, P.; Gourlay, C.W. Identification of new surfaces of cofilin that link mitochondrial function to the control of multi-drug resistance. J. Cell Sci. 2012, 125, 2288–2299. [Google Scholar] [CrossRef] [Green Version]
- Flinkman, D.; Hong, Y.; Gnjatovic, J.; Deshpande, P.; Ortutay, Z.; Peltonen, S.; Kaasinen, V.; James, P.; Coffey, E. Regulators of proteostasis are translationally repressed in fibroblasts from patients with sporadic and LRRK2-G2019S Parkinson’s disease. NPJ Park. Dis. 2023, 9, 20. [Google Scholar] [CrossRef]
- Fang, J.D.; Tung, H.H.; Lee, S.L. Mitochondrial localization of St14-encoding transmembrane serine protease is involved in neural stem/progenitor cell bioenergetics through binding to F(0)F(1)-ATP synthase complex. FASEB J. 2019, 33, 4327–4340. [Google Scholar] [CrossRef] [PubMed]
- Shulyakova, N.; Andreazza, A.C.; Mills, L.R.; Eubanks, J.H. Mitochondrial Dysfunction in the Pathogenesis of Rett Syndrome: Implications for Mitochondria-Targeted Therapies. Front. Cell Neurosci. 2017, 11, 58. [Google Scholar] [CrossRef] [Green Version]
- Higa, A.; Taouji, S.; Lhomond, S.; Jensen, D.; Fernandez-Zapico, M.E.; Simpson, J.C.; Pasquet, J.M.; Schekman, R.; Chevet, E. Endoplasmic reticulum stress-activated transcription factor ATF6alpha requires the disulfide isomerase PDIA5 to modulate chemoresistance. Mol. Cell Biol. 2014, 34, 1839–1849. [Google Scholar] [CrossRef] [Green Version]
- Farfariello, V.; Gordienko, D.V.; Mesilmany, L.; Touil, Y.; Germain, E.; Fliniaux, I.; Desruelles, E.; Gkika, D.; Roudbaraki, M.; Shapovalov, G.; et al. TRPC3 shapes the ER-mitochondria Ca(2+) transfer characterizing tumour-promoting senescence. Nat. Commun. 2022, 13, 956. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Vashisht, A.A.; Tchieu, J.; Wohlschlegel, J.A.; Dreier, L. Voltage-dependent anion channels (VDACs) recruit Parkin to defective mitochondria to promote mitochondrial autophagy. J. Biol. Chem. 2012, 287, 40652–40660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Keeffe, G.W.; Hegarty, S.V.; Sullivan, A.M. Targeting bone morphogenetic protein signalling in midbrain dopaminergic neurons as a therapeutic approach in Parkinson’s disease. Neuronal Signal. 2017, 1, NS20170027. [Google Scholar] [CrossRef] [Green Version]
- Urra, F.A.; Fuentes-Retamal, S.; Palominos, C.; Rodriguez-Lucart, Y.A.; Lopez-Torres, C.; Araya-Maturana, R. Extracellular Matrix Signals as Drivers of Mitochondrial Bioenergetics and Metabolic Plasticity of Cancer Cells During Metastasis. Front. Cell Dev. Biol. 2021, 9, 751301. [Google Scholar] [CrossRef] [PubMed]
- Pantner, Y.; Polavarapu, R.; Chin, L.S.; Li, L.; Shimizu, Y.; Calvert, J.W. DJ-1 attenuates the glycation of mitochondrial complex I and complex III in the post-ischemic heart. Sci. Rep. 2021, 11, 19408. [Google Scholar] [CrossRef]
- Yang, J.; Kim, K.S.; Iyirhiaro, G.O.; Marcogliese, P.C.; Callaghan, S.M.; Qu, D.; Kim, W.J.; Slack, R.S.; Park, D.S. DJ-1 modulates the unfolded protein response and cell death via upregulation of ATF4 following ER stress. Cell Death Dis. 2019, 10, 135. [Google Scholar] [CrossRef] [Green Version]
- Moscovitz, O.; Ben-Nissan, G.; Fainer, I.; Pollack, D.; Mizrachi, L.; Sharon, M. The Parkinson’s-associated protein DJ-1 regulates the 20S proteasome. Nat. Commun. 2015, 6, 6609. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Ma, X.; Fujioka, H.; Liu, J.; Chen, S.; Zhu, X. DJ-1 regulates the integrity and function of ER-mitochondria association through interaction with IP3R3-Grp75-VDAC1. Proc. Natl. Acad. Sci. USA 2019, 116, 25322–25328. [Google Scholar] [CrossRef]
- Cagalinec, M.; Liiv, M.; Hodurova, Z.; Hickey, M.A.; Vaarmann, A.; Mandel, M.; Zeb, A.; Choubey, V.; Kuum, M.; Safiulina, D.; et al. Role of Mitochondrial Dynamics in Neuronal Development: Mechanism for Wolfram Syndrome. PLoS Biol. 2016, 14, e1002511. [Google Scholar] [CrossRef] [Green Version]
- Koks, S.; Soomets, U.; Paya-Cano, J.L.; Fernandes, C.; Luuk, H.; Plaas, M.; Terasmaa, A.; Tillmann, V.; Noormets, K.; Vasar, E.; et al. Wfs1 gene deletion causes growth retardation in mice and interferes with the growth hormone pathway. Physiol. Genom. 2009, 37, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Luuk, H.; Plaas, M.; Raud, S.; Innos, J.; Sutt, S.; Lasner, H.; Abramov, U.; Kurrikoff, K.; Koks, S.; Vasar, E. Wfs1-deficient mice display impaired behavioural adaptation in stressful environment. Behav. Brain. Res. 2009, 198, 334–345. [Google Scholar] [CrossRef] [PubMed]
- Gharbi, N.; Roise, D.; Forre, J.E.; Edson, A.J.; Hushagen, H.A.; Tronci, V.; Froyset, A.K.; Fladmark, K.E. Reintroduction of DJ-1 in Muller Cells Inhibits Retinal Degeneration in the DJ-1 Deficient Retina. Antioxidants 2021, 10, 1862. [Google Scholar] [CrossRef] [PubMed]
- Imberechts, D.; Kinnart, I.; Wauters, F.; Terbeek, J.; Manders, L.; Wierda, K.; Eggermont, K.; Madeiro, R.F.; Sue, C.; Verfaillie, C.; et al. DJ-1 is an essential downstream mediator in PINK1/parkin-dependent mitophagy. Brain 2022, 145, 4368–4384. [Google Scholar] [CrossRef] [PubMed]
- Thomas, K.J.; McCoy, M.K.; Blackinton, J.; Beilina, A.; van der Brug, M.; Sandebring, A.; Miller, D.; Maric, D.; Cedazo-Minguez, A.; Cookson, M.R. DJ-1 acts in parallel to the PINK1/parkin pathway to control mitochondrial function and autophagy. Hum. Mol. Genet. 2011, 20, 40–50. [Google Scholar] [CrossRef] [Green Version]
- Abou-Sleiman, P.M.; Healy, D.G.; Quinn, N.; Lees, A.J.; Wood, N.W. The role of pathogenic DJ-1 mutations in Parkinson’s disease. Ann. Neurol. 2003, 54, 283–286. [Google Scholar] [CrossRef]
- Choi, J.; Sullards, M.C.; Olzmann, J.A.; Rees, H.D.; Weintraub, S.T.; Bostwick, D.E.; Gearing, M.; Levey, A.I.; Chin, L.S.; Li, L. Oxidative damage of DJ-1 is linked to sporadic Parkinson and Alzheimer diseases. J. Biol. Chem. 2006, 281, 10816–10824. [Google Scholar] [CrossRef] [Green Version]
- Egan, R.J.; Bergner, C.L.; Hart, P.C.; Cachat, J.M.; Canavello, P.R.; Elegante, M.F.; Elkhayat, S.I.; Bartels, B.K.; Tien, A.K.; Tien, D.H.; et al. Understanding behavioral and physiological phenotypes of stress and anxiety in zebrafish. Behav. Brain Res. 2009, 205, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Stewart, A.; Gaikwad, S.; Kyzar, E.; Green, J.; Roth, A.; Kalueff, A.V. Modeling anxiety using adult zebrafish: A conceptual review. Neuropharmacology 2012, 62, 135–143. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UniProt Acc a | Protein Names | Gene Name | p-Value | F.C b | KO c | WT d | Protein Description | |
|---|---|---|---|---|---|---|---|---|
| Transport and membrane trafficking | ||||||||
| A9JRA5 | Gelsolin | scinla | 0.0073 | 4.62 | 22.69 | 20.50 | Linked to OXPHOS defect [45]. | |
| Q6NZW3 | Cofilin 2 | cfl2 | 0.0482 | 4.47 | 20.54 | 17.87 | Mitochondrial homeostasis [46]. | |
| Q66HW2 | EH-domain containing 1 | ehd1b | 0.0419 | 0.45 | 19.90 | 21.11 | Downregulated in PD [47]. | |
| X1WC49 | Anoctamin | ano3 | 0.0204 | 0.32 | 19.99 | 20.94 | ||
| Synapse signaling | ||||||||
| A2CJ03 | Dystrobrevin | dtna | 0.0003 | 5.20 | 19.47 | 17.23 | ||
| A5WWH0 | Chromogranin B | chgb | 0.0078 | 3.74 | 21.13 | 19.05 | ||
| Metabolism | ||||||||
| Q1RLP8 | Suppressor of tumorigenicity 14 protein homolog | st14a | 0.0285 | 8.89 | 20.45 | 16.89 | Mitochondrial localization stress response [48]. | |
| Q6P962 | Glutathione transferase | gsta.1 | 0.0081 | 3.70 | 24.74 | 22.62 | Oxidative stress regulation | |
| Q66I52 | NADH dehydrogenase iron-sulfur protein 6 | ndufs6 | n.a. | n.a. | NaN | 25.20 | Complex I assembly | |
| A0A0R4IQ88 | D-glutamate cyclase | dglucy | 0.0012 | 0.27 | 22.68 | 24.50 | Metabolization od D-glutamate | |
| Q7ZU10 | Aldehyde dehydrogenase | aldh3b1 | 0.0135 | 0.47 | 17.71 | 18.61 | ||
| B3DJF3 | 3′-phosphoadenosine-5′-phosphosulfate synthase | papss1 | 0.0004 | 0.49 | 20.44 | 21.22 | ||
| Chaperone activity | ||||||||
| A0A2R8RNM0 | Mitochondrial import inner membrane translocase | timm8a | 0.0249 | 3.37 | 20.11 | 18.19 | ||
| Mitochondrial transcriptional and translational regulation | Oxidative stress regulation [49]. | |||||||
| Q7T2T7 | Methyl-CpG-binding protein 2 | mecp2 | 0.0417 | 6.71 | 21.22 | 18.47 | Ass. to neurologic developmental disorder | |
| Q502J9 | 39S ribosomal protein L30, mitochondrial | mrpl30 | 0.0266 | 0.14 | 19.09 | 21.94 | ||
| E9QEK4 | Mitochondrial ribosomal protein L45 | mrpl45 | 0.0158 | 0.36 | 21.05 | 22.11 | ||
| A2BEV1 | Mitochondrial translational initiation factor 3 | mtif3 | 0.0020 | 0.36 | 19.51 | 21.04 | ||
| Stress response regulation and unfolded protein response | Required for ER stress respons. | |||||||
| A8WG75 | Pdia5 protein | pdia5 | 0.0019 | 0.05 | 20.91 | 24.84 | ER stress regulation [50] | |
| Q7ZUW0 | DnaJ (Hsp40) homolog, subfamily A, member 3A | dnaja3a | 0.0089 | 0.47 | 21.19 | 21.72 | Reg. of mitochondrial apoptosis signaling | |
| U3N8Z0 | Transient receptor potential cation channel memb. 3 | trpc3 | 0.0071 | 0.48 | 18.53 | 19.44 | Mitochondrial Ca2+ control [51] | |
| Ubiquitin-Proteasome Dependent Proteolysis | ||||||||
| E7EZD6 | Ubiquitinyl hydrolase 1 | usp4 | 0.0165 | 0.21 | 20.09 | 21.63 | ||
| Q9PTH5 | Proteasome activator subunit 1 | psme1 | 0.0368 | 0.22 | 20.86 | 21.78 | ||
| Mitochondrial quality control | Linked to PD [52]. | |||||||
| B0R198 | Voltage-dependent anion-selective channel protein 3 | vdac3 | 0.0433 | 0.39 | 21.37 | 22.45 | ||
| A0A0R4IXA9 | Adenylyl cyclase-associated protein | cap2 | 0.0099 | 0.43 | 20.30 | 21.44 | ||
| Neuroinflammation | Neuroprotective. Linked to early loss of axons in PD models [53]. | |||||||
| O73736 | Receptor protein serine/threonine kinase | acvr1l | n.a. | n.a. | NaN | 21.29 | Regulation of mitochondrial function [54]. | |
| Q3YA99 | Integrin beta | itgb1b.1 | n.a. | n.a. | NaN | 18.90 | ||
| Other | ||||||||
| F1QYR3 | Uncharacterized protein | LOC100004199 | 0.0425 | 0.28 | 18.62 | 20.35 | ||
| A0A286YA42 | Si:ch73-366l1.5 | si:ch73-366l1.5 | 0.0174 | 0.19 | 20.67 | 22.83 | ||
| A5PMM7 | Si:dkey-193c22.1 | si:dkey-193c22.1 | 0.0355 | 0.38 | 17.95 | 19.25 | ||
| Compound | Accurate Mass | Formula [M + H]+ | Inclusion List | rt |
|---|---|---|---|---|
| NAD+ | 664.11695 | C21H28N7O14P2 | 665.0 ± 8 | 11.02 |
| NADH | 666.13205 | C21H30N7O14P2 | 665.0 ± 8 | 5.57 |
| NADP+ | 744.08273 | C21H29N7O17P3 | 745.0 ± 8 | 10.54 |
| NADPH | 746.09783 | C21H31N7O17P3 | 745.0 ± 8 | 10.57 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chavali, L.N.M.; Yddal, I.; Bifulco, E.; Mannsåker, S.; Røise, D.; Law, J.O.; Frøyset, A.-K.; Grellscheid, S.N.; Fladmark, K.E. Progressive Motor and Non-Motor Symptoms in Park7 Knockout Zebrafish. Int. J. Mol. Sci. 2023, 24, 6456. https://doi.org/10.3390/ijms24076456
Chavali LNM, Yddal I, Bifulco E, Mannsåker S, Røise D, Law JO, Frøyset A-K, Grellscheid SN, Fladmark KE. Progressive Motor and Non-Motor Symptoms in Park7 Knockout Zebrafish. International Journal of Molecular Sciences. 2023; 24(7):6456. https://doi.org/10.3390/ijms24076456
Chicago/Turabian StyleChavali, Lakshmi Narasimha Murthy, Ingeborg Yddal, Ersilia Bifulco, Simen Mannsåker, Dagne Røise, Jack O. Law, Ann-Kristin Frøyset, Sushma N. Grellscheid, and Kari E. Fladmark. 2023. "Progressive Motor and Non-Motor Symptoms in Park7 Knockout Zebrafish" International Journal of Molecular Sciences 24, no. 7: 6456. https://doi.org/10.3390/ijms24076456