
Effects of Curcumin on Oxidative Stress and Ferroptosis in Acute Ammonia Stress-Induced Liver Injury in Gibel Carp (Carassius gibelio)
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Growth Performance and Body Composition
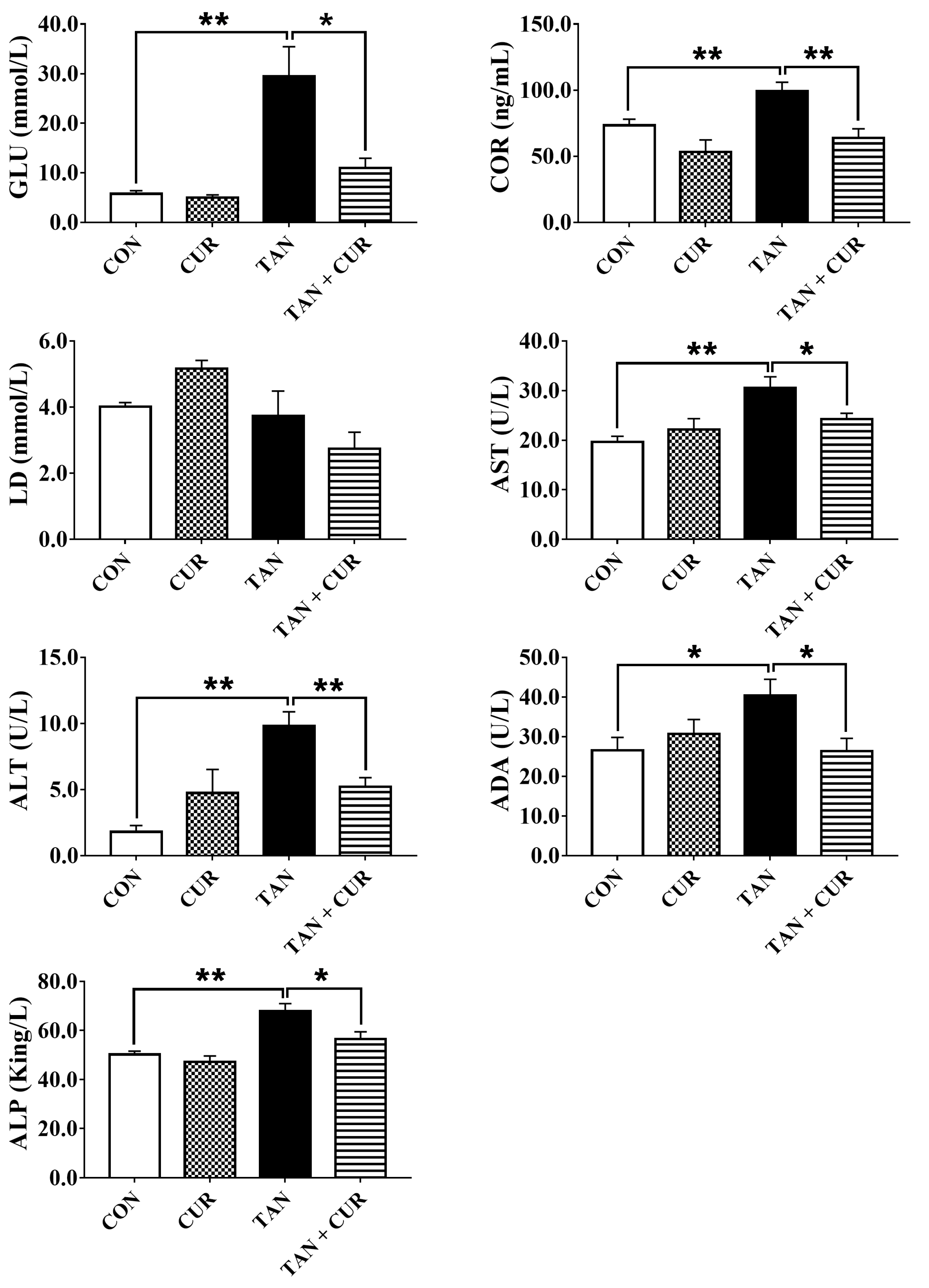
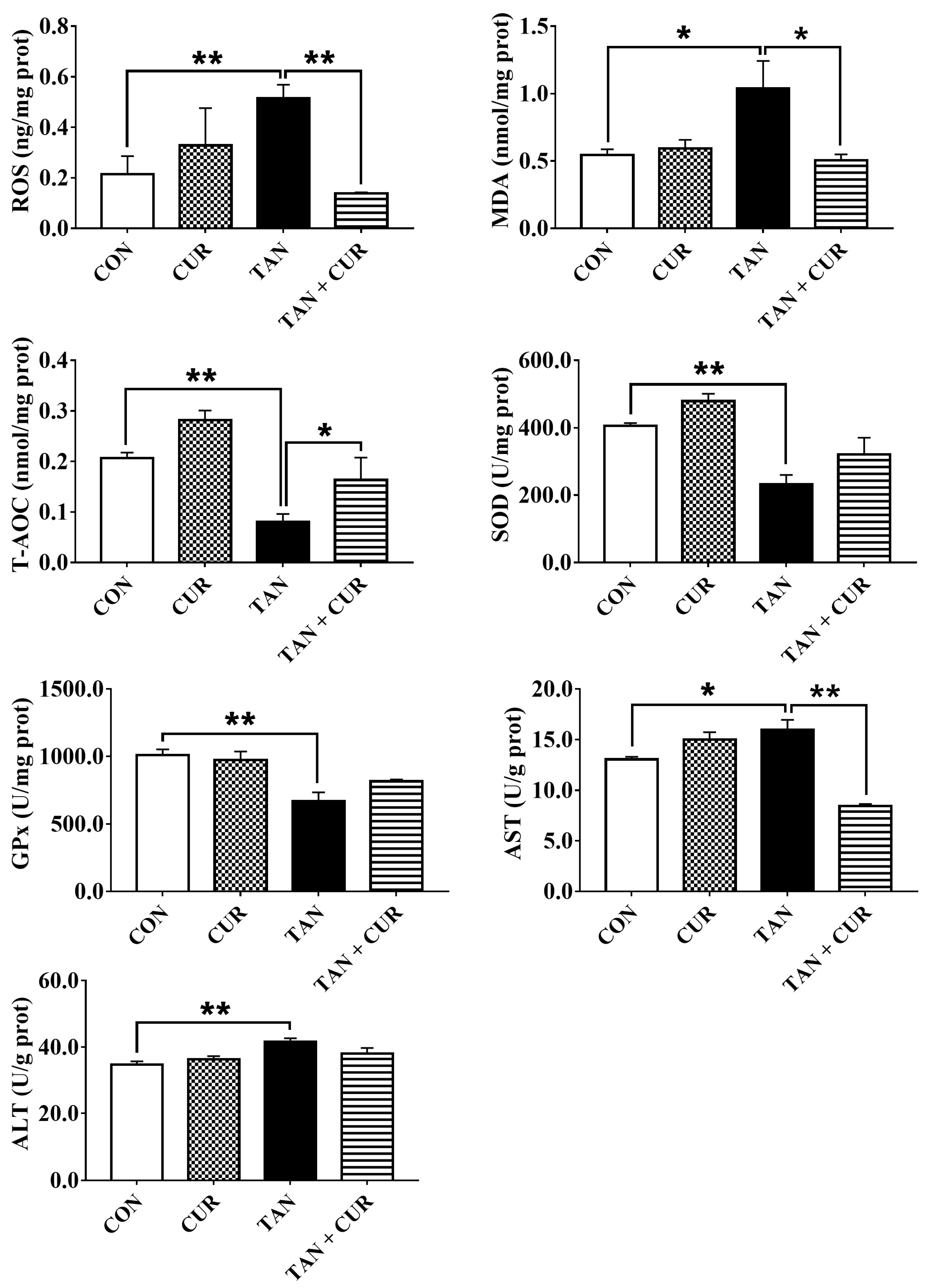
2.2. Plasma and Liver Metabolites
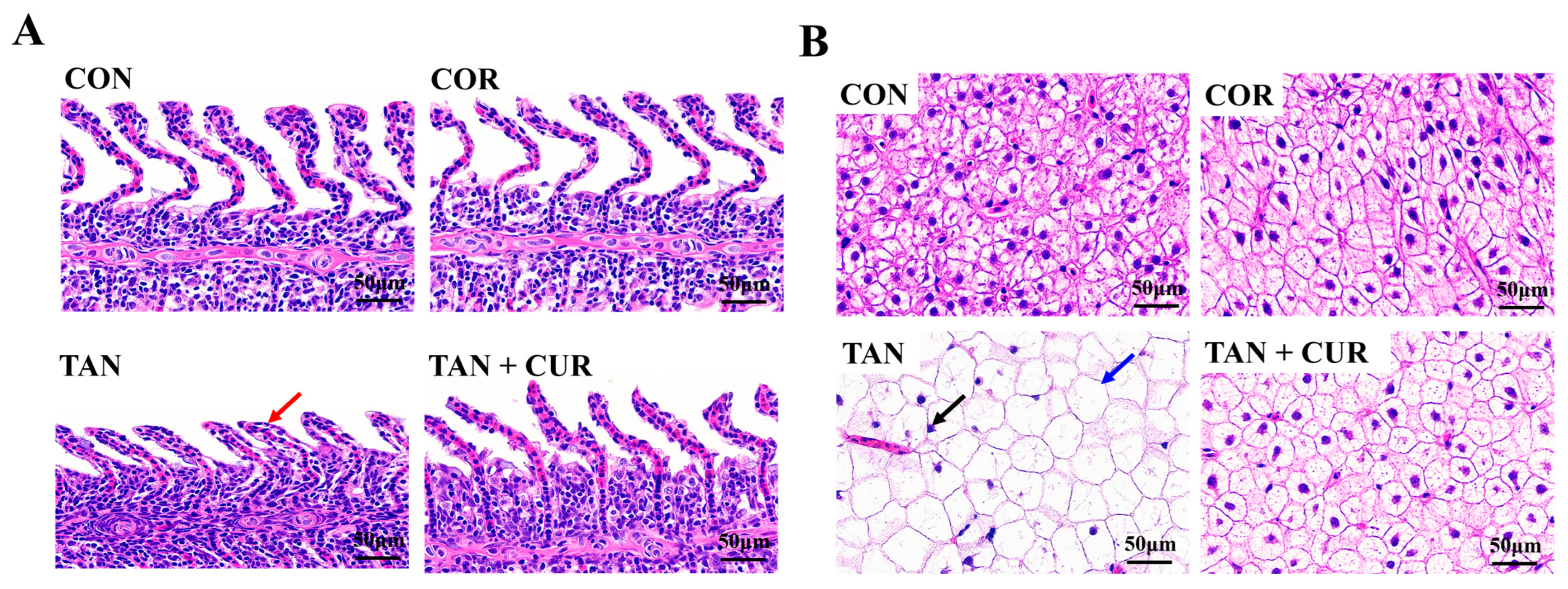
2.3. Tissue Histopathology
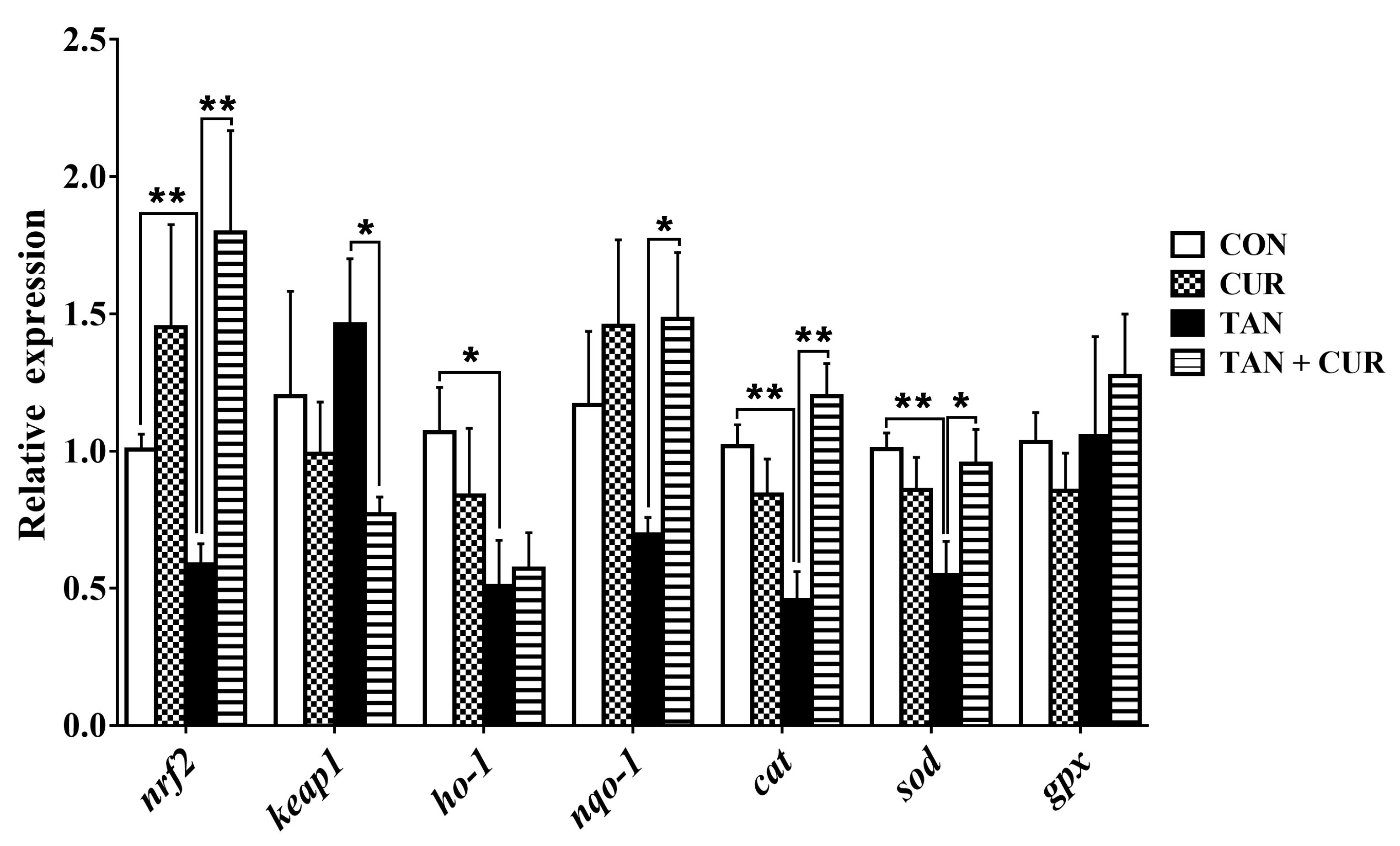
2.4. Nuclear Factor [Erythroid-Derived 2]-like 2 (Nrf2) Signaling Pathway
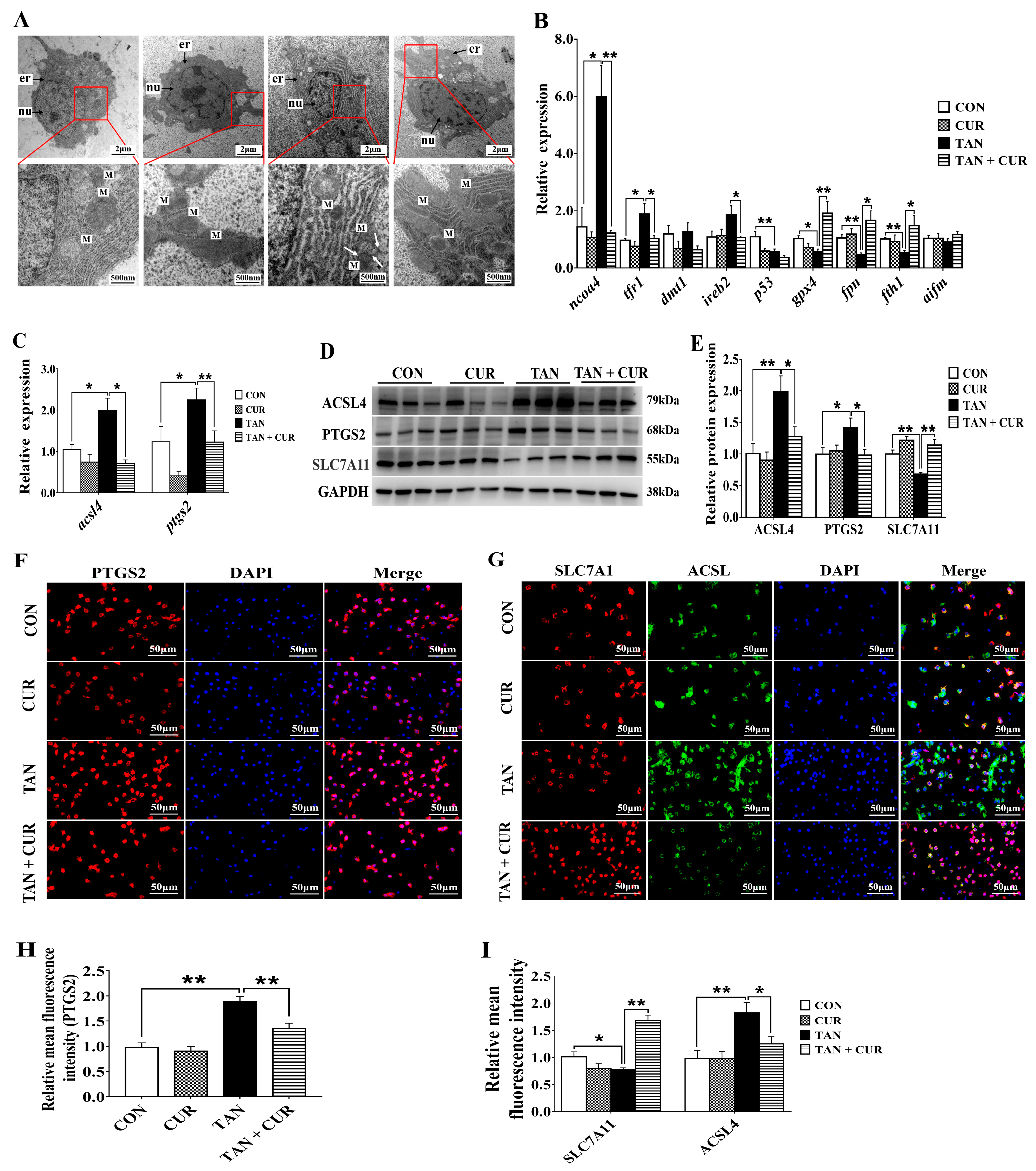
2.5. Ferroptosis
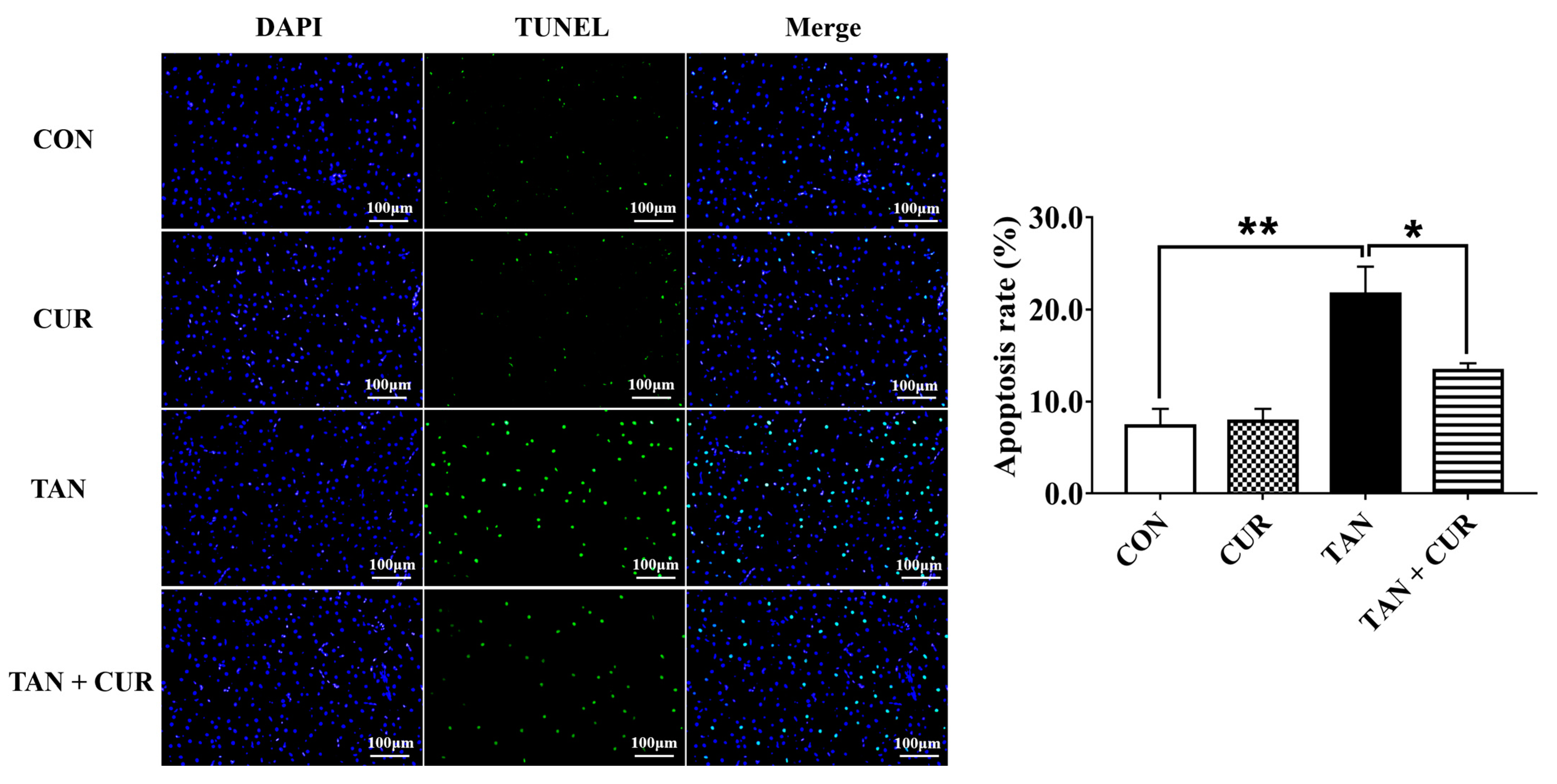
2.6. Terminal Deoxynucleotidyl Transferase dUTP Nick-End Labeling (TUNEL) Analysis
3. Discussion
4. Materials and Methods
4.1. Experimental Diets
4.2. Experimental Animals and Procedures
4.3. Ammonia Stress Experiments
4.4. Sample Collection
4.5. Biochemical Analysis
4.6. Histopathology
4.7. TUNEL Assay
4.8. TEM
4.9. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.10. Western Blot Analysis
4.11. Immunofluorescence Analysis
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheng, C.H.; Yang, F.F.; Ling, R.Z.; Liao, S.A.; Miao, Y.T.; Ye, C.X.; Wang, A.L. Effects of ammonia exposure on apoptosis, oxidative stress and immune response in pufferfish (Takifugu obscurus). Aquat. Toxicol. 2015, 164, 61–71. [Google Scholar] [CrossRef]
- Xu, Z.K.; Cao, J.; Qin, X.M.; Qiu, W.Q.; Mei, J.; Xie, J. Toxic effects on bioaccumulation, hematological parameters, oxidative stress, immune responses and tissue structure in fish exposed to ammonia nitrogen: A review. Animals 2021, 11, 3304. [Google Scholar] [CrossRef]
- Luo, Z.; Zou, G.Y.; Gao, Y.; Ye, H.M.; Xi, W.Q.; Liu, X. Effect of dietary iron (Fe) levels on growth performance, hepatic lipid metabolism and antioxidant responses in juvenile yellow catfish Pelteobagrus fulvidraco. Aquacult. Nutr. 2017, 23, 1475–1482. [Google Scholar] [CrossRef]
- Habte-Tsion, H.M.; Liu, B.; Ge, X.P.; Xie, J.; Xu, P.; Ren, M.C.; Zhou, Q.L.; Pan, L.K.; Chen, R.L. Effects of dietary protein level on growth performance, muscle composition, blood composition, and digestive enzyme activity of wuchang bream (Megalobrama amblycephala) fry. Isr. J. Aquacult-Bamid. 2013, 65, 9. [Google Scholar] [CrossRef]
- Hoseini, S.M.; Yousefi, M.; Hoseinifar, S.H.; Van Doan, H. Antioxidant, enzymatic and hematological responses of common carp (Cyprinus carpio) fed with myrcene- or menthol-supplemented diets and exposed to ambient ammonia. Aquaculture 2019, 506, 246–255. [Google Scholar] [CrossRef]
- Rojo de la Vega, M.; Chapman, E.; Zhang, D.D. Nrf2 and the hallmarks of cancer. Cancer Cell 2018, 34, 21–43. [Google Scholar] [CrossRef]
- Kong, Y.D.; Li, M.; Guo, G.L.; Yu, L.H.; Sun, L.; Yin, Z.; Li, R.M.; Chen, X.M.; Wang, G.Q. Effects of dietary curcumin inhibit deltamethrin-induced oxidative stress, inflammation and cell apoptosis in Channa argus via Nrf2 and NF-κB signaling pathways. Aquaculture 2021, 540, 736744. [Google Scholar] [CrossRef]
- Yang, W.S.; Stockwell, B.R. Ferroptosis: Death by lipid peroxidation. Trends Cell Biol. 2016, 26, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Gao, M.; Jiang, X. To eat or not to eat-the metabolic flavor of ferroptosis. Curr. Opin. Cell Biol. 2018, 51, 58–64. [Google Scholar] [CrossRef]
- Li, Q.; Han, X.N.; Lan, X.; Gao, Y.F.; Wan, J.R.; Durham, F.; Cheng, T.; Yang, J.; Wang, Z.Y.; Jiang, C.; et al. Inhibition of neuronal ferroptosis protects hemorrhagic brain. JCI Insight 2017, 2, e90777. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; An, P.; Xie, E.J.; Wu, Q.; Fang, X.X.; Gao, H.; Zhang, Z.Z.; Li, Y.Z.; Wang, X.D.; Zhang, J.Y.; et al. Characterization of ferroptosis in murine models of hemochromatosis. Hepatology 2017, 66, 449–465. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Li, J.B.; Kang, R.; Klionsky, D.J.; Tang, D.L. Ferroptosis: Machinery and regulation. Autophagy 2021, 17, 2054–2081. [Google Scholar] [CrossRef]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.S.; SriRamaratnam, R.; Welsch, M.E.; Shimada, K.; Skouta, R.; Viswanathan, V.S.; Cheah, J.H.; Clemons, P.A.; Shamji, A.F.; Clish, C.B.; et al. Regulation of ferroptotic cancer cell death by GPX4. Cell 2014, 156, 317–331. [Google Scholar] [CrossRef] [Green Version]
- Angeli, J.P.F.; Shah, R.; Pratt, D.A.; Conrad, M. Ferroptosis inhibition: Mechanisms and opportunities. Trends Pharmacol. Sci. 2017, 38, 489–498. [Google Scholar] [CrossRef]
- Alagawany, M.; Farag, M.R.; Abdelnour, S.A.; Dawood, M.A.O.; Elnesr, S.S.; Dhama, K. Curcumin and its different forms: A review on fish nutrition. Aquaculture 2021, 532, 736030. [Google Scholar] [CrossRef]
- Wang, Y.J.; Liu, F.J.; Liu, M.R.; Zhou, X.; Wang, M.; Cao, K.X.; Jin, S.J.; Shan, A.S.; Feng, X.J. Curcumin mitigates aflatoxin B1-induced liver injury via regulating the NLRP3 inflammasome and Nrf2 signaling pathway. Food Chem. Toxicol. 2022, 161, 112823. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations. The State of World Fisheries and Aquaculture; FAO Fisheries and Aquaculture Department: Rome, Italy, 2022. [Google Scholar]
- Qi, X.Z.; Xue, M.Y.; Yang, S.B.; Zha, J.W.; Wang, G.X.; Ling, F. Ammonia exposure alters the expression of immune-related and antioxidant enzymes-related genes and the gut microbial community of crucian carp (Carassius auratus). Fish Shellfish Immunol. 2017, 70, 485–492. [Google Scholar] [CrossRef]
- Jiang, J.; Wu, X.Y.; Zhou, X.Q.; Feng, L.; Liu, Y.; Jiang, W.D.; Wu, P.; Zhao, Y. Effects of dietary curcumin supplementation on growth performance, intestinal digestive enzyme activities and antioxidant capacity of crucian carp Carassius auratus. Aquaculture 2016, 463, 174–180. [Google Scholar] [CrossRef]
- Ming, J.; Ye, J.; Zhang, Y.; Xu, Q.; Yang, X.; Shao, X.; Qiang, J.; Xu, P. Optimal dietary curcumin improved growth performance, and modulated innate immunity, antioxidant capacity and related genes expression of NF-κB and Nrf2 signaling pathways in grass carp (Ctenopharyngodon idella) after infection with Aeromonas hydrophila. Fish Shellfish Immunol. 2020, 97, 540–553. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Gewaily, M.S.; Monier, M.N.; Younis, E.M.; Doan, H.V.; Sewilam, H. The regulatory roles of yucca extract on the growth rate, hepato-renal function, histopathological alterations, and immune-related genes in common carp exposed with acute ammonia stress. Aquaculture 2021, 534, 736287. [Google Scholar] [CrossRef]
- Goorden, S.M.; Buffart, T.E.; Bakker, A.; Buijs, M.M. Liver disorders in adults: ALT and AST. Ned. Tijdschr. Geneeskd. 2013, 157, A6443. [Google Scholar] [PubMed]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative Stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Kim, W.K.; Bae, K.H.; Lee, S.C.; Lee, E.W. Lipid metabolism and ferroptosis. Biology-Basel 2021, 10, 184. [Google Scholar] [CrossRef]
- Gan, L.; Yao, L.; Zhang, M.Z.; He, K.W.; Zhang, H.; Lin, Y.H.; Qin, C.J.; Shao, J.; Jiang, H.B.; Wen, M. Ammonia triggers ferroptosis in macrophages of yellow catfish Pelteobagrus fulvidraco. Aquacult. Res. 2022, 53, 568–575. [Google Scholar] [CrossRef]
- Jin, X.T.; Song, L.; Li, Z.Y.; Newton, I.P.; Zhao, M.R.; Liu, W.P. Dichlorodiphenyldichloroethylene exposure reduces r-GCS via suppressed Nrf2 in HepG2 cells. Environ. Toxicol. 2016, 31, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Huang, W.; Wang, S.; Sun, K.; Zhang, W.; Ding, Y.; Zhang, L.; Tumen, B.; Ji, L.; Liu, C. Astragaloside IV attenuates acetaminophen-induced liver injuries in mice by activating the Nrf2 signaling pathway. Molecules 2018, 23, 2023. [Google Scholar] [CrossRef] [Green Version]
- He, K.W.; Luo, X.P.; Wen, M.; Wang, C.G.; Qin, C.J.; Shao, J.; Gan, L.; Dong, R.R.; Jiang, H.B. Effect of acute ammonia toxicity on inflammation, oxidative stress and apoptosis in head kidney macrophage of Pelteobagrus fulvidraco and the alleviation of curcumin. Comp. Biochem. Phys. C 2021, 248, 109098. [Google Scholar] [CrossRef]
- Zhang, X.J.; Zhou, L.; Lu, W.J.; Du, W.X.; Mi, X.Y.; Li, Z.; Li, X.Y.; Wang, Z.W.; Wang, Y.; Duan, M.; et al. Comparative transcriptomic analysis reveals an association of gibel carp fatty liver with ferroptosis pathway. BMC Genom. 2021, 22, 328. [Google Scholar] [CrossRef]
- Cheng, C.M.; Wang, D.; Cao, X.; Luo, Q.Q.; Lu, Y.P.; Zhu, L. Iron regulatory protein 1 suppresses hypoxia-induced iron uptake proteins expression and decreases iron levels in HepG2 cells. J. Cell. Biochem. 2015, 116, 1919–1931. [Google Scholar] [CrossRef]
- Di Sanzo, M.; Quaresima, B.; Biamonte, F.; Palmieri, C.; Faniello, M.C. FTH1 pseudogenes in cancer and cell metabolism. Cells-Basel 2020, 9, 2554. [Google Scholar] [CrossRef]
- Zheng, J.S.; Conrad, M. The metabolic underpinnings of ferroptosis. Cell Metab. 2020, 32, 920–937. [Google Scholar] [CrossRef]
- Doll, S.; Proneth, B.; Tyurina, Y.Y.; Panzilius, E.; Kobayashi, S.; IngoId, I.; Irmler, M.; Beckers, J.; Aichler, M.; Walch, A.; et al. ACSL4 dictates ferroptosis sensitivity by shaping cellular lipid composition. Nat. Chem. Biol. 2017, 13, 91–98. [Google Scholar] [CrossRef]
- Jiang, L.; Kon, N.; Li, T.Y.; Wang, S.J.; Su, T.; Hibshoosh, H.; Baer, R.; Gu, W. Ferroptosis as a p53-mediated activity during tumour suppression. Nature 2015, 520, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Guerrero-Hue, M.; Garcia-Caballero, C.; Palomino-Antolin, A.; Rubio-Navarro, A.; Vazquez-Carballo, C.; Herencia, C.; Martin-Sanchez, D.; Farre-Alins, V.; Egea, J.; Cannata, P.; et al. Curcumin reduces renal damage associated with rhabdomyolysis by decreasing ferroptosis-mediated cell death. FASEB J. 2019, 33, 8961–8975. [Google Scholar] [CrossRef]
- Wu, L.Y.; Chen, Q.Z.; Dong, B.; Han, D.; Zhu, X.M.; Liu, H.K.; Yang, Y.X.; Xie, S.Q.; Jin, J.Y. Resveratrol attenuated oxidative stress and inflammatory and mitochondrial dysfunction induced by acute ammonia exposure in gibel carp (Carassius gibelio). Ecotoxicol. Environ. Saf. 2023, 251, 114544. [Google Scholar] [CrossRef]
- Cao, S.P.; Zou, T.; Zhang, P.Y.; Han, D.; Jin, J.Y.; Liu, H.K.; Yang, Y.X.; Zhu, X.M.; Xie, S.Q. Effects of dietary fishmeal replacement with Spirulina platensis on the growth, feed utilization, digestion and physiological parameters in juvenile gibel carp (Carassis auratus gibelio var. CAS III). Aquacult. Res. 2018, 49, 1320–1328. [Google Scholar] [CrossRef]
- Dong, B.; Wu, L.Y.; Chen, Q.Z.; Xu, W.J.; Li, D.G.; Han, D.; Zhu, X.M.; Liu, H.K.; Yang, Y.X.; Xie, S.Q.; et al. Tolerance assessment of Atractylodes macrocephala polysaccharide in the diet of largemouth bass (Micropterus salmoides). Antioxidants 2022, 11, 1581. [Google Scholar] [CrossRef]
- Li, H.Y.; Xu, W.J.; Wu, L.Y.; Dong, B.; Jin, J.Y.; Han, D.; Zhu, X.M.; Yang, Y.X.; Liu, H.K.; Xie, S.Q. Differential regulation of endoplasmic reticulum stress-induced autophagy and apoptosis in two strains of gibel carp (Carassius gibelio) exposed to acute waterborne cadmium. Aquat. Toxicol. 2021, 231, 105721. [Google Scholar] [CrossRef]
- Wu, L.Y.; Li, H.Y.; Xu, W.J.; Dong, B.; Geng, H.C.; Jin, J.Y.; Han, D.; Liu, H.K.; Zhu, X.M.; Yang, Y.X.; et al. Emodin alleviates acute hypoxia-induced apoptosis in gibel carp (Carassius gibelio) by upregulating autophagy through modulation of the AMPK/mTOR pathway. Aquaculture 2022, 548, 737689. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Parameter | Treatment | |
|---|---|---|---|
| CON | CUR | ||
| Growth performance | W0 1 (g) | 11.23 ± 0.07 | 11.20 ± 0.10 |
| Wt 2 (g) | 36.43 ± 2.29 | 37.63 ± 3.13 | |
| FR 3 (%) | 3.12 ± 0.05 a | 3.13 ± 0.01 b | |
| FE 4 (%) | 61.02 ± 0.97 | 60.42 ± 1.89 | |
| SGR 5 (%/d) | 2.10 ± 0.06 | 2.16 ± 0.09 | |
| Body composition | Crude protein (%) | 14.62 ± 0.22 | 14.41 ± 0.28 |
| Crude lipid (%) | 6.34 ± 0.17 | 6.13 ± 0.23 | |
| Ash (%) | 3.33 ± 0.32 a | 3.45 ± 0.33 b | |
| Moisture (%) | 72.24 ± 0.30 | 71.43 ± 0.43 | |
| Ingredients | CON | CUR |
|---|---|---|
| White fish meal 1 | 15 | 15 |
| Rapeseed meal 2 | 20 | 20 |
| Soybean meal 3 | 25 | 25 |
| Wheat flour 4 | 25.4 | 25.4 |
| Oil mixture 5 | 5.5 | 5.5 |
| CUR | 0 | 0.5 |
| Vitamin premix 6 | 0.39 | 0.39 |
| Choline chloride | 0.11 | 0.11 |
| Mineral premix 7 | 5 | 5 |
| Carboxy methyl cellulose sodium | 3 | 3 |
| Cellulose | 0.60 | 0.1 |
| Proximate chemical composition (%) | ||
| Crude protein (%) | 36.83 | 36.95 |
| Crude lipid (%) | 6.94 | 7.01 |
| Nitrogen free extract (%) | 19.96 | 21.48 |
| Gross energy (kJ/g) | 18.64 | 18.58 |
| Moisture (%) | 9.90 | 9.59 |
| Ash (%) | 6.58 | 6.43 |
| Gene Name | Sense and Antisense Primer (5′-3′) | Gene Bank | Product Length |
|---|---|---|---|
| Accession No. | (bp) | ||
| Elongation factor 1 alpha (ef1α) | GTTGGAGTCAACAAGATGGACTCCAC | AB056104 | 198 |
| CTTCCATCCCTTGAACCAGCCCAT | |||
| Nuclear factor [erythroid-derived 2]-like 2 (nrf2) | CCCTTCACCAAAGACAAGCA | MG759384 | 128 |
| TTGAAGTCATCCACAGGCAG | |||
| Kelch-like ECH-associated protein-1 (keap1) | CTCACCCCCAACTTCCTGCAG | MG759382 | 150 |
| GATGAGCTGCGGCACCTTGGG | |||
| Heme oxygenase-1 (ho-1) | GACAGGAGCATCTACCCACAG | KC758864.1 | 113 |
| GTGGCTGCTTTTATCTGCTCG | |||
| NADPH quinine oxidoreductase-1 (nqo-1) | AGCAACAGAGACAACGGCAC | XM_026268231.1 | 176 |
| GTGTGCACCAGTACAGAGGAG | |||
| Catalase (cat) | CTCCAACGGCAACTTCCCAT | JX477239.1 | 102 |
| CACACCTTAGTCAAATCAAA | |||
| Superoxide dismutase (sod) | GTCCGCACTACAACCCTCAT | JQ776518.1 | 134 |
| GGTCACCATTTTATCCACAA | |||
| Glutathione peroxidase (gpx) | GCCCACCCTCTGTTTGTGTT | DQ983598.1 | 244 |
| CAGGTTTATTTCGCCCTCTTC | |||
| Nuclear receptor coactivator 4 (ncoa4) | TTTTGCCAGCGATGAAGCAC | XM_026224501.1 | 198 |
| GCCACTTCTCTTTGTCCCCA | |||
| Transferrin receptor 1 (tfr1) | TATCCCAGAGTGACCGTAG | XM_026237902.1 | 148 |
| TTGTTGGGTTTGGACTTGT | |||
| Divalent metal transporter 1 (dmt1) | GCTGAACTCGTCTGATCTGGAA | XM_026240820.1 | 296 |
| CCTCGAAGTATGTGGAGATGGG | |||
| Iron-responsive element-binding protein 2 (ireb2) | TTGGTGCTGGATGCGAAGA | XM_026202373.1 | 99 |
| GCTCCATCAAATTGGCCGT | |||
| Cellular tumor antigen (p53) | GTGCAAGATAGTCAAGCTGGTG | XM_026210616.1 | 245 |
| CTGGGGTTCTCTCATGGTGG | |||
| Glutathione peroxidase 4 (gpx4) | ATCTAATACGCTGCCTGCCG | XR_003278905.1 | 211 |
| TGATGACAACTTTGCCCCTG | |||
| Ferroportin (fpn) | CCTCGGACATGCTCTGTCAA | XM_026271430.1 | 112 |
| CAGTCCATACACGGCTGTCA | |||
| Ferritin heavy chain 1 (fth1) | TTGCGAAGTTTTTCCGCCATC | XM_026216156.1 | 94 |
| AGAAAGATCCTCCCTCCCCTC | |||
| Apoptosis-inducing factor mitochondria-associated 2 (aifm2) | GTTCAAGCGGGTTTTGCGA | XM_026224404.1 | 216 |
| GGCTGCTTGGTAGGTGTCC | |||
| Prostaglandin-endoperoxide synthase 2 (ptgs2) | AAAAACATCGCCTCCTGCAAC | JX183400.1 | 178 |
| CACTGACTGGAAAGGGGAGAC | |||
| Long chain acyl CoA synthetase 4 (acsl4) | ACAGTATTTATAGCAGTGAGGT | NC_039256.1 | 173 |
| ATAATAATGTTTGCATGTCCGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, L.; Dong, B.; Chen, Q.; Wang, Y.; Han, D.; Zhu, X.; Liu, H.; Zhang, Z.; Yang, Y.; Xie, S.; et al. Effects of Curcumin on Oxidative Stress and Ferroptosis in Acute Ammonia Stress-Induced Liver Injury in Gibel Carp (Carassius gibelio). Int. J. Mol. Sci. 2023, 24, 6441. https://doi.org/10.3390/ijms24076441
Wu L, Dong B, Chen Q, Wang Y, Han D, Zhu X, Liu H, Zhang Z, Yang Y, Xie S, et al. Effects of Curcumin on Oxidative Stress and Ferroptosis in Acute Ammonia Stress-Induced Liver Injury in Gibel Carp (Carassius gibelio). International Journal of Molecular Sciences. 2023; 24(7):6441. https://doi.org/10.3390/ijms24076441
Chicago/Turabian StyleWu, Liyun, Bo Dong, Qiaozhen Chen, Yu Wang, Dong Han, Xiaoming Zhu, Haokun Liu, Zhimin Zhang, Yunxia Yang, Shouqi Xie, and et al. 2023. "Effects of Curcumin on Oxidative Stress and Ferroptosis in Acute Ammonia Stress-Induced Liver Injury in Gibel Carp (Carassius gibelio)" International Journal of Molecular Sciences 24, no. 7: 6441. https://doi.org/10.3390/ijms24076441