
Nitric Oxide Affects Heme Oxygenase-1, Hepcidin, and Transferrin Receptor Expression in the Placenta
 , and
, and 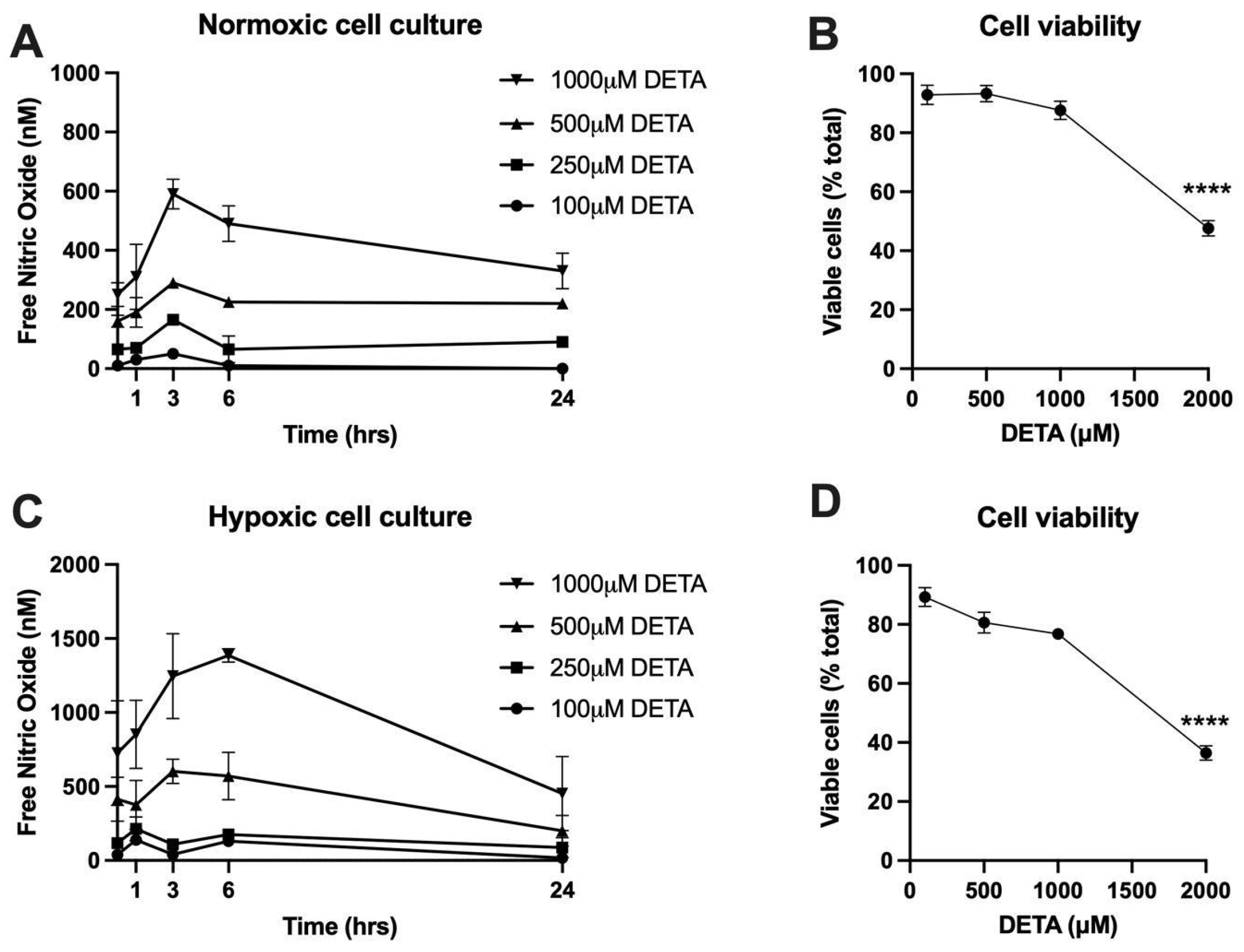{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Determination of NO Donor Dose
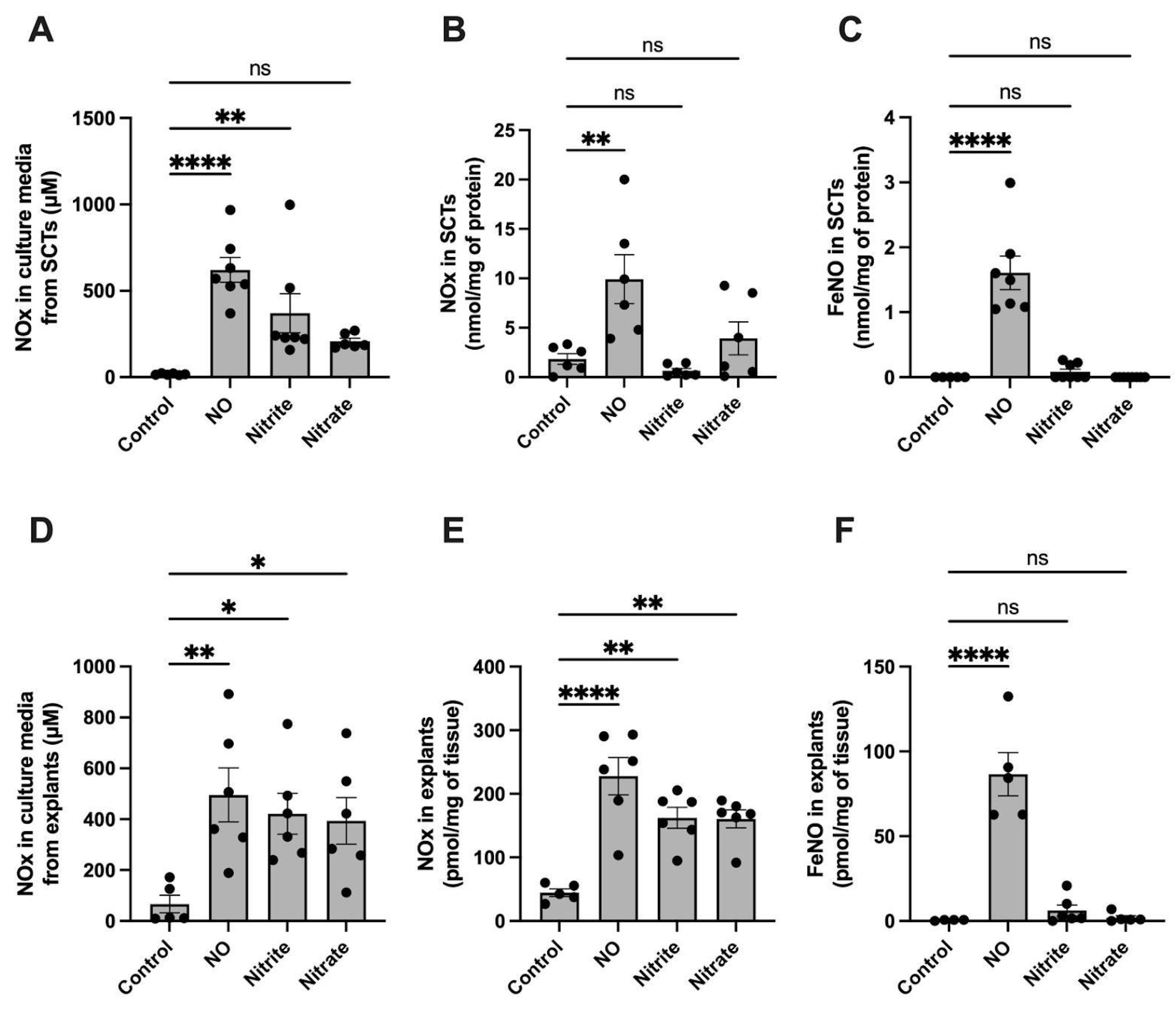
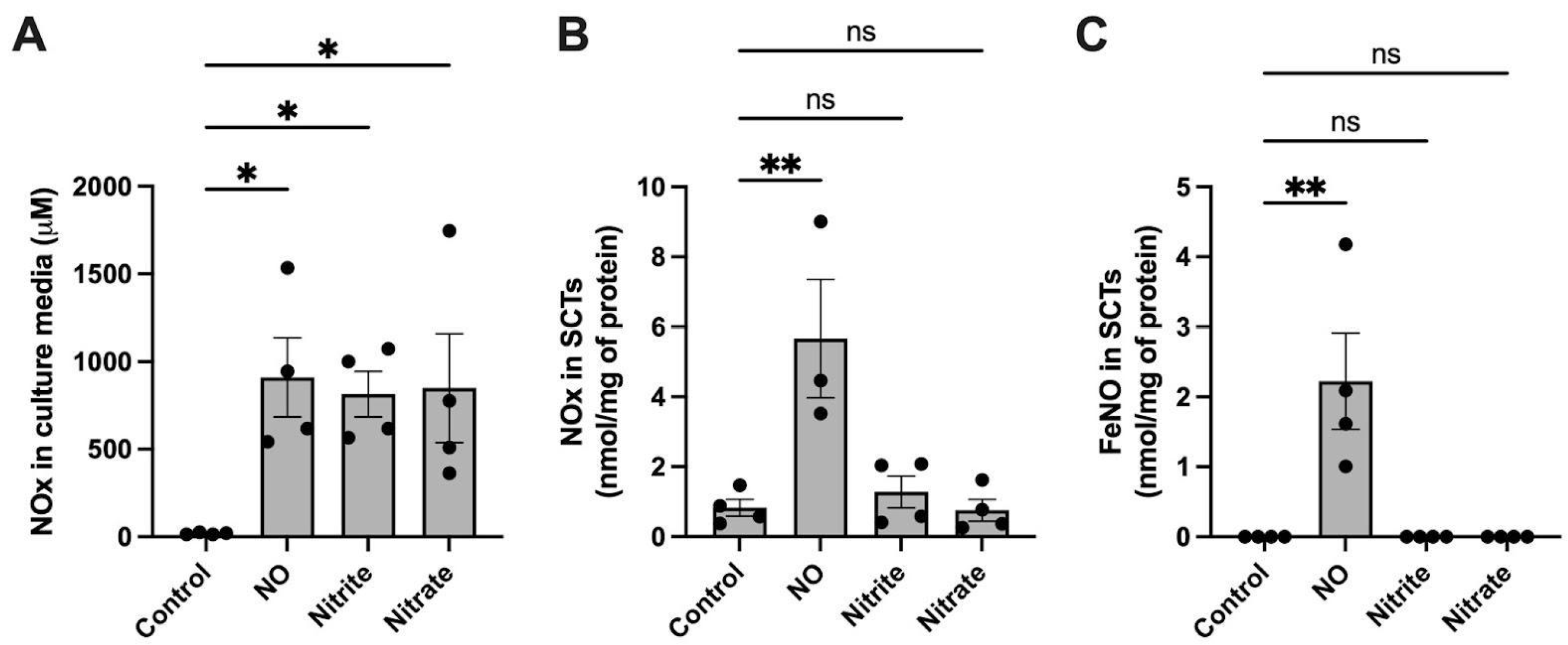
2.2. FeNO Formation in Placental Cell and Villous Explant Culture
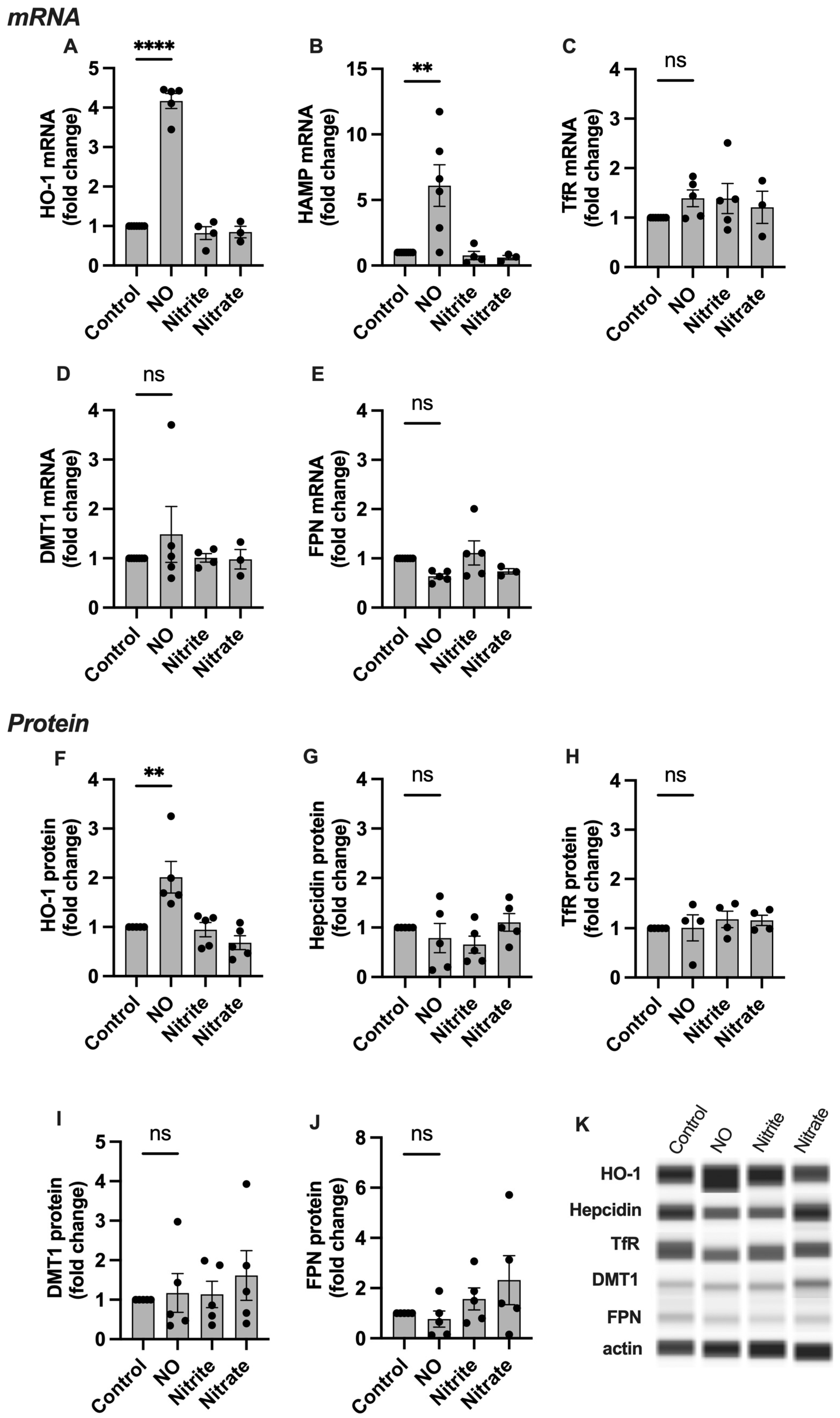
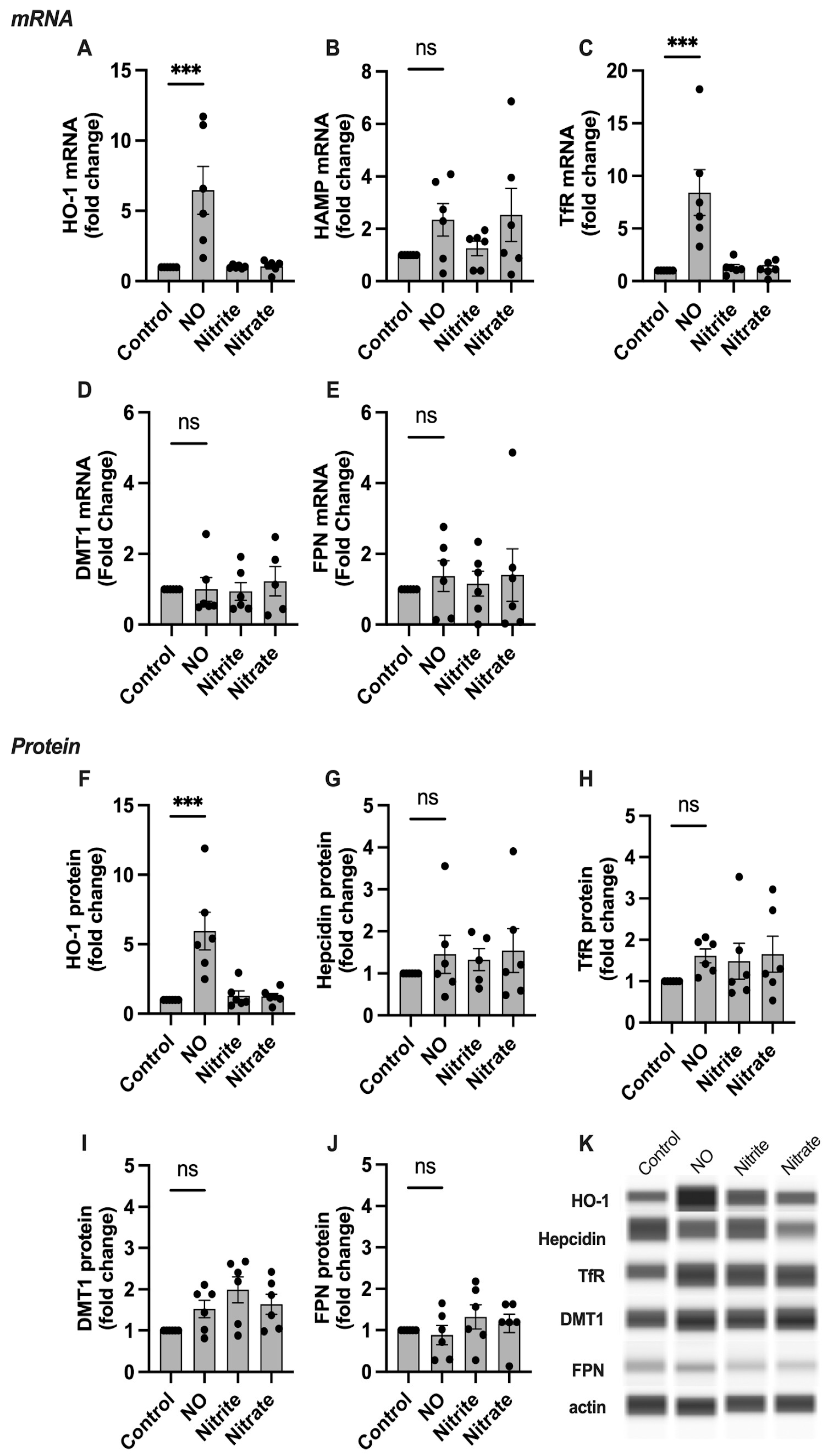
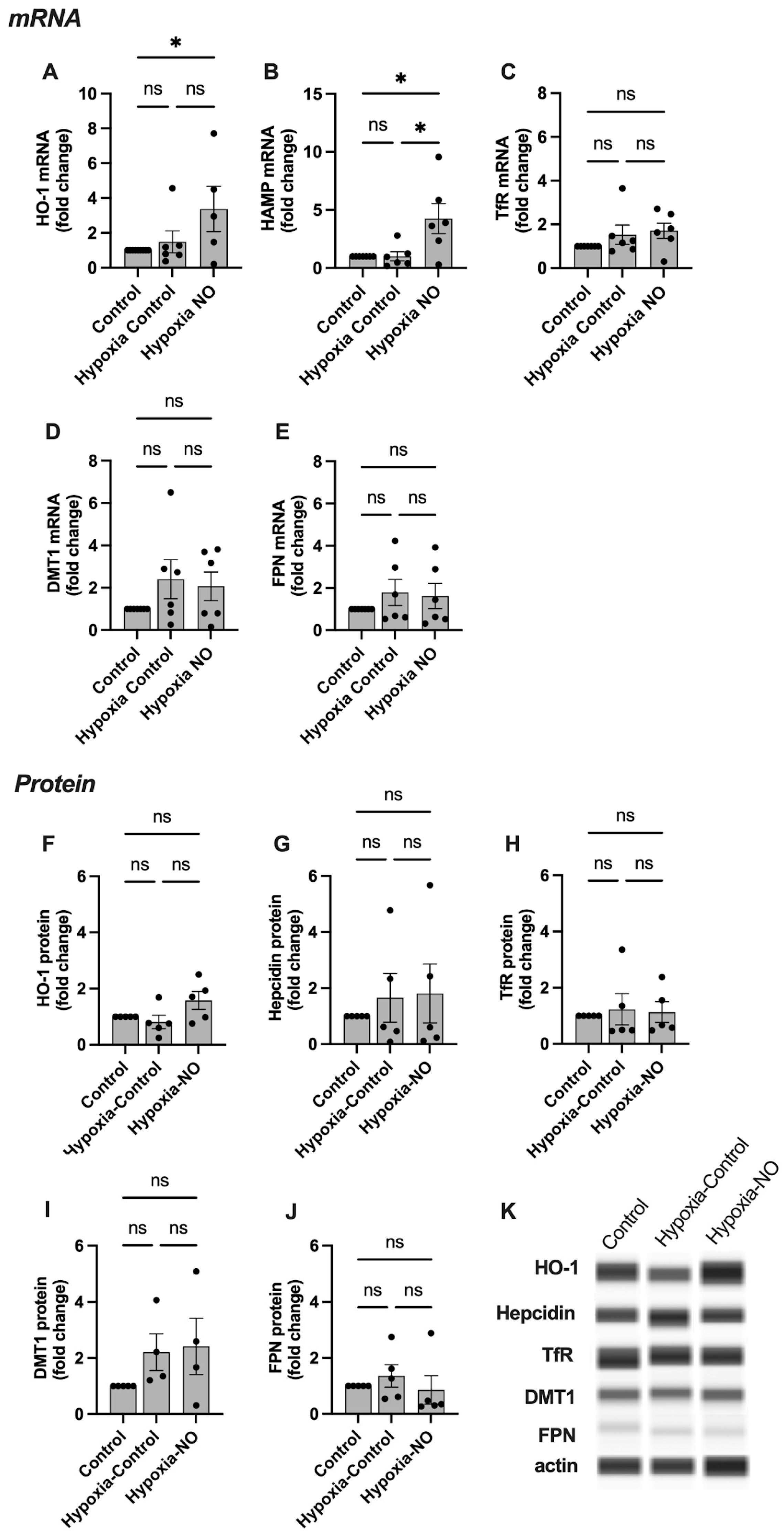
2.3. HO-1, Hepcidin, and TfR Levels Change in Response to NO
3. Discussion
4. Materials and Methods
4.1. BeWo Cell Culture
4.2. Placental Explant Culture
4.3. NO and Hypoxia Treatments
4.4. Cell Viability
4.5. Cell and Placental Explant Processing for NO and NOx Measurements
4.6. NO and NOx Measurement
4.7. qPCR
4.8. Protein Isolation and Quantification
4.9. Western Blot
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Sangkhae, V.; Fisher, A.L.; Wong, S.; Koenig, M.D.; Tussing-Humphreys, L.; Chu, A.; Lelić, M.; Ganz, T.; Nemeth, E. Effects of maternal iron status on placental and fetal iron homeostasis. J. Clin. Investig. 2020, 130, 625–640. [Google Scholar] [CrossRef] [PubMed]
- Barad, A.; Guillet, R.; Pressman, E.K.; Katzman, P.J.; Miller, R.K.; Darrah, T.H.; O’Brien, K.O. Placental Iron Content Is Lower than Previously Estimated and Is Associated with Maternal Iron Status in Women at Greater Risk of Gestational Iron Deficiency and Anemia. J. Nutr. 2022, 152, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Thornburg, K.L.; Marshall, N. The placenta is the center of the chronic disease universe. Am. J. Obstet. Gynecol. 2015, 213, S14–S20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sangkhae, V.; Nemeth, E. Placental iron transport: The mechanism and regulatory circuits. Free Radic. Biol. Med. 2019, 133, 254–261. [Google Scholar] [CrossRef]
- Richardson, D.R.; Lok, H.C. The nitric oxide-iron interplay in mammalian cells: Transport and storage of dinitrosyl iron complexes. Biochim. Biophys. Acta 2008, 1780, 638–651. [Google Scholar] [CrossRef]
- Erlandsson, L.; Masoumi, Z.; Hansson, L.R.; Hansson, S.R. The roles of free iron, heme, haemoglobin, and the scavenger proteins haemopexin and alpha-1-microglobulin in preeclampsia and fetal growth restriction. J. Intern. Med. 2021, 290, 952–968. [Google Scholar] [CrossRef] [PubMed]
- Yanatori, I.; Richardson, D.R.; Toyokuni, S.; Kishi, F. The new role of poly (rC)-binding proteins as iron transport chaperones: Proteins that could couple with inter-organelle interactions to safely traffic iron. Biochim. Biophys. Acta Gen. Subj. 2020, 1864, 129685. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.D. Breathing new life into nitric oxide signaling: A brief overview of the interplay between oxygen and nitric oxide. Redox Biol. 2015, 5, 225–233. [Google Scholar] [CrossRef] [Green Version]
- Tiravanti, E.; Samouilov, A.; Zweier, J.L. Nitrosyl-heme complexes are formed in the ischemic heart: Evidence of nitrite-derived nitric oxide formation, storage, and signaling in post-ischemic tissues. J. Biol. Chem. 2004, 279, 11065–11073. [Google Scholar] [CrossRef] [Green Version]
- Vanin, A.F. Dinitrosyl iron complexes with thiol-containing ligands as a ‘working form’ of endogenous nitric oxide. Nitric Oxide 2016, 54, 15–29. [Google Scholar] [CrossRef]
- Drapier, J.C.; Hirling, H.; Wietzerbin, J.; Kaldy, P.; Kühn, L.C. Biosynthesis of nitric oxide activates iron regulatory factor in macrophages. EMBO J. 1993, 12, 3643–3649. [Google Scholar] [CrossRef] [PubMed]
- Weiss, G.; Goossen, B.; Doppler, W.; Fuchs, D.; Pantopoulos, K.; Werner-Felmayer, G.; Wachter, H.; Hentze, M.W. Translational regulation via iron-responsive elements by the nitric oxide/NO-synthase pathway. EMBO J. 1993, 12, 3651–3657. [Google Scholar] [CrossRef] [PubMed]
- Richardson, D.R.; Neumannova, V.; Nagy, E.; Ponka, P. The effect of redox-related species of nitrogen monoxide on transferrin and iron uptake and cellular proliferation of erythroleukemia (K562) cells. In Blood; 1995; 86, pp. 3211–3219. Available online: https://www.ncbi.nlm.nih.gov/pubmed/7579417 (accessed on 16 March 2023).
- Cairo, G.; Ronchi, R.; Recalcati, S.; Campanella, A.; Minotti, G. Nitric oxide and peroxynitrite activate the iron regulatory protein-1 of J774A.1 macrophages by direct disassembly of the Fe-S cluster of cytoplasmic aconitase. Biochemistry 2002, 41, 7435–7442. [Google Scholar] [CrossRef] [PubMed]
- Wardrop, S.L.; Watts, R.N.; Richardson, D.R. Nitrogen monoxide activates iron regulatory protein 1 RNA-binding activity by two possible mechanisms: Effect on the [4Fe-4S] cluster and iron mobilization from cells. Biochemistry 2000, 39, 2748–2758. [Google Scholar] [CrossRef]
- Pantopoulos, K.; Hentze, M.W. Nitric oxide signaling to iron-regulatory protein: Direct control of ferritin mRNA translation and transferrin receptor mRNA stability in transfected fibroblasts. Proc. Natl. Acad. Sci. USA 1995, 92, 1267–1271. [Google Scholar] [CrossRef] [Green Version]
- Mukosera, G.T.; Clark, T.C.; Ngo, L.; Liu, T.; Schroeder, H.; Power, G.G.; Yellon, S.M.; Parast, M.M.; Blood, A.B. Nitric oxide metabolism in the human placenta during aberrant maternal inflammation. J. Physiol. 2020, 598, 2223–2241. [Google Scholar] [CrossRef] [Green Version]
- Mukosera, G.T.; Principe, P.; Mata-Greenwood, E.; Liu, T.; Schroeder, H.; Parast, M.; Blood, A.B. Iron nitrosyl complexes are formed from nitrite in the human placenta. J. Biol. Chem. 2022, 298, 102078. [Google Scholar] [CrossRef]
- Ignarro, L.J.; Fukuto, J.M.; Griscavage, J.M.; Rogers, N.E.; Byrns, R.E. Oxidation of nitric oxide in aqueous solution to nitrite but not nitrate: Comparison with enzymatically formed nitric oxide from L-arginine. Proc. Natl. Acad. Sci. USA 1993, 90, 8103–8107. [Google Scholar] [CrossRef] [Green Version]
- Blood, A.B.; Liu, T.; Mukosera, G.; Hanson, S.F.; Terry, M.H.; Schroeder, H.; Power, G.G. Evidence for placental-derived iron-nitrosyls in the circulation of the fetal lamb and against a role for nitrite in mediating the cardiovascular transition at birth. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2020, 319, R401–R411. [Google Scholar] [CrossRef]
- Hickok, J.R.; Sahni, S.; Shen, H.; Arvind, A.; Antoniou, C.; Fung, L.W.M.; Thomas, D.D. Dinitrosyliron complexes are the most abundant nitric oxide-derived cellular adduct: Biological parameters of assembly and disappearance. Free Radic. Biol. Med. 2011, 51, 1558–1566. [Google Scholar] [CrossRef] [Green Version]
- Mukosera, G.T.; Liu, T.; Ahmed, A.S.I.; Li, Q.; Sheng, M.H.-C.; Tipple, T.E.; Baylink, D.J.; Power, G.G.; Blood, A.B. Detection of dinitrosyl iron complexes by ozone-based chemiluminescence. Nitric Oxide 2018, 79, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Medina, M.V.; Sapochnik, D.; Garcia Solá, M.; Coso, O. Regulation of the Expression of Heme Oxygenase-1: Signal Transduction, Gene Promoter Activation, and Beyond. Antioxid. Redox Signal. 2020, 32, 1033–1044. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Akagi, R.; Mori, M.; Tsuchiya, T.; Sassa, S. Marked developmental changes in heme oxygenase-1 (HO-1) expression in the mouse placenta: Correlation between HO-1 expression and placental development. Placenta 2004, 25, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Inoue, R.; Irie, Y.; Akagi, R. Role of heme oxygenase-1 in human placenta on iron supply to fetus. Placenta 2021, 103, 53–58. [Google Scholar] [CrossRef]
- Hartsfield, C.L.; Alam, J.; Cook, J.L.; Choi, A.M. Regulation of heme oxygenase-1 gene expression in vascular smooth muscle cells by nitric oxide. Am. J. Physiol. 1997, 273, L980–L988. [Google Scholar] [CrossRef]
- Liu, X.-M.; Peyton, K.; Ensenat, D.; Wang, H.; Hannink, M.; Alam, J.; Durante, W. Nitric oxide stimulates heme oxygenase-1 gene transcription via the Nrf2/ARE complex to promote vascular smooth muscle cell survival. Cardiovasc. Res. 2007, 75, 381–389. [Google Scholar] [CrossRef] [Green Version]
- Bouton, C.; Demple, B. Nitric Oxide-inducible Expression of Heme Oxygenase-1 in Human Cells: Translation-Independent Stabilization of the Mrna and Evidence for Direct Action of Nitric Oxide. J. Biol. Chem. 2000, 275, 32688–32693. [Google Scholar] [CrossRef] [Green Version]
- McCaig, D.; Lyall, F. Heme oxygenase expression in human placental villous tissue in response to exposure to in vitro ischemia-reperfusion injury. Hypertens. Pregnancy 2009, 28, 256–272. [Google Scholar] [CrossRef]
- Bilban, M.; Haslinger, P.; Prast, J.; Klinglmüller, F.; Woelfel, T.; Haider, S.; Sachs, A.; Otterbein, L.E.; Desoye, G.; Hiden, U.; et al. Identification of novel trophoblast invasion-related genes: Heme oxygenase-1 controls motility via peroxisome proliferator-activated receptor gamma. Endocrinology 2009, 150, 1000–1013. [Google Scholar] [CrossRef] [Green Version]
- Miya, M.; Okamoto, A.; Nikaido, T.; Tachimoto-Kawaguchi, R.; Tanaka, T. Immunohistochemical localization of heme oxygenase-1 and bilirubin/biopyrrin of heme metabolites as antioxidants in human placenta with preeclampsia. Hypertens. Pregnancy 2020, 39, 33–42. [Google Scholar] [CrossRef]
- Li, J.; Zhou, J.; Ye, Y.; Liu, Q.; Wang, X.; Zhang, N.; Wang, X. Increased Heme Oxygenase-1 and Nuclear Factor Erythroid 2-Related Factor-2 in the Placenta Have a Cooperative Action on Preeclampsia. Gynecol. Obstet. Investig. 2016, 81, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganz, T.; Kaplan, J. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science 2004, 306, 2090–2093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Best, C.M.; Pressman, E.K.; Cao, C.; Cooper, E.; Guillet, R.; Yost, O.L.; Galati, J.; Kent, T.R.; O’Brien, K.O. Maternal iron status during pregnancy compared with neonatal iron status better predicts placental iron transporter expression in humans. FASEB J. 2016, 30, 3541–3550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kämmerer, L.; Mohammad, G.; Wolna, M.; Robbins, P.A.; Lakhal-Littleton, S. Fetal liver hepcidin secures iron stores in utero. Blood 2020, 136, 1549–1557. [Google Scholar] [CrossRef] [PubMed]
- Young, M.F.; Griffin, I.; Pressman, E.; McIntyre, A.W.; Cooper, E.; McNanley, T.; Harris, Z.L.; Westerman, M.; O’Brien, K.O. Maternal hepcidin is associated with placental transfer of iron derived from dietary heme and nonheme sources. J. Nutr. 2012, 142, 33–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, E.A.; Gundogan, F.; Olveda, R.M.; Bartnikas, T.B.; Kurtis, J.D.; Friedman, J.F. Iron transport across the human placenta is regulated by hepcidin. Pediatr. Res. 2020, 92, 396–402. [Google Scholar] [CrossRef]
- Cardaropoli, S.; Todros, T.; Nuzzo, A.M.; Rolfo, A. Maternal serum levels and placental expression of hepcidin in preeclampsia. Pregnancy Hypertens. 2018, 11, 47–53. [Google Scholar] [CrossRef]
- Silvestri, L.; Nai, A.; Dulja, A.; Pagani, A. Hepcidin and the BMP-SMAD pathway: An unexpected liaison. Vitam. Horm. 2019, 110, 71–99. [Google Scholar] [CrossRef]
- Differ, C.; Klatte-Schulz, F.; Bormann, N.; Minkwitz, S.; Knaus, P.; Wildemann, B. Is NO the Answer? The Nitric Oxide Pathway Can Support Bone Morphogenetic Protein 2 Mediated Signaling. Cells 2019, 1979, 8. [Google Scholar] [CrossRef] [Green Version]
- Wada, H.G.; Hass, P.E.; Sussman, H.H. Transferrin receptor in human placental brush border membranes. Studies on the binding of transferrin to placental membrane vesicles and the identification of a placental brush border glycoprotein with high affinity for transferrin. J. Biol. Chem. 1979, 254, 12629–12635. [Google Scholar] [CrossRef]
- Lakhal-Littleton, S. Advances in understanding the crosstalk between mother and fetus on iron utilization. Semin. Hematol. 2021, 58, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Gunshin, H.; Fujiwara, Y.; Custodio, A.O.; Direnzo, C.; Robine, S.; Andrews, N.C. Slc11a2 is required for intestinal iron absorption and erythropoiesis but dispensable in placenta and liver. J. Clin. Investig. 2005, 115, 1258–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mok, H.; Mendoza, M.; Prchal, J.T.; Balogh, P.; Schumacher, A. Dysregulation of ferroportin 1 interferes with spleen organogenesis in polycythaemia mice. Development 2004, 131, 4871–4881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paradkar, P.N.; Roth, J.A. Nitric oxide transcriptionally down-regulates specific isoforms of divalent metal transporter (DMT1) via NF-kappaB. J. Neurochem. 2006, 96, 1768–1777. [Google Scholar] [CrossRef]
- Feelisch, M. The use of nitric oxide donors in pharmacological studies. Naunyn. Schmiedebergs. Arch. Pharmacol. 1998, 358, 113–122. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Principe, P.; Mukosera, G.T.; Gray-Hutto, N.; Tugung, A.; Gheorghe, C.P.; Blood, A.B. Nitric Oxide Affects Heme Oxygenase-1, Hepcidin, and Transferrin Receptor Expression in the Placenta. Int. J. Mol. Sci. 2023, 24, 5887. https://doi.org/10.3390/ijms24065887
Principe P, Mukosera GT, Gray-Hutto N, Tugung A, Gheorghe CP, Blood AB. Nitric Oxide Affects Heme Oxygenase-1, Hepcidin, and Transferrin Receptor Expression in the Placenta. International Journal of Molecular Sciences. 2023; 24(6):5887. https://doi.org/10.3390/ijms24065887
Chicago/Turabian StylePrincipe, Patricia, George T. Mukosera, Nikia Gray-Hutto, Ashra Tugung, Ciprian P. Gheorghe, and Arlin B. Blood. 2023. "Nitric Oxide Affects Heme Oxygenase-1, Hepcidin, and Transferrin Receptor Expression in the Placenta" International Journal of Molecular Sciences 24, no. 6: 5887. https://doi.org/10.3390/ijms24065887