1. Introduction
AML comprises a group of malignant neoplastic disorders characterized by accumulation in the blood and/or in the bone marrow of immature white blood cells called leukemic cells, together with a deficiency in normal hematopoiesis. If conventional treatment based on intensive chemotherapy allows complete remission for 60 to 85% of patients, the prognosis of AML remains very negative, with a global survival rate of around 25 to 30%. In recent years, several studies linked this aggressiveness and the bad prognosis of AML to the ability of the leukemic cells to reprogram their metabolism in order to satisfy their high metabolic needs and to resist chemotherapy. In addition to the universally known Warburg effect, which involves a shift from oxidative phosphorylation as the main energy-producing pathway to high-rate aerobic glycolysis coupled with lactic acid fermentation in the cytosol, many metabolic pathways are deregulated in leukemic cells, sometimes in direct correlation with genetic alterations [
1]; this suggests multimodal metabolism in leukemogenesis. Among these metabolic pathways, lipid metabolism plays a particular role that has already been described in many solid tumors. Alterations, such as accentuated FAO and de novo lipid synthesis, could promote cancer cell survival and contribute to resistance to chemotherapeutic agents and metastasis [
2] through an increase in energy production and in lipid species necessary for the production of new cells. The overexpression of fatty acid synthase (FASN) gives cancer cells a proliferative advantage; conversely, lipolysis, through the activation of lipoprotein lipase (LPL), could increase the availability of FAs to levels necessary for cancer cell proliferation.
In AML, a recent study showed that patients with rapidly progressive AML who did not respond to treatment had alterations in their lipid metabolism [
3]. Interestingly, both aspects of lipid metabolism were implicated. On one hand, the degradation of FA through FAO may contribute to chemotherapy resistance. Indeed, resistant AML cells show overexpression of CD36, an FA transporter, at their membranes, and very intense FAO. This particular role of FAO in the survival of leukemic cells was also underlined in a demonstration of the overexpression of the enzyme 3-very long chain acyl-CoA dehydrogenase (VLCAD) in AML, of which a decrease in proliferation can be induced by gene attenuation or pharmacological inhibition via alteration of the Krebs cycle [
4]. Additionally, a recent study demonstrated that glycerophospholipid metabolism is involved in drug resistance in chronic myeloid leukemia through downregulation of the G0/G1 switch gene 2 (G0S2), which confers worse overall survival to patients [
5]. Similarly, Carnitine Palmitoyltransferase 1A (CPT1A), a key enzyme in carnitine-dependent transport across the mitochondrial inner membrane of FAs, was found to be overexpressed in bone marrow samples from AML patients with poor prognosis compared to those with normal bone marrow [
6]. These data provide a rationale for studying the impact of etomoxir (Eto), which irreversibly inhibits CPT1A, thereby preventing the entrance of fatty acids into the mitochondria, and their beta-oxidation.
On the other hand, the synthesis of FAs also plays a critical role. It was shown that low expression of ATP Citrate Lyase (ACL), the enzyme that initiates lipid synthesis through Acetyl-CoA production, is associated with favorable prognosis in patients with AML, and that discontinuation of cell growth was achieved when ACL was inhibited in leukemia cell models [
7]. Similarly, an anti-leukemic role of Acetyl-CoA Carboxylase 1 (ACC1), the enzyme that catalyzes the production of Malonyl-CoA from acetyl-CoA, has been shown [
8].
In AML patients, several lipid patterns representative of different subtypes of AML have recently been identified via mass spectrometry. Indeed, significant differences concerning modulation of the synthesis of ceramides and sphingolipids have been highlighted in patients presenting with t(8;21) compared to those presenting with an inv(16) or a normal karyotype [
9]. Additionally, Stuani et al. compared the dysregulation of lipid metabolism in a cellular model of AML mutated for the IDH1 gene to that in a non-mutated model [
10]. These differences are mainly characterized by an increase in phosphatidylinositol, sphingolipids, free cholesterol and monounsaturated fatty acids in IDH-mutated cells. This, again, highlights the impact of the cytogenetic characteristics and mutational status of AML patients on cellular metabolism. Finally, a recent study showed a decrease in total FAs in the plasma of patients with AML, as well as the attenuation of plasmatic phosphocholine, triglycerides and cholesterol esters [
11]. In addition, arachidonic acid appears to be increased in AML plasma samples, suggesting its involvement in the cancerous phenotype [
12].
Interestingly, lipids may play a role in the expression and functionality of certain oncogenes. It was shown that the aberrant localization of FLT3-ITD at the level of the endoplasmic reticulum depended on the palmitoylation of the receptor, and that the pharmacological inhibition of the depalmitoylation of the receptor had a synergistic effect on the inhibitor of FLT3 [
13]. Similarly, in an MLL-AF9 mouse model, a high-fat diet was shown to increase FLT3-dependent signaling via clusterization of the receptor in lipid rafts, and thus, promote the emergence of leukemic clones [
14]. Picou et al. tested the impact of polyunsaturated FAs (PUFAs) on acute myeloid leukemia cell lines and on fresh leukemic cells from patients, and induced the inhibition of mitochondrial respiration, and increased glycolysis and oxidative stress, which lead to the death of leukemic cells [
15]. However, Gyan et al. did not succeed in demonstrating the significant impact of FAs from fish oil emulsion on the complete remission of AML patients [
16].
Altogether, these data suggested that the reprogramming of lipid metabolism and the balance of lipid species could play a role in improving the prognosis of AML. In this context, we performed extensive characterization of the lipidome of human leukemic cell lines and plasma samples from AML patients, to decipher the characteristics of the lipid species and evaluate those of potential interest for targeting lipid metabolism in AML.
3. Discussion
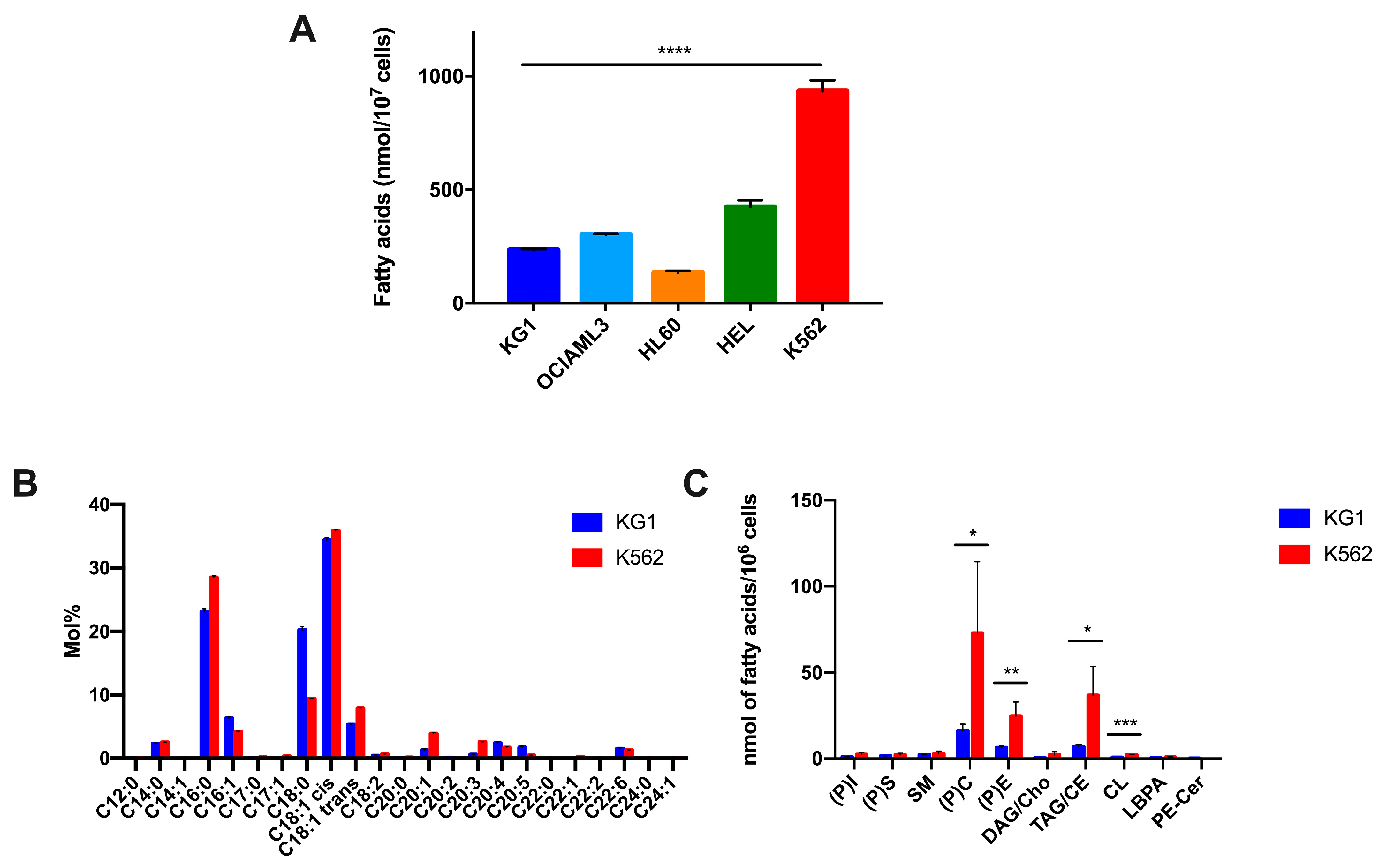
In this study, we firstly aimed to decipher the characteristics of lipid species in leukemic cells that could be of interest for targeting lipid metabolism for AML survival. Our first results show that, in their basic state, the five leukemic cell lines, which were representative of a certain diversity of AML, were characterized by very different total quantities of fatty acids, with a ratio of 1 to 7 between the K562 (the highest) and HL60 (the lowest) cell lines. To further characterize the relative proportion of fatty acids and lipid species, independently of the initial quantity of fatty acids, we chose to analyze K562 and KG1, which presented very different levels of FAs at baseline, suggesting very different lipids compositions. In fact, the qualitative analysis of their lipid contents showed very clear differences in the relative proportion of lipid species, demonstrating the diversity of the lipidome of leukemic cells. These results show that, in accordance with the results that we have previously published on the HRMAS profiles of these cell lines [
18], there are clear differences in the expression levels of metabolites between leukemic cell lines. The most probable hypothesis is a direct link between the metabolic profile and the genetic status of leukemic cells, as has already been shown in patients with t(8;21), inv(16) or IDH mutation [
9,
10].
These differences in the pattern of lipid expression suggest that leukemic cells might develop different lipid metabolism reprogramming under stress. As our goal was to study the adaptation of leukemic cells to global metabolic stress, and not to a particular deficiency, we chose, in our experiments, to use global serum deprivation. Although the use of serum-free medium that was selectively supplemented with a particular type of lipid species would be more precise, it would not have provided an overall picture of lipid reprogramming in this situation. Moreover, FBS is the major source of lipids in the medium, and the removal of serum is now valid and has been used in a number of publications; moreover, to date, there is no controlled medium whose exact and necessary FA composition is known.
Under these conditions, we confirmed our previous results, which showed the rapid adaptation of these two cell lines to starvation, allowing them to survive thanks to metabolic reprogramming [
18]. Regarding the lipid level, the first indicator of this adaptation was a significant decrease in their overall level of fatty acids after 48 h of serum deficiency, suggesting that the “basic quantitative reserves” of lipids in the cells are, in themselves, a protective mechanism. A decrease in lipid storage, such as DAG/cholesterol and TAG/cholesteryl ester, can also be attributed to the mobilization of lipids for survival. Interestingly, we observed significant variation in the lipid species that showed many similarities between the two cell lines, suggesting that under this stress condition, the leukemic cells set up common pathways, independently of the differences observed in their basal state. In particular, we observed a decrease in phosphatidyl serine, phosphatidyl-ethanolamine and phosphatidyl-choline, which suggests that leukemic cells use phosphatidyl-choline and its substrates to survive. The deregulation of phosphatidyl-choline metabolism has been described in many cancers [
19]. Several papers have noted changes in phosphatidyl-choline levels in cancers, particularly those of the prostate, ovaries and lung [
20,
21]. Similarly, the dysregulation of phosphatidyl-ethanolamine and phosphatidyl-inositol has been reported in prostate cancer. Moreover, Zheng et al. showed that under serum deprivation, human breast cancer cells exhibited increased phospholipase D (PLD) activity, leading to a decrease in (P)C (via the activation of its catabolism) and increased migration and metastasis potential [
22]. We also observed a significant decrease in arachidonic acid, high levels of which have been described in AML patients with a poor prognosis [
11]. Hence, our results show, for the first time, and to our knowledge, that the remodeling of lipid species is a major and shared mechanism of adaptation to stress in leukemic cells. This role was highlighted in the mechanism of sensitivity of acute promyelocytic leukemia (APL) to all-trans-retinoic acid (ATRA). Highly unsaturated fatty acids (HUFA) containing phosphatidyl-ethanolamine (HUFA-(P)E), such as those found in salmon testes, significantly enhanced dbcAMP-induced cell differentiation [
23]. This effect was suppressed by a protein kinase C inhibitor, suggesting that HUFA-(P)E might enhance dbcAMP-induced differentiation through the modulation of the protein kinase C signaling pathway in HL60 cells.
Overall, these results suggest that therapeutic targeting of lipid metabolism could be conceived as a complementary treatment to increase the therapeutic efficacy of induction chemotherapy. To assess this hypothesis, we studied the impacts of treating leukemic cells with etomoxir (Eto), which irreversibly inhibits CPT1A, thereby preventing the entrance of fatty acids into the mitochondria, and their beta-oxidation. In the presence of Eto, we observed a decrease in viability time and dose dependence in both cell lines, which increased dramatically in the absence of serum. However, sensitivity to Eto was significantly different between the two cell lines. The seahorse analysis of mitochondrial respiration demonstrated that in both cell lines, Eto—by inhibiting FAO—definitively reduces ATP production, even though KG1 was less impacted than K562, which was consistent with what we observed regarding viability.
The fact that K562 was significantly more sensitive compared to KG1, despite higher levels of FAs at steady state, could suggest deeper dependence of K562 cells on lipid metabolism, which is prevented by higher levels of lipid stock. Moreover, the IC
50 level in KG1 was more than 300 µM, and we cannot exclude that at these doses, as well as off-target effects such as the inhibition of Complex I of the electron transport chain (ETC) and the production of reactive species of oxygen (ROS), are the main factors responsible for cell death, as previously described [
24].
In addition, comparison of the modifications of lipid profiles induced by starvation or inhibition due to FAO, respectively, shows that if the two leukemia cell lines, despite their genotypic and phenotypic differences, have developed very closed mechanisms to fight against starvation, their levels of sensitivity to metabolic inhibitors are different and would depend on their initial lipid profiles. On one hand, K562 cells, with a dose close to their IC
50 value, present a blockade of the use of most of their fatty acids (in coherence with the mechanism of action of etomoxir), since their rates and relative proportions show very few differences compared to their basal profiles. In addition, no modification of the lipid species was observed in this cell line, suggesting that the blockade of FAO actually induced the death of K562 cells. Conversely, the KG1 cells, which are resistant to a dose of 100 µM of Eto (IC
50 = 320 µM), showed very significant variations (
p < 0.001) in the rates and relative proportions of several fatty acids. This decrease in the presence of Eto, which blocks FAO, together with the resistance of KG1 cells to etomoxir, suggests that KG1 cells have developed a mechanism of resistance against Eto that involves certain FAs and/or storage lipids. We can hypothesize that these lipid species are used for the renewal of new leukemic cells through their structural role, but also that they could be implicated in mechanism of drug resistance. Indeed, glycosphingolipid biosynthesis was shown to regulate drug sensitivity in clinically relevant models of drug-resistant chronic myeloid leukemias, through modulation of both the expression and activity of the drug efflux proteins ABCB1 and ABCC1 [
25].
Altogether, the results of this first part show that in AML, one can discriminate leukemic cells that are highly dependent on FAO, for which the use of an inhibitor of FAO should be efficient, and conversely, leukemic cells that are independent of FAO, for which an inhibitor of FAO would not be a reliable additional therapy. Therefore, the targeting of lipid metabolism should be proposed only for a subset of AML patients who display a sensitive “lipidic phenotype”.
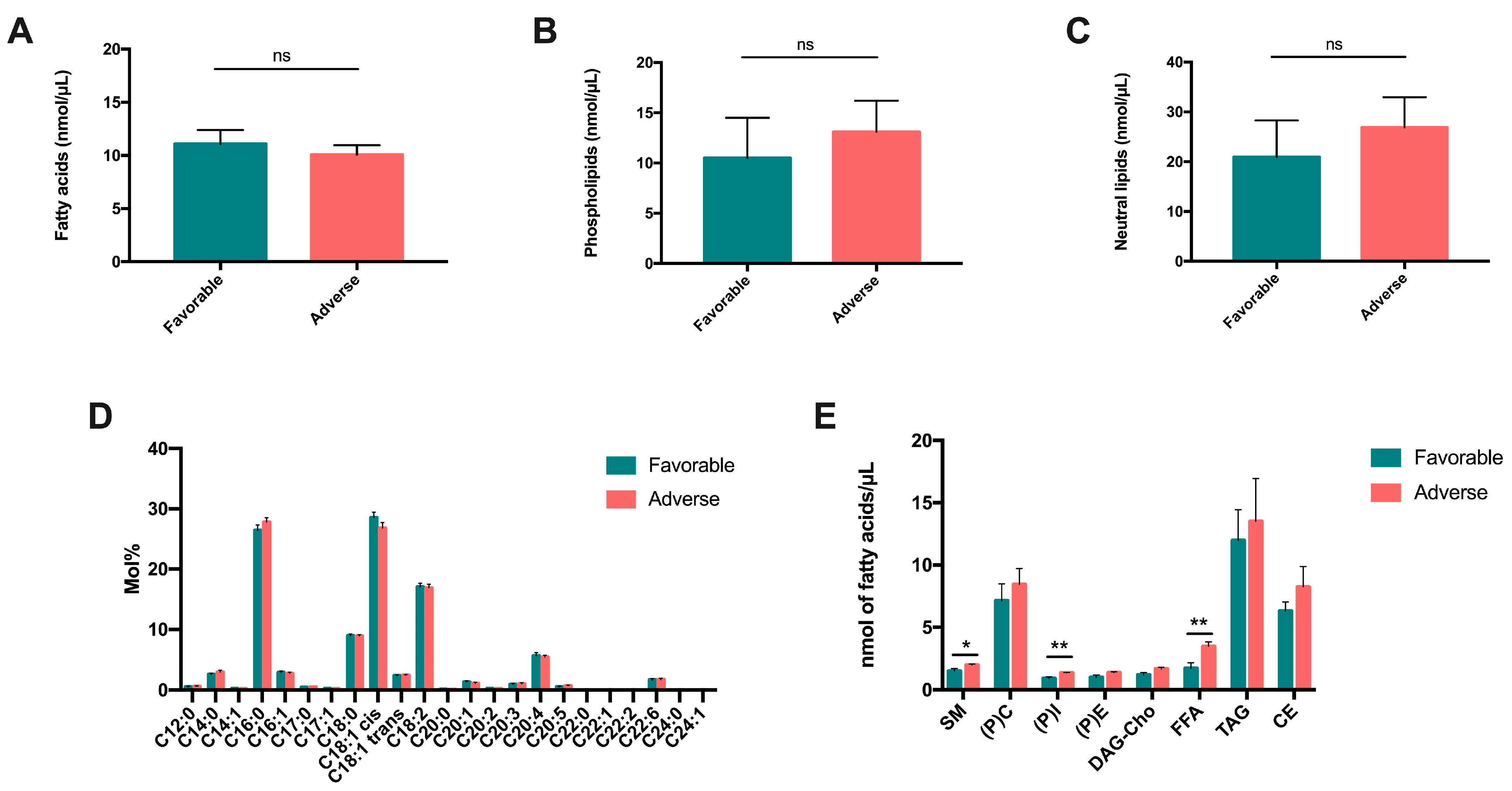
Leukemic cell lines represent a questionable model of what happens in patients. This is why we reinforced the in vitro analysis with an ex vivo analysis of patient samples, in order to assess the role of the patients’ lipidome on their survival. We firstly compared total fatty acids, phospholipids and neutral lipids among a cohort of 39 AML patients with high (n = 21) or low (n = 18) prognoses (according to ELN 2017 [
17]). In addition, as all investigations of lipid species (FA and phospholipid composition) using LC-MS and GC-MS are very time-consuming experiences, require very high-level expertise and could not be carried out on large series of patients, we further selected five patients belonging to the high or low prognostic subgroups, respectively, for extensive lipidome analysis. Although this number of patients may seem low (n = 10, 5 patients in each group), to our knowledge, this type of characterization of the lipidome in samples from AML patients has never before been published. In addition, the number of independent measurements for this type of assay is classically n = 3m and in our case, we had five independent values for each parameter; this gives statistical significance to our results, which are quite robust.
We found an increase in the quantity of neutral lipids (FFAs), which are storage lipids, in patients with an unfavorable prognosis, suggesting that the mobilization of these storage lipids improve the viability of leukemic cells. Moreover, the increase in the quantity of phospholipids, and in particular, of sphingomyelin and phosphocholine (PC), in patients with an unfavorable prognosis confirms that PLs are markers of aggressiveness and tumor progression, which are associated with a poor prognosis in AML. These results are consistent with those described in cancers [
26,
27]. Elevated levels of PC indicate intense cell proliferation, commonly observed in cancers, in association with an increase in phosphoethanolamine (PE) [
26]. At the neurological level, two studies have shown that patients with a low-grade glioma that progressed to a high-grade glioma had much higher levels of PC and PE [
28,
29]. Similarly, Wang et al. observed that PC was expressed at a higher level in the sera of patients with an intermediate prognosis compared to patients with a favorable prognosis. This is in line with an unfavorable impact of PC on prognosis [
30]. The fact that we observed, in a previous study, that an increase in PC was a common mechanism of survival against starvation in leukemic cell lines [
18] strongly suggests that its prognostic role in patients could be explained by a “protective” role of PC, which could increase the resistance of leukemic cells in stressful situations. It is very interesting to note that in this study, we observed a decrease in phosphatidyl-choline in a situation of metabolic stress in our cell lines. Taking into account that the catabolism of phosphatidyl-choline can produce phosphocholine, we can hypothesize that leukemic cells use phosphatidyl-choline catabolism to generate lipid mediators that will increase their survival. Moreover, a team recently created an inventory of lipidome in the plasma of patients with AML, and showed a decrease in total FFAs, including PC; this was probably be due to increased oxidation of FAs in AML cells. Altogether, these data underline the crucial role of phosphatidyl-choline and phosphocholine metabolism in the survival of leukemic cells, suggesting that it could be also an interesting therapeutic target.
In conclusion, our data show that balanced lipid metabolism is a phenotypic marker of the diversity (very probably genetic) of leukemic cells that significantly influences their proliferation and resistance to stress, and could be a fruitful therapeutic target to improve the survival of some AML patients harboring a particular “lipidic phenotype”.
4. Materials and Methods
4.1. Cell Culture and Conditions
The cells used for this experiment were purchased from the ATCC (American Type Culture Collection, Manassas, VA, USA) society. Five human leukemic cell lines were used in our study: HL60, HEL, OCI-AML3, KG1 and K562. The K562 line was derived from a patient who initially had chronic myeloid leukemia, but the cells were collected from this patient in the phase of transformation into AML. It is therefore considered a model of AML. The cells were cultured at a density of 0.5 × 105 cells/mL for exponential growth in a medium containing 10% fetal bovine serum (FBS), 1% L-glutamine, 1% penicillin/streptomycin and RPMI 1640. The cells were incubated in a laboratory oven at 37% and 5% CO2, and the media were changed every two–three days.
The conditions used for the experiments were a control, a culture with etomoxir, starvation (medium without serum) and a culture with etomoxir under the conditions of starvation. Etomoxir sodium salt hydrate was purchased from Sigma Aldrich (Saint-Quentin-Fallavier, France) and reconstituted in sterile water for injection into a stock solution of 10 mM. For the cultures with etomoxir, the cells were suspended in a routinely used medium, and etomoxir was diluted directly with the medium to the desired concentration (100 µM, 200 µM or 400 µM). For starvation, the cells were centrifuged and resuspended in a medium deprived of FBS, with 1% L-glutamine, 1% penicillin/streptomycin, RPMI 1640 and 1% insulin–transferrin–selenium. The cultures with etomoxir under the conditions of starvation were prepared using the same protocol as the cultures with etomoxir under non-starvation conditions.
4.2. Fluorescence-Activated Cell Sorting
An amount of 0.3 × 106 cells were centrifuged at 1400 rpm for 10 min and washed twice with phosphate-buffered saline (PBS) solution. The cells were labeled with 5 µL of Annexin V (FITC Annexin V Apoptosis Detection Kit I from BD Pharmingen, France) and 5 µL of Propidium iodide (PI), and incubated for 15 min at room temperature, protected from the light. Their acquisition was conducted using a FACS Lyric cytometer. Data analysis was performed using BD FACSuite Software. Handling was performed in parallel for each cell line and in three independent experiments.
4.3. Mitochondrial Respiration Assay
The bioenergetics assay was performed using the Seahorse XF Cell Mito stress test kit and the Seahorse XFe96 analyzer, purchased from Agilent Technologies. The day before the assay, the tissue culture plate was coated using Corning Cell-Tak adhesive, according to the supplier’s recommendations. The Seahorse cartridge was hydrated with sterile water and incubated overnight in a CO2-free incubator, at 37 °C. The day of the assay, the Seahorse medium was prepared, and the pH solution adjusted to 7.4. The following step involved seeding the cells on the previously coated culture plate. The cells were enumerated and centrifuged for 5 min at 2000 rpm, and the supernatant was removed and replaced with Seahorse medium. The cells were then incubated for 45 min in a CO2-free incubator, at 37 °C. During the incubation, the stock solutions were reconstituted and diluted to the appropriate concentrations as follows: ATP synthase inhibitor oligomycin at 1.5 µmol/L, uncoupling agent carbonyl cyanide p-trifluoro-methoxyphenyl hydrazone (FCCP) at 0.5 µmol/L and 2 µmol/L, fatty acid oxidation inhibitor etomoxir at 4 µmol/L, and inhibitors of complex I/III of the electron transport chain antimycin/rotenone at 0.5 µmol/L. The oxygen consumption rate (OCR) was measured at baseline and after sequential injections with the previously mentioned solutions.
4.4. Determination of IC50
The half-maximal inhibitory concentration (IC50) was determined using the PrestoBlue cell viability reagent, purchased from Thermo Fisher Scientific (Waltham, MA, USA). The cell line K562 was cultured at an initial density of 40,000 cells/well with increasing concentrations of etomoxir (0, 50, 100, 150, 200, 250, 300 µM), for 24 h, on a black culture plate with a flat, clear bottom. After 24 h, 10 µL of PrestoBlue reagent was added to each well. The cells were incubated for 30 min at 37 °C. The fluorescence was measured using a Varioskan microplate reader (Thermo Fisher Scientific).
The measurements were performed in triplicate for each condition. The average fluorescence was calculated for the triplicates corresponding to each concentration, and the background (blank) was subtracted.
4.5. Lipid Analysis
The cells were collected after 24 h of treatment, followed by quenching in an ethanol/dry ice bath, to stop cell metabolism. After quenching, the pellets were washed twice with PBS and stored at −80 °C until the day of the lipid extraction. Total lipids were extracted in chloroform/methanol (1:2) with an internal standard (10 nmol tridecanoic acid C13:0 and 10 nmol PC C23:0). By adding chloroform and 0.2% KCl/0.5% acetic acid, biphasic separation, at a chloroform/methanol/water ratio of 2:1:0.8 (v/v/v), was generated. The lowest organic phase that was recovered was the total lipid fraction. Total lipid equivalent of 0.2–0.5 M of the cells were dried in vacuo, and the derivatizing agent TMSH (Machery-Nagel, Hoerdt, France) in chloroform/methanol (1:1, v/v) was added, followed by gas chromatography–mass spectrometry (GC-MS, Agilent 5977A-7890B). The total fatty acid amount was quantified using Mass Hunter software (Agilent, Santa Clara, CA, USA) in the form of fatty acid methyl esters, using the calibration curve generated using the authentic standards, followed by normalization using the internal standards and cell number.
4.6. Plasma Analysis
We recovered the plasma of 39 adult patients with a diagnosis of de novo AML, who received an induction treatment according to the standards for “fit” patients. All subjects gave their informed consent for inclusion before they participated in the study. The study was conducted in accordance with the Declaration of Helsinki, and the protocol was approved by the Ethics Committee of CHUGA (Project 38RC13.209). The patients’ prognoses were assessed according to the ELN 2017 classification: 21 patients belonged to the favorable prognosis group and 18 to the unfavorable prognosis group. Plasma was collected upon diagnosis of AML. All the characteristics of the patients are summarized in
Supplemental Table S1. The analysis of total FAs and lipid species was carried out according to the protocol described in the section “Lipid Analysis”. Detailed information on the composition of the lipid species was determined for 5 patients representative of each of the two prognostic subgroups. Thereby, we were able to identify 8 compounds in these 10 patients. Sphingomyelin (SM), phosphatidyl-choline ((P)C), phosphatidyl-inositol ((P)I) and phosphatidyl-ethanolamine ((P)E), which correspond to phospholipids (PLs), appeared after the first migration. On the other hand, a mixture of diacylglycerol and cholesterol (DAG-Cho), free fatty acids (FFAs), triacylglycerol (TAG) and cholesteryl esters (CEs), which correspond to neutral lipids (NLs), appeared after the second migration.
4.7. Statistical Analysis
For statistical analysis, the Kruskal–Wallis test was used for three or more groups, and the Mann–Whitney test was used to compare two groups. Unless otherwise specified, the experiments were repeated at least three times. The data were analyzed using GraphPad Prism software (version 7). A p-value < 0.05 was considered significant.

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





