
Transcriptome Reveals the Molecular Mechanism of the ScALDH21 Gene from the Desert Moss Syntrichia caninervis Conferring Resistance to Salt Stress in Cotton

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
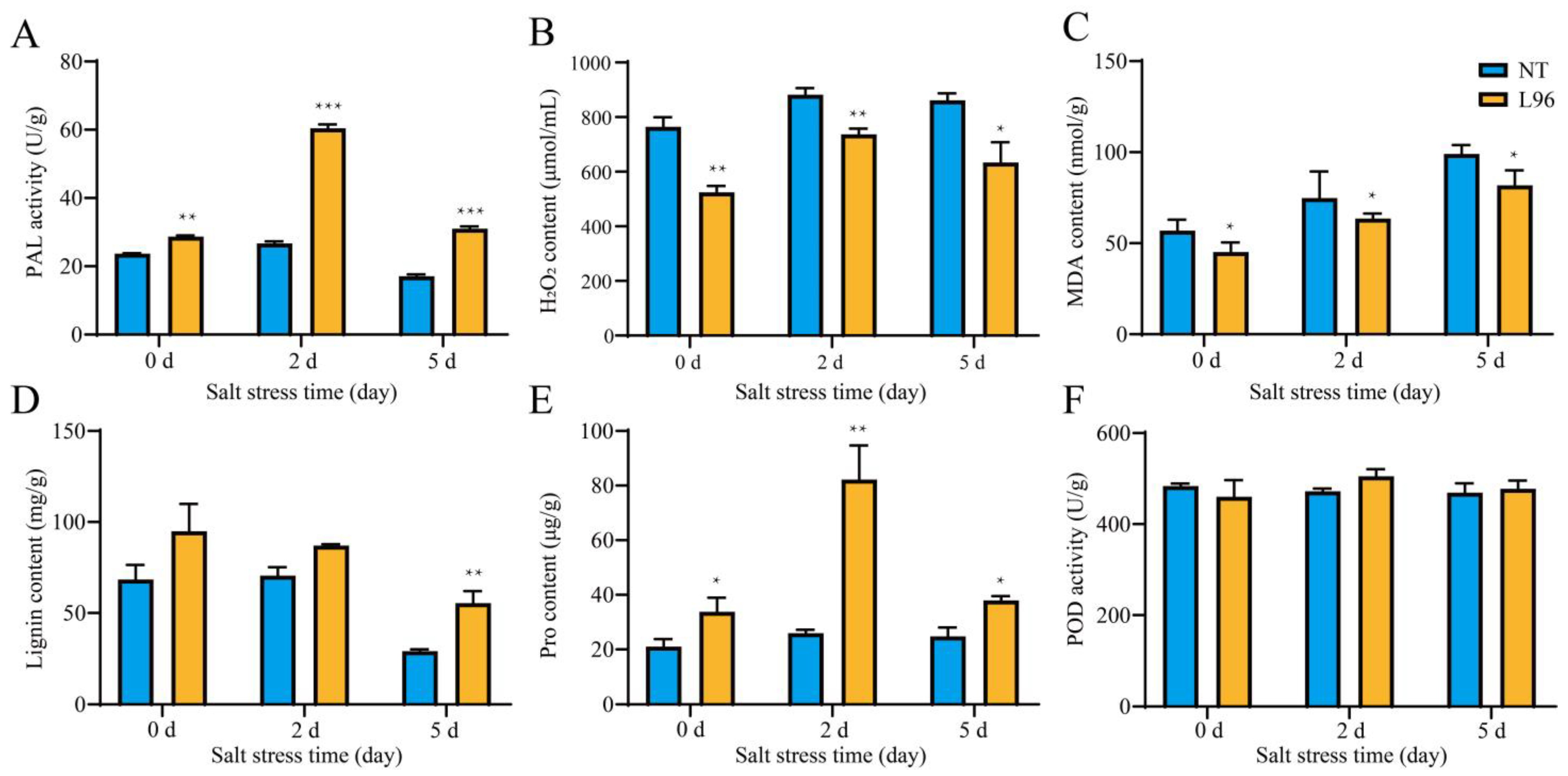
2.1. The Physiological Responses of ScALDH21 Cotton to Salt Stress
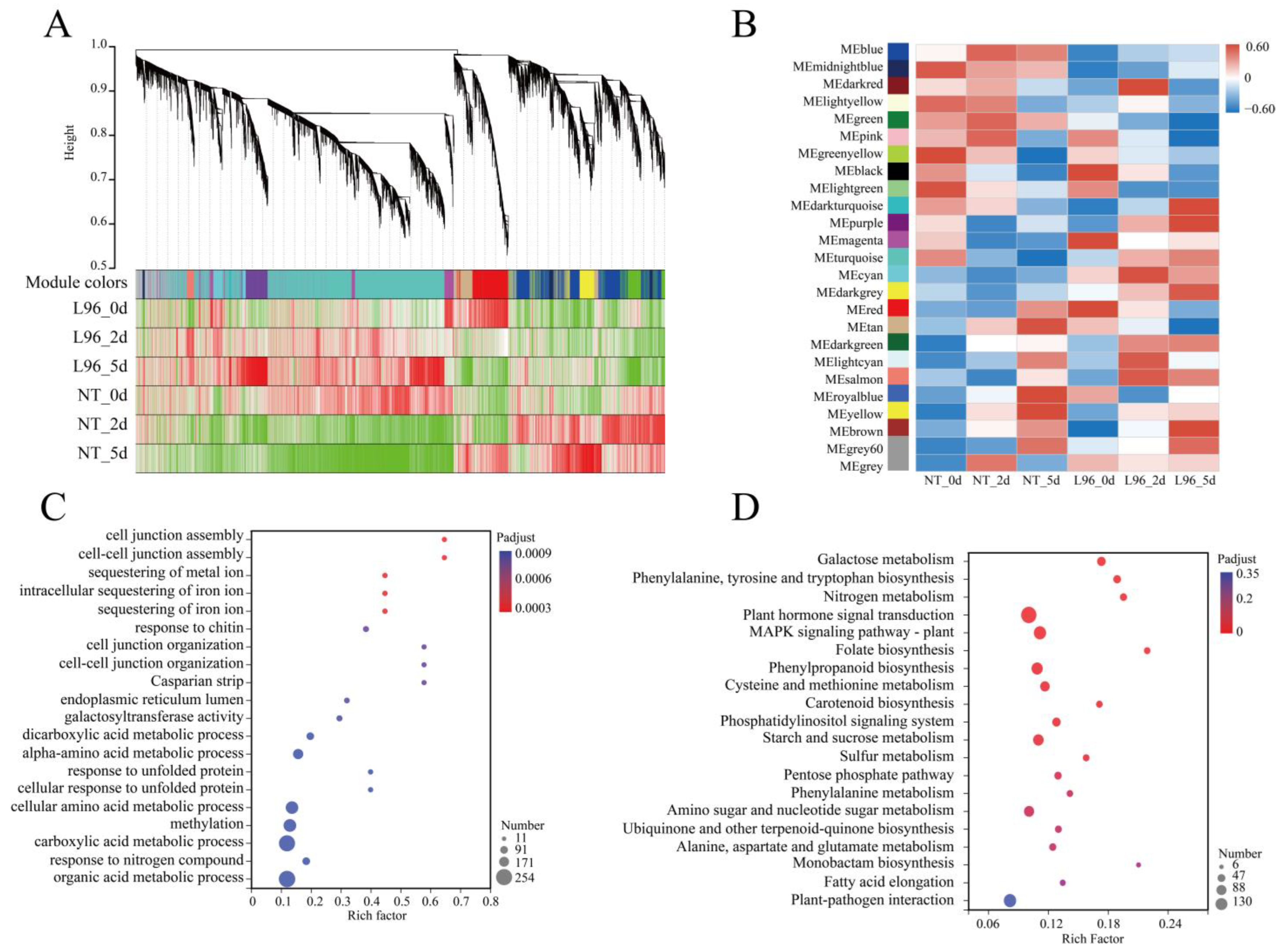
2.2. Analysis of Differentially Expressed Genes during Salt Stress
2.3. Comparison of Differentially Expressed Genes under Salt Stress Conditions
2.4. Weighted Correlation Network Analysis Mines Key Pathways in Transgenic Cotton
2.5. Plant Hormone Signal Transduction Pathway Was More Activated
2.6. Plant–Pathogen Interaction and MAPK Signaling Pathway under Salt Stress
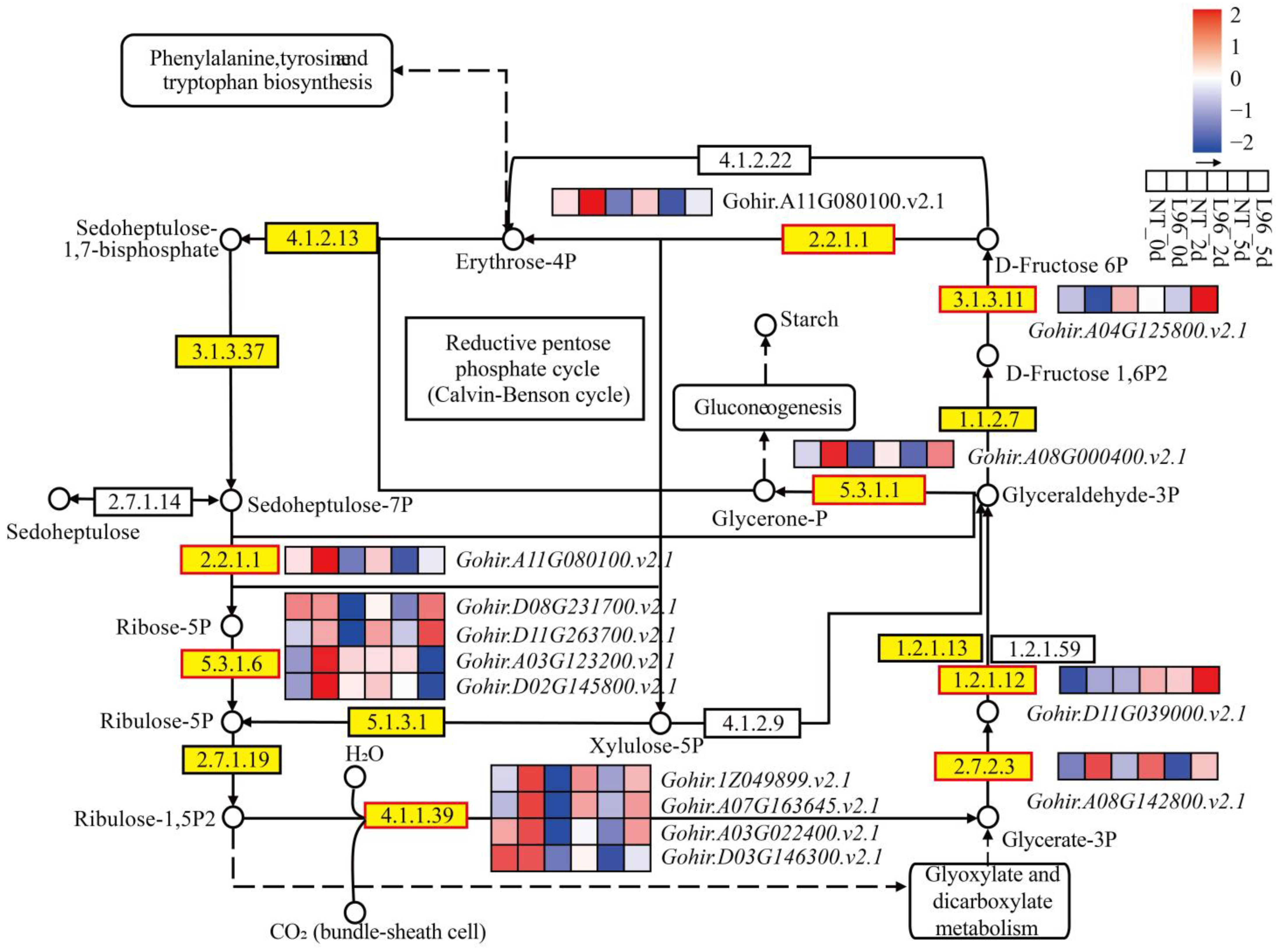
2.7. Carbon Fixation in Photosynthetic Organisms Is Benefit of Transgenic Line
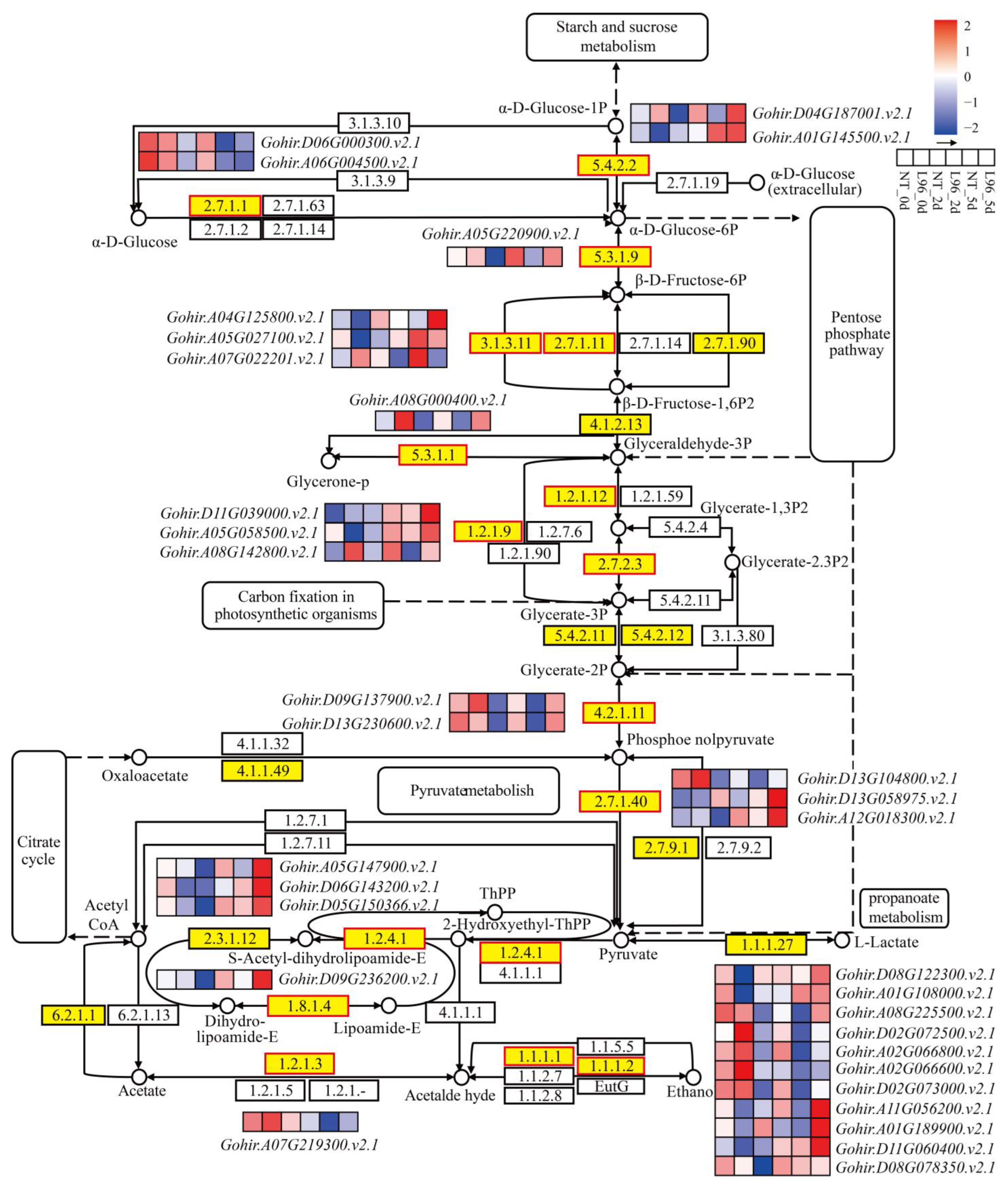
2.8. Carbohydrate Metabolism Is Better in Transgenic Line
3. Discussion
3.1. Effect of Salt Stress on Transcript Levels in Cotton
3.2. The Effects of Plant Hormones on the Response of Transgenic Cotton to Salt Stress
3.3. The Effects of Ca2+ Signaling and the MAPK Pathway on the Response of Transgenic Cotton to Salt Stress
3.4. The Effects of Photosynthesis on the Response of Transgenic Cotton to Salt Stress
3.5. The Effects of Carbohydrate Metabolism on the Response of Transgenic Cotton to Salt Stress
4. Materials and Methods
4.1. Plant Material and Cultured Conditions in Greenhouse
4.2. Measurement of Physiological Indicators
4.3. RNA Extraction and Quantitative PCR
4.4. cDNA Library Preparation and Sequencing
4.5. Quality Control and Read Mapping
4.6. Differential Expression Analysis and Functional Enrichment
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cramer, G.R.; Urano, K.; Delrot, S.; Pezzotti, M.; Shinozaki, K. Effects of abiotic stress on plants: A systems biology perspective. BMC Plant Biol. 2011, 11, 1471–2229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.M.; Zhu, J.H.; Gong, Z.Z.; Zhu, J.K. Abiotic stress responses in plants. Nat. Rev. Genet. 2022, 23, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Zorb, C.; Geilfus, C.M.; Dietz, K.J. Salinity and crop yield. Plant Biol. 2019, 21, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Safdar, H.; Amin, A.; Shafiq, Y.; Ali, A.; Yasin, R.; Shoukat, A.; Hussan, M.U.; Hussan, M.U. A review: Impact of salinity on plant growth. Nat. Sci. 2019, 17, 34–40. [Google Scholar]
- Rezvi, H.U.A.; Arif, M.T.U.; Azim, M.A.; Tumpa, T.A.; Tipu, M.M.H.; Najnine, F.; Dawood, M.F.A.; Skalicky, M.; Brestič, M. Rice and food security: Climate change implications and the future prospects for nutritional security. Food Energy Secur. 2022, 12, e430. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Parvin, K.; Bardhan, K.; Nahar, K.; Anee, T.I.; Masud, A.A.C.; Fotopoulos, V. Biostimulants for the regulation of reactive oxygen species metabolism in plants under abiotic stress. Cells 2021, 10, 2537. [Google Scholar] [CrossRef]
- Dmitrieva, V.A.; Tyutereva, E.V.; Voitsekhovskaja, O.V. Singlet oxygen in plants: Generation, detection, and signaling roles. Int. J. Mol. Sci. 2020, 21, 3237. [Google Scholar] [CrossRef]
- Pitzschke, A.; Forzani, C.; Hirt, H. Reactive oxygen species signaling in plants. Antioxid. Redox Signaling 2006, 8, 1757–1764. [Google Scholar] [CrossRef]
- Gapper, C.; Dolan, L. Control of plant development by reactive oxygen species. Plant Physiol. 2006, 141, 341–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waszczak, C.; Carmody, M.; Kangasjarvi, J. Reactive oxygen species in plant signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotchoni, S.O.; Kuhns, C.; Ditzer, A.; Kirch, H.H.; Bartels, D. Over-expression of different aldehyde dehydrogenase genes in Arabidopsis thaliana confers tolerance to abiotic stress and protects plants against lipid peroxidation and oxidative stress. Plant Cell Environ. 2006, 29, 1033–1048. [Google Scholar] [CrossRef]
- Chen, X.B.; Zeng, Q.; Wood, A.J. Aldh7B6 encodes a turgor-responsive aldehyde dehydrogenase homologue that is constitutively expressed in tortula ruralis gametophytes. Bryologist 2002, 105, 117–184. [Google Scholar] [CrossRef]
- Kirch, H.H.; Bartels, D.; Wei, Y.; Schnable, P.S.; Wood, A.J. The ALDH gene superfamily of Arabidopsis. Trends Plant Sci. 2004, 9, 371–377. [Google Scholar] [CrossRef] [Green Version]
- Tola, A.J.; Jaballi, A.; Germain, H.; Missihoun, T.D. Recent development on plant aldehyde dehydrogenase enzymes and their functions in plant development and stress signaling. Genes 2020, 12, 51. [Google Scholar] [CrossRef]
- Stiti, N.; Giarola, V.; Bartels, D. From algae to vascular plants: The multistep evolutionary trajectory of the ALDH superfamily towards functional promiscuity and the emergence of structural characteristics. Environ. Exp. Bot. 2021, 185, 104376. [Google Scholar] [CrossRef]
- Yang, Z.R.; Qanmbe, G.; Wang, Z.; Yang, Z.E.; Li, F.G. Gossypium Genomics: Trends, Scope, and Utilization for Cotton Improvement. Trends Plant Sci. 2020, 25, 488–500. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.L.; Dong, H.Z. Intensive cotton farming technologies in China: Achievements, challenges and countermeasures. Field Crop. Res. 2014, 155, 99–110. [Google Scholar] [CrossRef] [Green Version]
- Sharif, I.; Aleem, S.; Farooq, J.; Rizwan, M.; Younas, A.; Sarwar, G.; Chohan, S.M. Salinity stress in cotton: Effects, mechanism of tolerance and its management strategies. Physiol. Mol. Biol. Plants 2019, 25, 807–820. [Google Scholar] [CrossRef]
- Zafar, M.M.; Shakeel, A.; Haroon, M.; Manan, A.; Sahar, A.; Shoukat, A.; Mo, H.J.; Farooq, M.A.; Ren, M.Z. Effects of Salinity Stress on Some Growth, Physiological, and Biochemical Parameters in Cotton (Gossypium hirsutum L.) Germplasm. J. Nat. Fibers 2021, 19, 8854–8886. [Google Scholar] [CrossRef]
- Zhang, Y. The microstructure and for-mation of biological soil crusts in their early developmental stage. Chin. Sci. Bull. 2005, 50, 117–121. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Wang, H.L.; Wang, X.Q.; Yang, W.K.; Zhang, D.Y. The microstructure of microbiotic crust and its influence on wind erosion for a sandy soil surface in the gurbantunggut desert of northwestern china. Geoderma 2006, 132, 441–449. [Google Scholar] [CrossRef]
- Silva, A.T.; Gao, B.; Fisher, K.M.; Mishler, B.D.; Ekwealor, J.T.B.; Stark, L.R.; Li, X.; Zhang, D.; Bowker, M.A.; Brinda, J.C.; et al. To dry perchance to live: Insights from the genome of the desiccation-tolerant biocrust moss Syntrichia caninervis. The Plant Journal 2021, 105, 1339–1356. [Google Scholar] [CrossRef]
- Liang, Y.; Li, X.; Zhang, D.; Gao, B.; Yang, H.; Wang, Y.; Guan, K.; Wood, A.J. ScDREB8, a novel A-5 type of DREB gene in the desert moss Syntrichia caninervis, confers salt tolerance to Arabidopsis. Plant Physiol. Biochem. 2017, 120, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Zhan, D.; Li, X.; Yang, H.; Wood, A.J. De novo assembly and characterization of the transcriptome in the desiccation-tolerant moss Syntrichia caninervis. BMC Res. Notes 2014, 7, 490. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Liang, Y.; Gao, B.; Mijiti, M.; Bozorov, T.A.; Yang, H.; Zhang, D.; Wood, A.J. ScDREB10, an A-5c type of DREB gene of the desert moss Syntrichia caninervis, confers osmotic and salt tolerances to arabidopsis. Genes 2019, 10, 146. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhou, P.; Li, X.; Zhang, D. Propagation of desert moss Syntrichia caninervis in peat pellet: A method for rapidly obtaining large numbers of cloned gametophytes. Plant Methods 2021, 17, 42. [Google Scholar] [CrossRef]
- Liu, J.; Yang, R.; Liang, Y.; Wang, Y.; Li, X. The DREB A-5 transcription factor ScDREB5 from Syntrichia caninervis enhanced salt tolerance by regulating jasmonic acid biosynthesis in transgenic Arabidopsis. Front. Plant Sci. 2022, 13, 735. [Google Scholar] [CrossRef] [PubMed]
- Li, X.S.; Yang, R.R.; Liang, Y.Q.; Gao, B.; Li, S.M.; Bai, W.W.; Oliver, M.J.; Zhang, D.Y. The ScAPD1-like gene from the desert moss Syntrichia caninervis enhances resistance to Verticillium dahliae via phenylpropanoid gene regulation. Plant J. 2022, 113, 75–91. [Google Scholar] [CrossRef]
- Yang, H.L.; Zhang, D.Y.; Li, X.S.; Li, H.Y.; Zhang, D.W.; Lan, H.Y.; Wood, A.J.; Wang, J.C. Overexpression of ScALDH21 gene in cotton improves drought tolerance and growth in greenhouse and field conditions. Mol. Breed. 2016, 36, 34. [Google Scholar] [CrossRef]
- Yang, H.L.; Zhang, D.Y.; Wang, J.C.; Wood, A.J.; Zhang, Y.M. Molecular cloning of a stress-responsive aldehyde dehydrogenase gene ScALDH21 from the desiccation-tolerant moss Syntrichia caninervis and its responses to different stresses. Mol. Biol. Rep. 2012, 39, 2645–2652. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.L.; Bozorov, T.A.; Chen, X.P.; Zhang, D.W.; Wang, J.C.; Li, X.S.; Gui, D.W.; Qi, Z.M.; Zhang, D.Y. Yield comparisons between cotton variety xin nong mian 1 and its transgenic ScALDH21 lines under different water deficiencies in a desert-oasis ecotone. Agronomy 2021, 11, 1019. [Google Scholar] [CrossRef]
- Yang, H.L.; Zhang, D.Y.; Li, H.Y.; Dong, L.F.; Lan, H.Y. Ectopic overexpression of the aldehyde dehydrogenase ALDH21 from Syntrichia caninervis in tobacco confers salt and drought stress tolerance. Plant Physiol. Biochem. 2015, 95, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.C.; Yang, H.L.; Gao, B.; Bozorov, T.A.; Li, X.S.; Zhang, D.Y.; Wood, A.J. Analysis and characterization of the aldehyde dehydrogenase (ALDH) gene superfamily in the desert moss Syntrichia caninervis in response to abiotic stress. Environ. Exp. Bot. 2020, 178, 104176. [Google Scholar] [CrossRef]
- Yang, H.L.; Zhang, D.W.; Zhang, D.Y.; Bozorov, T.A.; Abdullaev, A.A.; Wood, A.J.; Wang, J.C.; Li, X.S.; Zhao, J.Y. Overexpression of ALDH21 from Syntrichia caninervis Moss in upland cotton enhances fiber quality, boll component traits, and physiological parameters during deficit Irrigation. Crop Sci. 2019, 59, 553–564. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.H.; Yan, H.L.; Zhang, D.W.; Ma, Q.Q.; Zhang, D.Y. Analysis of salt toerance of ScALDH21 transgenic cotton. Acta Agriculurae Boreali-Sinica 2022, 37, 31–36. [Google Scholar]
- Maury, L.L.; Marguerat, S.; Bähler, J. Tuning gene expression to changing environments: From rapid responses to evolutionary adaptation. Nat. Rev. Genet. 2008, 9, 583–593. [Google Scholar] [CrossRef]
- Mazzucotelli, E.; Mastrangelo, A.M.; Crosatti, C.; Guerra, D.; Stanca, A.M.; Cattivelli, L. Abiotic stress response in plants: When post-transcriptional and post-translational regulations control transcription. Plant Sci. 2008, 174, 420–431. [Google Scholar] [CrossRef]
- Jamil, A.; Riaz, S.; Ashraf, M.; Foolad, M.R. Gene Expression Profiling of Plants under Salt Stress. Crit. Rev. Plant Sci. 2011, 30, 435–458. [Google Scholar] [CrossRef]
- Allu, A.D.; Soja, A.M.; Wu, A.; Szymanski, J.; Balazadeh, S. Salt stress and senescence: Identification of cross-talk regulatory components. J. Exp. Bot. 2014, 65, 3993–4008. [Google Scholar] [CrossRef] [Green Version]
- Geng, Y.; Wu, R.; Wee, C.W.; Xie, F.; Wei, X.L.; Chan, P.M.Y.; Tham, C.; Duan, L.; Dinneny, J.R. A spatio-temporal understanding of growth regulation during the salt stress response in Arabidopsis. Plant Cell 2013, 25, 2132–2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devaraj, V.R.; Dsouza, M. Signaling Molecules and Their Involvement in Abiotic and Biotic Stress Response Crosstalk in Plants; CRC Press: Boca Raton, FL, USA, 2018; Volume 26, pp. 295–310. [Google Scholar]
- Sharma, D.; Verma, N.; Pandey, C.; Verma, D.; Bhagat, P.K.; Noryang, S.; Singh, K.; Tayyeba, S.; Banerjee, G.; Sinha, A.K. MAP kinase as regulators for stress responses in plants. In Protein Kinases and Stress Signaling in Plants; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2020; Volume 15, pp. 369–392. [Google Scholar]
- Golldack, D.; King, I.L.; Yang, O. Plant tolerance to drought and salinity: Stress regulating transcription factors and their functional significance in the cellular transcriptional network. Plant Cell Rep. 2011, 30, 1383–1391. [Google Scholar] [CrossRef]
- Shah, W.H.; Rasool, A.; Saleem, S.; Mushtaq, N.U.; Tahir, I.; Hakeem, K.R.; Rehman, R.U. Understanding the integrated pathways and mechanisms of transporters, protein kinases, and transcription factors in plants under salt stress. Int. J. Genomics 2021, 2021, 5578727. [Google Scholar] [CrossRef] [PubMed]
- Ke, Y.G.; Yuan, M.; Liu, H.B.; Hui, S.G.; Qin, X.F.; Chen, J.; Zhang, Q.L.; Li, X.H.; Xiao, J.H.; Zhang, Q.F.; et al. The versatile functions of OsALDH2B1 provide a genic basis for growth-defense trade-offs in rice. Proc. Natl. Acad. Sci. USA 2020, 117, 3867–3873. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Duan, X.; Luo, L.; Da, S.; Ding, Z.; Xia, G. How plant hormones mediate salt stress responses. Trends Plant Sci. 2020, 25, 1117–1130. [Google Scholar] [CrossRef] [PubMed]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef] [Green Version]
- Chandler, J.W. Auxin as compere in plant hormone crosstalk. Planta 2009, 231, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ribba, T.; Vargas, F.G.; Brien, J.A.O. Auxin-mediated responses under salt stress: From developmental regulation to biotechnological applications. J. Exp. Bot. 2020, 71, 3843–3853. [Google Scholar] [CrossRef]
- Ryu, H.; Cho, Y.G. Plant hormones in salt stress tolerance. J. Plant Biol. 2015, 58, 147–155. [Google Scholar] [CrossRef]
- Kanojia, A.; Dijkwel, P.P. Abiotic stress responses are governed by reactive oxygen species and age. Annu. Plant Rev. 2018, 1, 295–326. [Google Scholar]
- Wang, X.X.; Chen, X.D.; Wang, Q.J.; Chen, M.; Liu, X.; Gao, D.S.; Li, D.M.; Li, L. MdBZR1 and MdBZR1-2like transcription factors improves salt tolerance by regulating gibberellin biosynthesis in apple. Front. Plant Sci. 2019, 10, 1473. [Google Scholar] [CrossRef]
- Vafadar, F.; Amooaghaie, R.; Ehsanzadeh, P.; Ghanati, F.; Sajedi, R.H. Crosstalk between melatonin and Ca(2+)/CaM evokes systemic salt tolerance in Dracocephalum kotschyi. J. Plant Physiol. 2020, 252, 153237. [Google Scholar] [CrossRef]
- Wu, X.T.; Liu, Z.Y.; Liao, W.B. The involvement of gaseous signaling molecules in plant MAPK cascades: Function and signal transduction. Planta 2021, 254, 127. [Google Scholar] [CrossRef] [PubMed]
- Sudhir, P.; Murthy, S.D.S. Effects of salt stress on basic processes of photosynthesis. Photosynthetica 2004, 42, 481–486. [Google Scholar] [CrossRef]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Henry, E.; Fung, N.; Liu, J.; Drakakaki, G.; Coaker, G. Beyond glycolysis: GAPDHs are multi-functional enzymes involved in regulation of ROS, autophagy, and plant immune responses. PLoS Genet. 2015, 11, e1005199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wieloch, T. A cytosolic oxidation-reduction cycle in plant leaves. J. Exp. Bot. 2021, 72, 4186–4189. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Yu, G.; Shi, C.P.; Liu, L.M.; Guo, Q.; Han, C.; Zhang, D.; Zhang, L.; Liu, B.; Gao, H.; et al. Majorbio Cloud: A one-stop, comprehensive bioinformatic platform for multiomics analyses. iMeta 2022, 1, e12. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Yang, Q.; Zhang, D.; Wang, J.; Cao, T.; Bozorov, T.A.; Cheng, L.; Zhang, D. Transcriptome Reveals the Molecular Mechanism of the ScALDH21 Gene from the Desert Moss Syntrichia caninervis Conferring Resistance to Salt Stress in Cotton. Int. J. Mol. Sci. 2023, 24, 5822. https://doi.org/10.3390/ijms24065822
Yang H, Yang Q, Zhang D, Wang J, Cao T, Bozorov TA, Cheng L, Zhang D. Transcriptome Reveals the Molecular Mechanism of the ScALDH21 Gene from the Desert Moss Syntrichia caninervis Conferring Resistance to Salt Stress in Cotton. International Journal of Molecular Sciences. 2023; 24(6):5822. https://doi.org/10.3390/ijms24065822
Chicago/Turabian StyleYang, Honglan, Qilin Yang, Dawei Zhang, Jiancheng Wang, Ting Cao, Tohir A. Bozorov, Lihua Cheng, and Daoyuan Zhang. 2023. "Transcriptome Reveals the Molecular Mechanism of the ScALDH21 Gene from the Desert Moss Syntrichia caninervis Conferring Resistance to Salt Stress in Cotton" International Journal of Molecular Sciences 24, no. 6: 5822. https://doi.org/10.3390/ijms24065822