
Relevance of Sugar Transport across the Cell Membrane

Abstract
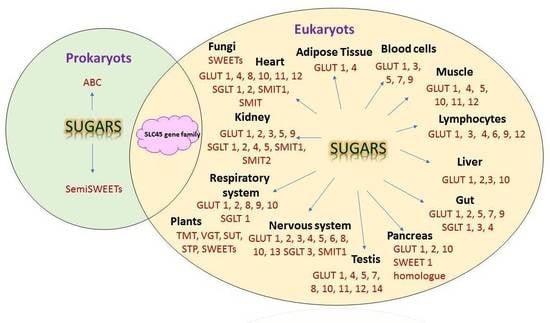
:
1. Introduction
2. Gateways through the Membrane
3. SGLTs
4. GLUTs
4.1. SGLT and GLUT Participation in Diverse Physiological Conditions
4.1.1. Reproductive System
4.1.2. Hepatic System
4.1.3. Renal System
4.1.4. Cardiovascular System
4.1.5. Skeletal System
4.1.6. Immune System
4.1.7. Nervous System
4.2. SGLT and GLUT Participation in Disease
4.2.1. Obesity
4.2.2. Diabetes
4.2.3. Inflammation and Immunodeficiency
4.2.4. Infertility and Miscarriage
4.2.5. Hypertension
4.2.6. Metabolic Disturbances
4.2.7. Cancer
4.2.8. Epilepsy
4.2.9. Alzheimer Disease
4.2.10. Aging
4.2.11. Sleep Disturbances
4.2.12. Osteoporosis
5. SWEETs
6. ABCs
7. Other Transporters
8. Economic Importance of Sugar Transporters
9. Targeting Glucose Transporters in Clinics
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 2-DG | 2-deoxy-D-glucose |
| αMG | α-methyl-D-glucopyranoside |
| ABC | ATP-Binding Cassette |
| AD | Alzheimer’s disease |
| AGEs | Advanced glycation end-products |
| ATP | Adenosine Triphosphate |
| BBB | Brain–blood barrier |
| CA1 | Hippocampal Pyramidal Cells |
| CDG-IIf | Congenital disorder of N-linked glycosylation IIf |
| CP | Choroid Plexus |
| CWI | Cell Wall Invertase |
| cDNADHA | Complementary Deoxyribonucleic AcidDehydroascorbic acid |
| E.c.MalK | Maltose transporter from E. coli |
| ECF | Eenergy-coupling factor transporters |
| EGFR | Epidermal Growth Factor Receptor |
| ER | Endoplasmic Reticulum |
| FfZ | Fungal Sugar Facilitators |
| FDG | Fluorodeoxyglucose |
| GLUT | Glucose Transporters |
| GLUT1DS | GLUT1 Deficiency Syndrome |
| HEX | Hexosaminidase |
| HG | Hyperglycemia |
| HIF-1 | Hypoxia Inducible factor 1 |
| HMIT | H-Mio-Inositol Transporter |
| IL-1β | Interleukin 1β |
| MFS | Major Facilitator Superfamily |
| MRCA | Most Recent Common Ancestor |
| MST | Monosaccharide Transporter |
| MtN3 | Bidirectional Sugar Transporter N3 |
| mTOR | Mammalian Target of Rapamycin |
| mRNA | Messenger Ribonucleic Acid |
| miRNA | Micro ribonucleic acid |
| MTP | Monosaccharide Transporter |
| MYC | Transcription factors c-Myc or MYC oncogene |
| NaGLT | Novel sodium-dependent sugar transporter |
| NBD | Nucleotide Binding Domains |
| ND | Not described |
| NST | Nucleotide sugar transporter |
| PET | Positron emission tomography |
| PglK | oligosaccharide ABC transporter |
| RAG1AP1 | Recombination Activating Gene 1 Activating Protein 1 |
| rCGU | regional cerebral glucose utilization |
| RiMST | Rhizophagus irregularis monosaccharide transporters |
| ROS | Reactive Oxygen Species |
| RUNX2 | Runt-related Osteoblast Transcription Factor |
| SGLT | Sodium-Dependent Glucose Transporters |
| shRNA | Short hairpin ribonucleic acid |
| siRNA | Silencing ribonucleic acid |
| SIRT | Sirtuins |
| SIX1 | Sine Oculis Homeobox 1 |
| SLC | Solute Carriers |
| SMIT | Osmolyte Transporters |
| SNAP | Synaptophysin |
| ST | Sugar Transporters |
| STF-3 | Specific transcription factor 3 |
| STP | Sugar Transporter Protein |
| SUT or /SUC | Sucrose Transporter or sucrose carrier |
| SWEET | Sugars Will Eventually be Exported Transporter |
| Swt-1 | Sugar Transporter SWEET1 |
| T2DM | Type 2 Diabetes Mellitus |
| TGF-β1 | Transforming growth factor β1 |
| TM | Transmembrane regions |
| TMD | Trans Membrane Domain |
| TMT | Tonoplast Monosaccharide Transporter |
| TNF-α | Tumor necrosis factor α |
| TRPV | Transient Receptor Potential Vanilloid |
| URAT | Ureate Transporter |
| VGT | Vacuolar Glucose Transporter |
| VLPO | Ventrolateral Preoptic Nucleus |
| WZB | Small molecule GLUT1 inhibitor |
References
- Freeman, C.R.; Zehra, A.; Ramirez, V.; Wiers, C.E.; Volkow, N.D.; Wang, G.-J. Impact of sugar on the body, brain, and behavior. Front. Biosci. 2018, 23, 2255–2266. [Google Scholar] [CrossRef]
- Fairweather, S.J.; Shah, N.; Brӧer, S. Heteromeric Solute Carriers: Function, Structure, Pathology and Pharmacology. Adv. Exp. Med. Biol. Protein Rev. 2021, 21, 13–127. [Google Scholar] [CrossRef]
- Gatenby, R.A. The Role of Cell Membrane Information Reception, Processing, and Communication in the Structure and Function of Multicellular Tissue. Int. J. Mol. Sci. 2019, 20, 3609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, C.; Coelho, M.A.; Salema-Oom, M.; Gonçalves, P. Stepwise Functional Evolution in a Fungal Sugar Transporter Family. Mol. Biol. Evol. 2016, 33, 352–366. [Google Scholar] [CrossRef]
- Scheepers, A.; Joost, H.G.; Schurmann, A. The glucose transporter families SGLT and GLUT: Molecular basis of normal and aberrant function. J. Parenter Enter. Nutr. 2004, 28, 364–371. [Google Scholar] [CrossRef] [Green Version]
- Wright, E.M.; Loo, D.D.F.; Hirayama, B.A. Biology of Human Sodium Glucose Transporters. Physiol. Rev. 2011, 91, 733–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, D.; Sun, P.; Yan, C.; Ke, M.; Jiang, X.; Xiong, L.; Ren, W.; Hirata, K.; Yamamoto, M.; Fan, S.; et al. Molecular basis of ligand recognition and transport by glucose transporters. Nature 2015, 526, 391–396. [Google Scholar] [CrossRef]
- Głuchowska, K.; Pliszka, M.; Szablewski, L. Expression of glucose transporters in human neurodegenerative diseases. Biochem. Biophys. Res. Commun. 2021, 540, 8–15. [Google Scholar] [CrossRef]
- Van Steenbergen, A.; Balteau, M.; Ginion, A.; Ferté, L.; Battault, S.; de Meester de Ravenstein, C.; Balligand, J.L.; Daskalopoulos, E.P.; Gilon, P.; Despa, F.; et al. Sodium-myoinositol cotransporter-1, SMIT1, mediates the production of reactive oxygen species induced by hyperglycemia in the heart. Sci. Rep. 2017, 7, 41166. [Google Scholar] [CrossRef] [PubMed]
- Cura, A.J.; Carruthers, A. The role of Monosaccharide Transport Proteins in carbohydrate assimilation, distribution, metabolism and homeostasis. Compr Physiol. 2012, 2, 863–914. [Google Scholar] [CrossRef] [Green Version]
- Holman, G.D. Structure, function and regulation of mammalian glucose transporters of the SLC2 family. Pflügers Arch. —Eur. J. Physiol. 2020, 472, 1155–1175. [Google Scholar] [CrossRef]
- Ceder, M.M.; Fredriksson, R. A phylogenetic analysis between humans and D. melanogaster: A repertoire of solute carriers in humans and flies. Gene 2022, 809, 146033. [Google Scholar] [CrossRef]
- Castrejón, V.; Carbó, R.; Martínez, M. Mecanismos macromoleculares que intervienen en el transporte de la glucosa. Revista de Educación Bioquímica 2007, 26, 49–57. [Google Scholar]
- Villagrán, M.; Burgos, C.F.; Rivas, C.I.; Mardones, L. Identification of Structural Determinants of the Transport of the Dehydroascorbic Acid Mediated by Glucose Transport GLUT1. Molecules 2023, 28, 521. [Google Scholar] [CrossRef] [PubMed]
- Ng, B.G.; Sosicka, P.; Xia, Z. Freeze HHGLUT1 is a highly efficient L-fucose transporter. J. Biol. Chem. 2023, 299, 102738. [Google Scholar] [CrossRef] [PubMed]
- Szablewski, L. Glucose transporters in healthy heart and in cardiac disease. Int. J. Cardiol. 2017, 230, 70–75. [Google Scholar] [CrossRef]
- Mueckler, M.; Thorens, B. The SLC2 (GLUT) family of membrane transporters. Mol. Asp. Med. 2013, 34, 121–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lizák, B.; Szarka, A.; Kim, Y.; Choi, K.; Németh, C.E.; Marcolongo, P.; Benedetti, A.; Bánhegyi, G.; Margittai, É. Glucose Transport and Transporters in the Endomembranes. Int. J. Mol. Sci. 2019, 20, 5898. [Google Scholar] [CrossRef] [Green Version]
- Vrhovac Madunić, I.; Madunić, J.; Breljak, D.; Karaica, D.; Sabolić, I. Sodium-glucose cotransporters: New targets of cancer therapy? Arhiv za Higijenu Rada i Toksikologiju 2018, 69, 278–285. [Google Scholar] [CrossRef] [Green Version]
- Ermini, L.; Nuzzo, A.M.; Ietta, F.; Romagnoli, R.; Moretti, L.; Masturzo, B.; Paulesu, L.; Rolfo, A. Placental Glucose Transporters and Response to Bisphenol A in Pregnancies from of Normal and Overweight Mothers. Int. J. Mol. Sci. 2021, 22, 6625. [Google Scholar] [CrossRef]
- Frolova, A.I.; Moley, K.H. Glucose transporters in the uterus: An analysis of tissue distribution and proposed physiological roles. Reproduction 2011, 142, 211–220. [Google Scholar] [CrossRef] [Green Version]
- Guillet-Deniau, I.; Leturque, A.; Girard, J. Expression and cellular localization of glucose transporters (GLUT1, GLUT3, GLUT4) during differentiation of myogenic cells isolated from rat fetuses. J. Cell Sci. 1994, 107, 487–496. [Google Scholar] [CrossRef]
- Soták, M.; Casselbrant, A.; Rath, E.; Zietek, T.; Strömstedt, M.; Adingupu, D.D.; Karlsson, D.; Fredin, M.F.; Ergang, P.; Pácha, J.; et al. Intestinal sodium/glucose cotransporter 3 expression is epithelial and downregulated in obesity. Life Sci. 2021, 267, 118974. [Google Scholar] [CrossRef]
- Leturque, A.; Brot-Laroche, E.; Le Gall, M. GLUT2 mutations, translocation, and receptor function in diet sugar managing. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E985–E992. [Google Scholar] [CrossRef]
- Keembiyehetty, C.; Augustin, R.; Carayannopoulos, M.O.; Steer, S.; Manolescu, A.; Cheeseman, C.I.; Moley, K.H. Mouse Glucose Transporter 9 Splice Variants Are Expressed in Adult Liver and Kidney and Are Upregulated in Diabetes. Mol. Endocrinol. 2006, 20, 686–697. [Google Scholar] [CrossRef] [Green Version]
- Wright, E.M. Glucose transport families SLC5 and SLC50. Mol. Asp. Med. 2013, 34, 183–196. [Google Scholar] [CrossRef]
- Nakano, D.; Akiba, J.; Tsutsumi, T.; Kawaguchi, M.; Yoshida, T.; Koga, H.; Kawaguchi, T. Hepatic expression of sodium–glucose cotransporter 2 (SGLT2) in patients with chronic liver disease. Med. Mol. Morphol. 2022, 55, 304–315. [Google Scholar] [CrossRef] [PubMed]
- Anzai, N.; Jutabha, P.; Amonpatumrat-Takahashi, S.; Sakurai, H. Recent advances in renal urate transport: Characterization of candidate transporters indicated by genome-wide association studies. Clin. Exp. Nephrol. 2012, 1, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Uthman, L.; Baartscheer, A.; Schumacher, C.A.; Fiolet, J.W.T.; Kuschma, M.C.; Hollmann, M.W.; Coronel, R.; Weber, N.C.; Zuurbier, C.J. Direct Cardiac Actions of Sodium Glucose Cotransporter 2 Inhibitors Target Pathogenic Mechanisms Underlying Heart Failure in Diabetic Patients. Front. Physiol. 2018, 9, 1575. [Google Scholar] [CrossRef]
- Schaan, B.D.A.; Machado, U.F. Glucose transporters in animal models of diabetes and hypertension. Am. J. Physiol. -Ren. Physiol. 2006, 291, F702–F704. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, L.; Auquier, J.; Renguet, E.; Angé, M.; Cumps, J.; Horman, S.; Beauloye, C. Glucose transporters in cardiovascular system in health and disease. Pflügers Arch. Eur. J. Physiol. 2020, 472, 1385–1399. [Google Scholar] [CrossRef] [PubMed]
- Taşdemir, A.; Belice, T.; Yüksel, A. Falls and SGLT-2 inhibitors in geriatric patients—A case report. J. Fam. Med. Prim. Care 2020, 9, 4454–4455. [Google Scholar] [CrossRef]
- Varin, C.; Rancillac, A.; Geoffroy, H.; Arthaud, S.; Fort, P.; Gallopin, T. Glucose Induces Slow-Wave Sleep by Exciting the Sleep-Promoting Neurons in the Ventrolateral Preoptic Nucleus: A New Link between Sleep and Metabolism. J. Neurosci. 2015, 35, 9900–9911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuncheddu, D.; Della Bella, E.; Petta, D.; Bärtschi, C.; Häckel, S.; Deml, M.C.; Stoddart, M.J.; Grad, S.; Basoli, V. Effect of glucose depletion and fructose administration during chondrogenic commitment in human bone marrow-derived stem cells. Stem Cell Res. Ther. 2022, 13, 533. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C.; Dimitriadis, G.; Newsholme, P. Glucose metabolism in lymphoid and inflammatory cells and tissues. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 531–540. [Google Scholar] [CrossRef]
- Carbó, R.; Guarner, V. Insulin effect on glucose transporters in thymocytes and splenocytes from rats with metabolic syndrome. Diabetol. Metab. Syndr. 2010, 2, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koepsell, H. Glucose transporters in brain in health and disease. Pflügers Arch.—Eur. J. Physiol. 2020, 472, 1299–1343. [Google Scholar] [CrossRef]
- Chiba, Y.; Murakami, R.; Matsumoto, K.; Wakamatsu, K.; Nonaka, W.; Uemura, N.; Yanase, K.; Kamada, M.; Ueno, M. Glucose, Fructose, and Urate Transporters in the Choroid Plexus Epithelium. Int. J. Mol. Sci. 2020, 21, 7230. [Google Scholar] [CrossRef]
- Pawlos, A.; Broncel, M.; Woźniak, E.; Gorzelak-Pabiś, P. Neuroprotective Effect of SGLT2 Inhibitors. Molecules 2021, 26, 7213. [Google Scholar] [CrossRef]
- Simpson, I.A.; Appel, N.M.; Hokari, M.; Oki, J.; Holman, G.D.; Maher, F.; Koehler-Stec, E.M.; Vannucci, S.J.; Smith, Q.R. Blood-brain barrier glucose transporter: Effects of hypo- and hyperglycemia revisited. J Neurochem. 1999, 72, 238–247. [Google Scholar] [CrossRef]
- Leão, L.L.; Tangen, G.; Barca, M.L.; Engedal, K.; Santos, S.H.S.; Machado, F.S.M.; de Paula, A.M.B.; Monteiro-Junior, R.S. Does hyperglycemia downregulate glucose transporters in the brain? Med. Hypotheses 2020, 139, 109614. [Google Scholar] [CrossRef] [PubMed]
- Donat, A.; Knapstein, P.R.; Jiang, S.; Baranowsky, A.; Ballhause, T.M.; Frosch, K.H.; Keller, J. Glucose Metabolism in Osteoblasts in Healthy and Pathophysiological Conditions. Int. J. Mol. Sci. 2021, 22, 4120. [Google Scholar] [CrossRef] [PubMed]
- Jurcovicova, J. Glucose transport in brain—effect of inflammation. Endocr. Regul. 2014, 48, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.; Hunter, L.; Trayhurn, P. Expression of Class III facilitative glucose transporter genes (GLUT-10 and GLUT-12) in mouse and human adipose tissues. Biochem. Biophys. Res. Commun. 2003, 308, 43–49. [Google Scholar] [CrossRef]
- Deal, R.A.; Tang, Y.; Fletcher, R.; Torquati, A.; Omotosho, P. Understanding intestinal glucose transporter expression in obese compared to non-obese subjects. Surg. Endosc. 2018, 32, 1755–1761. [Google Scholar] [CrossRef]
- Fiorentino, T.V.; DeVito, F.; Suraci, E.; Marasco, R.; Hribal, M.L.; Luzza, F.; Sesti, G. Obesity, and overweight are linked to increased sodium-glucose cotransporter 1 and glucose transporter 5 levels in duodenum. Obesity 2023, 31, 724–731. [Google Scholar] [CrossRef]
- Jha, D.; Mazumder, P.M. High fat diet administration leads to the mitochondrial dysfunction and selectively alters the expression of class 1 GLUT protein in mice. Mol. Biol. Rep. 2019, 46, 1727–1736. [Google Scholar] [CrossRef]
- Mourelatou, R.; Kostopoulou, E.; Rojas Gil, A.P.; Sinopidis, X.; Kehagias, J.; Linos, D.; Kalfarentzos, F.E.; Spiliotis, B.E. Impaired adipocyte glucose transport regulators in morbid obesity—Possible mechanisms contributing to obesity, an excessive energy condition, would induce GLUT12 downregulation. Eur. Rev. Med. Pharm. Sci. 2022, 26, 2134–2142. [Google Scholar] [CrossRef]
- Gil-Iturbe, E.; Félix-Soriano, E.; Sáinz, N.; Idoate-Bayón, A.; Castilla-Madrigal, R.; Moreno-Aliaga, M.J.; Lostao, M.P. Effect of aging and obesity on GLUT12 expression in small intestine, adipose tissue, muscle, and kidney and its regulation by docosahexaenoic acid and exercise in mice. Appl. Physiol. Nutr. Metab. 2020, 45, 957–967. [Google Scholar] [CrossRef]
- Baader-Paglera, T.; Eckhardtb, M.; Himmelsbachb, F.; Sauer, A.; Stierstorferc, B.E.; Hamiltona, B.S. SGLT6—A pharmacological target for the treatment of obesity? Adipocyte 2018, 7, 277–284. [Google Scholar] [CrossRef] [Green Version]
- Dilworth, L.; Facey, A.; Omoruyi, F. Diabetes Mellitus and Its Metabolic Complications: The Role of Adipose Tissues. Int. J. Mol. Sci. 2021, 2, 7644. [Google Scholar] [CrossRef] [PubMed]
- Klip, A.; McGraw, T.E.; James, D.E. Thirty sweet years of GLUT4. J. Biol. Chem. 2019, 294, 11369–11381. [Google Scholar] [CrossRef] [Green Version]
- Maria, Z.; Campolo, A.R.; Lacombe, V.A. Diabetes Alters the Expression and Translocation of the Insulin-Sensitive Glucose Transporters 4 and 8 in the Atria. PLoS ONE 2015, 10, e0146033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hediger, M.A.; Rhoads, D.B. Molecular physiology of sodium-glucose cotransporters. Physiol. Rev. 1994, 74, 993–1026. [Google Scholar] [CrossRef]
- Banerjee, S.K.; McGaffin, K.R.; Pastor-Soler, N.M.; Ahmad, F. SGLT1 is a novel cardiac glucose transporter that is perturbed in disease states. Cardiovasc. Res. 2009, 84, 111–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Y.C.; Zheng, C.M.; Yen, T.H.; Lu, K.C. Molecular Mechanisms of SGLT2 Inhibitor on Cardiorenal Protection. Int. J. Mol. Sci. 2020, 21, 7833. [Google Scholar] [CrossRef]
- Unno, K.; Yamamoto, H.; Toda, M.; Hagiwara, S.; Iguchi, K.; Hoshino, M.; Takabayashi, F.; Hasegawa-Ishii, S.; Shimada, A.; Hosokawa, M.; et al. Novel frame-shift mutation in Slc5a2 encoding SGLT2 in a strain of senescence-accelerated mouse SAMP10. Biochem. Biophys. Res. Commun. 2014, 454, 89–94. [Google Scholar] [CrossRef]
- Cipriani, C.; Colangelo, L.; Santori, R.; Renella, M.; Mastrantonio, M.; Minisola, S.; Pepe, J. The Interplay Between Bone and Glucose Metabolism. Front. Endocrinol. 2020, 11, 122. [Google Scholar] [CrossRef] [Green Version]
- Bray, J.J.H.; Foster-Davies, H.; Stephens, J.W. A systematic review examining the effects of sodium-glucose cotransporter-2 inhibitors (SGLT2is) on biomarkers of inflammation and oxidative stress. Diabetes Res. Clin. Pr. 2020, 168, 108368. [Google Scholar] [CrossRef]
- Gnudi, L.; Viberti, G.C.; Raij, L.; Rodriguez, V.; Burt, D.; Cortes, P.; Hartley, B.; Thomas, S.; Maestrini, S.; Gruden, G. GLUT-1 Overexpression: Link Between Hemodynamic and Metabolic Factors in Glomerular Injury? Hypertension 2003, 42, 19–24. [Google Scholar] [CrossRef] [Green Version]
- De Cingolani, G.E.C.; Caldiz, C.I. Insulin resistance and GLUT-4 glucose transporter in adipocytes from hypertensive rats. Metabolism 2004, 53, 382–387. [Google Scholar] [CrossRef] [PubMed]
- Na, K.J.; Choi, H.; Oh, H.R.; Kim, Y.H.; Lee, S.B.; Jung, Y.J.; Koh, J.; Park, S.; Lee, H.J.; Jeon, Y.K.; et al. Reciprocal change in Glucose metabolism of Cancer and Immune Cells mediated by different Glucose Transporters predicts Immunotherapy response. Theranostics 2020, 10, 9579–9590. [Google Scholar] [CrossRef] [PubMed]
- Navale, A.M.; Paranjape, A.N. Glucose transporters: Physiological and pathological roles. Biophys. Rev. 2016, 8, 5–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, J.; Mei, J.; Zhang, M.Z.; Yuan, F.; Li, S.Z.; Yu, G.R.; Chen, L.H.; Tan, Q.; Xian, G.C.J. Clinicopathological significance of glucose transporter protein-1 overexpression in human osteosarcoma. Oncol. Lett. 2017, 14, 2439–2445. [Google Scholar] [CrossRef] [Green Version]
- Barron, C.C.; Bilan, P.J.; Tsakiridis, T.; Tsiani, E. Facilitative glucose transporters: Implications for cancer detection, prognosis and treatment. Metabolism 2016, 65, 124–139. [Google Scholar] [CrossRef]
- Echeverría, C.; Nualart, F.; Ferrada, L.; Smith, G.J.; Godoy, A.S. Hexose Transporters in Cancer: From Multifunctionality to Diagnosis and Therapy. Trends Endocrinol. Metab. 2021, 32, 198–211. [Google Scholar] [CrossRef]
- Beylerli, O.; Sufianova, G.; Shumadalova, A.; Zhang, D.; Gareev, I. MicroRNAs-mediated regulation of glucose transporter (GLUT) expression in glioblastoma. Non-Coding RNA Res. 2022, 7, 205–211. [Google Scholar] [CrossRef]
- López-Ramírez, G.; Méndez-Flores, O.G. El transporte de la glucosa en las células gliales del cerebro obeso. Glucose transport in glial cells from obese brain. Revista eNeurobiología 2022, 13, 011022. [Google Scholar]
- Zambrano, A.; Molt, M.; Uribe, E.; Salas, M. Glut 1 in Cancer Cells and the Inhibitory Action of Resveratrol as A Potential Therapeutic Strategy. Int. J. Mol. Sci. 2019, 20, 3374. [Google Scholar] [CrossRef] [Green Version]
- Chai, Y.J.; Yi, J.W.; Oh, S.W.; Kim, Y.A.; Yi, K.H.; Kim, J.H.; Lee, K.E. Upregulation of SLC2 (GLUT) family genes is related to poor survival outcomes in papillary thyroid carcinoma: Analysis of data from The Cancer Genome Atlas. Surgery 2017, 161, 188–194. [Google Scholar] [CrossRef]
- Feng, Z.; Ou, Y.; Hao, L. The roles of glycolysis in osteosarcoma. Front. Pharmacol. 2022, 13, 950886. [Google Scholar] [CrossRef]
- Baraban, S.C. Emerging epilepsy models: Insights from mice, flies, worms and fish. Opin Neurol. 2007, 20, 164–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuse, T.; Mizuma, H.; Hirose, Y.; Kushida, T.; Yamada, I.; Miura, I.; Masuya, H.; Funato, H.; Yanagisawa, M.; Onoe, H.; et al. A new mouse model of GLUT1 deficiency syndrome exhibits abnormal sleep-wake patterns and alterations of glucose kinetics in the brain. Dis. Model. Mech. 2019, 12, dmm038828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, J.; Johannesen, K.M.; Ek, J.; Tang, S.; Marini, C.; Blichfeldt, S.; Kibæk, M.; von Spiczak, S.; Weckhuysen, S.; Frangu, M.; et al. The role of SLC2A1 mutations in myoclonic astatic epilepsy and absence epilepsy, and the estimated frequency of GLUT1 deficiency syndrome. Epilepsia 2015, 56, e203–e208. [Google Scholar] [CrossRef] [PubMed]
- Bourque, D.K.; Cordeiro, D.; Nimmo, G.A.M.; Kobayashi, J.; Mercimek-Andrews, S. Phenotypic and Genotypic Spectrum of Glucose Transporter-1 Deficiency Syndrome. Can. J. Neurol. Sci. 2021, 48, 826–830. [Google Scholar] [CrossRef]
- Shah, K.; DeSilva, S.; Abbruscato, T. The Role of Glucose Transporters in Brain Disease: Diabetes and Alzheimer’s Disease. Int. J. Mol. Sci. 2012, 13, 12629–12655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, Y.; Varma, V.R.; Varma, S.; Casanova, R.; Dammer, E.; Pletnikova, O.; Chia, C.W.; Egan, J.M.; Ferrucci, L.; Troncoso, J.; et al. Evidence for brain glucose dysregulation in Alzheimer’s disease. Alzheimer’s Dement. 2018, 14, 318–329. [Google Scholar] [CrossRef]
- Santos, J.M.; Benite-Ribeiro, S.A.; Queiroz, G.; Duarte, J.A. The effect of age on glucose uptake and GLUT1 and GLUT4 expression in rat skeletal muscle. Cell Biochem. Funct. 2012, 30, 191–197. [Google Scholar] [CrossRef]
- Wang, Q.; Duan, L.; Li, X.; Wang, Y.; Guo, W.; Guan, F.; Ma, S. Glucose Metabolism, Neural Cell Senescence and Alzheimer’s Disease. Int. J. Mol. Sci. 2022, 23, 4351. [Google Scholar] [CrossRef]
- Buysse, D.J. Insomnia. JAMA 2013, 309, 706–716. [Google Scholar] [CrossRef] [Green Version]
- Kordestani Moghadam, P.; Nasehi, M.; Khodagholi, F.; Zarrindast, M.R. Vulnerability of Left Amygdala to Total Sleep Deprivation and Reversed Circadian Rhythm in Molecular Level: Glut1 as a Metabolic Biomarker. Galen Med. J. 2019, 8, e970. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Liu, Y.; Wang, X. TNF-α promotes insulin resistance in obstructive sleep apnea-hypopnea syndrome. Exp. Ther. Med. 2021, 21, 568. [Google Scholar] [CrossRef]
- Lee, S.Y.; Abel, E.D.; Long, F. Glucose metabolism induced by Bmp signaling is essential for murine skeletal development. Nat. Commun. 2018, 9, 4831. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.-Y.; Cheng, Y.-Z.; Yang, S.-L.; An, M.; Zhang, H.; Chen, H.; Yang, L. Dapagliflozin Attenuates Hyperglycemia Related Osteoporosis in ZDF Rats by Alleviating Hypercalciuria. Front. Endocrinol. 2019, 10, 700. [Google Scholar] [CrossRef] [Green Version]
- Havukainen, S.; Pujol-Giménez, J.; Valkonen, M.; Westerholm-Parvinen, A.; Hediger, M.A.; Landowski, C.P. Electrophysiological characterization of a diverse group of sugar transporters from Trichoderma reesei. Sci. Rep. 2021, 11, 14678. [Google Scholar] [CrossRef]
- Yoshino, K.; Yamamoto, K.; Hara, K.; Sonoda, M.; Yamamoto, Y.; Sakamoto, K. The conservation of polyol transporter proteins and their involvement in lichenized Ascomycota. Fungal Biol. 2019, 123, 318–329. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Frommer, W.B. Structure and function of SemiSWEET and SWEET sugar transporters. Trend Biochem. Sci. 2015, 40, P480–P486. [Google Scholar] [CrossRef] [Green Version]
- Jeena, G.S.; Kumar, S.; Shukla, R.K. Structure, evolution and diverse physiological roles of SWEET sugar transporters in plants. Plant Mol. Biol. 2019, 100, 351–365. [Google Scholar] [CrossRef]
- Ancey, P.B.; Contat, C.; Meylan, E. Glucose transporters in cancer—From tumor cells to the tumor microenvironment. FEBS J. 2018, 285, 2926–2943. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Q.; Hou, B.H.; Lalonde, S.; Takanaga, H.; Hartung, M.L.; Qu, X.Q.; Guo, W.J.; Kim, J.G.; Underwood, W.; Chaudhuri, B.; et al. Sugar transporters for intercellular exchange and nutrition of Pathogens. Nature 2010, 468, 527–532. [Google Scholar] [CrossRef] [Green Version]
- Xuan, Y.H.; Hu, Y.B.; Chen, L.Q.; Sosso, D.; Ducat, D.C.; Hou, B.H.; Frommer, W.B. Functional role of oligomerization for bacterial and plant SWEET sugar transporter family. Proc. Natl. Acad. Sci. USA 2013, 110, E3685–E3694. [Google Scholar] [CrossRef] [Green Version]
- Doidy, J.; Vidal, U.; Lemoine, R. Sugar transporters in Fabaceae, featuring SUT MST and SWEET families of the model plant Medicago truncatula and the agricultural crop Pisum sativum. PLoS ONE 2019, 14, e0223173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahmidi, N.A.; Courty, P.E.; Brulé, D.; Chatagnier, O.; Arnould, C.; Doidy, J.; Berta, G.; Lingua, G.; Wipf, D.; Bonneau, L. Sugar exchanges in arbuscular mycorrhiza: RiMST5 and RiMST6, two novel Rhizophagus irregularis monosaccharide transporters, are involved in both sugar uptake from the soil and from the plant partner. Plant Physiol. Biochem. 2016, 107, 354–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linka, N.; Weber, A.P.M. Intracellular Metabolite Transporters in Plants. Mol. Plant 2010, 3, 21–53. [Google Scholar] [CrossRef]
- Julius, B.T.; Leach, K.A.; Tran, T.M.; Mertz, R.A.; Braun, D.M. Sugar Transporters in Plants: New Insights and Discoveries. Plant Cell Physiol. 2017, 58, 1442–1460. [Google Scholar] [CrossRef] [Green Version]
- Slewinski, T.L. Diverse Functional Roles of Monosaccharide Transporters and their Homologs in Vascular Plants: A Physiological Perspective. Mol. Plant 2011, 4, 641–662. [Google Scholar] [CrossRef] [PubMed]
- Williams, L.E.; Lemoine, R.; Sauer, N. Sugar transporters in higher plants—A diversity of roles and complex regulation. Trends Plant Sci. 2000, 5, S1360–S1385. [Google Scholar] [CrossRef]
- Doidy, J.; Grace, E.; Kühn, C.; Simon-Plas, F.; Casieri, L.; Wipf, D. Sugar transporters in plants and in their interactions with fungi. Trends Plant Sci. 2012, 17, 413–422. [Google Scholar] [CrossRef]
- Chen, L.Q.; Cheung, L.S.; Feng, L.; Tanner, W.; Frommer, W.B. Transport of Sugars. Ann. Rev. Biochem. 2015, 84, 865–894. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.J.; Song, S.; Mason, K.; Pinkett, H.W. Selective substrate uptake: The role of ATP-binding cassette (ABC) importers in pathogenesis. Biochim. Biophys. Acta 2018, 1860, 868–877. [Google Scholar] [CrossRef]
- Thomas, C.; Aller, S.G.; Beis, K.; Carpenter, E.P.; Chang, G.; Chen, L.; Dassa, E.; Dean, M.; Van Hoa, F.D.; Ekiert, D.; et al. Structural and functional diversity calls for a new classification of ABC transporters. FEBS Lett. 2020, 594, 3767–3775. [Google Scholar] [CrossRef]
- Perez, C.; Gerber, S.; Boilevin, J.; Bucher, M.; Darbre, T.; Aebi, M.; Reymond, J.L.; Locher, K.P. Structure and mechanism of an active lipid-linked oligosaccharide flippase. Nature 2015, 524, 433–438. [Google Scholar] [CrossRef] [Green Version]
- Davidson, A.L.; Chen, J. ATP-binding cassette transporters in bacteria. Annu. Rev. Biochem. 2004, 73, 241–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitavska, O.; Wieczorek, H. The SLC45 gene family of putative sugar transporters. Mol. Asp. Med. 2013, 34, 655–660. [Google Scholar] [CrossRef]
- Hadley, B.; Litfin, T.; Day, C.J.; Haselhorst, T.; Zhou, Y.; Tiralongo, J. Nucleotide Sugar Transporter SLC35 Family Structure and Function. Comput. Struct. Biotechnol. J. 2019, 17, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Motlhalamme, T.Y.; Zhou, N.; Gamero, A.; Mehlomakulu, N.N.; Jolly, N.; Albertyn-Pohl, C.; Setati, M.E. Origin and Evolution of Yeasts. In CHAPTER 1, Yeasts: From Nature to Bioprocesses; Bentham Science Publishers: Oak Park, IL, USA, 2022; pp. 1–26. ISBN 9789815051063. [Google Scholar]
- Pliszka, M.; Szablewski, L. Glucose Transporters as a Target for Anticancer Therapy. Cancers 2021, 13, 4184. [Google Scholar] [CrossRef] [PubMed]
- Zezina, E.; Sercan-Alp, O.; Herrmann, M.; Biesemann, N. Glucose transporter 1 in rheumatoid arthritis and autoimmunity. Wiley Interdiscip. Rev. Syst. Biol. Med. 2020, 12, e1483. [Google Scholar] [CrossRef] [PubMed]
- Varghese, E.; Samuel, S.M.; Líšková, A.; Samec, M.; Kubatka, P.; Büsselberg, D. Targeting Glucose Metabolism to Overcome Resistance to Anticancer Chemotherapy in Breast Cancer. Cancers 2020, 12, 2252. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Transporter | Transports | Km | Localization | Ref. |
| SGLT1 (SLC5A1) | 1 Glucose/1 Galactose:2 Na+ αMG | 0.5/1 mM | intestine, trachea, kidney, heart, and colon | [7,8] |
| SGLT2 (SLC5A2) | 1 Glucose:1 Na+ αMG | 5 mM | heart and kidney cortex | [7,8] |
| SGLT3 (SLC5A4) | 1 Glucose:2 Na+ imino sugars αMG | 20 mM | glucose sensor in the enteric nervous system, uterus, lung, brain and autonomic nervous system, spleen, thyroid, kidney, and trachea | [6,7,8] |
| SGLT4 (SLC5A9) | Mannose, fructose, and glucose αMG | 2 mM | small intestine, brain, and kidney | [6,8] |
| SGLT5 (SLC5A10) | Mannose, fructose, and glucose αMG | ND | kidney cortex | [6] |
| SMIT1 (SLC5A3) | Myo-inositol:Na+, 1 Glucose/1 Galactose: Na+ αMG | 0.05 mM | heart, brain, and kidney | [9] |
| SGLT6 or SMIT2 (SLC5A11) | Myo-inositol: Na+ αMG, xylose and glucose | 0.3 mM | spinal cord, kidney, and brain | [6,8] |
| Transporter | Transports | Km | Localization | Ref. |
|---|---|---|---|---|
| CLASS I | ||||
| GLUT1 (SLC2A1) | Glucose, mannose, glucosamine, DHA, fucose, and galactose | 2 mM | erythrocytes, brain endothelial cells, neurons, kidney, heart, lymphocytes, mainly all cells | [12,13,14,15,16] |
| GLUT2 (SLC2A2) | Glucose, mannose, glucosamine, fructose, and galactose | 17 mM | β-pancreatic cells, liver, brain, kidney, small intestine | [8,12,13] |
| GLUT3 (SLC2A3) | Glucose, mannose, DHA, xylose, and galactose | 2 mM | central nervous system, placenta, liver, kidney, heart, lymphocytes | [12,13,14] |
| GLUT4 (SLC2A4) | Glucose, DHA, and glucosamine | 5 mM | insulin-sensitive tissues, heart, central nervous system, heart, lymphocytes | [8,12,13,14,16] |
| GLUT14 (SLC2A14) | Glucose | ND | testis | [13] |
| CLASS II | ||||
| GLUT5 (SLC2A5) | Fructose | 10 mM | small intestine, testis, kidney, central nervous system | [8,12,13] |
| GLUT7 (SLC2A7) | Glucose and fructose | 0.3 mM and 0.06 mM | small intestine, colon, testis, prostate | [12,13] |
| GLUT9 (SLC2A9) | Fructose and uric acid | urate ~0.6 mM | kidney, liver, small intestine, placenta, lungs, leukocytes | [13,17] |
| GLUT11 (SLC2A11) | Fructose and glucose | Fructose high affinity, and glucose low affinity | heart, skeletal muscle, kidney, adipose tissue, placenta, pancreas | [13] |
| CLASS III | ||||
| GLUT6 (SLC2A6) | Glucose | 5 mM | brain, spleen, leukocytes, lysosomal membranes | [10,12,13] |
| GLUT8 (SLC2A8) | Glucose, fructose, DHA, and galactose | 2 mM | testis, insulin-sensitive tissues | [13,14,16] |
| GLUT10 (SLC2A10) | Glucose and galactose | 0.3 mM | liver, heart, pancreas, brain, ER glucose transporter | [8,16,18] |
| GLUT12 (SLC2A12) | Glucose, galactose, and fructose | Glucose high affinity | skeletal muscle, central nervous system, adipose tissue, small intestine, heart | [8,12,16] |
| HMIT (GLUT13) (SLC2A13) | Mio-Inositol H+- coupled | 100 μM | brain | [13] |
| Tissue | Transporter Modified in Malignancy | Ref. |
|---|---|---|
| Nervous system | ||
| Astrocytes | 1, 3, 4, 12 | [66,67,68] |
| Brain | 1, 3 | [66,68] |
| Brain–blood barrier | 1 | [69] |
| Glia | 1, 3, 4, 5 | [66,67,68,70] |
| Meninges | 1 | [70] |
| Oligodendrocytes | 12 | [66,69] |
| Retina | 1 | [70] |
| Digestive system | ||
| Colon | 1, 2, 4, 5, 12 | [66,68,70] |
| Esophagus | 1 | [70] |
| Larynx | 1, 3 | [66,70] |
| Liver | 1, 2, 5, 9 | [68,70] |
| Mouth | 1, 3 | [68,70] |
| Pancreas | 1, 2, 4, 5, 6 | [66,67,68,70] |
| Rectum | 1, 2, 5 | [68,70] |
| Stomach | 1, 2, 3, 4, 6, 10 | [66,67,68] |
| Tongue | 1 | [70] |
| Reproductive system | ||
| Breast | 1, 2, 3, 4, 5, 6, 12, HMIT | [66,67,68] |
| Cervix | 1 | [66,67,68] |
| Endometrium | 4, 6, 8 | [67,68] |
| Ovary | 1, 3, | [66,67,68] |
| Prostate | 1, 3, 5, 7, 9, 11, 12 | [66,67,68] |
| Testis | 5, 6, 9, 14 | [66,67,68,70] |
| Uterus | 1, 6 | [66,70] |
| Respiratory system | ||
| Lung | 1, 3, 5, 9, 12 | [66,70] |
| Nasopharynx | 1, | [68,70] |
| Urinary system | ||
| Bladder | 1, 3 | [66] |
| Kidney | 1, 5, 9, 13, HMIT | [66,68,70] |
| Urethra | 1 | [70] |
| Immune system | ||
| Plasmatic cells | 4, 7, 8, 10, 11 | [66,67] |
| Lymphoid tissue | 1, 4, 5, 8 | [66,70] |
| Dermal system | ||
| Melanocytes | 1, 3 | [68,70] |
| Skin | 1, 3, 9 | [66,68,70] |
| Glandular system | ||
| Thyroid | 1, 2, 3, 4, 9, 14 | [66,70,71] |
| Muscular system | ||
| Heart | 1, 9 | [68] |
| Muscle | 1, 12 | [68] |
| Skeletal system | ||
| Bone | 1, 3 | [72] |
| Transporter | Transports | Function | Refs |
|---|---|---|---|
| Clade I SWEETs | Hexoses | ||
| SWEET1 (RAG1AP1) (SLC50A1) | Glucose | β-cells, pathogens, and symbionts | [87,88] |
| SWEET2 (SLC50A2) | 2-Deoxyglucose | Plants | |
| SWEET3 (SLC50A3) | 2-Deoxyglucose | ||
| Clade II SWEETs | Hexoses | ||
| SWEET4 (SLC50A4) | Glucose | ||
| SWEET5 (SLC50A5) | Glucose | ||
| SWEET6 (SLC50A6) | Glucose | ||
| SWEET7 (SLC50A7) | Glucose | ||
| SWEET8 (SLC50A8) | Glucose | ||
| Clade III SWEETs | Sucrose | ||
| SWEET9 (SLC50A9) | Sucrose | ||
| SWEET10 (SLC50A10) | Sucrose | ||
| SWEET11 (SLC50A11) | Sucrose | ||
| SWEET12 (SLC50A12) | Sucrose | ||
| SWEET13 (SLC50A13) | Sucrose | ||
| SWEET14 (SLC50A14) | Sucrose | ||
| SWEET15 (SLC50A15) | Sucrose | ||
| Clade IV SWEETs | Fructose | ||
| SWEET16 (SLC50A16) | Glucose, sucrose, fructose | ||
| SWEET17 (SLC50A17) | Fructose | ||
| SemiSWEETS | |||
| BjSemiSWEET | Sucrose | Bacteria | |
| LbSemiSWEET | Glucose | ||
| VsSemiSWEET | ND | ||
| TySemiSWEET | ND | ||
| EcSemiSWEET | Sucrose | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carbó, R.; Rodríguez, E. Relevance of Sugar Transport across the Cell Membrane. Int. J. Mol. Sci. 2023, 24, 6085. https://doi.org/10.3390/ijms24076085
Carbó R, Rodríguez E. Relevance of Sugar Transport across the Cell Membrane. International Journal of Molecular Sciences. 2023; 24(7):6085. https://doi.org/10.3390/ijms24076085
Chicago/Turabian StyleCarbó, Roxana, and Emma Rodríguez. 2023. "Relevance of Sugar Transport across the Cell Membrane" International Journal of Molecular Sciences 24, no. 7: 6085. https://doi.org/10.3390/ijms24076085