
StaR Is a Positive Regulator of Topoisomerase I Activity Involved in Supercoiling Maintenance in Streptococcus pneumoniae

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
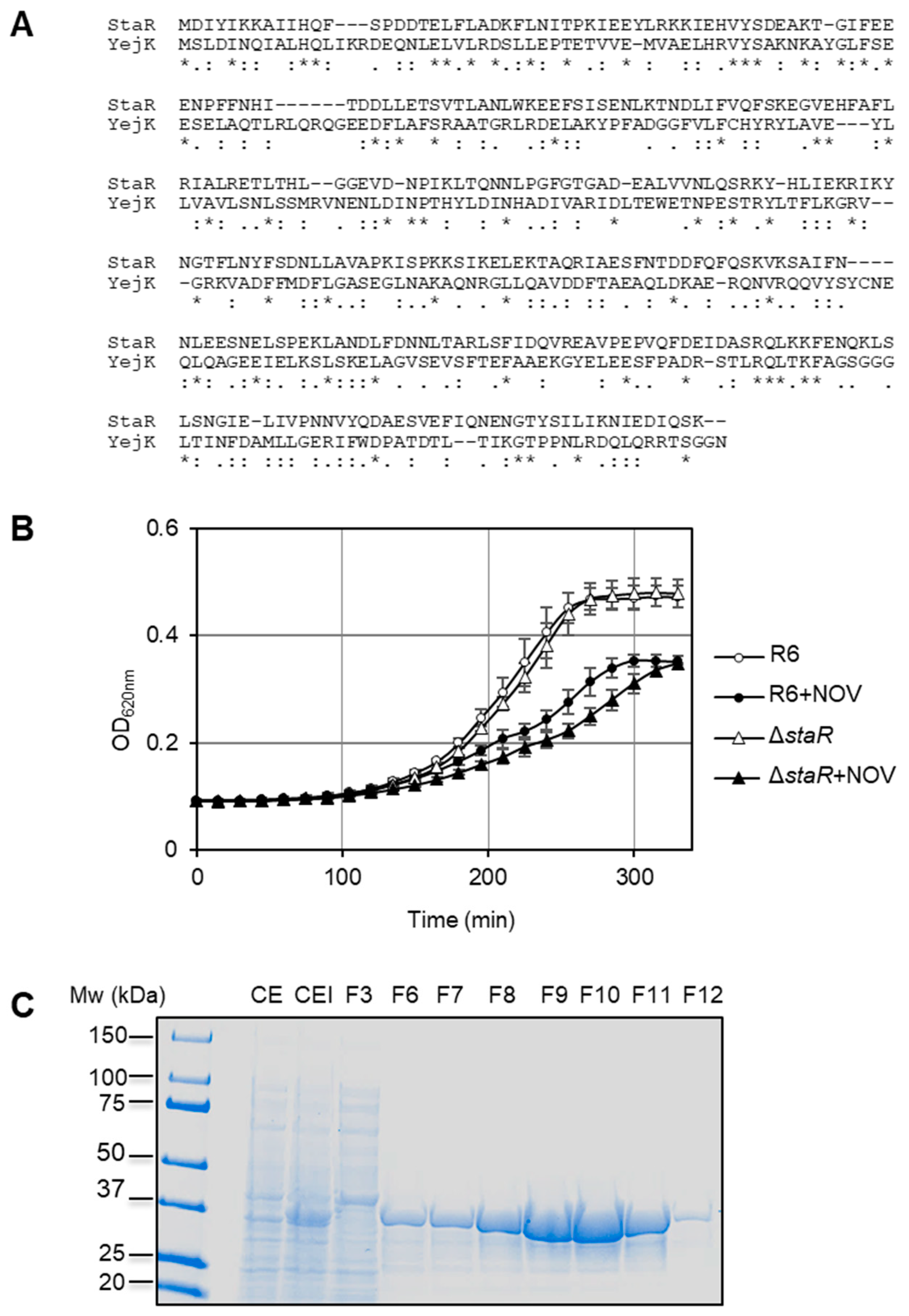
2.1. The Absence of StaR Affects Novobiocin Susceptibility in S. pneumoniae
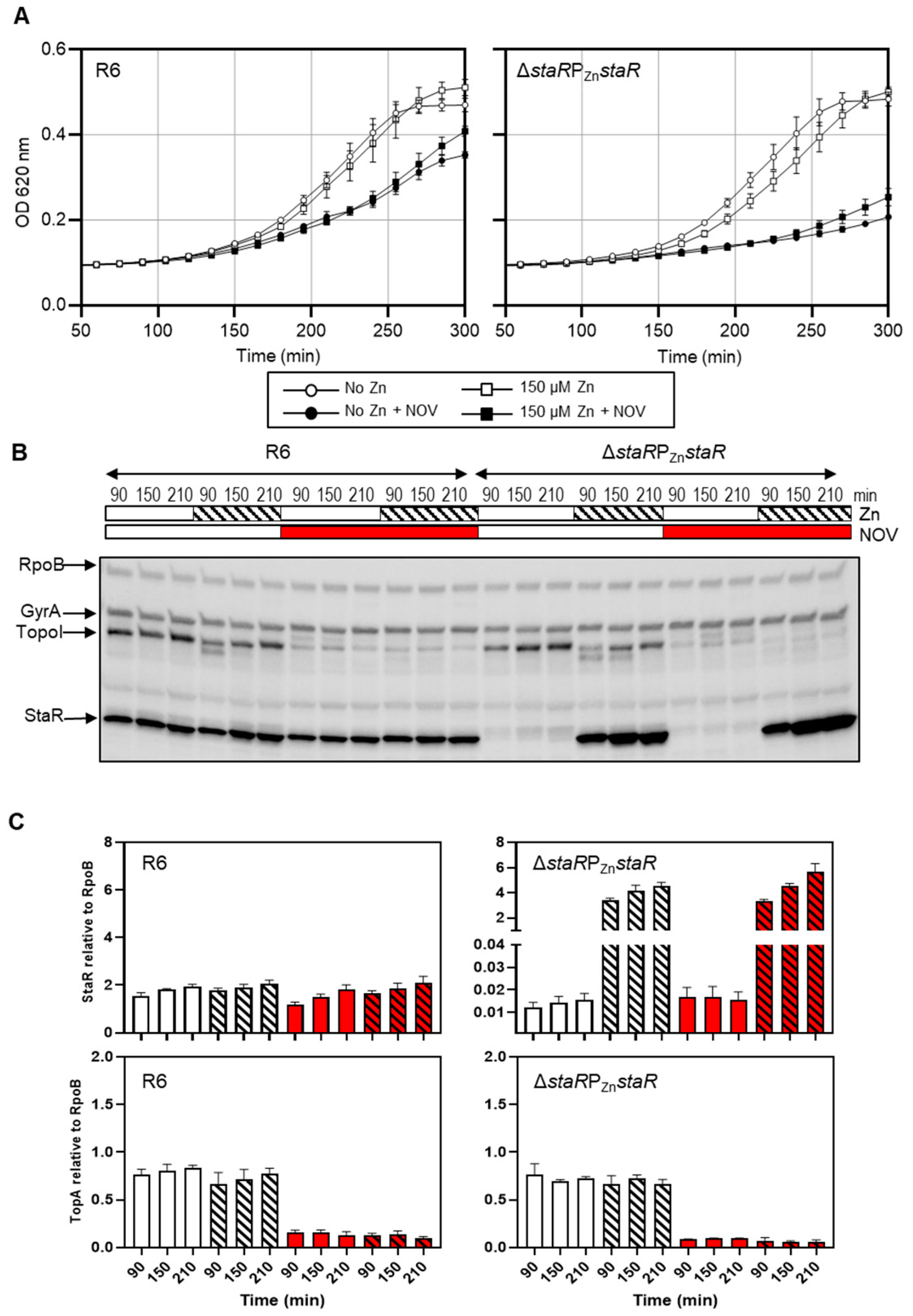
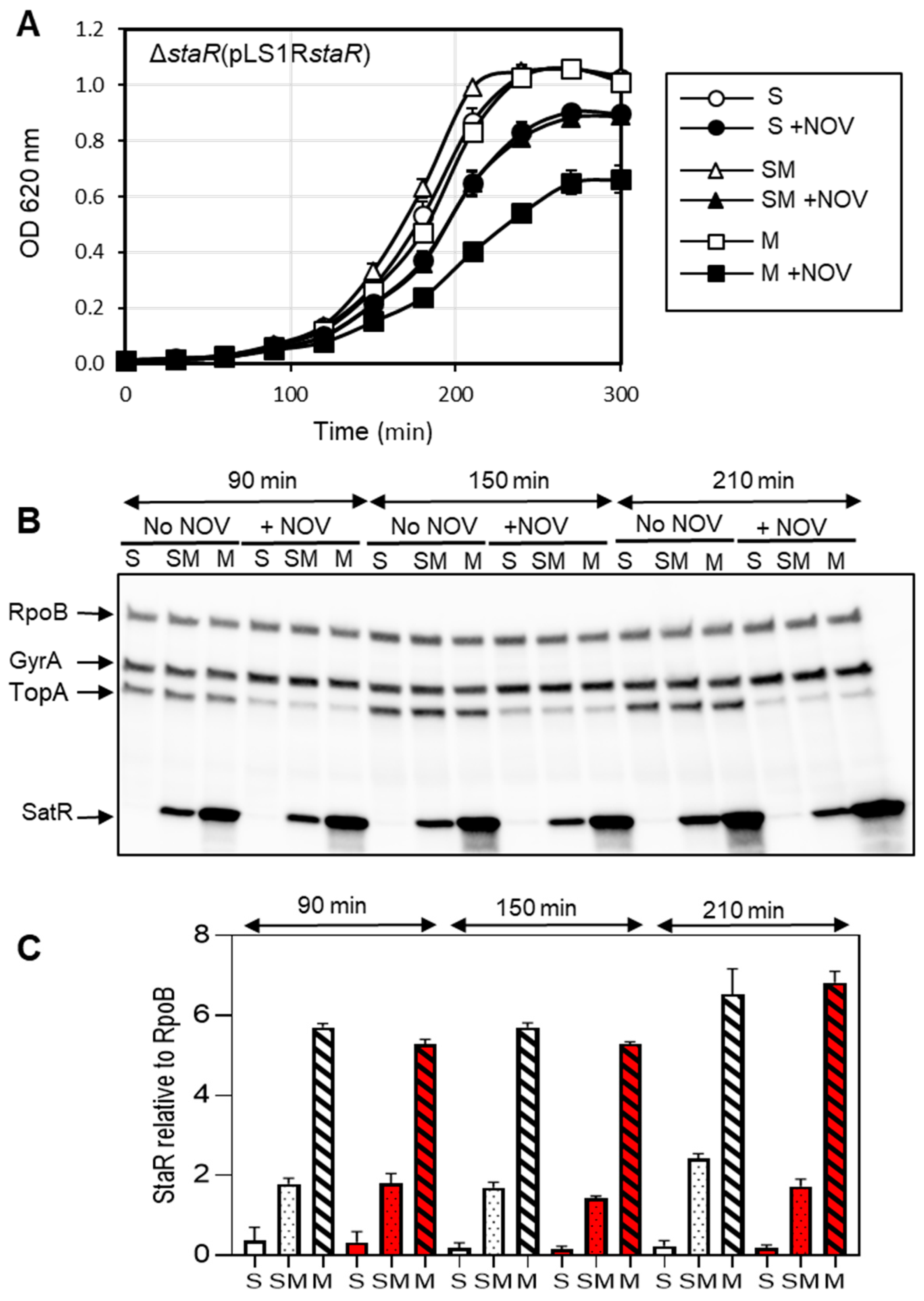
2.2. The Expression Level of StaR Affects Novobiocin Susceptibility
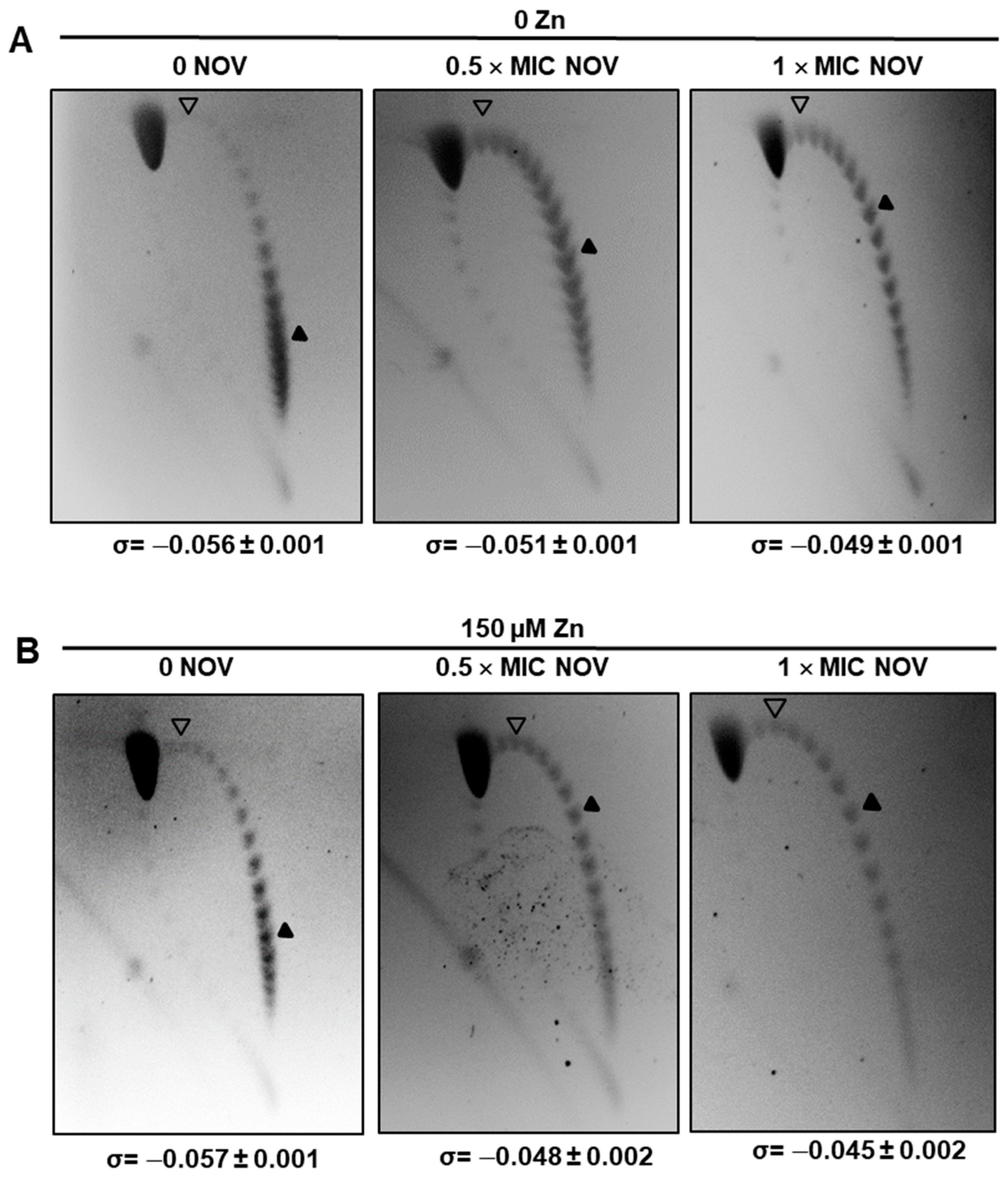
2.3. The Supercoiling Level Is Affected by StaR under Novobiocin Treatment
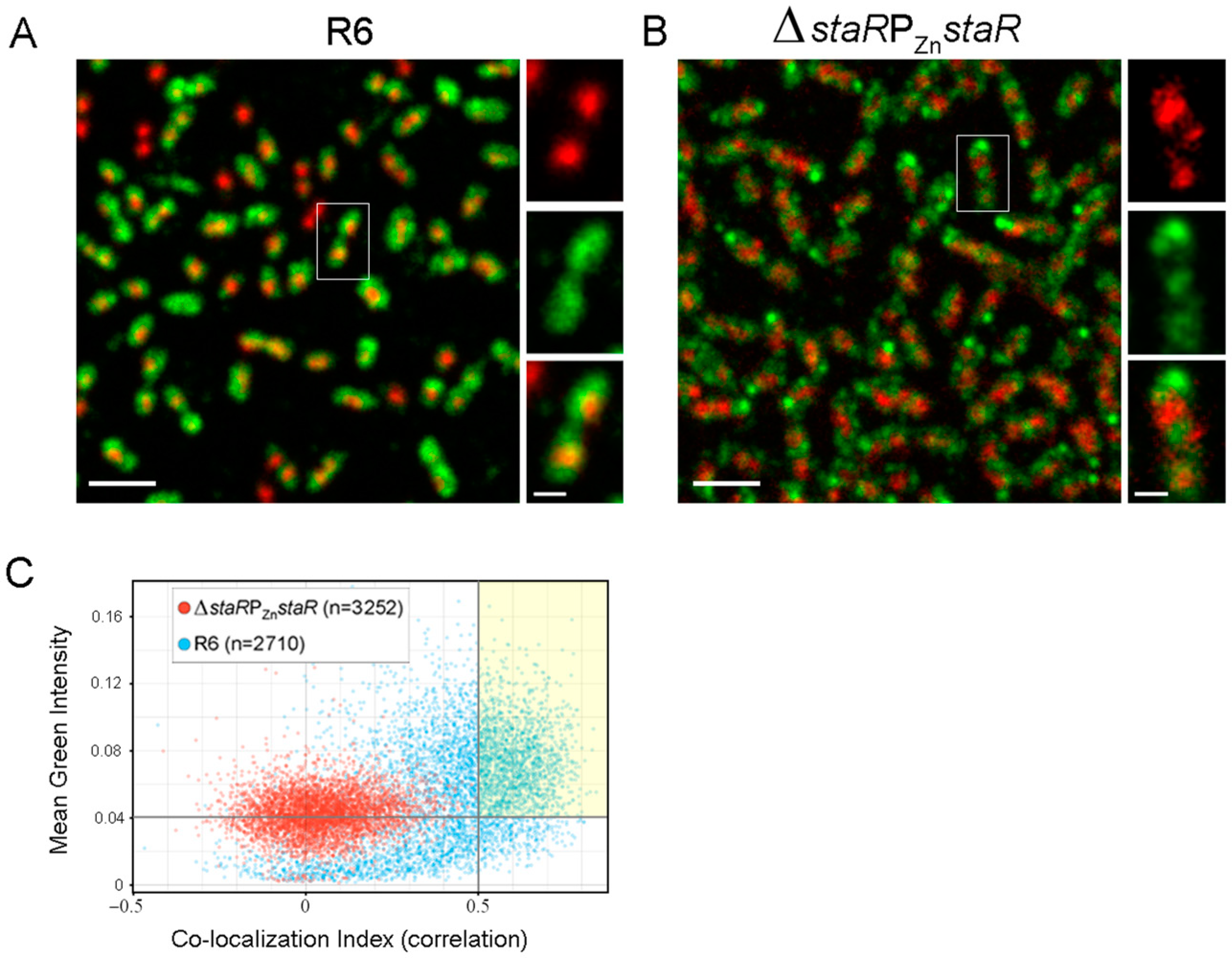
2.4. StaR Is a Nucleoid-Associated Protein
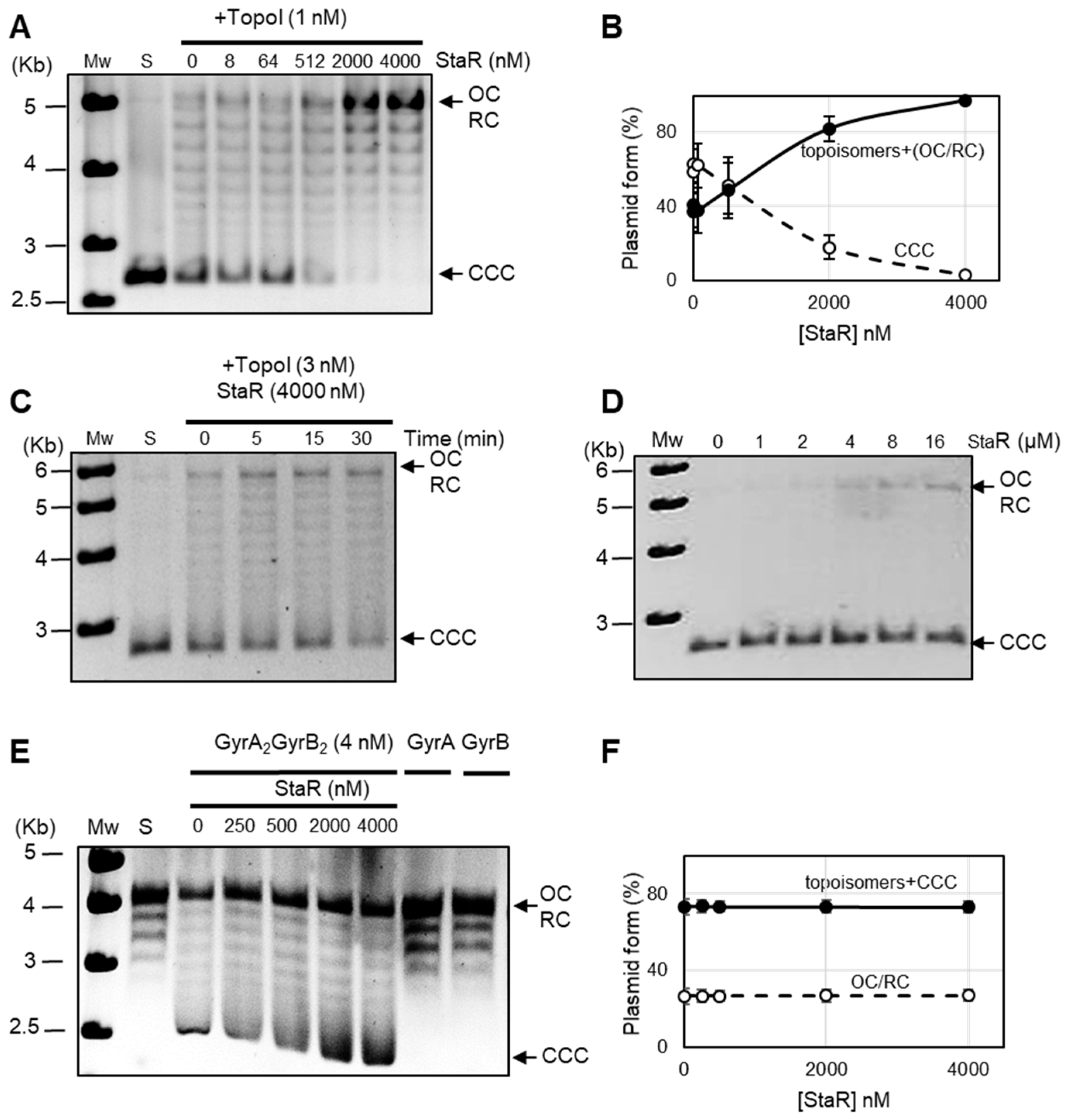
2.5. StaR Specifically Activates S. pneumoniae TopoI
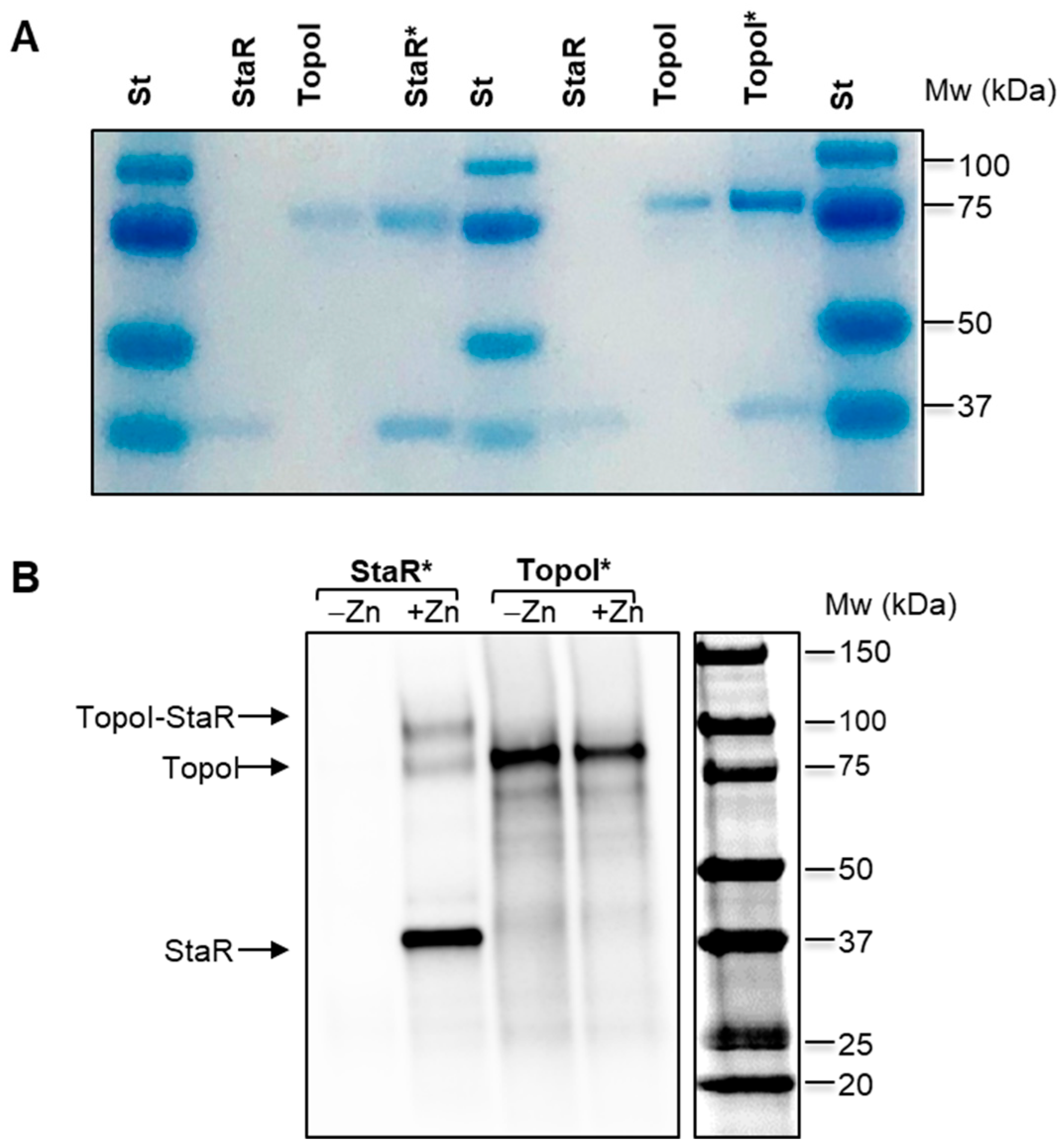
2.6. Physical Interaction between StaR and TopoI
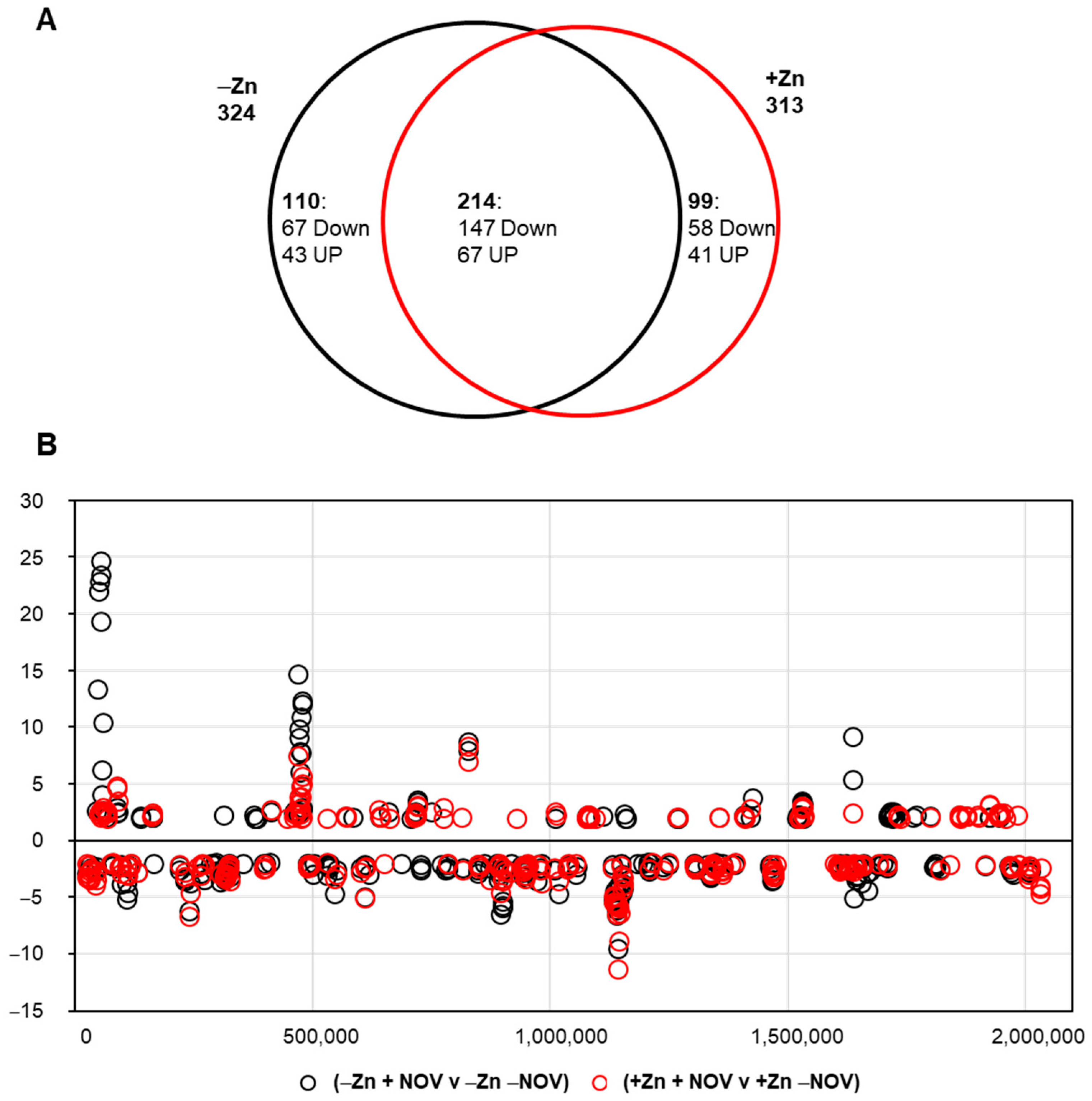
2.7. No Global Changes in Transcriptome Are Induced by the Absence or Presence of StaR
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Growth Conditions and Transformation
4.2. Construction of Strains and Plasmids
4.3. Cloning, Expression and Purification of Proteins
4.4. Confocal Microscopy
4.5. Western Blot Assays and Antibody Purification
4.6. DNA Topoisomerase Assays
4.7. Co-Immunoprecipitation (Co-IP) Assays
4.8. Analysis of the Topology of Plasmids
4.9. RNA Library Preparation for RNA-Seq
4.10. RNA Seq Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Holmes, V.F.; Cozzarelli, N.R. Closing the ring: Links between SMC proteins and chromosome partitioning, condensation, and supercoiling. Proc. Natl. Acad. Sci. USA 2000, 97, 1322–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Champoux, J.J. DNA Topoisomerases: Structure, Function, and Mechanism. Annu. Rev. Biochem. 2001, 70, 369–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gellert, M.; Mizuuchi, K.; O’Dea, M.H.; Nash, H.A. DNA gyrase: An enzyme that introduces superhelical turns into DNA. Proc. Natl. Acad. Sci. USA 1976, 73, 3872–3876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizuuchi, K.; O’Dea, M.H.; Gellert, M. DNA gyrase: Subunit structure and ATPase activity of the purified enzyme. Proc. Natl. Acad. Sci. USA 1978, 75, 5960–5963. [Google Scholar] [CrossRef] [Green Version]
- DiNardo, S.; Voelkel, K.A.; Sternglanz, R.; Reynolds, A.; Wright, A. Escherichia coli DNA topoisomerase I mutants have compensatory mutations in DNA gyrase genes. Cell 1982, 31, 43–51. [Google Scholar] [CrossRef]
- Pruss, G.J.; Manes, S.H.; Drlica, K. Escherichia coli DNA topoisomerase I mutants: Increased supercoiling is corrected by mutations near gyrase genes. Cell 1982, 31, 35–42. [Google Scholar] [CrossRef]
- Ferrándiz, M.-J.; Martín-Galiano, A.J.; Schvartzman, J.B.; de la Campa, A.G. The genome of Streptococcus pneumoniae is organized in topology-reacting gene clusters. Nucleic Acids Res. 2010, 38, 3570–3581. [Google Scholar] [CrossRef] [Green Version]
- de la Campa, A.G.; Ferrándiz, M.J.; Martín-Galiano, A.J.; García, M.T.; Tirado-Vélez, J.M. The Transcriptome of Streptococcus pneumoniae Induced by Local and Global Changes in Supercoiling. Front. Microbiol. 2017, 8, 1447. [Google Scholar] [CrossRef] [Green Version]
- García-López, M.; Megias, D.; Ferrándiz, M.-J.; de la Campa, A.G. The balance between gyrase and topoisomerase I activities determines levels of supercoiling, nucleoid compaction, and viability in bacteria. Front. Microbiol. 2022, 13, 1094692. [Google Scholar] [CrossRef]
- Wang, X.; Llopis, P.M.; Rudner, D.Z. Organization and segregation of bacterial chromosomes. Nat. Rev. Genet. 2013, 14, 191–203. [Google Scholar] [CrossRef] [Green Version]
- Dorman, C.J. Genome architecture and global gene regulation in bacteria: Making progress towards a unified model? Nat. Rev. Microbiol. 2013, 11, 349–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.F.; Wang, J.C. Supercoiling of the DNA template during transcription. Proc. Natl. Acad. Sci. USA 1987, 84, 7024–7027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, B.; Zhu, C.-X.; Ji, C.; Ahumada, A.; Tse-Dinh, Y.-C. Direct Interaction between Escherichia coli RNA Polymerase and the Zinc Ribbon Domains of DNA Topoisomerase I. J. Biol. Chem. 2003, 278, 30705–30710. [Google Scholar] [CrossRef] [Green Version]
- Ferrándiz, M.-J.; Hernández, P.; de la Campa, A.G. Genome-wide proximity between RNA polymerase and DNA topoisomerase I supports transcription in Streptococcus pneumoniae. PLoS Genet. 2021, 17, e1009542. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.; Sala, C.; Hegde, S.R.; Jha, R.K.; Cole, S.T.; Nagaraja, V. Transcription facilitated genome-wide recruitment of topoisomerase I and DNA gyrase. PLoS Genet. 2017, 13, e1006754. [Google Scholar] [CrossRef] [Green Version]
- Ferrándiz, M.-J.; Martín-Galiano, A.J.; Arnanz, C.; Camacho-Soguero, I.; Tirado-Vélez, J.-M.; de la Campa, A.G. An increase in negative supercoiling in bacteria reveals topology-reacting gene clusters and a homeostatic response mediated by the DNA topoisomerase I gene. Nucleic Acids Res. 2016, 44, 7292–7303. [Google Scholar] [CrossRef] [Green Version]
- Higgins, N.P.; Yang, X.; Fu, Q.; Roth, J.R. Surveying a supercoil domain by using the gamma delta resolution system in Salmonella typhimurium. J. Bacteriol. 1996, 178, 2825–2835. [Google Scholar] [CrossRef] [Green Version]
- Sinden, R.R.; Pettijohn, D.E. Chromosomes in living Escherichia coli cells are segregated into domains of supercoiling. Proc. Natl. Acad. Sci. USA 1981, 78, 224–228. [Google Scholar] [CrossRef] [Green Version]
- Worcel, A.; Burgi, E. On the structure of the folded chromosome of Escherichia coli. J. Mol. Biol. 1972, 71, 127–147. [Google Scholar] [CrossRef]
- Martín-Galiano, A.J.; Ferrándiz, M.J.; de la Campa, A.G. Bridging Chromosomal Architecture and Pathophysiology of Streptococcus pneumoniae. Genome Biol. Evol. 2017, 9, 350–361. [Google Scholar] [CrossRef] [Green Version]
- Ferrandiz, M.-J.; Arnanz, C.; Martin-Galiano, A.J.; Rodríguez-Martín, C.; De La Campa, A.G. Role of Global and Local Topology in the Regulation of Gene Expression in Streptococcus pneumoniae. PLoS ONE 2014, 9, e101574. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Pneumococcal conjugate vaccine for childhood immunization—WHO position paper. Wkly. Epidemiol. Rec. 2007, 82, 93–104. [Google Scholar]
- Jacobs, M.R.; Felmingham, D.; Appelbaum, P.C.; Grüneberg, R.N.; The Alexander Project Group. The Alexander Project 1998-2000: Susceptibility of pathogens isolated from community-acquired respiratory tract infection to commonly used antimicrobial agents. J. Antimicrob. Chemother. 2003, 52, 229–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domenech, A.; Tirado-Vélez, J.M.; Fenoll, A.; Ardanuy, C.; Yuste, J.; Liñares, J.; de la Campa, A.G. Fluoroquinolone-Resistant Pneumococci: Dynamics of Serotypes and Clones in Spain in 2012 Compared with Those from 2002 and 2006. Antimicrob. Agents Chemother. 2014, 58, 2393–2399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ardanuy, C.; de la Campa, A.G.; García, E.; Fenoll, A.; Calatayud, L.; Cercenado, E.; Pérez-Trallero, E.; Bouza, E.; Liñares, J. Spread of Streptococcus pneumoniae Serotype 8-ST63 Multidrug-Resistant Recombinant Clone, Spain. Emerg. Infect. Dis. 2014, 20, 1848–1856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balsalobre, L.; Ferrandiz, M.-J.; de Alba, G.; de la Campa, A.G. Nonoptimal DNA Topoisomerases Allow Maintenance of Supercoiling Levels and Improve Fitness of Streptococcus pneumoniae. Antimicrob. Agents Chemother. 2011, 55, 1097–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balsalobre, L.; Ferrándiz, M.J.; Liñares, J.; Tubau, F.; de la Campa, A.G. Viridans Group Streptococci Are Donors in Horizontal Transfer of Topoisomerase IV Genes to Streptococcus pneumoniae. Antimicrob. Agents Chemother. 2003, 47, 2072–2081. [Google Scholar] [CrossRef] [Green Version]
- de la Campa, A.G.; Ardanuy, C.; Balsalobre, L.; Pérez-Trallero, E.; Marimón, J.M.; Fenoll, A.; Liñares, J. Changes in Fluoroquinolone-ResistantStreptococcus pneumoniaafter 7-Valent Conjugate Vaccination, Spain. Emerg. Infect. Dis. 2009, 15, 905–911. [Google Scholar] [CrossRef]
- Alvarado, M.; Martin-Galiano, A.J.; Ferrandiz, M.-J.; Zaballos, Á.; De La Campa, A.G. Upregulation of the PatAB Transporter Confers Fluoroquinolone Resistance to Streptococcus pseudopneumoniae. Front. Microbiol. 2017, 8, 2074. [Google Scholar] [CrossRef] [Green Version]
- Amblar, M.; Zaballos, Á.; de la Campa, A.G. Role of PatAB Transporter in Efflux of Levofloxacin in Streptococcus pneumoniae. Antibiotics 2022, 11, 1837. [Google Scholar] [CrossRef]
- Dillon, S.C.; Dorman, C.J. Bacterial nucleoid-associated proteins, nucleoid structure and gene expression. Nat. Rev. Microbiol. 2010, 8, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Ferrándiz, M.-J.; Carreño, D.; Ayora, S.; de la Campa, A. HU of Streptococcus pneumoniae Is Essential for the Preservation of DNA Supercoiling. Front. Microbiol. 2018, 9, 493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minnen, A.; Attaiech, L.; Thon, M.; Gruber, S.; Jan-Willem Veening, J.-W. SMC is recruited to oriC by ParB and promotes chromosome segregation in Streptococcus pneumoniae. Mol. Microbiol. 2011, 81, 676–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín-Galiano, A.J.; Yuste, J.; Cercenado, M.I.; De La Campa, A.G. Inspecting the potential physiological and biomedical value of 44 conserved uncharacterised proteins of Streptococcus pneumoniae. BMC Genom. 2014, 15, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, L.D.; Rosner, J.L.; Zimmerman, S.B.; Esposito, D. Identification of Two New Proteins in Spermidine Nucleoids Isolated from Escherichia coli. J. Bacteriol. 1999, 181, 3842–3844. [Google Scholar] [CrossRef] [Green Version]
- Gellert, M.; Mizuuchi, K.; O’Dea, M.H.; Itoh, T.; Tomizawa, J.-I. Nalidixic acid resistance: A second genetic character involved in DNA gyrase activity. Proc. Natl. Acad. Sci. USA 1977, 74, 4772–4776. [Google Scholar] [CrossRef] [Green Version]
- Pruss, G.J.; Franco, R.J.; Chevalier, S.G.; Manes, S.H.; Drlica, K. Effects of DNA gyrase inhibitors in Escherichia coli topoisomerase I mutants. J. Bacteriol. 1986, 168, 276–282. [Google Scholar] [CrossRef] [Green Version]
- Azam, T.A.; Hiraga, S.; Ishihama, A. Two types of localization of the DNA-binding proteins within the Escherichia coli nucleoid. Genes Cells 2000, 5, 613–626. [Google Scholar] [CrossRef]
- Nakanishi, A.; Oshida, T.; Matsushita, T.; Imajoh-Ohmi, S.; Ohnuki, T. Identification of DNA Gyrase Inhibitor (GyrI) in Escherichia coli. J. Biol. Chem. 1998, 273, 1933–1938. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.S.; Haakonsen, D.L.; Zeng, W.; Schumacher, M.A.; Laub, M.T. A Bacterial Chromosome Structuring Protein Binds Overtwisted DNA to Stimulate Type II Topoisomerases and Enable DNA Replication. Cell 2018, 175, 583–597.e23. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Mallick, B.; Nagaraja, V. Direct regulation of topoisomerase activity by a nucleoid-associated protein. Nucleic Acids Res. 2014, 42, 11156–11165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.; Marians, K.J. Characterization of the Nucleoid-associated Protein YejK. J. Biol. Chem. 2013, 288, 31503–31516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baranello, L.; Wojtowicz, D.; Cui, K.; Devaiah, B.N.; Chung, H.-J.; Chan-Salis, K.Y.; Guha, R.; Wilson, K.; Zhang, X.; Zhang, H.; et al. RNA Polymerase II Regulates Topoisomerase 1 Activity to Favor Efficient Transcription. Cell 2016, 165, 357–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Lacks, S.A.; López, P.; Greenberg, B.; Espinosa, M. Identification and analysis of genes for tetracycline resistance and replication functions in the broad-host-range plasmid pLS1. J. Mol. Biol. 1986, 192, 753–765. [Google Scholar] [CrossRef]
- Kloosterman, T.G.; Witwicki, R.M.; van der Kooi-Pol, M.M.; Bijlsma, J.J.E.; Kuipers, O.P. Opposite Effects of Mn2+ and Zn2+ on PsaR-Mediated Expression of the Virulence Genes pcpA, prtA, and psaBCA of Streptococcus pneumoniae. J. Bacteriol. 2008, 190, 5382–5393. [Google Scholar] [CrossRef] [Green Version]
- Escolano-Martínez, M.S.; Domenech, A.; Yuste, J.; Cercenado, M.I.; Ardanuy, C.; Liñares, J.; de la Campa, A.G.; Martin-Galiano, A.J. DiiA is a novel dimorphic cell wall protein of Streptococcus pneumoniae involved in invasive disease. J. Infect. 2016, 73, 71–81. [Google Scholar] [CrossRef]
- Ruiz-Masó, J.A.; López-Aguilar, C.; Nieto, C.; Sanz, M.; Burón, P.; Espinosa, M.; del Solar, G. Construction of a plasmid vector based on the pMV158 replicon for cloning and inducible gene expression in Streptococcus pneumoniae. Plasmid 2012, 67, 53–59. [Google Scholar] [CrossRef] [Green Version]
- García, M.T.; Blázquez, M.A.; Ferrandiz, M.-J.; Sanz, M.J.; Silva-Martín, N.; Hermoso, J.; de la Campa, A.G. New Alkaloid Antibiotics That Target the DNA Topoisomerase I of Streptococcus pneumoniae. J. Biol. Chem. 2011, 286, 6402–6413. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Moreira, E.; Balas, D.; González, I.; DE LA Campa, A.G. Fluoroquinolones Inhibit PreferentiallyStreptococcus pneumoniaeDNA Topoisomerase IV Than DNA Gyrase Native Proteins. Microb. Drug Resist. 2000, 6, 259–267. [Google Scholar] [CrossRef]
- Schvartzman, J.B.; Martínez-Robles, M.-L.; Hernández, P.; Krimer, D.B. Plasmid DNA Topology Assayed by Two-Dimensional Agarose Gel Electrophoresis. In DNA Electrophoresis; Methods in Molecular Biology; Humana: Totowa, NJ, USA, 2013; Volume 1054, pp. 121–132. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Role o Subrole | Common Name | R6 Locus (Gene) a | Relative Fold Variation b |
|---|---|---|---|
| Carbohydrate metabolism | Beta-galactosidase | spr0059 (bgaC) | −2.2 |
| Hypothetical proteins | spr0066 | 3.9 | |
| Transport: Carbohydrates | ABCT membrane permease | spr0082 | −2.2 |
| Hypothetical proteins | spr0085 | −2.0 | |
| Carbohydrate metabolism | 6-P-beta-glucosidase | spr0276 (celA) | −3.0 |
| Transcription factors | Antiterminator | spr0279 (bglG) | −3.2 |
| Transport: Carbohydrates | PTS sugar-specific EII component | spr0280 (celC) | −2.3 |
| Hypothetical proteins | spr0281 | −2.3 | |
| Transcription factors | Antiterminator | spr0504 | −2.2 |
| Pathogenesis | LPxTG protein | spr0561 (prtA) | 2.3 |
| Hypothetical proteins | spr0600 | −2.0 | |
| Hypothetical proteins | spr0601 | −2.1 | |
| Hypothetical proteins | spr0929 (staR) | 438.1 | |
| Transport: antibiotics | ABCT ATP-binding/membrane | spr1289 | −2.2 |
| Hypothetical proteins | spr1402 | 2.3 | |
| Transport: cations | ABCT ATP-binding-Mn | spr1492 (psaB) | 4.1 |
| Transport: cations | ABCT membrane permease -Mn | spr1493 (psaC) | 4.7 |
| Transport: cations | ABCT substrate-binding-Mn | spr1494 (psaA) | 4.8 |
| Carbohydrate metabolism | Galactose-1-P uridylyltransferase | spr1667 | 2.1 |
| Carbohydrate metabolism | Galactokinase | spr1668 (galK) | 2.3 |
| Carbohydrate metabolism | Alcohol dehydrogenase | spr1670 (adhB) | 73.7 |
| Hypothetical proteins | spr1671 | 105.3 | |
| Transport: cations | Cation diffusion facilitator | spr1672 (czcD) | 507.5 |
| Carbohydrate metabolism | Dextran glucosidase | spr1698 | −3.2 |
| Hypothetical proteins | spr1940 | −2.2 | |
| Hypothetical proteins | spr1966 | 2.6 | |
| Transport: other | Glycerol uptake facilitator protein | spr1988 (glpF) | −2.2 |
| Carbohydrate metabolism | Glycerol-3-P dehydrogenase | spr1989 | −2.6 |
| Carbohydrate metabolism | Glycerol-3-P dehydrogenase | spr1990 | −3.1 |
| Carbohydrate metabolism | Glycerol kinase | spr1991 (glpK) | −2.0 |
| Hypothetical proteins | spr2037 | 3.1 | |
| Hypothetical proteins | spr2038 | 3.4 | |
| Hypothetical proteins | spr2039 | 3.4 | |
| Hypothetical proteins | spr2040 | 2.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Vasconcelos Junior, A.A.; Tirado-Vélez, J.M.; Martín-Galiano, A.J.; Megias, D.; Ferrándiz, M.-J.; Hernández, P.; Amblar, M.; de la Campa, A.G. StaR Is a Positive Regulator of Topoisomerase I Activity Involved in Supercoiling Maintenance in Streptococcus pneumoniae. Int. J. Mol. Sci. 2023, 24, 5973. https://doi.org/10.3390/ijms24065973
de Vasconcelos Junior AA, Tirado-Vélez JM, Martín-Galiano AJ, Megias D, Ferrándiz M-J, Hernández P, Amblar M, de la Campa AG. StaR Is a Positive Regulator of Topoisomerase I Activity Involved in Supercoiling Maintenance in Streptococcus pneumoniae. International Journal of Molecular Sciences. 2023; 24(6):5973. https://doi.org/10.3390/ijms24065973
Chicago/Turabian Stylede Vasconcelos Junior, Antonio A., Jose M. Tirado-Vélez, Antonio J. Martín-Galiano, Diego Megias, María-José Ferrándiz, Pablo Hernández, Mónica Amblar, and Adela G. de la Campa. 2023. "StaR Is a Positive Regulator of Topoisomerase I Activity Involved in Supercoiling Maintenance in Streptococcus pneumoniae" International Journal of Molecular Sciences 24, no. 6: 5973. https://doi.org/10.3390/ijms24065973