
The Downregulation of Opioid Receptors and Neuropathic Pain


Abstract
:1. Introduction
2. The Opioid System and Opioids in NP
2.1. Endogenous Opioid System
2.2. Opioid Receptor-Targeted Drugs for NP
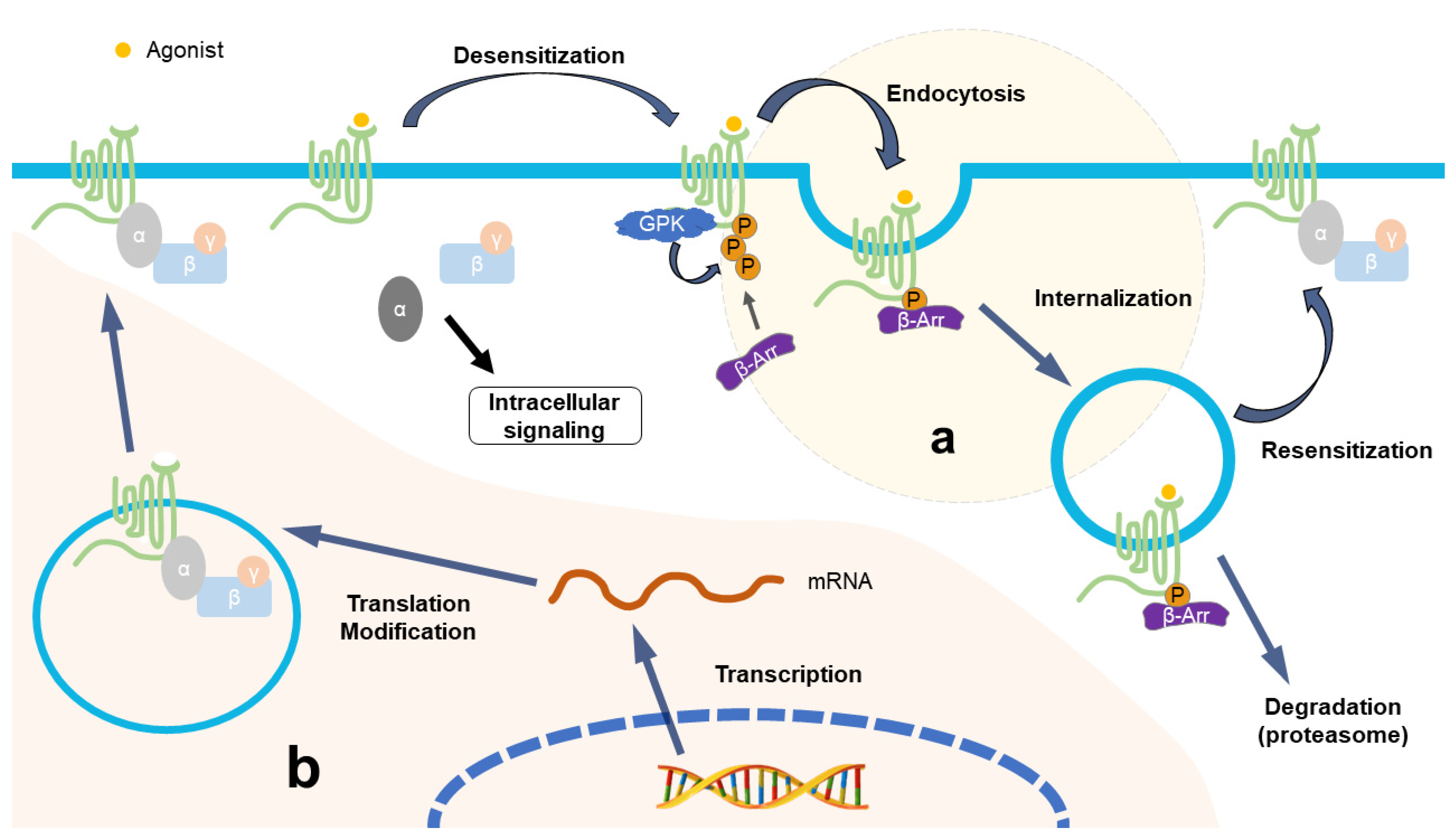
2.3. The Same Outcome of NP and Long-Term Opioid Exposure
3. Downregulation of Opioid Receptors after Repeated Opioid Treatment
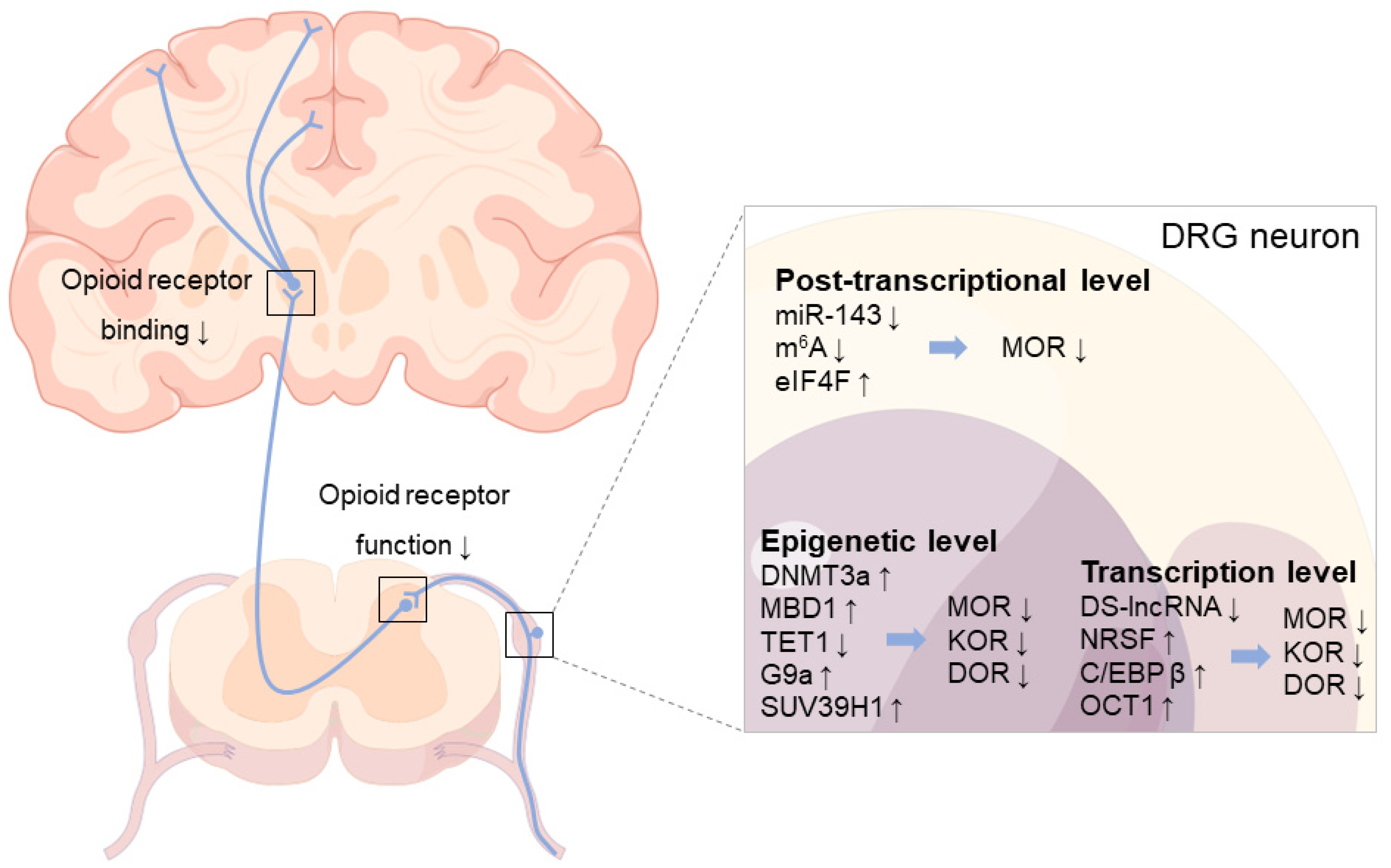
4. Downregulation of Opioid Receptors in DRG following Nerve Injury
4.1. Epigenetic Level
4.2. Transcription Level
4.3. Post-Transcriptional Level
5. Downregulation of Opioid Receptors in the CNS following Nerve Injury
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Jensen, T.S.; Baron, R.; Haanpää, M.; Kalso, E.; Loeser, J.D.; Rice, A.S.C.; Treede, R.D. A new definition of neuropathic pain. Pain 2011, 152, 2204–2205. [Google Scholar] [CrossRef] [PubMed]
- IASP. 2017. Available online: https://www.iasp-pain.org/resources/terminology/#pain (accessed on 16 July 2020).
- Torrance, N.; Smith, B.H.; Bennett, M.I.; Lee, A.J. The epidemiology of chronic pain of predominantly neuropathic origin. Results from a general population survey. J. Pain 2006, 7, 281–289. [Google Scholar] [CrossRef]
- Bouhassira, D.; Lantéri-Minet, M.; Attal, N.; Laurent, B.; Touboul, C. Prevalence of chronic pain with neuropathic characteristics in the general population. Pain 2008, 136, 380–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Hecke, O.; Austin, S.K.; Khan, R.A.; Smith, B.H.; Torrance, N. Neuropathic pain in the general population: A systematic review of epidemiological studies. Pain 2014, 155, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Attal, N.; Lanteri-Minet, M.; Laurent, B.; Fermanian, J.; Bouhassira, D. The specific disease burden of neuropathic pain: Results of a French nationwide survey. Pain 2011, 152, 2836–2843. [Google Scholar] [CrossRef]
- Finnerup, N.B.; Haroutounian, S.; Kamerman, P.; Baron, R.; Bennett, D.L.H.; Bouhassira, D.; Cruccu, G.; Freeman, R.; Hansson, P.; Nurmikko, T.; et al. Neuropathic pain: An updated grading system for research and clinical practice. Pain 2016, 157, 1599–1606. [Google Scholar] [CrossRef] [Green Version]
- Blyth, F.M. Global burden of neuropathic pain. Pain 2018, 159, 614–617. [Google Scholar] [CrossRef] [PubMed]
- Loh, E.; Mirkowski, M.; Agudelo, A.R.; Allison, D.J.; Benton, B.; Bryce, T.N.; Guilcher, S.; Jeji, T.; Kras-Dupuis, A.; Kreutzwiser, D.; et al. The CanPain SCI clinical practice guidelines for rehabilitation management of neuropathic pain after spinal cord injury: 2021 update. Spinal Cord. 2022, 60, 548–566. [Google Scholar] [CrossRef]
- Colloca, L.; Ludman, T.; Bouhassira, D.; Baron, R.; Dickenson, A.H.; Yarnitsky, D.; Freeman, R.; Truini, A.; Attal, N.; Finnerup, N.B.; et al. Neuropathic pain. Nat. Rev. Dis. Prim. 2017, 3, 17002. [Google Scholar] [CrossRef] [Green Version]
- Finnerup, N.B.; Kuner, R.; Jensen, T.S. Neuropathic pain: From mechanisms to treatment. Physiol. Rev. 2021, 101, 259–301. [Google Scholar] [CrossRef]
- Cox, B.M.; Christie, M.J.; Devi, L.; Toll, L.; Traynor, J.R. Challenges for opioid receptor nomenclature: IUPHAR Review 9. Br. J. Pharmacol. 2015, 172, 317–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, C. Opioid receptors. Annu. Rev. Med. 2016, 67, 433–451. [Google Scholar] [CrossRef]
- Poon, A.; Ing, J.; Hsu, E. Opioid-related side effects and management. Cancer Treat. Res. 2021, 182, 97–105. [Google Scholar]
- Cahill, C.M.; Walwyn, W.; Taylor, A.M.W.; Pradhan, A.A.A.; Evans, C.J. Allostatic mechanisms of opioid tolerance beyond desensitization and downregulation. Trends Pharmacol. Sci. 2016, 37, 963–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volkow, N.D.; McLellan, A.T. Opioid abuse in chronic pain-misconceptions and mitigation strategies. N. Engl. J. Med. 2016, 374, 1253–1263. [Google Scholar] [CrossRef]
- Stein, C.; Machelska, H. Modulation of peripheral sensory neurons by the immune system: Implications for pain therapy. Pharmacol. Rev. 2011, 63, 860–881. [Google Scholar] [CrossRef] [Green Version]
- Tsao, P.; von Zastrow, M. Downregulation of G protein-coupled receptors. Curr. Opin. Neurobiol. 2000, 10, 365–369. [Google Scholar] [CrossRef]
- Di Guglielmo, G.M.; Drake, P.G.; Baass, P.C.; Authier, F.; Posner, B.I.; Bergeron, J.J. Insulin receptor internalization and signalling. Mol. Cell. Biochem. 1998, 182, 59–63. [Google Scholar] [CrossRef]
- Koch, T.; Höllt, V. Role of receptor internalization in opioid tolerance and dependence. Pharmacol. Ther. 2008, 117, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.T.; Ingram, S.L.; Henderson, G.; Chavkin, C.; von Zastrow, M.; Schulz, S.; Koch, T.; Evans, C.J.; Christie, M.J. Regulation of μ-opioid receptors: Desensitization, phosphorylation, internalization, and tolerance. Pharmacol. Rev. 2013, 65, 223–254. [Google Scholar] [CrossRef] [Green Version]
- Mansour, A.; Hoversten, M.T.; Taylor, L.P.; Watson, S.J.; Akil, H. The cloned mu, delta and kappa receptors and their endogenous ligands: Evidence for two opioid peptide recognition cores. Brain Res. 1995, 700, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Massaly, N.; Morón, J.A. Pain and opioid systems, implications in the opioid epidemic. Curr. Opin. Behav. Sci. 2019, 26, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, R.J. Endogenous opiates and behavior: 2019. Peptides 2021, 141, 170547. [Google Scholar] [CrossRef] [PubMed]
- Corder, G.; Castro, D.C.; Bruchas, M.R.; Scherrer, G. Endogenous and exogenous opioids in pain. Annu. Rev. Neurosci. 2018, 41, 453–473. [Google Scholar] [CrossRef]
- Le Merrer, J.; Becker, J.A.; Befort, K.; Kieffer, B.L. Reward processing by the opioid system in the brain. Physiol. Rev. 2009, 89, 1379–1412. [Google Scholar] [CrossRef]
- Vicente-Sanchez, A.; Pradhan, A.A. Ligand-directed signaling at the delta opioid receptor. Handb. Exp. Pharmacol. 2018, 247, 73–85. [Google Scholar] [PubMed]
- Land, B.B.; Bruchas, M.R.; Lemos, J.C.; Xu, M.; Melief, E.J.; Chavkin, C. The dysphoric component of stress is encoded by activation of the dynorphin kappa-opioid system. J. Neurosci. 2008, 28, 407–414. [Google Scholar] [CrossRef] [Green Version]
- Leitl, M.D.; Onvani, S.; Bowers, M.S.; Cheng, K.; Rice, K.C.; Carlezon Jr, W.A.; Banks, M.L.; Negus, S.S. Pain-related depression of the mesolimbic dopamine system in rats: Expression, blockade by analgesics, and role of endogenous κ-opioids. Neuropsychopharmacology 2014, 39, 614–624. [Google Scholar] [CrossRef]
- Al-Hasani, R.; McCall, J.G.; Shin, G.; Gomez, A.M.; Schmitz, G.P.; Bernardi, J.M.; Pyo, C.O.; Park, S.I.; Marcinkiewcz, C.M.; Crowley, N.A.; et al. Distinct subpopulations of nucleus accumbens dynorphin neurons drive aversion and reward. Neuron 2015, 87, 1063–1077. [Google Scholar] [CrossRef] [Green Version]
- Meunier, J.C.; Mollereau, C.; Toll, L.; Suaudeau, C.; Moisand, C.; Alvinerie, P.; Butour, J.L.; Guillemot, J.C.; Ferrara, P.; Monsarrat, B.; et al. Isolation and structure of the endogenous agonist of opioid receptor-like ORL1 receptor. Nature 1995, 377, 532–535. [Google Scholar] [CrossRef]
- Mollereau, C.; Mouledous, L. Tissue distribution of the opioid receptor-like (ORL1) receptor. Peptides 2000, 21, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, A.; Brunori, G.; Cippitelli, A.; Toll, N.; Schoch, J.; Kieffer, B.L.; Toll, L. Analysis of the distribution of spinal NOP receptors in a chronic pain model using NOP-eGFP knock-in mice. Br. J. Pharmacol. 2018, 175, 2662–2675. [Google Scholar] [CrossRef] [Green Version]
- Toll, L.; Bruchas, M.R.; Calo’, G.; Cox, B.M.; Zaveri, N.T. Nociceptin/Orphanin FQ receptor structure, signaling, ligands, functions, and interactions with opioid systems. Pharmacol. Rev. 2016, 68, 419–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, K.E.; Pedersen, C.E.; Gomez, A.M.; Spangler, S.M.; Walicki, M.C.; Feng, S.Y.; Stewart, S.L.; Otis, J.M.; Al-Hasani, R.; McCall, J.G.; et al. A paranigral VTA nociceptin circuit that constrains motivation for reward. Cell 2019, 178, 653–671.e19. [Google Scholar] [CrossRef] [PubMed]
- Kallupi, M.; Scuppa, G.; de Guglielmo, G.; Calò, G.; Weiss, F.; Statnick, M.A.; Rorick-Kehn, L.M.; Ciccocioppo, R. Genetic deletion of the nociceptin/orphanin FQ receptor in the rat confers resilience to the development of drug addiction. Neuropsychopharmacology 2017, 42, 695–706. [Google Scholar] [CrossRef] [Green Version]
- de Guglielmo, G.; Matzeu, A.; Kononoff, J.; Mattioni, J.; Martin-Fardon, R.; George, O. Cebranopadol blocks the escalation of cocaine intake and conditioned reinstatement of cocaine seeking in rats. J. Pharmacol. Exp. Ther. 2017, 362, 378–384. [Google Scholar] [CrossRef] [Green Version]
- Fields, H. State-dependent opioid control of pain. Nat. Rev. Neurosci. 2004, 5, 565–575. [Google Scholar] [CrossRef]
- Ossipov, M.H.; Dussor, G.O.; Porreca, F. Central modulation of pain. J. Clin. Investig. 2010, 120, 3779–3787. [Google Scholar] [CrossRef] [Green Version]
- Bagley, E.E.; Ingram, S.L. Endogenous opioid peptides in the descending pain modulatory circuit. Neuropharmacology 2020, 173, 108131. [Google Scholar] [CrossRef]
- Ringkamp, M.; Dougherty, P.M.; Raja, S.N. Anatomy and physiology of the pain signaling process. In Essentials of Pain Medicine; Elsevier: Amsterdam, The Netherlands, 2018; pp. 3–10. [Google Scholar]
- Benyamin, R.; Trescot, A.M.; Datta, S.; Buenaventura, R.; Adlaka, R.; Sehgal, N.; Glaser, S.E.; Vallejo, R. Opioid complications and side effects. Pain Physician 2008, 11 (Suppl. S2), S105–S120. [Google Scholar] [CrossRef]
- Nafziger, A.N.; Barkin, R.L. Opioid therapy in acute and chronic pain. J. Clin. Pharmacol. 2018, 58, 1111–1122. [Google Scholar] [CrossRef]
- Rosenblum, A.; Marsch, L.A.; Joseph, H.; Portenoy, R.K. Opioids and the treatment of chronic pain: Controversies, current status, and future directions. Exp. Clin. Psychopharmacol. 2008, 16, 405–416. [Google Scholar] [CrossRef] [Green Version]
- Rittner, H.L.; Brack, A.; Stein, C. Pain and the immune system. Br. J. Anaesth. 2008, 101, 40–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthes, H.W.; Maldonado, R.; Simonin, F.; Valverde, O.; Slowe, S.; Kitchen, I.; Befort, K.; Dierich, A.; Le Meur, M.; Dollé, P.; et al. Loss of morphine-induced analgesia, reward effect and withdrawal symptoms in mice lacking the mu-opioid-receptor gene. Nature 1996, 383, 819–823. [Google Scholar] [CrossRef] [PubMed]
- McDougall, J.J. Peripheral analgesia: Hitting pain where it hurts. Biochim. Biophys. Acta 2011, 1812, 459–467. [Google Scholar] [CrossRef] [Green Version]
- Stein, C. New concepts in opioid analgesia. Expert Opin. Investig. Drugs 2018, 27, 765–775. [Google Scholar] [CrossRef] [PubMed]
- Electronic Medicines Compendium (eMC). 2018. Available online: https://www.medicines.org.uk/emc (accessed on 23 November 2018).
- Smith, H.S. Opioids and neuropathic pain. Pain Physician 2012, 15 (Suppl. S3), ES93–ES110. [Google Scholar] [CrossRef]
- Sommer, C.; Klose, P.; Welsch, P.; Petzke, F.; Häuser, W. Opioids for chronic non-cancer neuropathic pain. An updated systematic review and meta-analysis of efficacy, tolerability and safety in randomized placebo-controlled studies of at least 4 weeks duration. Eur. J. Pain 2020, 24, 3–18. [Google Scholar] [CrossRef]
- Finnerup, N.B.; Attal, N.; Haroutounian, S.; McNicol, E.; Baron, R.; Dworkin, R.H.; Gilron, I.; Haanpää, M.; Hansson, P.; Jensen, T.S.; et al. Pharmacotherapy for neuropathic pain in adults: A systematic review and meta-analysis. Lancet Neurol. 2015, 14, 162–173. [Google Scholar] [CrossRef] [Green Version]
- Ray, W.A.; Chung, C.P.; Murray, K.T.; Hall, K.; Stein, C.M. Prescription of long-acting opioids and mortality in patients with chronic noncancer pain. JAMA 2016, 315, 2415–2423. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, A.B.; Dworkin, R.H. Treatment of neuropathic pain: An overview of recent guidelines. Am. J. Med. 2009, 122 (Suppl. S10), S22–S32. [Google Scholar] [CrossRef] [PubMed]
- Azzam, A.A.H.; McDonald, J.; Lambert, D.G. Hot topics in opioid pharmacology: Mixed and biased opioids. Br. J. Anaesth. 2019, 122, e136–e145. [Google Scholar] [CrossRef]
- Starnowska-Sokół, J.; Przewłocka, B. Multifunctional opioid-derived hybrids in neuropathic pain: Preclinical evidence, ideas and challenges. Molecules 2020, 25, 5520. [Google Scholar] [CrossRef] [PubMed]
- Vicario, N.; Pasquinucci, L.; Spitale, F.M.; Chiechio, S.; Turnaturi, R.; Caraci, F.; Tibullo, D.; Avola, R.; Gulino, R.; Parenti, R.; et al. Simultaneous activation of mu and delta opioid receptors reduces allodynia and astrocytic connexin 43 in an animal model of neuropathic pain. Mol. Neurobiol. 2019, 56, 7338–7354. [Google Scholar] [CrossRef] [PubMed]
- Podolsky, A.T.; Sandweiss, A.; Hu, J.; Bilsky, E.J.; Cain, J.P.; Kumirov, V.K.; Lee, Y.S.; Hruby, V.J.; Vardanyan, R.S.; Vanderah, T.W. Novel fentanyl-based dual μ/δ-opioid agonists for the treatment of acute and chronic pain. Life Sci. 2013, 93, 1010–1016. [Google Scholar] [CrossRef] [Green Version]
- Lei, W.; Vekariya, R.H.; Ananthan, S.; Streicher, J.M. A novel mu-delta opioid agonist demonstrates enhanced efficacy with reduced tolerance and dependence in mouse neuropathic pain models. J. Pain 2020, 21, 146–160. [Google Scholar] [CrossRef]
- Xu, B.; Zhang, M.; Shi, X.; Zhang, R.; Chen, D.; Chen, Y.; Wang, Z.; Qiu, Y.; Zhang, T.; Xu, K.; et al. The multifunctional peptide DN-9 produced peripherally acting antinociception in inflammatory and neuropathic pain via μ- and κ-opioid receptors. Br. J. Pharmacol. 2020, 177, 93–109. [Google Scholar] [CrossRef]
- Albert-Vartanian, A.; Boyd, M.R.; Hall, A.L.; Morgado, S.J.; Nguyen, E.; Nguyen, V.P.H.; Patel, S.P.; Russo, L.J.; Shao, A.J.; Raffa, R.B. Will peripherally restricted kappa-opioid receptor agonists (pKORAs) relieve pain with less opioid adverse effects and abuse potential? J. Clin. Pharm. Ther. 2016, 41, 371–382. [Google Scholar] [CrossRef] [Green Version]
- Vekariya, R.H.; Lei, W.; Ray, A.; Saini, S.K.; Zhang, S.; Molnar, G.; Barlow, D.; Karlage, K.L.; Bilsky, E.J.; Houseknecht, K.L.; et al. Synthesis and structure-activity relationships of 5′-Aryl-14-alkoxypyridomorphinans: Identification of a μ opioid receptor agonist/δ opioid receptor antagonist ligand with systemic antinociceptive activity and diminished opioid side effects. J. Med. Chem. 2020, 63, 7663–7694. [Google Scholar] [CrossRef]
- Ballantyne, J.C.; Chavkin, C. Will biased ligands be the opioids of the future? Pain 2020, 161, 1945–1948. [Google Scholar] [CrossRef]
- Che, T.; Dwivedi-Agnihotri, H.; Shukla, A.K.; Roth, B.L. Biased ligands at opioid receptors: Current status and future directions. Sci. Signal 2021, 14, eaav0320. [Google Scholar] [CrossRef]
- Martínez-Navarro, M.; Maldonado, R.; Baños, J.E. Why mu-opioid agonists have less analgesic efficacy in neuropathic pain? Eur. J. Pain 2019, 23, 435–454. [Google Scholar] [CrossRef] [Green Version]
- Santoni, A.; Mercadante, S.; Arcuri, E. Chronic cancer and non-cancer pain and opioid-induced hyperalgesia share common mechanisms: Neuroinflammation and central sensitization. Minerva Anestesiol. 2021, 87, 210–222. [Google Scholar] [CrossRef]
- Vanderah, T.W.; Gardell, L.R.; Burgess, S.E.; Ibrahim, M.; Dogrul, A.; Zhong, C.-M.; Zhang, E.-T.; Malan, T.P.J.; Ossipov, M.H.; Lai, J.; et al. Dynorphin promotes abnormal pain and spinal opioid antinociceptive tolerance. J. Neurosci. 2000, 20, 7074–7079. [Google Scholar] [CrossRef] [Green Version]
- Bai, X.; Wang, C.; Zhang, X.; Feng, Y.; Zhang, X. The role of testosterone in mu-opioid receptor expression in the trigeminal ganglia of opioid-tolerant rats. Neurosci. Lett. 2020, 723, 134868. [Google Scholar] [CrossRef] [PubMed]
- Ossipov, M.H.; Lai, J.; King, T.; Vanderah, T.W.; Porreca, F. Underlying mechanisms of pronociceptive consequences of prolonged morphine exposure. Biopolymers 2005, 80, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Rogers, T.J. Bidirectional regulation of opioid and chemokine function. Front. Immunol. 2020, 11, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhodin, A.; Grönbladh, A.; Ginya, H.; Nilsson, K.W.; Rosenblad, A.; Zhou, Q.; Enlund, M.; Hallberg, M.; Gordh, T.; Nyberg, F. Combined analysis of circulating β-endorphin with gene polymorphisms in OPRM1, CACNAD2 and ABCB1 reveals correlation with pain, opioid sensitivity and opioid-related side effects. Mol. Brain 2013, 6, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruehl, S.; Burns, J.W.; Gupta, R.; Buvanendran, A.; Chont, M.; Orlowska, D.; Schuster, E.; France, C.R. Do resting plasma β-endorphin levels predict responses to opioid analgesics? Clin. J. Pain 2017, 33, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Wawrzczak-Bargieła, A.; Ziółkowska, B.; Piotrowska, A.; Starnowska-Sokół, J.; Rojewska, E.; Mika, J.; Przewłocka, B.; Przewłocki, R. Neuropathic pain dysregulates gene expression of the forebrain opioid and dopamine systems. Neurotox. Res. 2020, 37, 800–814. [Google Scholar] [CrossRef] [Green Version]
- Navratilova, E.; Ji, G.; Phelps, C.; Qu, C.; Hein, M.; Yakhnitsa, V.; Neugebauer, V.; Porreca, F. Kappa opioid signaling in the central nucleus of the amygdala promotes disinhibition and aversiveness of chronic neuropathic pain. Pain 2019, 160, 824–832. [Google Scholar] [CrossRef]
- Briscini, L.; Corradini, L.; Ongini, E.; Bertorelli, R. Up-regulation of ORL-1 receptors in spinal tissue of allodynic rats after sciatic nerve injury. Eur. J. Pharmacol. 2002, 447, 59–65. [Google Scholar] [CrossRef]
- Back, S.K.; Lee, J.; Hong, S.K.; Na, H.S. Loss of spinal mu-opioid receptor is associated with mechanical allodynia in a rat model of peripheral neuropathy. Pain 2006, 123, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Obara, I.; Parkitna, J.R.; Korostynski, M.; Makuch, W.; Kaminska, D.; Przewlocka, B.; Przewlocki, R. Local peripheral opioid effects and expression of opioid genes in the spinal cord and dorsal root ganglia in neuropathic and inflammatory pain. Pain 2009, 141, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.; Perez, F.M.; Wang, W.; Guan, X.; Zhao, X.; Fisher, J.L.; Guan, Y.; Sweitzer, S.M.; Raja, S.N.; Tao, Y.X. Dynamic temporal and spatial regulation of mu opioid receptor expression in primary afferent neurons following spinal nerve injury. Eur. J. Pain 2011, 15, 669–675. [Google Scholar]
- Rashid, M.H.; Inoue, M.; Toda, K.; Ueda, H. Loss of peripheral morphine analgesia contributes to the reduced effectiveness of systemic morphine in neuropathic pain. J. Pharmacol. Exp. Ther. 2004, 309, 380–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, M.; Obata, H.; Saito, S. Peripheral nerve injury reduces analgesic effects of systemic morphine via spinal 5-hydroxytryptamine 3 receptors. Anesthesiology 2014, 121, 362–371. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.L.; Yu, L.N.; Wang, Y.; Tang, L.H.; Peng, Y.N.; Cao, J.L.; Yan, M. Increased methylation of the MOR gene proximal promoter in primary sensory neurons plays a crucial role in the decreased analgesic effect of opioids in neuropathic pain. Mol. Pain 2014, 10, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roeckel, L.A.; Utard, V.; Reiss, D.; Mouheiche, J.; Maurin, H.; Robé, A.; Audouard, E.; Wood, J.N.; Goumon, Y.; Simonin, F.; et al. Morphine-induced hyperalgesia involves mu opioid receptors and the metabolite morphine-3-glucuronide. Sci. Rep. 2017, 7, 10406. [Google Scholar] [CrossRef]
- Green-Fulgham, S.M.; Ball, J.B.; Kwilasz, A.J.; Fabisiak, T.; Maier, S.F.; Watkins, L.R.; Grace, P.M. Oxycodone, fentanyl, and morphine amplify established neuropathic pain in male rats. Pain 2019, 160, 2634–2640. [Google Scholar] [CrossRef]
- Grace, P.M.; Strand, K.A.; Galer, E.L.; Urban, D.J.; Wang, X.; Baratta, M.V.; Fabisiak, T.J.; Anderson, N.D.; Cheng, K.; Greene, L.I.; et al. Morphine paradoxically prolongs neuropathic pain in rats by amplifying spinal NLRP3 inflammasome activation. Proc. Natl. Acad. Sci. USA 2016, 113, E3441–E3450. [Google Scholar] [CrossRef] [Green Version]
- Grace, P.M.; Strand, K.A.; Galer, E.L.; Rice, K.C.; Maier, S.F.; Watkins, L.R. Protraction of neuropathic pain by morphine is mediated by spinal damage associated molecular patterns (DAMPs) in male rats. Brain Behav. Immun. 2018, 72, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, W.; Kaneta, M.; Nagae, M.; Yuzuhara, A.; Li, X.; Suzuki, H.; Hanagata, M.; Kitaoka, S.; Suto, W.; Kusunoki, Y.; et al. Mice with neuropathic pain exhibit morphine tolerance due to a decrease in the morphine concentration in the brain. Eur. J. Pharm. Sci. 2016, 92, 298–304. [Google Scholar] [CrossRef]
- Latapy, C.; Beaulieu, J.M. β-Arrestins in the central nervous system. Prog. Mol. Biol. Transl. Sci. 2013, 118, 267–295. [Google Scholar]
- He, L.; Fong, J.; von Zastrow, M.; Whistler, J.L. Regulation of opioid receptor trafficking and morphine tolerance by receptor oligomerization. Cell 2002, 108, 271–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kliewer, A.; Schmiedel, F.; Sianati, S.; Bailey, A.; Bateman, J.T.; Levitt, E.S.; Williams, J.T.; Christie, M.J.; Schulz, S. Phosphorylation-deficient G-protein-biased μ-opioid receptors improve analgesia and diminish tolerance but worsen opioid side effects. Nat. Commun. 2019, 10, 367. [Google Scholar] [CrossRef] [Green Version]
- Quillinan, N.; Lau, E.K.; Virk, M.; von Zastrow, M.; Williams, J.T. Recovery from mu-opioid receptor desensitization after chronic treatment with morphine and methadone. J. Neurosci. 2011, 31, 4434–4443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.A.; Bartlett, S.; He, L.; Nielsen, C.K.; Chang, A.M.; Kharazia, V.; Waldhoer, M.; Ou, C.J.; Taylor, S.; Ferwerda, M.; et al. Morphine-induced receptor endocytosis in a novel knockin mouse reduces tolerance and dependence. Curr. Biol. 2008, 18, 129–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McPherson, J.; Rivero, G.; Baptist, M.; Llorente, J.; Al-Sabah, S.; Krasel, C.; Dewey, W.L.; Bailey, C.P.; Rosethorne, E.M.; Charlton, S.J.; et al. μ-opioid receptors: Correlation of agonist efficacy for signalling with ability to activate internalization. Mol. Pharmacol. 2010, 78, 756–766. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Chen, R.; Huang, M.; Wang, W.; Luo, L.; Kim, D.K.; Jiang, W.; Xu, T. DAMGO-induced μ opioid receptor internalization and recycling restore morphine sensitivity in tolerant rat. Eur. J. Pharmacol. 2020, 878, 173118. [Google Scholar] [CrossRef]
- Bohn, L.M.; Lefkowitz, R.J.; Caron, M.G. Differential mechanisms of morphine antinociceptive tolerance revealed in (beta)arrestin-2 knock-out mice. J. Neurosci. 2002, 22, 10494–10500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, V.C.; Chieng, B.; Azriel, Y.; Christie, M.J. Cellular morphine tolerance produced by βarrestin-2-dependent impairment of μ-opioid receptor resensitization. J. Neurosci. 2011, 31, 7122–7130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muchhala, K.H.; Jacob, J.C.; Dewey, W.L.; Akbarali, H.I. Role of β-arrestin-2 in short- and long-term opioid tolerance in the dorsal root ganglia. Eur. J. Pharmacol. 2021, 899, 174007. [Google Scholar] [CrossRef]
- Krames, E.S. The role of the dorsal root ganglion in the development of neuropathic pain. Pain Med. 2014, 15, 1669–1685. [Google Scholar] [CrossRef] [PubMed]
- Berger, A.A.; Liu, Y.; Possoit, H.; Rogers, A.C.; Moore, W.; Gress, K.; Cornett, E.M.; Kaye, A.D.; Imani, F.; Sadegi, K.; et al. Dorsal root ganglion (DRG) and chronic pain. Anesth. Pain Med. 2021, 11, e113020. [Google Scholar] [CrossRef] [PubMed]
- Kohno, T.; Ji, R.R.; Ito, N.; Allchorne, A.J.; Befort, K.; Karchewski, L.A.; Woolf, C.J. Peripheral axonal injury results in reduced mu opioid receptor pre- and post-synaptic action in the spinal cord. Pain 2005, 117, 77–87. [Google Scholar] [CrossRef]
- Zhang, X.; Bao, L.; Shi, T.J.; Ju, G.; Elde, R.; Hökfelt, T. Down-regulation of mu-opioid receptors in rat and monkey dorsal root ganglion neurons and spinal cord after peripheral axotomy. Neuroscience 1998, 82, 223–240. [Google Scholar] [CrossRef]
- Obara, I.; Gunduz Cinar, O.; Starowicz, K.; Benyhe, S.; Borsodi, A.; Przewlocka, B. Agonist-dependent attenuation of mu-opioid receptor-mediated G-protein activation in the dorsal root ganglia of neuropathic rats. J. Neural Transm. 2010, 117, 421–429. [Google Scholar] [CrossRef]
- Santos, F.M.; Silva, J.T.; Rocha, I.R.C.; Martins, D.O.; Chacur, M. Non-pharmacological treatment affects neuropeptide expression in neuropathic pain model. Brain Res. 2018, 1687, 60–65. [Google Scholar] [CrossRef]
- Zheng, B.X.; Malik, A.; Xiong, M.; Bekker, A.; Tao, Y.X. Nerve trauma-caused downregulation of opioid receptors in primary afferent neurons: Molecular mechanisms and potential managements. Exp. Neurol. 2021, 337, 113572. [Google Scholar] [CrossRef]
- Zhao, J.Y.; Liang, L.; Gu, X.; Li, Z.; Wu, S.; Sun, L.; Atianjoh, F.E.; Feng, J.; Mo, K.; Jia, S.; et al. DNA methyltransferase DNMT3a contributes to neuropathic pain by repressing Kcna2 in primary afferent neurons. Nat. Commun. 2017, 8, 14712. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhao, J.Y.; Gu, X.; Liang, L.; Wu, S.; Mo, K.; Feng, J.; Guo, W.; Zhang, J.; Bekker, A.; et al. Nerve injury-induced epigenetic silencing of opioid receptors controlled by DNMT3a in primary afferent neurons. Pain 2017, 158, 1153–1165. [Google Scholar] [CrossRef]
- Mo, K.; Wu, S.; Gu, X.; Xiong, M.; Cai, W.; Atianjoh, F.E.; Jobe, E.E.; Zhao, X.; Tu, W.F.; Tao, Y.X. MBD1 contributes to the genesis of acute pain and neuropathic pain by epigenetic silencing of Oprm1 and Kcna2 genes in primary sensory neurons. J. Neurosci. 2018, 38, 9883–9899. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Wei, G.; Ji, F.; Jia, S.; Wu, S.; Guo, X.; He, L.; Pan, Z.; Miao, X.; Mao, Q.; et al. TET1 overexpression mitigates neuropathic pain through rescuing the expression of μ-opioid receptor and Kv1.2 in the primary sensory neurons. Neurotherapeutics 2019, 16, 491–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, L.; Zhao, J.Y.; Gu, X.; Wu, S.; Mo, K.; Xiong, M.; Marie Lutz, B.; Bekker, A.; Tao, Y.X. G9a inhibits CREB-triggered expression of mu opioid receptor in primary sensory neurons following peripheral nerve injury. Mol. Pain 2016, 12, 1744806916682242. [Google Scholar] [CrossRef] [Green Version]
- Liang, L.; Gu, X.; Zhao, J.Y.; Wu, S.; Miao, X.; Xiao, J.; Mo, K.; Zhang, J.; Lutz, B.M.; Bekker, A.; et al. G9a participates in nerve injury-induced Kcna2 downregulation in primary sensory neurons. Sci. Rep. 2016, 6, 37704. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chen, S.R.; Laumet, G.; Chen, H.; Pan, H.L. Nerve injury diminishes opioid analgesia through lysine methyltransferase-mediated transcriptional repression of μ-opioid receptors in primary sensory neurons. J. Biol. Chem. 2016, 291, 8475–8485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Liang, L.; Miao, X.; Wu, S.; Cao, J.; Tao, B.; Mao, Q.; Mo, K.; Xiong, M.; Lutz, B.M.; et al. Contribution of the suppressor of variegation 3-9 homolog 1 in dorsal root ganglia and spinal cord dorsal horn to nerve injury-induced nociceptive hypersensitivity. Anesthesiology 2016, 125, 765–778. [Google Scholar] [CrossRef] [Green Version]
- Pan, Z.; Du, S.; Wang, K.; Guo, X.; Mao, Q.; Feng, X.; Huang, L.; Wu, S.; Hou, B.; Chang, Y.J.; et al. Downregulation of a dorsal root ganglion-specifically enriched long noncoding RNA is required for neuropathic pain by negatively regulating RALY-triggered Ehmt2 expression. Adv. Sci. 2021, 8, e2004515. [Google Scholar] [CrossRef]
- Uchida, H.; Ma, L.; Ueda, H. Epigenetic gene silencing underlies C-fiber dysfunctions in neuropathic pain. J. Neurosci. 2010, 30, 4806–4814. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.; Tang, J.; Ding, T.; Chen, L.; Wang, W.; Mei, X.P.; He, X.T.; Wang, W.; Zhang, L.D.; Dong, Y.L.; et al. Neuron-restrictive silencer factor-mediated downregulation of μ-opioid receptor contributes to the reduced morphine analgesia in bone cancer pain. Pain 2017, 158, 879–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Mao, Y.; Liang, L.; Wu, S.; Yuan, J.; Mo, K.; Cai, W.; Mao, Q.; Cao, J.; Bekker, A.; et al. The transcription factor C/EBPβ in the dorsal root ganglion contributes to peripheral nerve trauma-induced nociceptive hypersensitivity. Sci. Signal 2017, 10, eaam5345. [Google Scholar] [CrossRef]
- Yuan, J.; Wen, J.; Wu, S.; Mao, Y.; Mo, K.; Li, Z.; Su, S.; Gu, H.; Ai, Y.; Bekker, A.; et al. Contribution of dorsal root ganglion octamer transcription factor 1 to neuropathic pain after peripheral nerve injury. Pain 2019, 160, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Cao, J.; Zhang, J.; Jia, S.; Wu, S.; Mo, K.; Wei, G.; Liang, L.; Miao, X.; Bekker, A.; et al. Role of microRNA-143 in nerve injury-induced upregulation of Dnmt3a expression in primary sensory neurons. Front. Mol. Neurosci. 2017, 10, 350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Guo, X.; Sun, L.; Xiao, J.; Su, S.; Du, S.; Li, Z.; Wu, S.; Liu, W.; Mo, K.; et al. N6-Methyladenosine demethylase FTO contributes to neuropathic pain by stabilizing G9a expression in primary sensory neurons. Adv. Sci. 2020, 7, 1902402. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zheng, B.; Du, S.; Han, G.; Zhao, H.; Wu, S.; Jia, S.; Bachmann, T.; Bekker, A.; Tao, Y.X. Eukaryotic initiation factor 4 gamma 2 contributes to neuropathic pain through down-regulation of Kv1.2 and the mu opioid receptor in mouse primary sensory neurones. Br. J. Anaesth. 2021, 126, 706–719. [Google Scholar] [CrossRef]
- Liang, L.; Lutz, B.M.; Bekker, A.; Tao, Y.X. Epigenetic regulation of chronic pain. Epigenomics. 2015, 7, 235–245. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Gu, X.; Pan, Z.; Guo, X.; Liu, J.; Atianjoh, F.E.; Wu, S.; Mo, K.; Xu, B.; Liang, L.; et al. Contribution of DNMT1 to neuropathic pain genesis partially through epigenetically repressing Kcna2 in primary afferent neurons. J. Neurosci. 2019, 39, 6595–6607. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.; Zhou, J.; Jing, H.; Liao, M.; Lin, S.; Huang, Z.; Huang, T.; Zhong, J.; Wang, H. Functional roles of lncRNAs and its potential mechanisms in neuropathic pain. Clin. Epigenet. 2019, 11, 78. [Google Scholar] [CrossRef]
- Ng, E.K.; Li, R.; Shin, V.Y.; Siu, J.M.; Ma, E.S.; Kwong, A. MicroRNA-143 is downregulated in breast cancer and regulates DNA methyltransferases 3A in breast cancer cells. Tumour. Biol. 2014, 35, 2591–2598. [Google Scholar] [CrossRef] [Green Version]
- Lence, T.; Soller, M.; Roignant, J.Y. A fly view on the roles and mechanisms of the m6A mRNA modification and its players. RNA Biol. 2017, 14, 1232–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prévôt, D.; Darlix, J.L.; Ohlmann, T. Conducting the initiation of protein synthesis: The role of eIF4G. Biol. Cell 2003, 95, 141–156. [Google Scholar] [CrossRef]
- Chen, Y.; Sommer, C. Nociceptin and its receptor in rat dorsal root ganglion neurons in neuropathic and inflammatory pain models: Implications on pain processing. J. Peripher. Nerv. Syst. 2006, 11, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Zaveri, N. Nociceptin opioid receptor (NOP) as a therapeutic target: Progress in translation from preclinical research to clinical utility. J. Med. Chem. 2016, 59, 7011–7028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corder, G.; Tawfik, V.L.; Wang, D.; Sypek, E.I.; Low, S.A.; Dickinson, J.R.; Sotoudeh, C.; Clark, J.D.; Barres, B.A.; Bohlen, C.J.; et al. Loss of μ opioid receptor signaling in nociceptors, but not microglia, abrogates morphine tolerance without disrupting analgesia. Nat. Med. 2017, 23, 164–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walczak, J.S.; Pichette, V.; Leblond, F.; Desbiens, K.; Beaulieu, P. Behavioral, pharmacological and molecular characterization of the saphenous nerve partial ligation: A new model of neuropathic pain. Neuroscience 2005, 132, 1093–1102. [Google Scholar] [CrossRef] [PubMed]
- Porreca, F.; Tang, Q.B.; Bian, D.; Riedl, M.; Elde, R.; Lai, J. Spinal opioid mu receptor expression in lumbar spinal cord of rats following nerve injury. Brain Res. 1998, 795, 197–203. [Google Scholar] [CrossRef]
- Chen, S.R.; Sweigart, K.L.; Lakoski, J.M.; Pan, H.L. Functional mu opioid receptors are reduced in the spinal cord dorsal horn of diabetic rats. Anesthesiology 2002, 97, 1602–1608. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.R.; Pan, H.L. Antinociceptive effect of morphine, but not mu opioid receptor number, is attenuated in the spinal cord of diabetic rats. Anesthesiology 2003, 99, 1409–1414. [Google Scholar] [CrossRef]
- Jones, A.K.; Watabe, H.; Cunningham, V.J.; Jones, T. Cerebral decreases in opioid receptor binding in patients with central neuropathic pain measured by [11C]diprenorphine binding and PET. Eur. J. Pain. 2004, 8, 479–485. [Google Scholar] [CrossRef]
- Maarrawi, J.; Peyron, R.; Mertens, P.; Costes, N.; Magnin, M.; Sindou, M.; Laurent, B.; Garcia-Larrea, L. Differential brain opioid receptor availability in central and peripheral neuropathic pain. Pain 2007, 127, 183–194. [Google Scholar] [CrossRef]
- Rojewska, E.; Wawrzczak-Bargiela, A.; Szucs, E.; Benyhe, S.; Starnowska, J.; Mika, J.; Przewlocki, R.; Przewlocka, B. Alterations in the activity of spinal and thalamic opioid systems in a mice neuropathic pain model. Neuroscience 2018, 390, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Costa, A.R.; Carvalho, P.; Flik, G.; Wilson, S.P.; Reguenga, C.; Martins, I.; Tavares, I. Neuropathic pain induced alterations in the opioidergic modulation of a descending pain facilitatory area of the brain. Front. Cell. Neurosci. 2019, 13, 287. [Google Scholar] [CrossRef]
- Thompson, S.J.; Pitcher, M.H.; Stone, L.S.; Tarum, F.; Niu, G.; Chen, X.; Kiesewetter, D.O.; Schweinhardt, P.; Bushnell, M.C. Chronic neuropathic pain reduces opioid receptor availability with associated anhedonia in rat. Pain 2018, 159, 1856–1866. [Google Scholar] [CrossRef]
- Schembri, E. Are opioids effective in relieving neuropathic pain? SN Compr. Clin. Med. 2019, 1, 30–46. [Google Scholar] [CrossRef] [Green Version]
- Llorca-Torralba, M.; Pilar-Cuéllar, F.; da Silva Borges, G.; Mico, J.A.; Berrocoso, E. Opioid receptors mRNAs expression and opioids agonist-dependent G-protein activation in the rat brain following neuropathy. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2020, 99, 109857. [Google Scholar] [CrossRef]
- Niikura, K.; Narita, M.; Butelman, E.R.; Kreek, M.J.; Suzuki, T. Neuropathic and chronic pain stimuli downregulate central mu-opioid and dopaminergic transmission. Trends Pharmacol. Sci. 2010, 31, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Hoot, M.R.; Sim-Selley, L.J.; Selley, D.E.; Scoggins, K.L.; Dewey, W.L. Chronic neuropathic pain in mice reduces μ-opioid receptor-mediated G-protein activity in the thalamus. Brain Res. 2011, 1406, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connor, A.B. Neuropathic pain: Quality-of-life impact, costs and cost effectiveness of therapy. Pharmacoeconomics 2009, 27, 95–112. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| MOR |
| Central |
|
| Peripheral |
|
| DOR |
| Central |
|
| Peripheral |
|
| KOR |
| Central |
|
| Peripheral |
|
| Changes in Key Factors | Relative Targets | Changes in Opioid Receptors | Efficacy of Opioids after Blocking Changes | References | ||
|---|---|---|---|---|---|---|
| Epigenetic level | ||||||
| DNMTs | DNMT3a | ↑ | Oprm1 and Oprk1 | MOR ↓ KOR ↓ | ↑ | [104,105] |
| MBDs | MBD1 | ↑ | Oprm1 | MOR ↓ | ↑ | [106] |
| TETs | TET1 | ↓ | Oprm1 | MOR ↓ | ↑ | [107] |
| Histone methyltransferase | G9a | ↑ | Oprm1, Oprk1, and Oprd1 | MOR ↓ KOR ↓ DOR ↓ | ↑ | [108,109,110] |
| SUV39H1 | ↑ | Oprm1 | MOR ↓ | Untested | [111] | |
| Transcription level | ||||||
| LncRNA | DS-lncRNA | ↓ | Ehmt2/G9a | MOR ↓ KOR ↓ DOR ↓ | Untested | [112] |
| TFs | NRSF | ↑ | Oprm1 | MOR ↓ | ↑ | [113,114] |
| C/EBP β | ↑ | Ehmt2/G9a | MOR ↓ | ↑ | [115] | |
| OCT1 | ↑ | Dnmt3a mRNA | MOR ↓ | ↑ | [116] | |
| Post-transcriptional level | ||||||
| MicroRNA | miR-143 | ↓ | Dnmt3a mRNA | MOR ↓ | ↑ | [117] |
| RNA modification | m6A | ↓ | Ehmt2 mRNA | MOR ↓ | ↑ | [118] |
| Protein complex | eIF4F | ↑ | MOR | MOR ↓ | Untested | [119] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Chen, J.; Li, Y.-Q. The Downregulation of Opioid Receptors and Neuropathic Pain. Int. J. Mol. Sci. 2023, 24, 5981. https://doi.org/10.3390/ijms24065981
Li L, Chen J, Li Y-Q. The Downregulation of Opioid Receptors and Neuropathic Pain. International Journal of Molecular Sciences. 2023; 24(6):5981. https://doi.org/10.3390/ijms24065981
Chicago/Turabian StyleLi, Lin, Jing Chen, and Yun-Qing Li. 2023. "The Downregulation of Opioid Receptors and Neuropathic Pain" International Journal of Molecular Sciences 24, no. 6: 5981. https://doi.org/10.3390/ijms24065981