
Influence of Scaffold Microarchitecture on Angiogenesis and Regulation of Cell Differentiation during the Early Phase of Bone Healing: A Transcriptomics and Histological Analysis
 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Gene Ontology
2.2. Pathway Analysis in Fil050G (Highly Osteoconductive) Scaffolds in the Early Phase of Bone Healing
2.2.1. Angiogenesis
2.2.2. Regulation of Cell Differentiation
2.2.3. Bone Formation
2.3. Analysis of Relative Gene Expression by qPCR in the Early Phase of Bone Healing
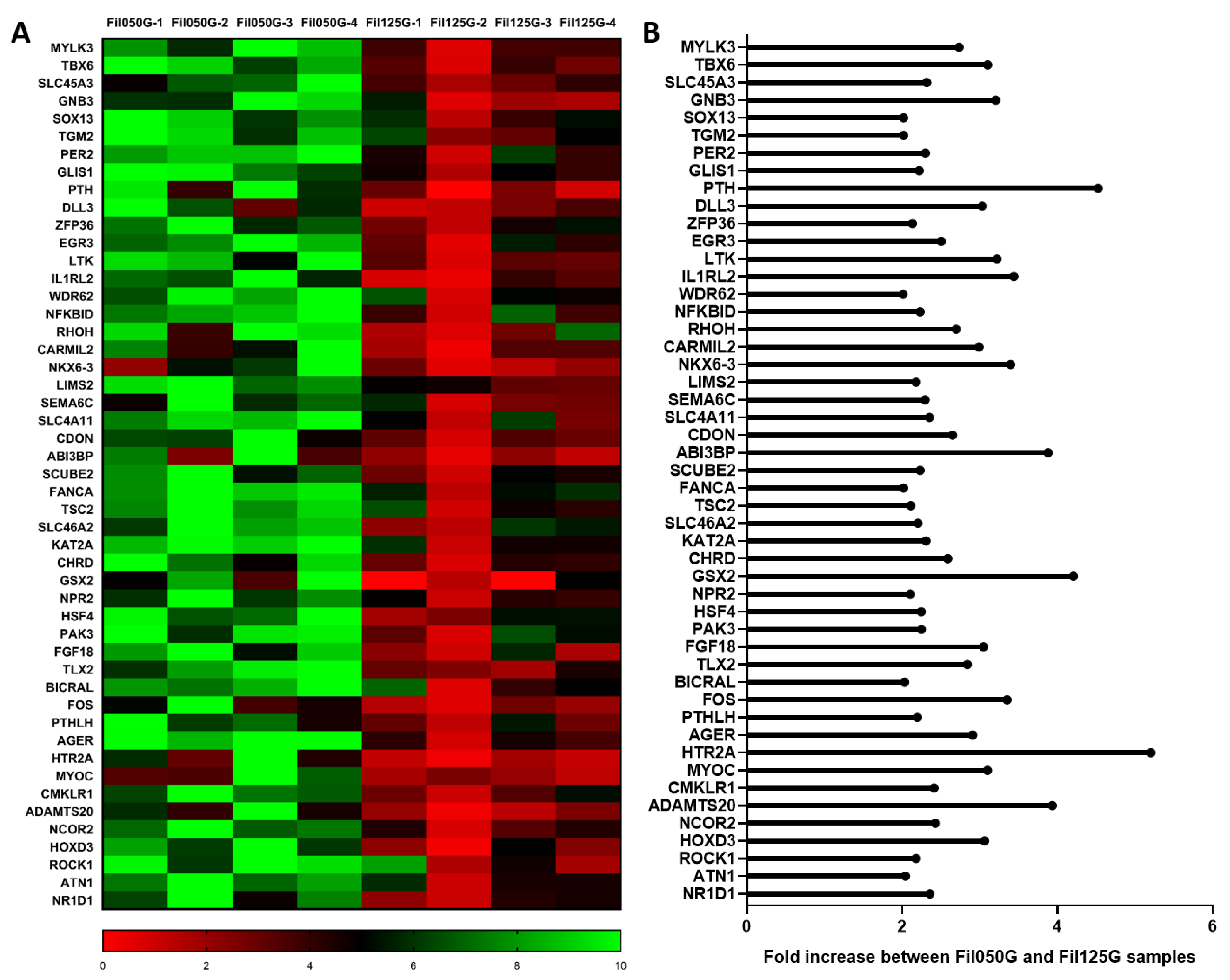
2.3.1. Cell Differentiation/Mesenchymal Stromal Cells Differentiation
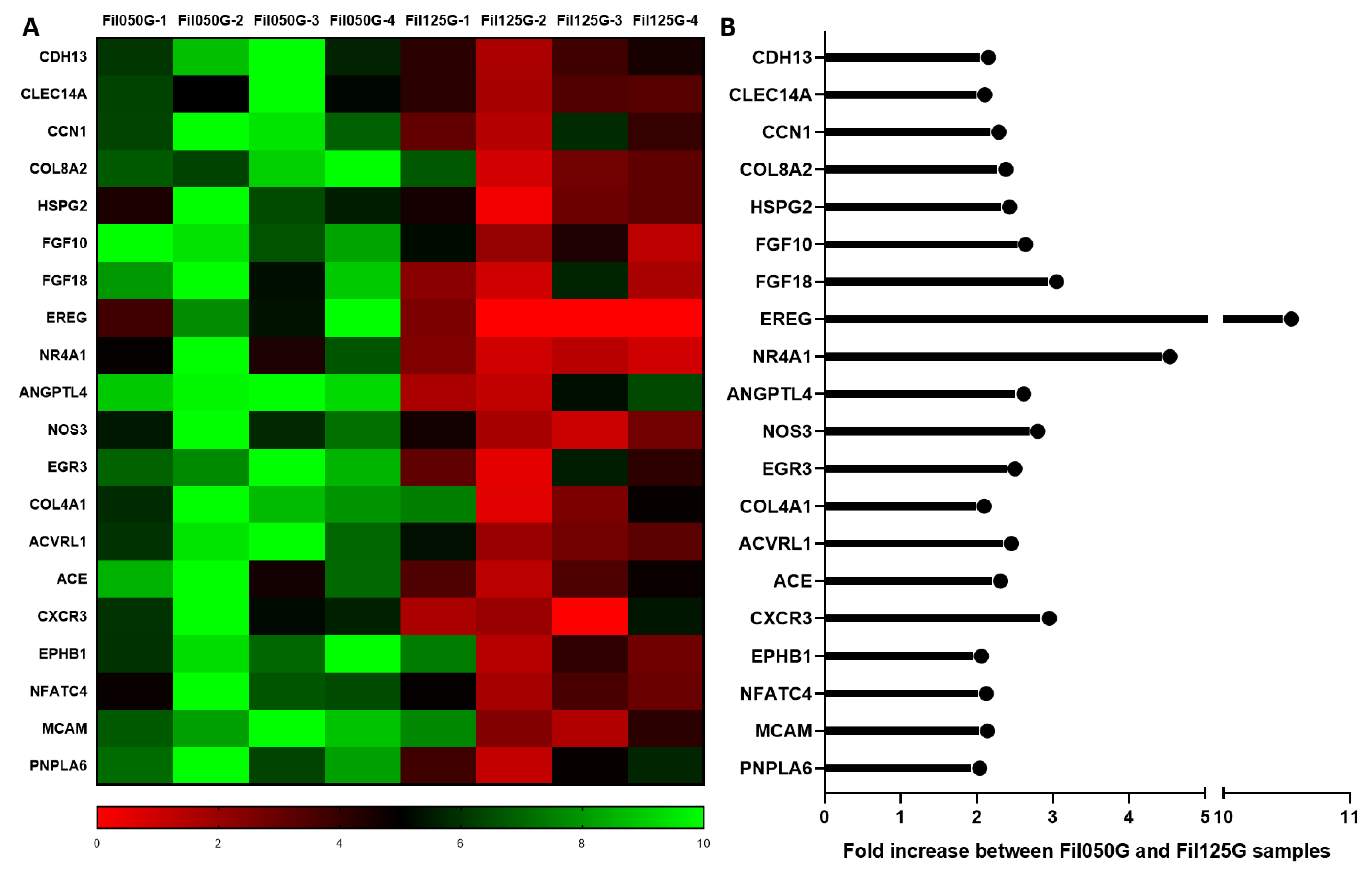
2.3.2. Angiogenesis
2.3.3. Highly Expressed in Fil125G
2.3.4. Bone Formation
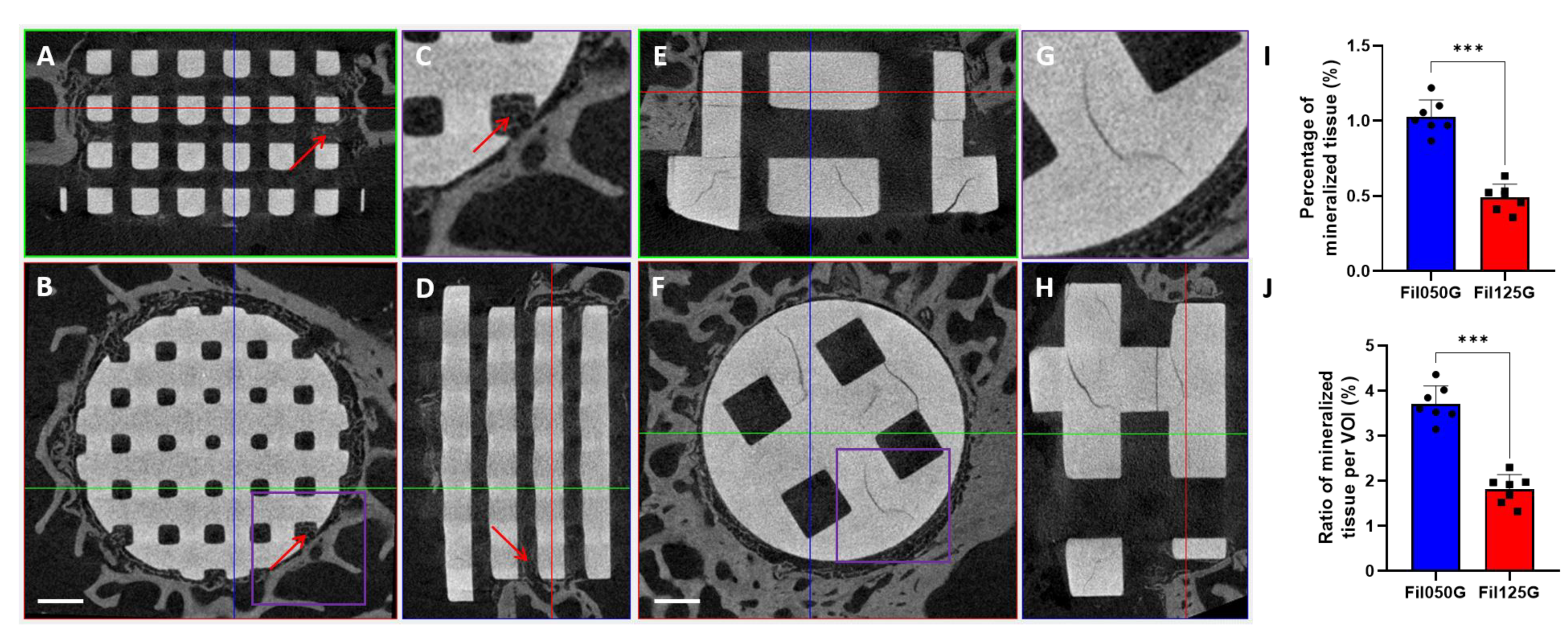
2.4. Mineralized Tissue after 10 Days of Implantation
2.5. Blood Vessel Analysis after 4 Weeks of Implantation
Microvasculature and Capillaries
3. Discussion
4. Materials and Methods
4.1. Scaffold Production
4.2. Surgical Procedure
4.3. RNA Extraction and Purification
4.4. RNA Sequencing
4.5. Gene Ontology (GO) Enrichment Analysis
4.6. Quantitative Real-Time Polymerase Chain Reaction
4.7. Histomorphometry by µCT
4.8. Assessment of In Vivo Vascularization by Immunohistofluorescence
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dimitriou, R.; Jones, E.; McGonagle, D.; Giannoudis, P.V. Bone regeneration: Current concepts and future directions. BMC Med. 2011, 9, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt-Bleek, K.; Kwee, B.J.; Mooney, D.J.; Duda, G.N. Boon and Bane of Inflammation in Bone Tissue Regeneration and Its Link with Angiogenesis. Tissue Eng. Part B Rev. 2015, 21, 354–364. [Google Scholar] [CrossRef] [Green Version]
- Kolar, P.; Schmidt-Bleek, K.; Schell, H.; Gaber, T.; Toben, D.; Schmidmaier, G.; Perka, C.; Buttgereit, F.; Duda, G.N. The early fracture hematoma and its potential role in fracture healing. Tissue Eng. Part B Rev. 2010, 16, 427–434. [Google Scholar] [CrossRef]
- Einhorn, T.A.; Majeska, R.J.; Rush, E.B.; Levine, P.M.; Horowitz, M.C. The expression of cytokine activity by fracture callus. J. Bone Miner. Res. 1995, 10, 1272–1281. [Google Scholar] [CrossRef] [PubMed]
- Kon, T.; Cho, T.J.; Aizawa, T.; Yamazaki, M.; Nooh, N.; Graves, D.; Gerstenfeld, L.C.; Einhorn, T.A. Expression of osteoprotegerin, receptor activator of NF-kappaB ligand (osteoprotegerin ligand) and related proinflammatory cytokines during fracture healing. J. Bone Miner. Res. 2001, 16, 1004–1014. [Google Scholar] [CrossRef] [PubMed]
- Holmes, D. Non-union bone fracture: A quicker fix. Nature 2017, 550, S193. [Google Scholar] [CrossRef] [Green Version]
- Ghayor, C.; Weber, F.E. Osteoconductive Microarchitecture of Bone Substitutes for Bone Regeneration Revisited. Front. Physiol. 2018, 9, 960. [Google Scholar] [CrossRef] [PubMed]
- Menger, M.M.; Laschke, M.W.; Nussler, A.K.; Menger, M.D.; Histing, T. The vascularization paradox of non-union formation. Angiogenesis 2022, 25, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Khalaf, A.T.; Wei, Y.; Wan, J.; Zhu, J.; Peng, Y.; Abdul Kadir, S.Y.; Zainol, J.; Oglah, Z.; Cheng, L.; Shi, Z. Bone Tissue Engineering through 3D Bioprinting of Bioceramic Scaffolds: A Review and Update. Life 2022, 12, 903. [Google Scholar] [CrossRef]
- McKibbin, B. The biology of fracture healing in long bones. J. Bone Jt. Surg. Br. 1978, 60-B, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Huang, E.E.; Zhang, N.; Shen, H.; Li, X.; Maruyama, M.; Utsunomiya, T.; Gao, Q.; Guzman, R.A.; Goodman, S.B. Novel Techniques and Future Perspective for Investigating Critical-Size Bone Defects. Bioengineering 2022, 9, 171. [Google Scholar] [CrossRef] [PubMed]
- Donos, N.; Dereka, X.; Mardas, N. Experimental models for guided bone regeneration in healthy and medically compromised conditions. Periodontol. 2000 2015, 68, 99–121. [Google Scholar] [CrossRef] [PubMed]
- Vajgel, A.; Mardas, N.; Farias, B.C.; Petrie, A.; Cimoes, R.; Donos, N. A systematic review on the critical size defect model. Clin. Oral Implant. Res. 2014, 25, 879–893. [Google Scholar] [CrossRef]
- Gomes, P.S.; Fernandes, M.H. Rodent models in bone-related research: The relevance of calvarial defects in the assessment of bone regeneration strategies. Lab. Anim. 2011, 45, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Bais, M.; McLean, J.; Sebastiani, P.; Young, M.; Wigner, N.; Smith, T.; Kotton, D.N.; Einhorn, T.A.; Gerstenfeld, L.C. Transcriptional analysis of fracture healing and the induction of embryonic stem cell-related genes. PLoS ONE 2009, 4, e5393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Quigg, R.J.; Zhou, J.; Ryaby, J.T.; Wang, H. Early signals for fracture healing. J. Cell. Biochem. 2005, 95, 189–205. [Google Scholar] [CrossRef] [PubMed]
- Rundle, C.H.; Wang, H.; Yu, H.; Chadwick, R.B.; Davis, E.I.; Wergedal, J.E.; Lau, K.H.; Mohan, S.; Ryaby, J.T.; Baylink, D.J. Microarray analysis of gene expression during the inflammation and endochondral bone formation stages of rat femur fracture repair. Bone 2006, 38, 521–529. [Google Scholar] [CrossRef]
- Kelly, N.H.; Schimenti, J.C.; Ross, F.P.; van der Meulen, M.C. Transcriptional profiling of cortical versus cancellous bone from mechanically-loaded murine tibiae reveals differential gene expression. Bone 2016, 86, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Mantila Roosa, S.M.; Liu, Y.; Turner, C.H. Gene expression patterns in bone following mechanical loading. J. Bone Miner. Res. 2011, 26, 100–112. [Google Scholar] [CrossRef] [Green Version]
- McKenzie, J.A.; Bixby, E.C.; Silva, M.J. Differential gene expression from microarray analysis distinguishes woven and lamellar bone formation in the rat ulna following mechanical loading. PLoS ONE 2011, 6, e29328. [Google Scholar] [CrossRef]
- Hadjiargyrou, M.; Lombardo, F.; Zhao, S.; Ahrens, W.; Joo, J.; Ahn, H.; Jurman, M.; White, D.W.; Rubin, C.T. Transcriptional profiling of bone regeneration. Insight into the molecular complexity of wound repair. J. Biol. Chem. 2002, 277, 30177–30182. [Google Scholar] [CrossRef] [Green Version]
- Al-Kattan, R.; Retzepi, M.; Calciolari, E.; Donos, N. Microarray gene expression during early healing of GBR-treated calvarial critical size defects. Clin. Oral Implant. Res. 2017, 28, 1248–1257. [Google Scholar] [CrossRef] [PubMed]
- Bucher, C.H.; Schlundt, C.; Wulsten, D.; Sass, F.A.; Wendler, S.; Ellinghaus, A.; Thiele, T.; Seemann, R.; Willie, B.M.; Volk, H.D.; et al. Experience in the Adaptive Immunity Impacts Bone Homeostasis, Remodeling, and Healing. Front. Immunol. 2019, 10, 797. [Google Scholar] [CrossRef] [Green Version]
- Schmidt-Bleek, K.; Schell, H.; Schulz, N.; Hoff, P.; Perka, C.; Buttgereit, F.; Volk, H.D.; Lienau, J.; Duda, G.N. Inflammatory phase of bone healing initiates the regenerative healing cascade. Cell Tissue Res. 2012, 347, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, J.; Dasen, B.; Frismantiene, A.; Pigeot, S.; Ismail, T.; Schaefer, D.J.; Philippova, M.; Resink, T.J.; Martin, I.; Scherberich, A. T-cadherin Expressing Cells in the Stromal Vascular Fraction of Human Adipose Tissue: Role in Osteogenesis and Angiogenesis. Stem Cells Transl. Med. 2022, 11, 213–229. [Google Scholar] [CrossRef]
- Weber, F.E. Reconsidering Osteoconduction in the Era of Additive Manufacturing. Tissue Eng. Part B Rev. 2019, 25, 375–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanz-Herrera, J.A.; Doblare, M.; Garcia-Aznar, J.M. Scaffold microarchitecture determines internal bone directional growth structure: A numerical study. J. Biomech. 2010, 43, 2480–2486. [Google Scholar] [CrossRef]
- Tadano, S.; Giri, B. X-ray diffraction as a promising tool to characterize bone nanocomposites. Sci. Technol. Adv. Mater. 2011, 12, 064708. [Google Scholar] [CrossRef] [PubMed]
- de Wild, M.; Schumacher, R.; Mayer, K.; Schkommodau, E.; Thoma, D.; Bredell, M.; Kruse Gujer, A.; Gratz, K.W.; Weber, F.E. Bone regeneration by the osteoconductivity of porous titanium implants manufactured by selective laser melting: A histological and micro computed tomography study in the rabbit. Tissue Eng. Part. A 2013, 19, 2645–2654. [Google Scholar] [CrossRef] [Green Version]
- Ghayor, C.; Chen, T.H.; Bhattacharya, I.; Ozcan, M.; Weber, F.E. Microporosities in 3D-Printed Tricalcium-Phosphate-Based Bone Substitutes Enhance Osteoconduction and Affect Osteoclastic Resorption. Int. J. Mol. Sci. 2020, 21, 9270. [Google Scholar] [CrossRef]
- Gong, T.; Xie, J.; Liao, J.; Zhang, T.; Lin, S.; Lin, Y. Nanomaterials and bone regeneration. Bone Res. 2015, 3, 15029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karageorgiou, V.; Kaplan, D. Porosity of 3D biomaterial scaffolds and osteogenesis. Biomaterials 2005, 26, 5474–5491. [Google Scholar] [CrossRef] [PubMed]
- Cornell, C.N.; Lane, J.M. Current understanding of osteoconduction in bone regeneration. Clin. Orthop. Relat. Res. 1998, 355, S267–S273. [Google Scholar] [CrossRef]
- Albrektsson, T.; Johansson, C. Osteoinduction, osteoconduction and osseointegration. Eur. Spine J. 2001, 10 (Suppl. S2), S96–S101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Liu, Y.; Zhang, Y.; Yao, B.; Enhejirigala; Li, Z.; Song, W.; Wang, Y.; Duan, X.; Yuan, X.; et al. Biophysical and Biochemical Cues of Biomaterials Guide Mesenchymal Stem Cell Behaviors. Front. Cell Dev. Biol. 2021, 9, 640388. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, R.; Webb, D. Cell migration. Curr. Biol. 2003, 13, R756–R759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayor, R.; Etienne-Manneville, S. The front and rear of collective cell migration. Nat. Rev. Mol. Cell Biol. 2016, 17, 97–109. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, M.A.; Horwitz, A.R. Integrating adhesion, protrusion, and contraction during cell migration. Cell 2006, 125, 1223–1225. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.; Wu, T.; Xia, Y. Perspective: Aligned arrays of electrospun nanofibers for directing cell migration. APL Mater. 2018, 6, 120902. [Google Scholar] [CrossRef] [Green Version]
- Guerrero, J.; Ghayor, C.; Bhattacharya, I.; Weber, F.E. Osteoconductivity of bone substitutes with filament-based microarchitectures: Influence of directionality, filament dimension, and distance. Int. J. Bioprint 2022, 9, 626. [Google Scholar] [CrossRef]
- Doniger, S.W.; Salomonis, N.; Dahlquist, K.D.; Vranizan, K.; Lawlor, S.C.; Conklin, B.R. MAPPFinder: Using Gene Ontology and GenMAPP to create a global gene-expression profile from microarray data. Genome Biol. 2003, 4, R7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donos, N.; Retzepi, M.; Wall, I.; Hamlet, S.; Ivanovski, S. In vivo gene expression profile of guided bone regeneration associated with a microrough titanium surface. Clin. Oral Implant. Res. 2011, 22, 390–398. [Google Scholar] [CrossRef]
- Ivanovski, S.; Hamlet, S.; Retzepi, M.; Wall, I.; Donos, N. Transcriptional profiling of “guided bone regeneration” in a critical-size calvarial defect. Clin. Oral Implant. Res. 2011, 22, 382–389. [Google Scholar] [CrossRef]
- Kostopoulos, L.; Karring, T.; Uraguchi, R. Formation of jawbone tuberosities by guided tissue regeneration. An experimental study in the rat. Clin. Oral Implant. Res. 1994, 5, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Muire, P.J.; Mangum, L.H.; Wenke, J.C. Time Course of Immune Response and Immunomodulation During Normal and Delayed Healing of Musculoskeletal Wounds. Front. Immunol. 2020, 11, 1056. [Google Scholar] [CrossRef]
- Xia, L.; Lin, K.; Jiang, X.; Xu, Y.; Zhang, M.; Chang, J.; Zhang, Z. Enhanced osteogenesis through nano-structured surface design of macroporous hydroxyapatite bioceramic scaffolds via activation of ERK and p38 MAPK signaling pathways. J. Mater. Chem. B 2013, 1, 5403–5416. [Google Scholar] [CrossRef]
- Malhan, D.; Schmidt-Bleek, K.; Duda, G.N.; El Khassawna, T. Landscape of Well-Coordinated Fracture Healing in a Mouse Model Using Molecular and Cellular Analysis. Int. J. Mol. Sci. 2023, 24, 3569. [Google Scholar] [CrossRef] [PubMed]
- Rapp, A.E.; Bindl, R.; Recknagel, S.; Erbacher, A.; Muller, I.; Schrezenmeier, H.; Ehrnthaller, C.; Gebhard, F.; Ignatius, A. Fracture Healing Is Delayed in Immunodeficient NOD/scid-IL2RIL2Rγcnull Mice. PLoS ONE 2016, 11, e0147465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Xia, H.; Wang, Y.; Peng, C.; Li, Y.; Pan, X. Gene expression of four adhesive proteins in the early healing of bone defect and bone-implant interface. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2006, 2006, 2087–2090. [Google Scholar] [CrossRef]
- Dong, C.H.; Deng, Y.S.; Yang, X.J.; Liu, J.; Liu, R.; Hou, F.Y.; Li, S.S.; Zhen, P. The interplay of transcriptional and post-transcriptional regulation of migration of mesenchymal stem cells during early stages of bone fracture healing. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 5542–5547. [Google Scholar] [CrossRef]
- Liu, F.; Li, G.; Deng, L.; Kuang, B.; Li, X. The roles of FGF10 in vasculogenesis and angiogenesis. Biomed. Res. 2017, 28, 1329–1332. [Google Scholar]
- Ong, L.L.; Li, W.; Oldigs, J.K.; Kaminski, A.; Gerstmayer, B.; Piechaczek, C.; Wagner, W.; Li, R.K.; Ma, N.; Steinhoff, G. Hypoxic/normoxic preconditioning increases endothelial differentiation potential of human bone marrow CD133+ cells. Tissue Eng. Part C Methods 2010, 16, 1069–1081. [Google Scholar] [CrossRef] [PubMed]
- Scholtysek, C.; Ipseiz, N.; Bohm, C.; Krishnacoumar, B.; Stenzel, M.; Czerwinski, T.; Palumbo-Zerr, K.; Rothe, T.; Weidner, D.; Klej, A.; et al. NR4A1 Regulates Motility of Osteoclast Precursors and Serves as Target for the Modulation of Systemic Bone Turnover. J. Bone Miner. Res. 2018, 33, 2035–2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, C.; Lu, M.; Hong, Y.; Mao, C.; Chen, J.; Ren, C.; Lin, M.; Chen, W. Osteogenic and angiogenic profiles of the palatal process of the maxilla and the palatal process of the palatine bone. J. Anat. 2022, 240, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Beheshtizadeh, N.; Asgari, Y.; Nasiri, N.; Farzin, A.; Ghorbani, M.; Lotfibakhshaiesh, N.; Azami, M. A network analysis of angiogenesis/osteogenesis-related growth factors in bone tissue engineering based on in-vitro and in-vivo data: A systems biology approach. Tissue Cell 2021, 72, 101553. [Google Scholar] [CrossRef]
- Khan, K.A.; McMurray, J.L.; Mohammed, F.; Bicknell, R. C-type lectin domain group 14 proteins in vascular biology, cancer and inflammation. FEBS J. 2019, 286, 3299–3332. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Murrell, G.A.; Lin, J.; Gardiner, E.M.; Diwan, A.D. Localization of nitric oxide synthases during fracture healing. J. Bone Miner. Res. 2002, 17, 1470–1477. [Google Scholar] [CrossRef]
- Tual-Chalot, S.; Mahmoud, M.; Allinson, K.R.; Redgrave, R.E.; Zhai, Z.; Oh, S.P.; Fruttiger, M.; Arthur, H.M. Endothelial depletion of Acvrl1 in mice leads to arteriovenous malformations associated with reduced endoglin expression. PLoS ONE 2014, 9, e98646. [Google Scholar] [CrossRef] [Green Version]
- Grosso, A.; Burger, M.G.; Lunger, A.; Schaefer, D.J.; Banfi, A.; Di Maggio, N. It Takes Two to Tango: Coupling of Angiogenesis and Osteogenesis for Bone Regeneration. Front. Bioeng. Biotechnol. 2017, 5, 68. [Google Scholar] [CrossRef] [Green Version]
- Stegen, S.; van Gastel, N.; Carmeliet, G. Bringing new life to damaged bone: The importance of angiogenesis in bone repair and regeneration. Bone 2015, 70, 19–27. [Google Scholar] [CrossRef]
- Hankenson, K.D.; Dishowitz, M.; Gray, C.; Schenker, M. Angiogenesis in bone regeneration. Injury 2011, 42, 556–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klenke, F.M.; Liu, Y.; Yuan, H.; Hunziker, E.B.; Siebenrock, K.A.; Hofstetter, W. Impact of pore size on the vascularization and osseointegration of ceramic bone substitutes in vivo. J. Biomed. Mater. Res. A 2008, 85, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.S.; Wong, A.; Toupadakis, C.A.; Yellowley, C.E. Expression of angiopoietin-like protein 4 at the fracture site: Regulation by hypoxia and osteoblastic differentiation. J. Orthop. Res. 2015, 33, 1364–1373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Hernando, C.; Suarez, Y. ANGPTL4: A multifunctional protein involved in metabolism and vascular homeostasis. Curr. Opin. Hematol. 2020, 27, 206–213. [Google Scholar] [CrossRef]
- Wang, W.; Wang, B. Expression of angiogenesis-related proteins in bone marrow mesenchymal stem cells induced by osteoprotegerin during osteogenic differentiation in rats. Int. Immunopharmacol. 2021, 98, 107821. [Google Scholar] [CrossRef]
- Bliziotes, M. Update in serotonin and bone. J. Clin. Endocrinol. Metab. 2010, 95, 4124–4132. [Google Scholar] [CrossRef] [Green Version]
- Lavoie, B.; Lian, J.B.; Mawe, G.M. Regulation of Bone Metabolism by Serotonin. Adv. Exp. Med. Biol. 2017, 1033, 35–46. [Google Scholar] [CrossRef]
- Yu, B.; Zhao, X.; Yang, C.; Crane, J.; Xian, L.; Lu, W.; Wan, M.; Cao, X. Parathyroid hormone induces differentiation of mesenchymal stromal/stem cells by enhancing bone morphogenetic protein signaling. J. Bone Miner. Res. 2012, 27, 2001–2014. [Google Scholar] [CrossRef] [Green Version]
- Hodgkinson, C.P.; Naidoo, V.; Patti, K.G.; Gomez, J.A.; Schmeckpeper, J.; Zhang, Z.; Davis, B.; Pratt, R.E.; Mirotsou, M.; Dzau, V.J. Abi3bp is a multifunctional autocrine/paracrine factor that regulates mesenchymal stem cell biology. Stem Cells 2013, 31, 1669–1682. [Google Scholar] [CrossRef] [Green Version]
- Muruganandan, S.; Roman, A.A.; Sinal, C.J. Role of chemerin/CMKLR1 signaling in adipogenesis and osteoblastogenesis of bone marrow stem cells. J. Bone Miner. Res. 2010, 25, 222–234. [Google Scholar] [CrossRef]
- Sen, R.; Pezoa, S.A.; Carpio Shull, L.; Hernandez-Lagunas, L.; Niswander, L.A.; Artinger, K.B. Kat2a and Kat2b Acetyltransferase Activity Regulates Craniofacial Cartilage and Bone Differentiation in Zebrafish and Mice. J. Dev. Biol. 2018, 6, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, H.S.; Johnson, T.V.; Tomarev, S.I. Myocilin stimulates osteogenic differentiation of mesenchymal stem cells through mitogen-activated protein kinase signaling. J. Biol. Chem. 2013, 288, 16882–16894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connor, J.P.; Manigrasso, M.B.; Kim, B.D.; Subramanian, S. Fracture healing and lipid mediators. Bonekey Rep. 2014, 3, 517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahney, C.S.; Zondervan, R.L.; Allison, P.; Theologis, A.; Ashley, J.W.; Ahn, J.; Miclau, T.; Marcucio, R.S.; Hankenson, K.D. Cellular biology of fracture healing. J. Orthop. Res. 2019, 37, 35–50. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Patil, S.; Gao, Y.G.; Qian, A. The Bone Extracellular Matrix in Bone Formation and Regeneration. Front. Pharmacol. 2020, 11, 757. [Google Scholar] [CrossRef]
- Turajane, K.; Ji, G.; Chinenov, Y.; Chao, M.; Ayturk, U.; Suhardi, V.J.; Greenblatt, M.B.; Ivashkiv, L.B.; Bostrom, M.P.; Yang, X. RNA-seq Analysis of Peri-Implant Tissue Shows Differences in Immune, Notch, Wnt, and Angiogenesis Pathways in Aged Versus Young Mice. JBMR Plus 2021, 5, e10535. [Google Scholar] [CrossRef]
- Kelly, N.H.; Schimenti, J.C.; Patrick Ross, F.; van der Meulen, M.C. A method for isolating high quality RNA from mouse cortical and cancellous bone. Bone 2014, 68, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- The Gene Ontology Consortium. The Gene Ontology resource: Enriching a GOld mine. Nucleic Acids Res. 2021, 49, D325–D334. [Google Scholar] [CrossRef]
- Mi, H.; Muruganujan, A.; Ebert, D.; Huang, X.; Thomas, P.D. PANTHER version 14: More genomes, a new PANTHER GO-slim and improvements in enrichment analysis tools. Nucleic Acids Res. 2019, 47, D419–D426. [Google Scholar] [CrossRef]
- Klemmer, V.A.; Khera, N.; Siegenthaler, B.M.; Bhattacharya, I.; Weber, F.E.; Ghayor, C. Effect of N-Vinyl-2-Pyrrolidone (NVP), a Bromodomain-Binding Small Chemical, on Osteoblast and Osteoclast Differentiation and Its Potential Application for Bone Regeneration. Int. J. Mol. Sci. 2021, 22, 1052. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biological Pathways Overrepresented in 3 Out of 3 Databases | Biological Pathways Overrepresented in 2 Out of 3 Databases | Biological Pathways Overrepresented in 1 Out of 3 Databases |
|---|---|---|
| Adaptive immune response | Cell motility (Rn, Mm) | Cell surface receptor signaling pathway (Rn) |
| Positive regulation of cell adhesion | Cellular process (Rn, Mm) | Dendrite morphogenesis (Rn) |
| Regulation of cell adhesion | Negative regulation of cell communication (Rn, Mm) | Detection of chemical stimulus (Rn) |
| Cell adhesion | Negative regulation of signaling (Rn, Mm) | Detection of chemical stimulus involved in sensory perception (Rn) |
| Regulation of cell migration | Nervous system development (Rn, Mm) | Detection of chemical stimulus involved in sensory perception of smell (Rn) |
| Cell–matrix adhesion | Positive regulation of cell communication (Rn, Mm) | Detection of stimulus (Rn) |
| Cell–substrate adhesion | Positive regulation of signal transduction (Rn, Mm) | Detection of stimulus involved in sensory perception (Rn) |
| Anatomical structure development | Regulation of intracellular signal transduction (Rn, Mm) | Intracellular receptor signaling pathway (Rn) |
| Anatomical structure formation involved in morphogenesis | Regulation of response to stimulus (Rn, Mm) | Negative regulation of response to stimulus (Rn) |
| Anatomical structure morphogenesis | Sensory perception of chemical stimulus (Rn, Mm) | Positive regulation of cellular metabolic process (Rn) |
| Animal organ development | Sensory perception of smell (Rn, Mm) | Positive regulation of macromolecule metabolic process (Rn) |
| Developmental process | System development (Rn, Mm) | Positive regulation of metabolic process (Rn) |
| Enzyme-linked receptor protein signaling pathway | Cell motility (Rn, Mm) | Positive regulation of nucleobase-containing compound metabolic process (Rn) |
| Multicellular organismal process | Cellular process (Rn, Mm) | Positive regulation of response to stimulus (Rn) |
| Muscle system process | Negative regulation of cell communication (Rn, Mm) | Regulation of cell motility (Rn) |
| Regulation of cell communication | Negative regulation of signaling (Rn, Mm) | Regulation of cell population proliferation (Rn) |
| Regulation of multicellular organismal process | Nervous system development (Rn, Mm) | Regulation of IP-10 production (Rn) |
| Regulation of signal transduction | Positive regulation of cell communication (Rn, Mm) | Cell projection organization (Mm) |
| Regulation of signaling | Positive regulation of signal transduction (Rn, Mm) | Negative regulation of response to stimulus (Mm) |
| Regulation of intracellular signal transduction (Rn, Mm) | Cellular response to endogenous stimulus (Mm) |
| Biological Pathways Overrepresented in 3 Out of 3 Databases | Biological Pathways Overrepresented in 2 Out of 3 Databases | Biological Pathways Overrepresented in 1 Out of 3 Databases |
|---|---|---|
| Circulatory system development | Angiogenesis (Rn, Mm) | Blood circulation (Rn) |
| Phospholipid catabolic process | Cell development (Rn, Mm) | Blood vessel development (Rn) |
| Skeletal muscle cell differentiation | Cell differentiation (Rn, Mm) | Detection of stimulus involved in sensory perception (Rn) |
| Tissue development | Embryonic morphogenesis (Rn, Mm) | Embryo development (Rn) |
| Multicellular organism development (Rn, Mm) | Formation of primary germ layer (Rn) | |
| Positive regulation of biological process (Rn, Mm) | Heart development (Rn) | |
| Positive regulation of cellular process (Rn, Mm) | Mesoderm formation (Rn) | |
| Regulation of cell differentiation (Rn, Mm) | Mesoderm morphogenesis (Rn) | |
| Regulation of developmental process (Rn, Mm) | Muscle tissue development (Rn) | |
| Regulation of MAPK cascade (Rn, Mm) | Positive regulation of signaling (Rn) | |
| Tube morphogenesis (Rn, Mm) | Regulation of biological quality (Rn) | |
| Regulation of cell migration (Rn) | ||
| Regulation of ERK1 and ERK2 cascade (Rn) | ||
| Skeletal muscle tissue development (Rn) | ||
| Tissue morphogenesis (Rn) | ||
| Animal organ morphogenesis (Mm) | ||
| Cell adhesion (Mm) |
| Molecular Functions Overrepresented in 3 Out of 3 Databases | Molecular Functions Overrepresented in 2 Out of 3 Databases | Molecular Functions Overrepresented in 1 Out of 3 Databases |
|---|---|---|
| Phospholipase activity | Calcium-dependent phospholipase A2 activity (Rn, Mm) | Actin binding (Rn) |
| Lipase activity | Phospholipase A2 activity (Rn, Mm) | Ion binding (Mm) |
| External encapsulating structure | Carboxylic ester hydrolase activity (Rn, Mm) | Odorant binding (Mm) |
| Nuclear glucocorticoid receptor binding (Rn, Mm) | ||
| Protein binding (Rn, Mm) | ||
| Binding (Rn, Mm) | ||
| Metal ion binding (Rn, Mm) | ||
| Cation binding (Rn, Mm) | ||
| Olfactory receptor activity (Rn, Mm) |
| Cellular Components Overrepresented in 3 Out of 3 Databases | Cellular Components Overrepresented in 2 Out of 3 Databases | Cellular Components Overrepresented in 1 Out of 3 Databases |
|---|---|---|
| Collagen-containing extracellular matrix | Plasma membrane bounded cell projection (Rn, Mm) | Sarcolemma (Rn) |
| Extracellular matrix | Cell projection (Rn, Mm) | Extracellular region (Rn) |
| External encapsulating structure | Cellular anatomical entity (Rn, Mm) | Ribonucleoprotein complex (Rn) |
| Caveola (Rn) | ||
| Cell periphery (Mm) |
| Fold Change | Symbol | Full Name |
|---|---|---|
| 3.047949166 | FGF18 | Fibroblast growth factor 18 |
| 3.097075048 | MYOC | Myocilin |
| 2.23006642 | SCUBE2 | Signal peptide, CUB and EGF-like domain-containing protein 2 |
| 2.103939633 | NPR2 | Atrial natriuretic peptide receptor 2 |
| 2.017002613 | TGM2 | Protein-glutamine gamma-glutamyltransferase 2 |
| 2.288859577 | CCN1 | CCN family member 1 |
| 2.194750496 | PTHLH | Parathyroid hormone-related protein |
| 2.219050295 | GLIS1 | Zinc finger protein GLIS1 |
| 4.521904971 | PTH | Parathyroid hormone |
| 2.429593561 | HSPG2 | Basement membrane-specific heparan sulfate proteoglycan core protein |
| 2.584840247 | CHRD | Chordin |
| 2.802297373 | NOS3 | Nitric oxide synthase, endothelial |
| Short Name | Full Name | Ensembl Number; NCBI Entrez Gene Number | BioRad Unique Assay ID |
|---|---|---|---|
| GAPDH | Glyceraldehyde-3-Phosphate Dehydrogenase | ENSOCUG00000025023; 100009074 | qOcuCED0019227 |
| 18S | Ribosomal Protein S18 | ENSOCUG00000025023; 100009074 | qOcuCED0018100 |
| ABI3BP | ABI Family Member 3 Binding Protein | ENSOCUG00000014699; 100349921 | qOcuCID0005404 |
| CMKLR1 | Chemerin Chemokine-Like Receptor 1 | ENSOCUG00000005336; 100358278 | qOcuCED0007914 |
| KAT2A | Lysine Acetyltransferase 2A | ENSOCUG00000012259; 100355497 | qOcuCED0011401 |
| MYOC | Myocilin | ENSOCUG00000013285; 100347774 | qOcuCED0012306 |
| TSC2 | TSC Complex Subunit 2 | ENSOCUG00000015957; 100008897 | qOcuCID0001468 |
| FGF10 | Fibroblast Growth Factor 10 | ENSOCUG00000012834; 100351640 | qOcuCED0010180 |
| ANGPTL4 | Angiopoietin-Like 4 | ENSOCUG00000008499; 100342802 | qOcuCED0008856 |
| MT-ND4L | Mitochondrially Encoded NADH:Ubiquinone Oxidoreductase Core Subunit 4L | ENSOCUG00000016603; 100339191 | qOcuCED0019933 |
| TSPO2 | Translocator Protein 2 | ENSOCUG00000029107; 808222 | qOcuCED0019792 |
| ALPL | Alkaline Phosphatase, Biomineralization Associated | ENSOCUG00000027644; 100347991 | qOcuCED0012587 |
| COL1A1 | Collagen Type I Alpha 1 Chain | ENSOCUG00000004447; 100341109 | qOcuCED0015663 |
| RUNX2 | RUNX Family Transcription Factor 2 | ENSOCUG00000012881; 100347598 | qOcuCID0006259 |
| OPN | Osteopontin, Secreted Phosphoprotein 1 | ENSOCUG00000011739; 100008943 | qOcuCED0010864 |
| SP7 | Osterix, Sp7 Transcription Factor | ENSOCUG00000022536; 100008982 | qOcuCED0015349 |
| CAV1 | Caveolin 1 | ENSOCUG00000003811; 100356871 | qOcuCID0002632 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerrero, J.; Maevskaia, E.; Ghayor, C.; Bhattacharya, I.; Weber, F.E. Influence of Scaffold Microarchitecture on Angiogenesis and Regulation of Cell Differentiation during the Early Phase of Bone Healing: A Transcriptomics and Histological Analysis. Int. J. Mol. Sci. 2023, 24, 6000. https://doi.org/10.3390/ijms24066000
Guerrero J, Maevskaia E, Ghayor C, Bhattacharya I, Weber FE. Influence of Scaffold Microarchitecture on Angiogenesis and Regulation of Cell Differentiation during the Early Phase of Bone Healing: A Transcriptomics and Histological Analysis. International Journal of Molecular Sciences. 2023; 24(6):6000. https://doi.org/10.3390/ijms24066000
Chicago/Turabian StyleGuerrero, Julien, Ekaterina Maevskaia, Chafik Ghayor, Indranil Bhattacharya, and Franz E. Weber. 2023. "Influence of Scaffold Microarchitecture on Angiogenesis and Regulation of Cell Differentiation during the Early Phase of Bone Healing: A Transcriptomics and Histological Analysis" International Journal of Molecular Sciences 24, no. 6: 6000. https://doi.org/10.3390/ijms24066000