
The Cytosolic Acetoacetyl-CoA Thiolase TaAACT1 Is Required for Defense against Fusarium pseudograminearum in Wheat

Abstract
:1. Introduction
2. Results
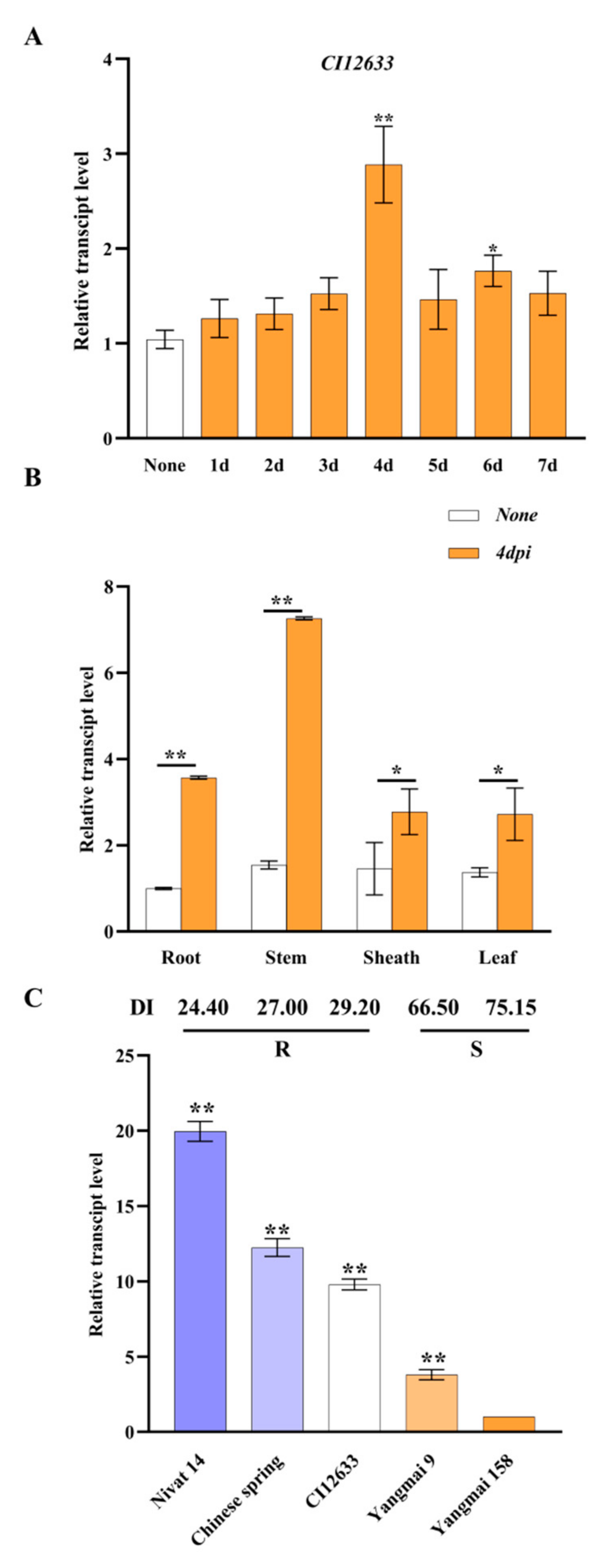
2.1. Identification of TaAACT1 Gene Involved in Wheat Resistance to FCR
2.2. Sequence and Phylogenetic Characteristics of TaAACT1
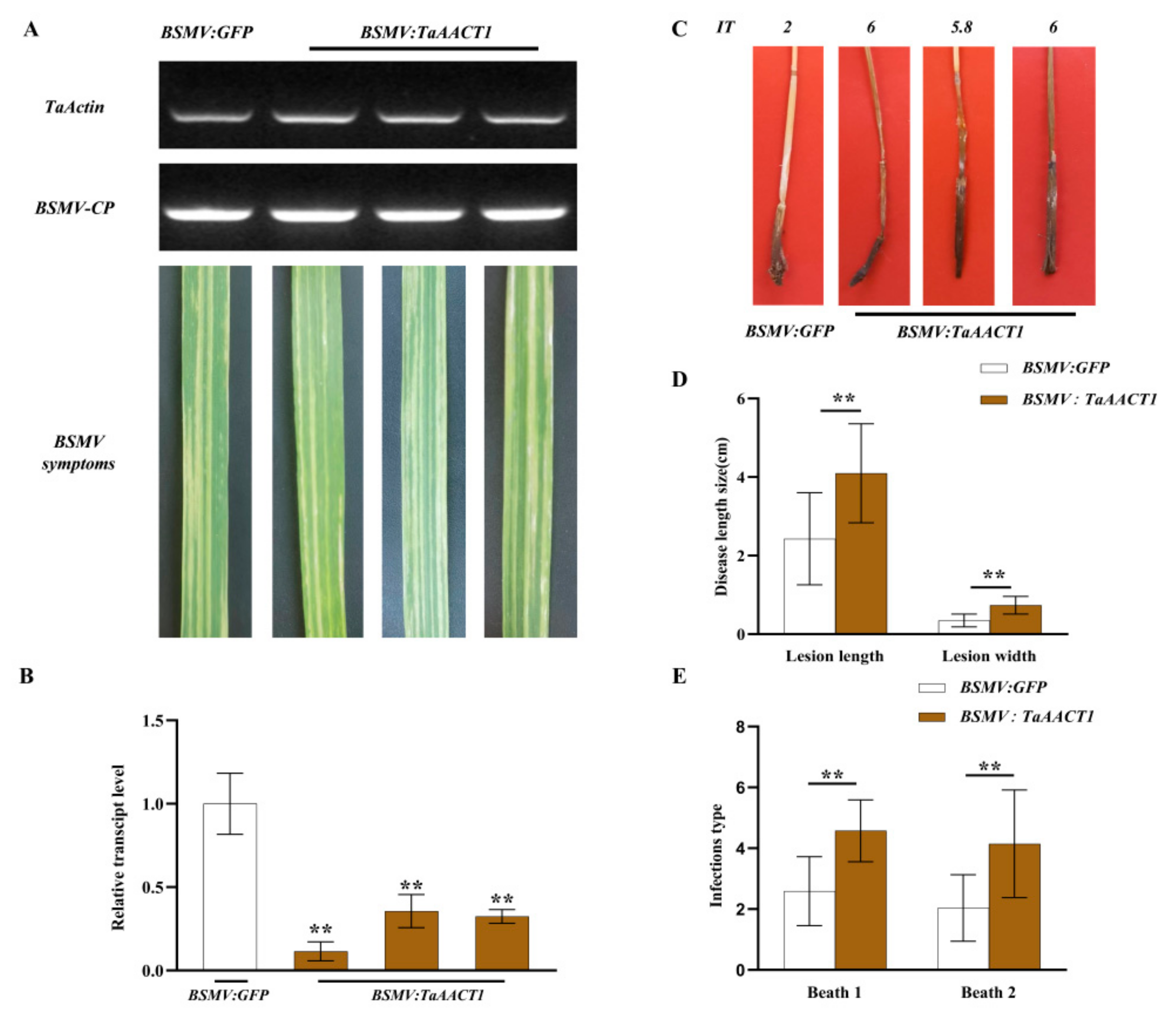
2.3. TaAACT1 Is Required for Wheat Resistance to F. pseudograminearum Infection
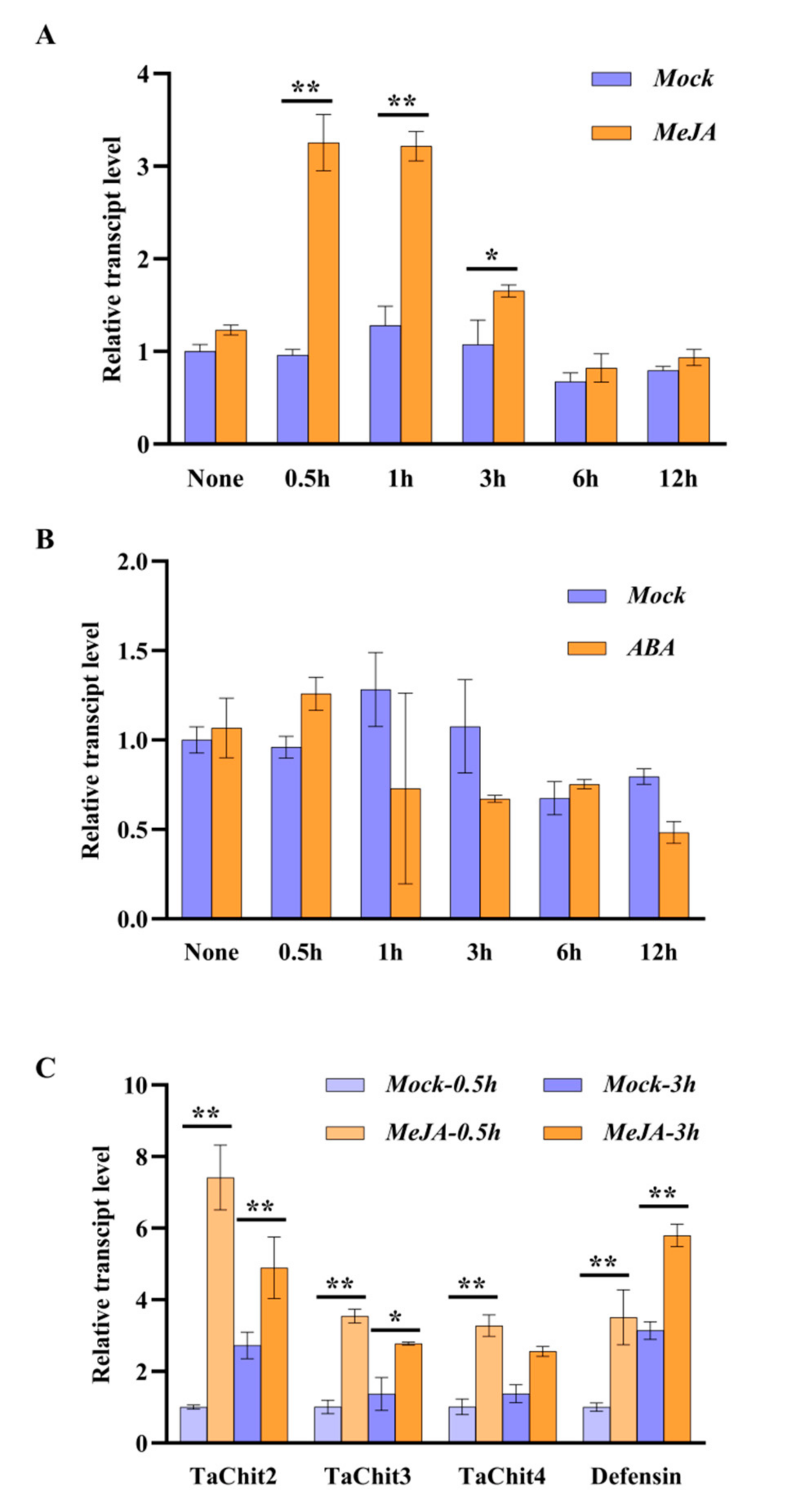
2.4. TaAACT1 Positively Regulates the Expression of Defense Genes
2.5. TaAACT1 and Its Modulated Defense Genes Are Responsive to an Exogenous Jasmonic Acid Stimulus
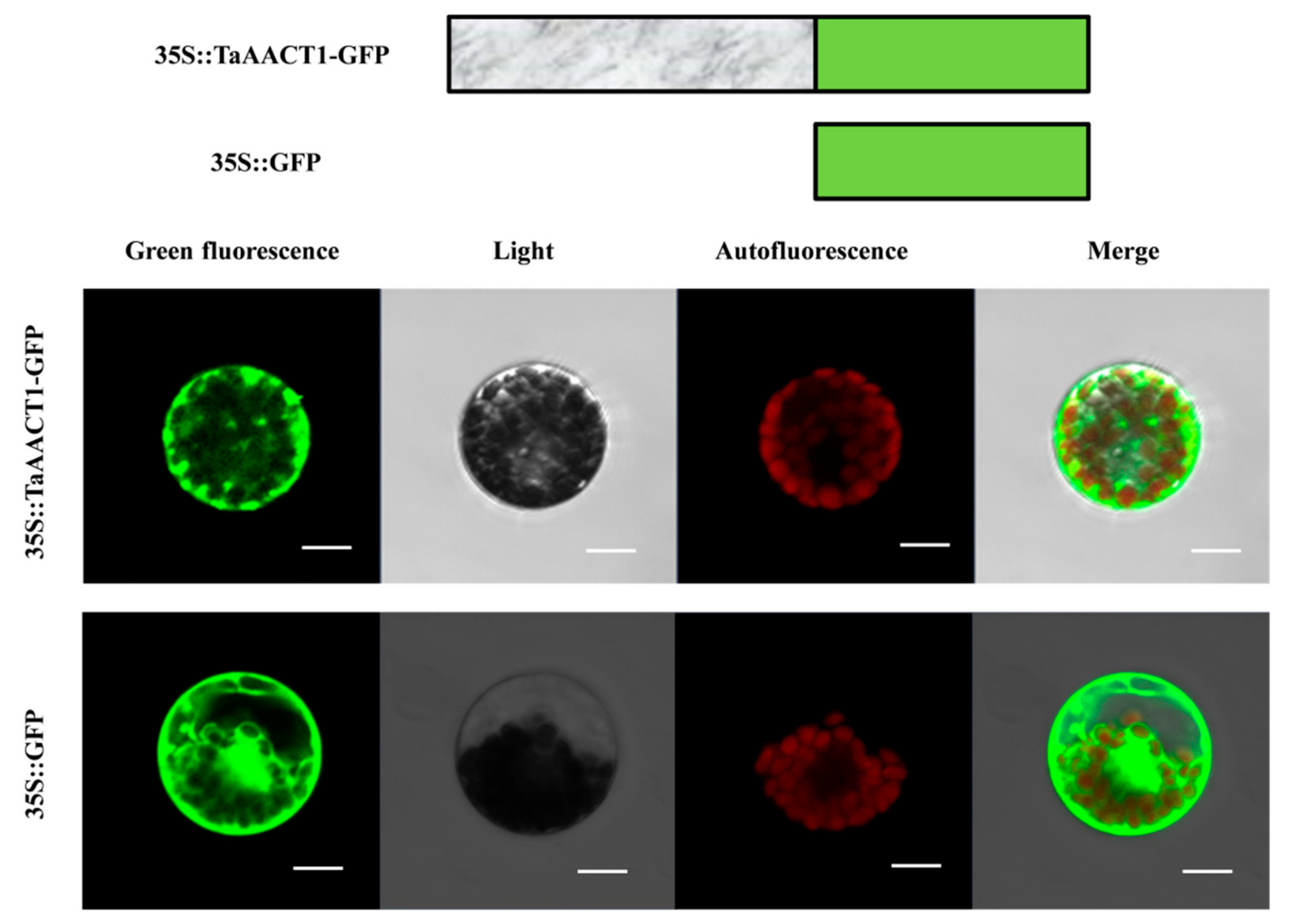
2.6. TaAACT1 Protein Localizes in the Cytosol in Wheat
3. Discussion
4. Materials and Methods
4.1. Plant and Fungal Materials, Primers, and Treatments
4.2. RNA Extraction, cDNA Synthesis, and RT-qPCR Analysis
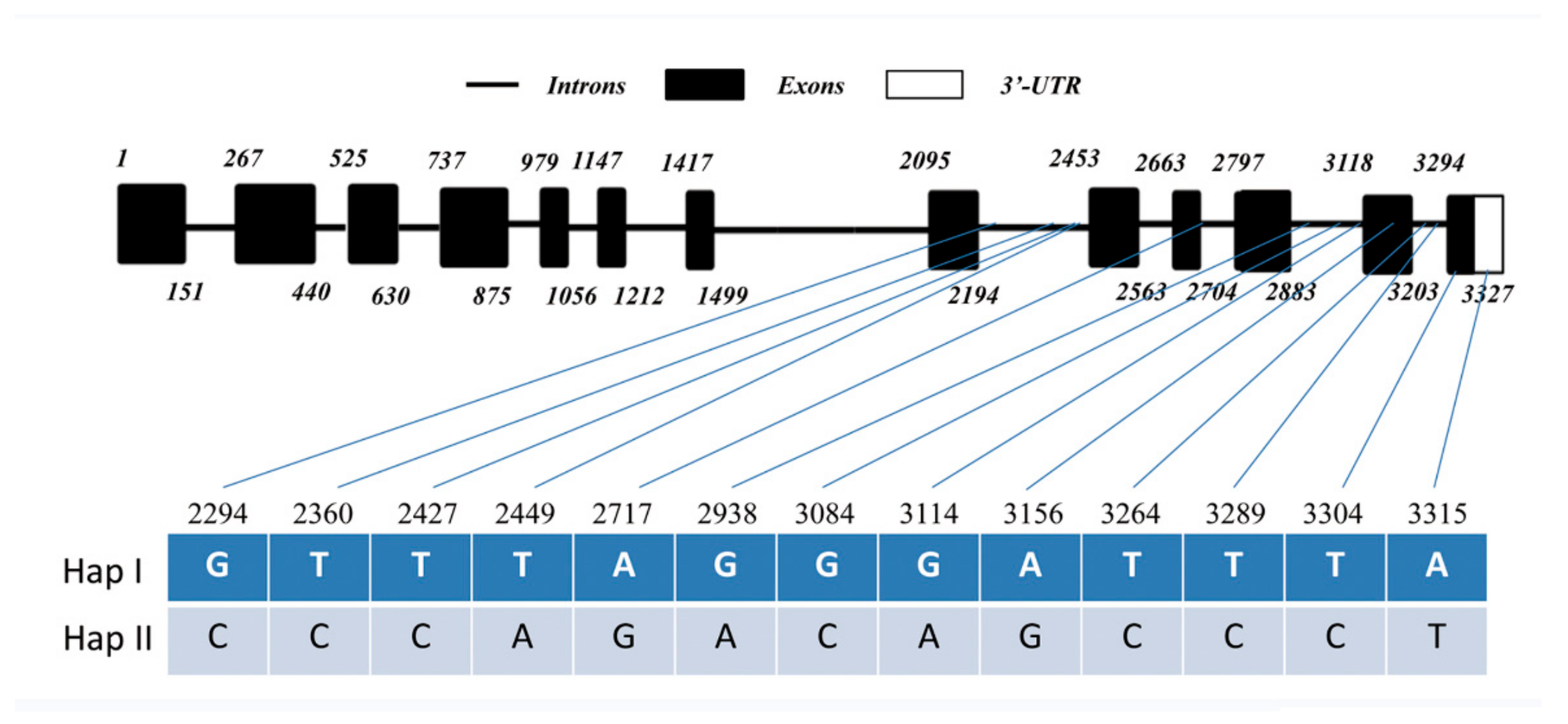
4.3. SNP Analysis
4.4. Sequence and Phylogenetic Analyses of TaAACT1
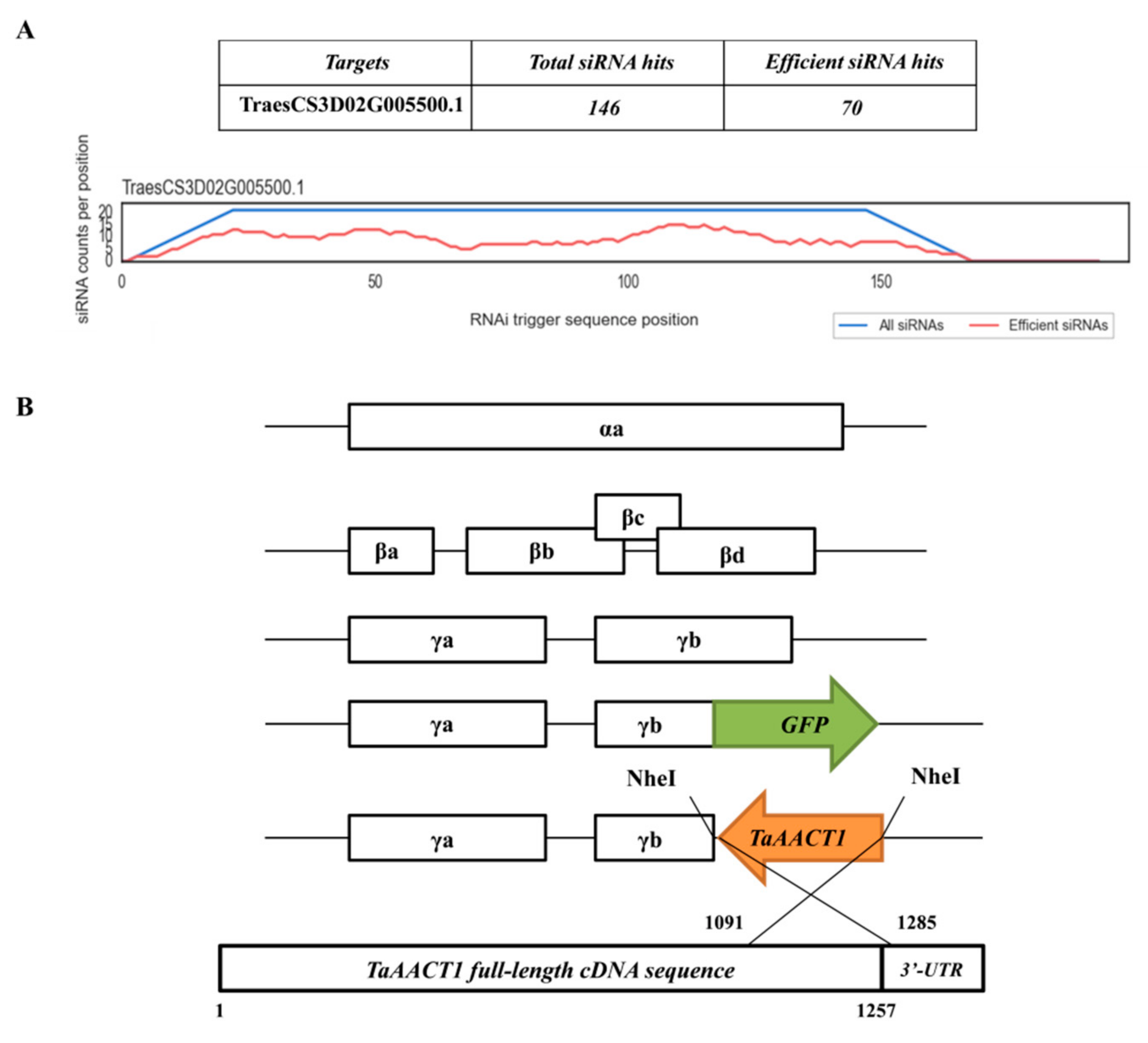
4.5. VIGS of TaAACT1 in Wheat CI12633
4.6. Assessment and Scoring of FCR Disease in BSMV-VIGS Wheat Plants
4.7. Subcellular Localization of TaAACT1 Protein
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Soto, G.; Stritzler, M.; Lisi, C.; Alleva, K.; Pagano, M.E.; Ardila, F.; Mozzicafreddo, M.; Cuccioloni, M.; Angeletti, M.; Ayub, N.D. Acetoacetyl-CoA thiolase regulates the mevalonate pathway during abiotic stress adaptation. J. Exp. Bot. 2011, 62, 5699–5711. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.H.; Norvienyeku, J.; Yu, J.; Chen, M.L.; Cai, R.L.; Hong, Y.H.; Chen, L.M.; Zhang, D.M.; Wang, B.H.; Zhou, J.; et al. Two different subcellular-localized Acetoacetyl-CoA acetyltransferases differentiate diverse functions in Magnaporthe oryzae. Fungal Genet. Biol. 2015, 83, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Wilding, E.I.; Brown, J.R.; Bryant, A.P.; Chalker, A.F.; Holmes, D.J.; Ingraham, K.A.; Iordanescu, S.; So, C.Y.; Rosenberg, M.; Gwynn, M.N. Identification, evolution, and essentiality of the mevalonate pathway for isopentenyl diphosphate biosynthesis in gram-positive cocci. J. Bacteriol. 2000, 182, 4319–4327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Disch, A.; Hemmerlin, A.; Bach, T.J.; Rohmer, M. Mevalonate-derived isopentenyl diphosphate is the biosynthetic precursor of ubiquinone prenyl side chain in tobacco BY-2 cells. Biochem. J. 1998, 331 Pt 2, 615–621. [Google Scholar] [CrossRef] [Green Version]
- Laule, O.; Furholz, A.; Chang, H.-S.; Zhu, T.; Wang, X.; Heifetz, P.B.; Gruissem, W.; Lange, M. Crosstalk between cytosolic and plastidial pathways of isoprenoid biosynthesis in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2003, 100, 6866–6871. [Google Scholar] [CrossRef] [Green Version]
- Carrie, C.; Murcha, M.W.; Millar, A.H.; Smith, S.M.; Whelan, J. Nine 3-ketoacyl-CoA thiolases (KATs) and acetoacetyl-CoA thiolases (ACATs) encoded by five genes in Arabidopsis thaliana are targeted either to peroxisomes or cytosol but not to mitochondria. Plant Mol. Biol. 2007, 63, 97–108. [Google Scholar] [CrossRef]
- Ahumada, I.; Cairo, A.; Hemmerlin, A.; Gonzalez, V.; Pateraki, I.; Bach, T.J.; Rodriguez-Concepcion, M.; Campos, N.; Boronat, A. Characterisation of the gene family encoding acetoacetyl-CoA thiolase in Arabidopsis. Funct. Plant Biol. FPB 2008, 35, 1100–1111. [Google Scholar] [CrossRef]
- Yasmin, S.; Alcazar-Fuoli, L.; Grundlinger, M.; Puempel, T.; Cairns, T.; Blatzer, M.; Lopez, J.F.; Grimalt, J.O.; Bignell, E.; Haas, H. Mevalonate governs interdependency of ergosterol and siderophore biosyntheses in the fungal pathogen Aspergillus fumigatus. Proc. Natl. Acad. Sci. USA 2012, 109, E497–E504. [Google Scholar] [CrossRef] [Green Version]
- Moses, T.; Pollier, J.; Thevelein, J.M.; Goossens, A. Bioengineering of plant (tri)terpenoids: From metabolic engineering of plants to synthetic biology invivo and invitro. New Phytol. 2013, 200, 27–43. [Google Scholar] [CrossRef]
- Dyer, J.H.; Maina, A.; Gomez, I.D.; Cadet, M.; Oeljeklaus, S.; Schiedel, A.C. Cloning, expression and purification of an acetoacetyl CoA thiolase from sunflower cotyledon. Int. J. Biol. Sci. 2009, 5, 736–744. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zheng, Z.; Tian, Z.N.; Zhang, H.; Zhu, C.Y.; Yao, X.Y.; Yang, Y.X.; Cai, X. Molecular cloning and analysis of an Acetyl-CoA C-acetyltransferase gene (EkAACT) from Euphorbia kansui Liou. Plants 2022, 11, 1539. [Google Scholar] [CrossRef]
- Maeda, H.; Song, W.; Sage, T.L.; Dellapenna, D. Tocopherols play a crucial role in low-temperature adaptation and phloem loading in Arabidopsis. Plant Cell 2006, 18, 2710–2732. [Google Scholar] [CrossRef] [Green Version]
- Abbasi, A.-R.; Hajirezaei, M.; Hofius, D.; Sonnewald, U.; Voll, L.M. Specific roles of alpha- and gamma-tocopherol in abiotic stress responses of transgenic tobacco. Plant Physiol. 2007, 143, 1720–1738. [Google Scholar] [CrossRef] [Green Version]
- Matringe, M.; Ksas, B.; Rey, P.; Havaux, M. Tocotrienols, the unsaturated forms of vitamin E, can function as antioxidants and lipid protectors in tobacco leaves. Plant Physiol. 2008, 147, 764–778. [Google Scholar] [CrossRef] [Green Version]
- Bajda, A.; Konopka-Postupolska, D.; Krzymowska, M.; Henning, J.; Skorupinska-Tudek, K.; Surmacz, L.; Wojcik, J.; Matysiak, Z.; Chojnacki, T.; Skorzynska-Polit, E.; et al. Role of polyisoprenoids in tobacco resistance against biotic stresses. Physiol. Plant. 2009, 135, 351–364. [Google Scholar] [CrossRef]
- Vickers, C.E.; Gershenzon, J.; Lerdau, M.T.; Loreto, F. A unified mechanism of action for volatile isoprenoids in plant abiotic stress. Nat. Chem. Biol. 2009, 5, 283–291. [Google Scholar] [CrossRef]
- Suzuki, M.; Nakagawa, S.; Kamide, Y.; Kobayashi, K.; Ohyama, K.; Hashinokuchi, H.; Kiuchi, R.; Saito, K.; Muranaka, T.; Nagata, N. Complete blockage of the mevalonate pathway results in male gametophyte lethality. J. Exp. Bot. 2009, 60, 2055–2064. [Google Scholar] [CrossRef] [Green Version]
- Bohmert, K.; Balbo, I.; Steinbuchel, A.; Tischendorf, G.; Willmitzer, L. Constitutive expression of the beta-ketothiolase gene in transgenic plants. A major obstacle for obtaining polyhydroxybutyrate-producing plants. Plant Physiol. 2002, 128, 1282–1290. [Google Scholar] [CrossRef] [Green Version]
- Poole, G.J.; Smiley, R.W.; Paulitz, T.C.; Walker, C.A.; Carter, A.H.; See, D.R.; Garland-Campbell, K. Identification of quantitative trait loci (QTL) for resistance to Fusarium crown rot (Fusarium pseudograminearum) in multiple assay environments in the Pacific Northwestern US. Theor. Appl. Genet. 2012, 125, 91–107. [Google Scholar] [CrossRef] [Green Version]
- Smiley, R.W.; Gourlie, J.A.; Easley, S.A.; Patterson, L.-M.; Whittaker, R.G. Crop damage estimates for crown rot of wheat and barley in the Pacific Northwest. Plant Dis. 2005, 89, 595–604. [Google Scholar] [CrossRef] [Green Version]
- Kazan, K.; Gardiner, D.M. Fusarium crown rot caused by Fusarium pseudograminearum in cereal crops: Recent progress and future prospects. Mol. Plant Pathol. 2018, 19, 1547–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Pan, Y.B.; Singh, P.K.; He, X.Y.; Ren, Y.; Zhao, L.; Zhang, N.; Cheng, S.H.; Chen, F. Investigation and genome-wide association study for Fusarium crown rot resistance in Chinese common wheat. BMC Plant Biol. 2019, 19, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.J.; Ogbonnaya, F.C. Resistance to Fusarium crown rot in wheat and barley: A review. Plant Breed. 2015, 134, 365–372. [Google Scholar] [CrossRef]
- Jin, J.J.; Duan, S.N.; Qi, Y.Z.; Yan, S.H.; Li, W.; Li, B.Y.; Xie, C.J.; Zhen, W.C.; Ma, J. Identification of a novel genomic region associated with resistance to Fusarium crown rot in wheat. Theor. Appl. Genet. 2020, 133, 2063–2073. [Google Scholar] [CrossRef]
- Yang, X.; Zhong, S.B.; Zhang, Q.J.; Ren, Y.; Sun, C.W.; Chen, F. A loss-of-function of the dirigent gene TaDIR-B1 improves resistance to Fusarium crown rot in wheat. Plant Biotechnol. J. 2021, 19, 866–868. [Google Scholar] [CrossRef]
- Qi, H.J.; Guo, F.L.; Lv, L.J.; Zhu, X.L.; Zhang, L.; Yu, J.F.; Wei, X.N.; Zhang, Z.Y. The wheat wall-associated receptor-like kinase TaWAK-6D mediates broad resistance to two fungal pathogens Fusarium pseudograminearum and Rhizoctonia cerealis. Front. Plant Sci. 2021, 12, 2322. [Google Scholar] [CrossRef]
- Wu, T.C.; Guo, F.L.; Xu, G.B.; Yu, J.F.; Zhang, L.; Wei, X.N.; Zhu, X.L.; Zhang, Z.Y. The receptor-like kinase TaCRK-7A inhibits Fusarium pseudograminearum growth and mediates resistance to Fusarium crown rot in wheat. Biology 2021, 10, 1122. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, J.Z.; Zhang, H.Z.; Guo, B.M.; Ning, S.Z.; Chen, Y.X.; Lu, P.; Wu, Q.H.; Li, M.M.; Zhang, D.Y.; et al. Mapping stripe rust resistance gene YrZH22 in Chinese wheat cultivar Zhoumai 22 by bulked segregant RNA-Seq (BSR-Seq) and comparative genomics analyses. Theor. Appl. Genet. 2017, 130, 2191–2201. [Google Scholar] [CrossRef]
- Benhamou, N.; Broglie, K.; Broglie, R.; Chet, I. Antifungal effect of bean endochitinase on Rhizoctonia solani: Ultrastructural changes and cytochemical aspects of chitin breakdown. Can. J. Microbiol. 1993, 39, 318–328. [Google Scholar] [CrossRef]
- Thevissen, K.; Terras, F.R.; Broekaert, W.F. Permeabilization of fungal membranes by plant defensins inhibits fungal growth. Appl. Environ. Microbiol. 1999, 65, 5451–5458. [Google Scholar] [CrossRef] [Green Version]
- Thomma, B.P.H.J.; Cammue, B.P.A.; Thevissen, K. Plant defensins. Planta 2002, 216, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Kirubakaran, S.I.; Sakthivel, N. Heterologous expression of new antifungal chitinase from wheat. Protein Expr. Purif. 2007, 56, 100–109. [Google Scholar] [CrossRef]
- Lacerda, A.F.; Vasconcelos, E.a.R.; Pelegrini, P.B.; De Sa, M.F.G. Antifungal defensins and their role inplant defense. Front. Microbiol. 2014, 5, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Xu, X.Y.; Tian, L.L.; Wang, G.L.; Zhang, X.Y.; Wang, X.Y.; Guo, W.Z. Discovery and identification of candidate genes from the chitinase gene family for Verticillium dahliae resistance in cotton. Sci. Rep. 2016, 6, 29022. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; Wang, K.; Zhang, Z.Y. Ecotopic expression of the antimicrobial peptide DmAMP1W improves resistance of transgenic wheat to two diseases: Sharp eyespot and common root rot. Int. J. Mol. Sci. 2020, 21, 647. [Google Scholar] [CrossRef] [Green Version]
- FAO. World Food and Agriculture—Statistical Yearbook 2020; FAO: Rome, Italy, 2020. [Google Scholar]
- Shen, Q.-H.; Saijo, Y.; Mauch, S.; Biskup, C.; Bieri, S.; Keller, B.; Seki, H.; Ulker, B.; Somssich, I.E.; Schulze-Lefert, P. Nuclear activity of MLA immune receptors links isolate-specific and basal disease-resistance responses. Science 2007, 315, 1098–1103. [Google Scholar] [CrossRef] [Green Version]
- Gunupuru, L.R.; Arunachalam, C.; Malla, K.B.; Kahla, A.; Perochon, A.; Jia, J.G.; Thapa, G.; Doohan, F.M. A wheat cytochrome P450 enhances both resistance to deoxynivalenol and grain yield. PLoS ONE 2018, 13, e0204992. [Google Scholar] [CrossRef] [Green Version]
- Thevissen, K.; Osborn, R.W.; Acland, D.P.; Broekaert, W.F. Specific binding sites for an antifungal plant defensin from Dahlia (Dahlia merckii) on fungal cells are required for antifungal activity. Mol. Plant-Microbe Interact. 2000, 13, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Sanz-Martin, J.M.; Pacheco-Arjona, J.R.; Bello-Rico, V.; Vargas, W.A.; Monod, M.; Diaz-Minguez, J.M.; Thon, M.R.; Sukno, S.A. A highly conserved metalloprotease effector enhances virulence in the maize anthracnose fungus Colletotrichum graminicola. Mol. Plant Pathol. 2016, 17, 1048–1062. [Google Scholar] [CrossRef] [Green Version]
- Kasprzewska, A. Plant chitinases--regulation and function. Cell. Mol. Biol. Lett. 2003, 8, 809–824. [Google Scholar]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.Y.; Liu, X.; Wang, X.D.; Zhou, M.P.; Zhou, X.Y.; Ye, X.G.; Wei, X.N. An R2R3 MYB transcription factor in wheat, TaPIMP1, mediates host resistance to Bipolaris sorokiniana and drought stresses through regulation of defense- and stress-related genes. New Phytol. 2012, 196, 1155–1170. [Google Scholar] [CrossRef]
- Zhu, X.L.; Qi, L.; Liu, X.; Cai, S.B.; Xu, H.J.; Huang, R.F.; Li, J.R.; Wei, X.N.; Zhang, Z.Y. The wheat ethylene response factor transcription factor PATHOGEN-INDUCED ERF1 mediates host responses to both the necrotrophic pathogen Rhizoctonia cerealis and freezing stresses. Plant Physiol. 2014, 164, 1499–1514. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.L.; Lu, C.G.; Du, L.P.; Ye, X.G.; Liu, X.; Coules, A.; Zhang, Z.Y. The wheat NB-LRR gene TaRCR1 is required for host defence response to the necrotrophic fungal pathogen Rhizoctonia cerealis. Plant Biotechnol. J. 2017, 15, 674–687. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.L.; Rong, W.; Wang, K.; Guo, W.; Zhou, M.P.; Wu, J.Z.; Ye, X.G.; Wei, X.N.; Zhang, Z.Y. Overexpression of TaSTT3b-2B improves resistance to sharp eyespot and increases grain weight in wheat. Plant Biotechnol. J. 2022, 20, 777–793. [Google Scholar] [CrossRef]
- Liu, X.; Zhu, X.L.; Wei, X.N.; Lu, C.G.; Shen, F.D.; Zhang, X.W.; Zhang, Z.Y. The wheat LLM-domain-containing transcription factor TaGATA1 positively modulates host immune response to Rhizoctonia cerealis. J. Exp. Bot. 2020, 71, 344–355. [Google Scholar] [CrossRef]
- Zheng, H.Y.; Dong, L.L.; Han, X.Y.; Jin, H.B.; Yin, C.C.; Han, Y.L.; Li, B.; Qin, H.J.; Zhang, J.S.; Shen, Q.H.; et al. The TuMYB46L-TuACO3 module regulates ethylene biosynthesis in einkorn wheat defense to powdery mildew. New Phytol. 2020, 225, 2526–2541. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.L.; Wu, T.C.; Shen, F.D.; Xu, G.B.A.; Qi, H.J.; Zhang, Z.Y. The cysteine-rich receptor-like kinase TaCRK3 contributes to defense against Rhizoctonia cerealis in wheat. J. Exp. Bot. 2021, 72, 6904–6919. [Google Scholar] [CrossRef]
- Wang, K.; Shao, Z.Y.; Guo, F.L.; Wang, K.; Zhang, Z.Y. The mitogen-activated protein kinase kinase TaMKK5 mediates immunity via the TaMKK5-TaMPK3-TaERF3 module. Plant Physiol. 2021, 187, 2323–2337. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Leon-Reyes, A.; Van Der Ent, S.; Van Wees, S.C.M. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 2009, 5, 308–316. [Google Scholar] [CrossRef] [Green Version]
- Peng, X.X.; Wang, H.H.; Jang, J.C.; Xiao, T.; He, H.H.; Jiang, D.; Tang, X.K. OsWRKY80-OsWRKY4 module as a positive regulatory circuit in rice resistance against Rhizoctonia solani. Rice 2016, 9, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.W.X.; Zhang, X.; Zhang, Z. Establishment of a highly-efficient transformation system of wheat protoplasts. J. Plant Genet. Resour. 2017, 18, 117–124. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SNP Site (no.) | Nucleotide in Haplotype I | Nucleotide in Haplotype II | Amino Acid Change |
|---|---|---|---|
| 2294 | G | C | - |
| 2360 | T | C | - |
| 2427 | T | C | - |
| 2449 | T | A | - |
| 2717 | A | G | - |
| 2938 | G | A | - |
| 3084 | G | C | - |
| 3114 | G | A | - |
| 3156 | A | G | - |
| 3264 | T | C | - |
| 3289 | T | C | - |
| 3304 | T | C | S→P |
| 3315 | A | T | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, F.; Zhu, X.; Luo, C.; Liu, Z.; Zhang, Z. The Cytosolic Acetoacetyl-CoA Thiolase TaAACT1 Is Required for Defense against Fusarium pseudograminearum in Wheat. Int. J. Mol. Sci. 2023, 24, 6165. https://doi.org/10.3390/ijms24076165
Xiong F, Zhu X, Luo C, Liu Z, Zhang Z. The Cytosolic Acetoacetyl-CoA Thiolase TaAACT1 Is Required for Defense against Fusarium pseudograminearum in Wheat. International Journal of Molecular Sciences. 2023; 24(7):6165. https://doi.org/10.3390/ijms24076165
Chicago/Turabian StyleXiong, Feng, Xiuliang Zhu, Changsha Luo, Zhixiang Liu, and Zengyan Zhang. 2023. "The Cytosolic Acetoacetyl-CoA Thiolase TaAACT1 Is Required for Defense against Fusarium pseudograminearum in Wheat" International Journal of Molecular Sciences 24, no. 7: 6165. https://doi.org/10.3390/ijms24076165