
Differences in Whole-Blood Transcriptional Profiles in Inflammatory Bowel Disease Patients Responding to Vedolizumab Compared with Non-Responders †

Abstract
:1. Introduction
2. Results
2.1. Clinical Characteristics
2.2. Plasma Concentration of VDZ vs. Response to Treatment
2.3. Whole-Genome Expression Analysis
2.4. Deconvolution
2.5. Gene-Expression in Responders Compared with Non-Responders
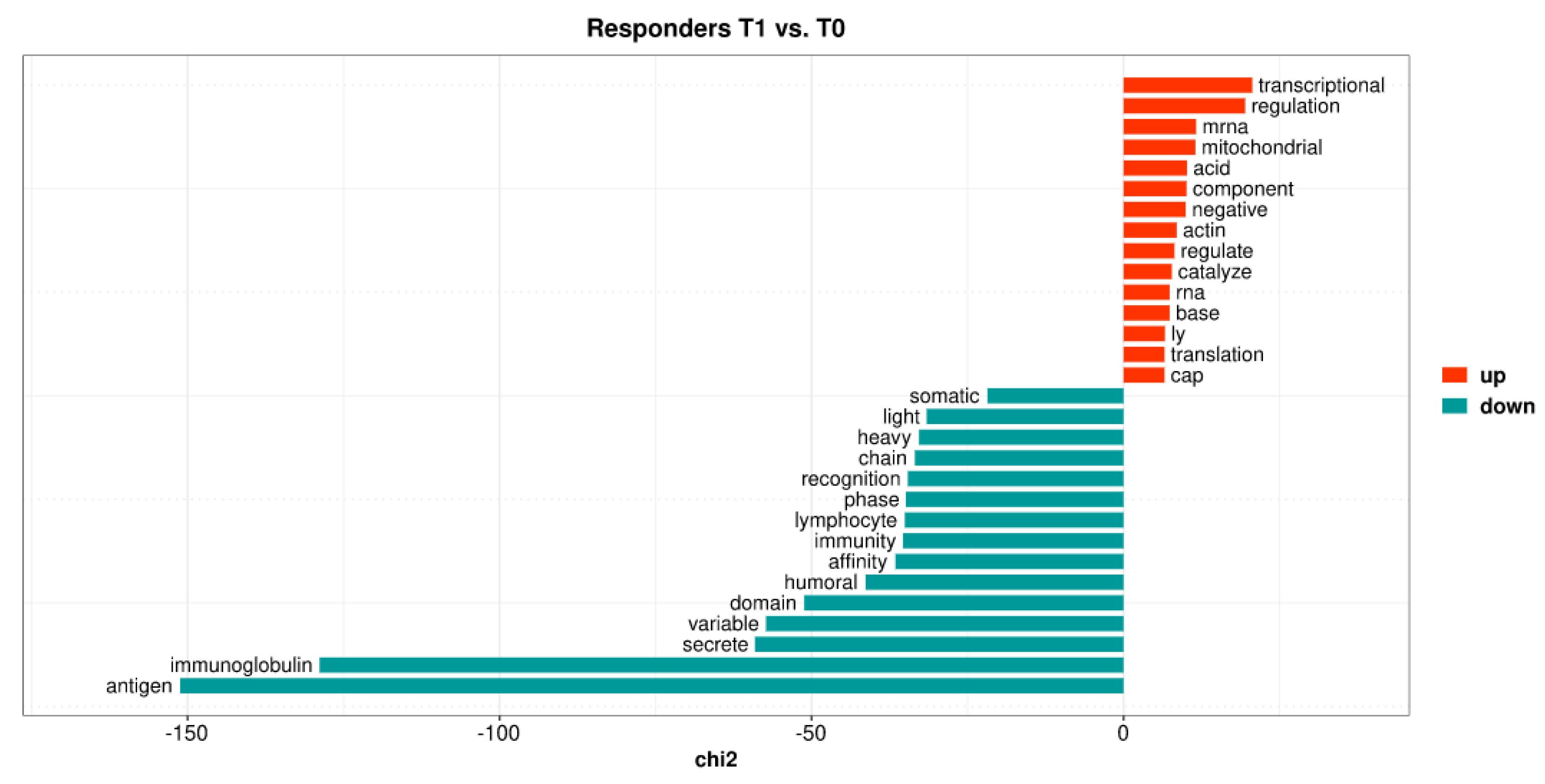
2.6. VDZ Regulates the Expression of Genes in Whole Blood Only in Patients Responding to Treatment
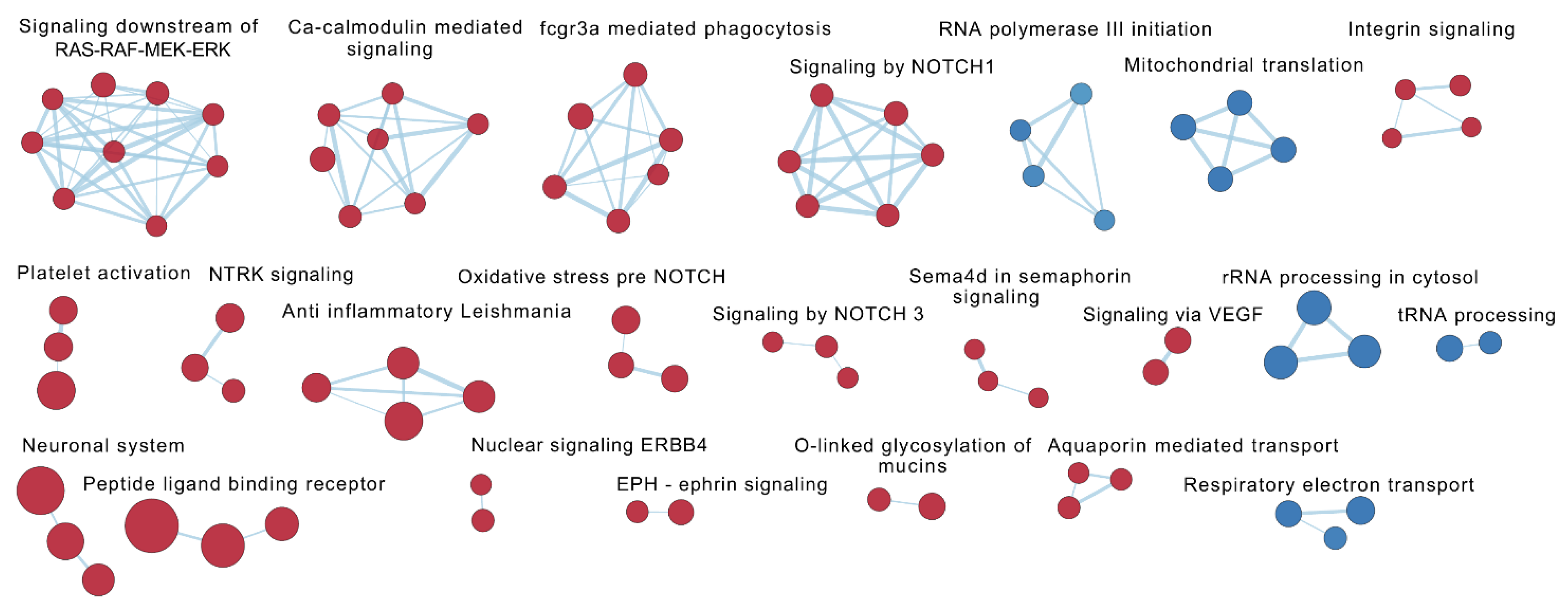
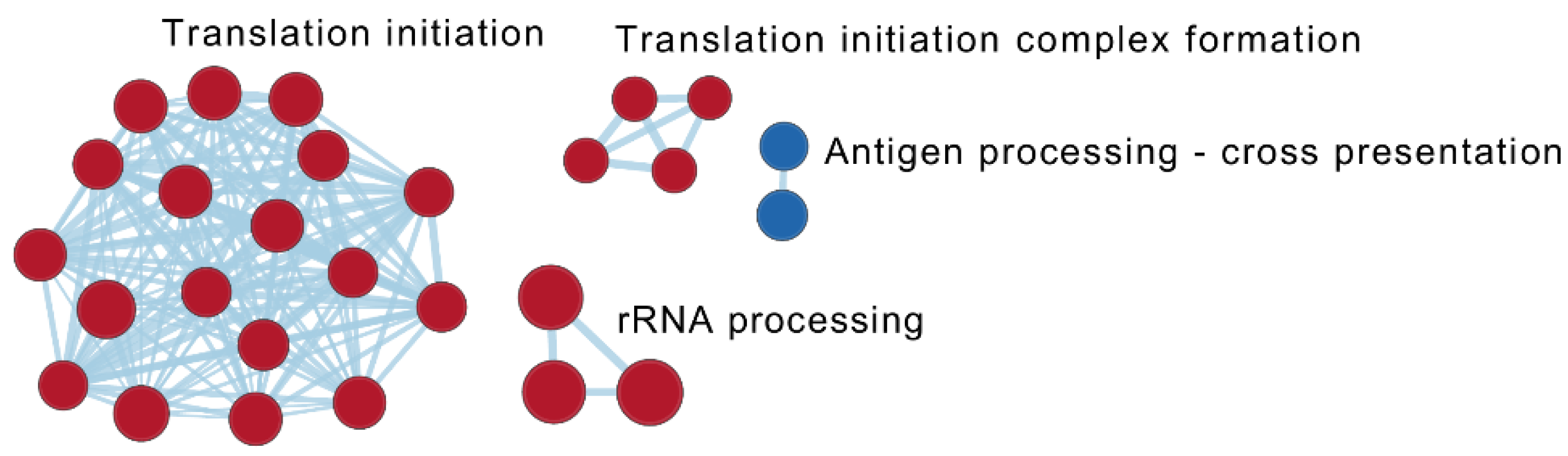
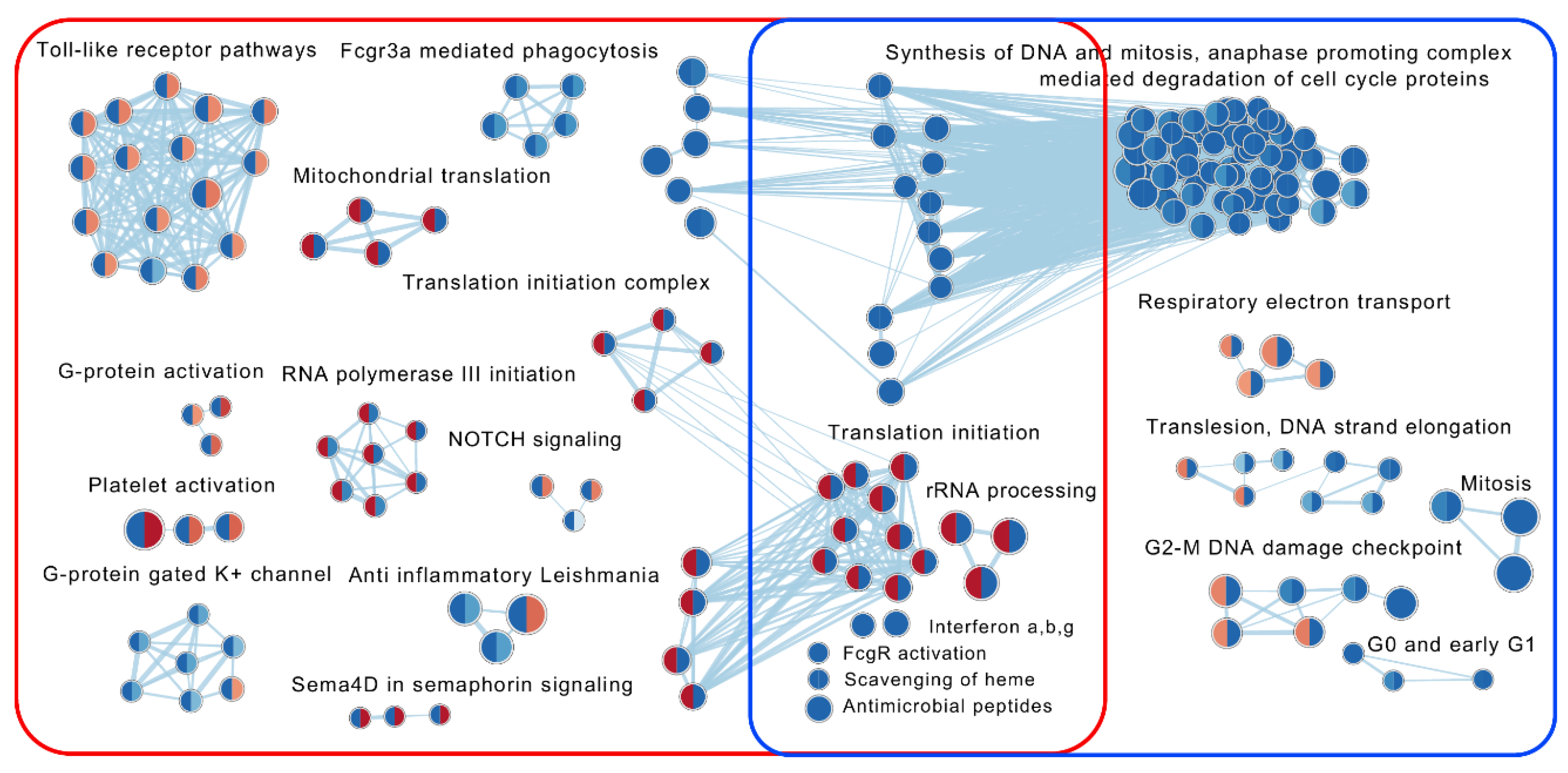
2.7. Gene Set Enrichment Analyses
2.7.1. Enrichment Analysis in Responders Compared with Non-Responders
2.7.2. The Effect of VDZ Treatment on Biological Pathways within Responders and Non-Responders
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Sample Collection
4.3. Plasma Concentration of Vedolizumab
4.4. RNA Sequencing
4.5. Statistical Analyses
4.5.1. Basic Statistics
4.5.2. Differentially Expressed Genes
4.5.3. Pathway Analyses
4.5.4. Deconvolution
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| IBD | inflammatory bowel disease |
| UC | ulcerative colitis |
| CD | Crohn’s disease |
| VDZ | vedolizumab |
| sHBI | simplified Harvey Bradshaw index |
| SCCAI | Simple Clinical Colitis Activity Index |
| PGA | physician global assessment |
| DEGs | differentially expressed genes |
| FC | fold-change |
| FDR | false discovery rate |
| ORA | over-representation analysis |
| GSEA | gene set enrichment analysis |
| NES | normalized enrichment score |
| CRP | C-reactive protein |
| Hb | haemoglobin |
| Alb | albumin |
References
- Colombel, J.F.; Sandborn, W.J.; Reinisch, W.; Mantzaris, G.J.; Kornbluth, A.; Rachmilewitz, D.; Lichtiger, S.; D’Haens, G.; Diamond, R.H.; Broussard, D.L.; et al. Infliximab, azathioprine, or combination therapy for Crohn’s disease. N. Engl. J. Med. 2010, 362, 1383–1395. [Google Scholar] [CrossRef] [Green Version]
- Duijvestein, M.; Battat, R.; Vande Casteele, N.; D’Haens, G.R.; Sandborn, W.J.; Khanna, R.; Jairath, V.; Feagan, B.G. Novel Therapies and Treatment Strategies for Patients with Inflammatory Bowel Disease. Curr. Treat. Options Gastroenterol. 2018, 16, 129–146. [Google Scholar] [CrossRef] [PubMed]
- Gisbert, J.P.; Chaparro, M. Predictors of Primary Response to Biologic Treatment [Anti-TNF, Vedolizumab, and Ustekinumab] in Patients with Inflammatory Bowel Disease: From Basic Science to Clinical Practice. J. Crohns Colitis 2020, 14, 694–709. [Google Scholar] [CrossRef]
- Elhag, D.A.; Kumar, M.; Saadaoui, M.; Akobeng, A.K.; Al-Mudahka, F.; Elawad, M.; Al Khodor, S. Inflammatory Bowel Disease Treatments and Predictive Biomarkers of Therapeutic Response. Int. J. Mol. Sci. 2022, 23, 6966. [Google Scholar] [CrossRef] [PubMed]
- Habtezion, A.; Nguyen, L.P.; Hadeiba, H.; Butcher, E.C. Leukocyte Trafficking to the Small Intestine and Colon. Gastroenterology 2016, 150, 340–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feagan, B.G.; Rutgeerts, P.; Sands, B.E.; Hanauer, S.; Colombel, J.F.; Sandborn, W.J.; Van Assche, G.; Axler, J.; Kim, H.J.; Danese, S.; et al. Vedolizumab as induction and maintenance therapy for ulcerative colitis. N. Engl. J. Med. 2013, 369, 699–710. [Google Scholar] [CrossRef] [Green Version]
- Sandborn, W.J.; Feagan, B.G.; Rutgeerts, P.; Hanauer, S.; Colombel, J.F.; Sands, B.E.; Lukas, M.; Fedorak, R.N.; Lee, S.; Bressler, B.; et al. Vedolizumab as induction and maintenance therapy for Crohn’s disease. N. Engl. J. Med. 2013, 369, 711–721. [Google Scholar] [CrossRef] [Green Version]
- Sands, B.E.; Sandborn, W.J.; Van Assche, G.; Lukas, M.; Xu, J.; James, A.; Abhyankar, B.; Lasch, K. Vedolizumab as Induction and Maintenance Therapy for Crohn’s Disease in Patients Naive to or Who Have Failed Tumor Necrosis Factor Antagonist Therapy. Inflamm. Bowel Dis. 2017, 23, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Gisbert, J.P.; Chaparro, M. Primary Failure to an Anti-TNF Agent in Inflammatory Bowel Disease: Switch (to a Second Anti-TNF Agent) or Swap (for Another Mechanism of Action)? J. Clin. Med. 2021, 10, 5318. [Google Scholar] [CrossRef] [PubMed]
- Arijs, I.; De Hertogh, G.; Lemmens, B.; Van Lommel, L.; de Bruyn, M.; Vanhove, W.; Cleynen, I.; Machiels, K.; Ferrante, M.; Schuit, F.; et al. Effect of vedolizumab (anti-alpha4beta7-integrin) therapy on histological healing and mucosal gene expression in patients with UC. Gut 2018, 67, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Zeissig, S.; Rosati, E.; Dowds, C.M.; Aden, K.; Bethge, J.; Schulte, B.; Pan, W.H.; Mishra, N.; Zuhayra, M.; Marx, M.; et al. Vedolizumab is associated with changes in innate rather than adaptive immunity in patients with inflammatory bowel disease. Gut 2019, 68, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Rath, T.; Billmeier, U.; Ferrazzi, F.; Vieth, M.; Ekici, A.; Neurath, M.F.; Atreya, R. Effects of Anti-Integrin Treatment with Vedolizumab on Immune Pathways and Cytokines in Inflammatory Bowel Diseases. Front. Immunol. 2018, 9, 1700. [Google Scholar] [CrossRef] [Green Version]
- Boden, E.K.; Shows, D.M.; Chiorean, M.V.; Lord, J.D. Identification of Candidate Biomarkers Associated with Response to Vedolizumab in Inflammatory Bowel Disease. Dig. Dis. Sci. 2018, 63, 2419–2429. [Google Scholar] [CrossRef]
- Ungar, B.; Kopylov, U.; Yavzori, M.; Fudim, E.; Picard, O.; Lahat, A.; Coscas, D.; Waterman, M.; Haj-Natour, O.; Orbach-Zingboim, N.; et al. Association of Vedolizumab Level, Anti-Drug Antibodies, and alpha4beta7 Occupancy with Response in Patients With Inflammatory Bowel Diseases. Clin. Gastroenterol. Hepatol. 2018, 16, 697–705.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, I.; Allner, C.; Muhl, L.; Melde, M.; Lissner, D.; Mantzivi, E.; Glauben, R.; Vitali, F.; Becker, E.; Atreya, I.; et al. Expression and function of alpha4beta7 integrin predict the success of vedolizumab treatment in inflammatory bowel disease. Transl. Res. 2022, 253, 8–15. [Google Scholar] [CrossRef]
- Zwicker, S.; Lira-Junior, R.; Hoog, C.; Almer, S.; Bostrom, E.A. Systemic Chemokine Levels with “Gut-Specific” Vedolizumab in Patients with Inflammatory Bowel Disease-A Pilot Study. Int. J. Mol. Sci. 2017, 18, 1827. [Google Scholar] [CrossRef] [Green Version]
- Soendergaard, C.; Seidelin, J.B.; Steenholdt, C.; Nielsen, O.H. Putative biomarkers of vedolizumab resistance and underlying inflammatory pathways involved in IBD. BMJ Open Gastroenterol. 2018, 5, e000208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juzenas, S.; Hubenthal, M.; Lindqvist, C.M.; Kruse, R.; Steiert, T.A.; Degenhardt, F.; Schulte, D.; Nikolaus, S.; Zeissig, S.; Bergemalm, D.; et al. Detailed Transcriptional Landscape of Peripheral Blood Points to Increased Neutrophil Activation in Treatment-Naive Inflammatory Bowel Disease. J. Crohns Colitis 2022, 16, 1097–1109. [Google Scholar] [CrossRef] [PubMed]
- Burakoff, R.; Chao, S.; Perencevich, M.; Ying, J.; Friedman, S.; Makrauer, F.; Odze, R.; Khurana, H.; Liew, C.C. Blood-based biomarkers can differentiate ulcerative colitis from Crohn’s disease and noninflammatory diarrhea. Inflamm. Bowel Dis. 2011, 17, 1719–1725. [Google Scholar] [CrossRef]
- Shi, W.; Ye, Z.; Zhuang, L.; Li, Y.; Shuai, W.; Zuo, Z.; Mao, X.; Liu, R.; Wu, J.; Chen, S.; et al. Olfactomedin 1 negatively regulates NF-kappaB signalling and suppresses the growth and metastasis of colorectal cancer cells. J. Pathol. 2016, 240, 352–365. [Google Scholar] [CrossRef] [Green Version]
- Lovisa, S.; Genovese, G.; Danese, S. Role of Epithelial-to-Mesenchymal Transition in Inflammatory Bowel Disease. J. Crohns Colitis 2019, 13, 659–668. [Google Scholar] [CrossRef]
- Wang, C.Y.; Babitt, J.L. Hepcidin regulation in the anemia of inflammation. Curr. Opin. Hematol. 2016, 23, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Zhang, Y.; Qian, L.; Wang, P. Emerging strategies to target RAS signaling in human cancer therapy. J. Hematol. Oncol. 2021, 14, 116. [Google Scholar] [CrossRef]
- Shi, Y.; Suh, Y.H.; Milstein, A.D.; Isozaki, K.; Schmid, S.M.; Roche, K.W.; Nicoll, R.A. Functional comparison of the effects of TARPs and cornichons on AMPA receptor trafficking and gating. Proc. Natl. Acad. Sci. USA 2010, 107, 16315–16319. [Google Scholar] [CrossRef] [Green Version]
- Bunker, R.D.; Bulloch, E.M.; Dickson, J.M.; Loomes, K.M.; Baker, E.N. Structure and function of human xylulokinase, an enzyme with important roles in carbohydrate metabolism. J. Biol. Chem. 2013, 288, 1643–1652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verstockt, B.; Verstockt, S.; Veny, M.; Dehairs, J.; Arnauts, K.; Van Assche, G.; De Hertogh, G.; Vermeire, S.; Salas, A.; Ferrante, M. Expression Levels of 4 Genes in Colon Tissue Might Be Used to Predict Which Patients Will Enter Endoscopic Remission After Vedolizumab Therapy for Inflammatory Bowel Diseases. Clin. Gastroenterol. Hepatol. 2020, 18, 1142–1151.e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feagan, B.G.; Greenberg, G.R.; Wild, G.; Fedorak, R.N.; Pare, P.; McDonald, J.W.; Cohen, A.; Bitton, A.; Baker, J.; Dube, R.; et al. Treatment of active Crohn’s disease with MLN0002, a humanized antibody to the alpha4beta7 integrin. Clin. Gastroenterol. Hepatol. 2008, 6, 1370–1377. [Google Scholar] [CrossRef]
- Feagan, B.G.; Greenberg, G.R.; Wild, G.; Fedorak, R.N.; Pare, P.; McDonald, J.W.; Dube, R.; Cohen, A.; Steinhart, A.H.; Landau, S.; et al. Treatment of ulcerative colitis with a humanized antibody to the alpha4beta7 integrin. N. Engl. J. Med. 2005, 352, 2499–2507. [Google Scholar] [CrossRef]
- Xiong, E.; Li, Y.; Min, Q.; Cui, C.; Liu, J.; Hong, R.; Lai, N.; Wang, Y.; Sun, J.; Matsumoto, R.; et al. MZB1 promotes the secretion of J-chain-containing dimeric IgA and is critical for the suppression of gut inflammation. Proc. Natl. Acad. Sci. USA 2019, 116, 13480–13489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soler, D.; Chapman, T.; Yang, L.L.; Wyant, T.; Egan, R.; Fedyk, E.R. The binding specificity and selective antagonism of vedolizumab, an anti-alpha4beta7 integrin therapeutic antibody in development for inflammatory bowel diseases. J. Pharmacol. Exp. Ther. 2009, 330, 864–875. [Google Scholar] [CrossRef] [Green Version]
- Postigo, A.A.; Sanchez-Mateos, P.; Lazarovits, A.I.; Sanchez-Madrid, F.; de Landazuri, M.O. Alpha 4 beta 7 integrin mediates B cell binding to fibronectin and vascular cell adhesion molecule-1. Expression and function of alpha 4 integrins on human B lymphocytes. J. Immunol. 1993, 151, 2471–2483. [Google Scholar] [CrossRef] [PubMed]
- Uzzan, M.; Tokuyama, M.; Rosenstein, A.K.; Tomescu, C.; SahBandar, I.N.; Ko, H.M.; Leyre, L.; Chokola, A.; Kaplan-Lewis, E.; Rodriguez, G.; et al. Anti-alpha4beta7 therapy targets lymphoid aggregates in the gastrointestinal tract of HIV-1-infected individuals. Sci. Transl. Med. 2018, 10, eaau4711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyler, C.J.; Guzman, M.; Lundborg, L.R.; Yeasmin, S.; Zgajnar, N.; Jedlicka, P.; Bamias, G.; Rivera-Nieves, J. Antibody secreting cells are critically dependent on integrin alpha4beta7/MAdCAM-1 for intestinal recruitment and control of the microbiota during chronic colitis. Mucosal. Immunol. 2022, 15, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Abraham, C.; Abreu, M.T.; Turner, J.R. Pattern Recognition Receptor Signaling and Cytokine Networks in Microbial Defenses and Regulation of Intestinal Barriers: Implications for Inflammatory Bowel Disease. Gastroenterology 2022, 162, 1602–1616.e6. [Google Scholar] [CrossRef]
- Alzoghaibi, M.A. Concepts of oxidative stress and antioxidant defense in Crohn’s disease. World J. Gastroenterol. 2013, 19, 6540–6547. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.; Gracio, D.; Teixeira, J.P.; Magro, F. Oxidative Stress and DNA Damage: Implications in Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2015, 21, 2403–2417. [Google Scholar] [CrossRef]
- Schippers, A.; Muschaweck, M.; Clahsen, T.; Tautorat, S.; Grieb, L.; Tenbrock, K.; Gassler, N.; Wagner, N. beta7-Integrin exacerbates experimental DSS-induced colitis in mice by directing inflammatory monocytes into the colon. Mucosal Immunol. 2016, 9, 527–538. [Google Scholar] [CrossRef] [Green Version]
- Schleier, L.; Wiendl, M.; Heidbreder, K.; Binder, M.T.; Atreya, R.; Rath, T.; Becker, E.; Schulz-Kuhnt, A.; Stahl, A.; Schulze, L.L.; et al. Non-classical monocyte homing to the gut via alpha4beta7 integrin mediates macrophage-dependent intestinal wound healing. Gut 2020, 69, 252–263. [Google Scholar] [CrossRef]
- Keir, M.E.; Fuh, F.; Ichikawa, R.; Acres, M.; Hackney, J.A.; Hulme, G.; Carey, C.D.; Palmer, J.; Jones, C.J.; Long, A.K.; et al. Regulation and Role of alphaE Integrin and Gut Homing Integrins in Migration and Retention of Intestinal Lymphocytes during Inflammatory Bowel Disease. J. Immunol. 2021, 207, 2245–2254. [Google Scholar] [CrossRef]
- Dai, B.; Hackney, J.A.; Ichikawa, R.; Nguyen, A.; Elstrott, J.; Orozco, L.D.; Sun, K.H.; Modrusan, Z.; Gogineni, A.; Scherl, A.; et al. Dual targeting of lymphocyte homing and retention through alpha4beta7 and alphaEbeta7 inhibition in inflammatory bowel disease. Cell Rep. Med. 2021, 2, 100381. [Google Scholar] [CrossRef] [PubMed]
- Novak, E.A.; Mollen, K.P. Mitochondrial dysfunction in inflammatory bowel disease. Front. Cell Dev. Biol. 2015, 3, 62. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Nag, S.; Zhang, X.; Wang, M.H.; Wang, H.; Zhou, J.; Zhang, R. Ribosomal proteins and human diseases: Pathogenesis, molecular mechanisms, and therapeutic implications. Med. Res. Rev. 2015, 35, 225–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grummt, I. The nucleolus-guardian of cellular homeostasis and genome integrity. Chromosoma 2013, 122, 487–497. [Google Scholar] [CrossRef]
- Houten, S.M.; Wanders, R.J.A.; Ranea-Robles, P. Metabolic interactions between peroxisomes and mitochondria with a special focus on acylcarnitine metabolism. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165720. [Google Scholar] [CrossRef]
- Soderman, J.; Berglind, L.; Almer, S. Inverse and Concordant Mucosal Pathway Gene Expressions in Inflamed and Non-Inflamed Ulcerative Colitis Patients: Potential Relevance to Aetiology and Pathogenesis. Int. J. Mol. Sci. 2022, 23, 6944. [Google Scholar] [CrossRef]
- Murdoch, T.; O’Donnell, S.; Silverberg, M.S.; Panaccione, R. Biomarkers as potential treatment targets in inflammatory bowel disease: A systematic review. Can. J. Gastroenterol. Hepatol. 2015, 29, 203–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.; Battat, R.; Parker, C.E.; Khanna, R.; Jairath, V.; Feagan, B.G. Update on C-reactive protein and fecal calprotectin: Are they accurate measures of disease activity in Crohn’s disease? Expert Rev. Gastroenterol. Hepatol. 2019, 13, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Arijs, I.; Li, K.; Toedter, G.; Quintens, R.; Van Lommel, L.; Van Steen, K.; Leemans, P.; De Hertogh, G.; Lemaire, K.; Ferrante, M.; et al. Mucosal gene signatures to predict response to infliximab in patients with ulcerative colitis. Gut 2009, 58, 1612–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poplawski, A.; Binder, H. Feasibility of sample size calculation for RNA-seq studies. Brief. Bioinform. 2018, 19, 713–720. [Google Scholar] [CrossRef]
- Lee, J.C.; Lyons, P.A.; McKinney, E.F.; Sowerby, J.M.; Carr, E.J.; Bredin, F.; Rickman, H.M.; Ratlamwala, H.; Hatton, A.; Rayner, T.F.; et al. Gene expression profiling of CD8+ T cells predicts prognosis in patients with Crohn disease and ulcerative colitis. J. Clin. Investig. 2011, 121, 4170–4179. [Google Scholar] [CrossRef] [Green Version]
- Schurch, N.J.; Schofield, P.; Gierlinski, M.; Cole, C.; Sherstnev, A.; Singh, V.; Wrobel, N.; Gharbi, K.; Simpson, G.G.; Owen-Hughes, T.; et al. How many biological replicates are needed in an RNA-seq experiment and which differential expression tool should you use? RNA 2016, 22, 839–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, R.; Liu, P. Sample size calculation while controlling false discovery rate for differential expression analysis with RNA-sequencing experiments. BMC Bioinform. 2016, 17, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, R.F.; Bradshaw, J.M. A simple index of Crohn’s-disease activity. Lancet 1980, 1, 514. [Google Scholar] [CrossRef]
- Walmsley, R.S.; Ayres, R.C.; Pounder, R.E.; Allan, R.N. A simple clinical colitis activity index. Gut 1998, 43, 29–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allegretti, J.R.; Barnes, E.L.; Stevens, B.; Storm, M.; Ananthakrishnan, A.; Yajnik, V.; Korzenik, J. Predictors of Clinical Response and Remission at 1 Year Among a Multicenter Cohort of Patients with Inflammatory Bowel Disease Treated with Vedolizumab. Dig. Dis. Sci. 2017, 62, 1590–1596. [Google Scholar] [CrossRef] [PubMed]
- Bragde, H.G.; Jansson, U.; Fredrikson, M.; Grodzinsky, E.; Soderman, J. Characterisation of gene and pathway expression in stabilised blood from children with coeliac disease. BMJ Open Gastroenterol. 2020, 7, e000536. [Google Scholar] [CrossRef]
- Law, C.W.; Alhamdoosh, M.; Su, S.; Dong, X.; Tian, L.; Smyth, G.K.; Ritchie, M.E. RNA-seq analysis is easy as 1-2-3 with limma, Glimma and edgeR. F1000Res 2016, 5, J-1408. [Google Scholar] [CrossRef] [Green Version]
- RStudioTeam. RStudio: Integrated Development Environment for R; RStudio, PBC: Boston, MA, USA, 2020; Available online: http://www.rstudio.com/ (accessed on 1 November 2022).
- Su, S.; Law, C.W.; Ah-Cann, C.; Asselin-Labat, M.L.; Blewitt, M.E.; Ritchie, M.E. Glimma: Interactive graphics for gene expression analysis. Bioinformatics 2017, 33, 2050–2052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Yu, G.; He, Q.Y. ReactomePA: An R/Bioconductor package for reactome pathway analysis and visualization. Mol. Biosyst. 2016, 12, 477–479. [Google Scholar] [CrossRef]
- Jassal, B.; Matthews, L.; Viteri, G.; Gong, C.; Lorente, P.; Fabregat, A.; Sidiropoulos, K.; Cook, J.; Gillespie, M.; Haw, R.; et al. The reactome pathway knowledgebase. Nucleic Acids Res. 2020, 48, D498–D503. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merico, D.; Isserlin, R.; Stueker, O.; Emili, A.; Bader, G.D. Enrichment map: A network-based method for gene-set enrichment visualization and interpretation. PLoS ONE 2010, 5, e13984. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Morris, J.H.; Apeltsin, L.; Newman, A.M.; Baumbach, J.; Wittkop, T.; Su, G.; Bader, G.D.; Ferrin, T.E. clusterMaker: A multi-algorithm clustering plugin for Cytoscape. BMC Bioinform. 2011, 12, 436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallania, F.; Tam, A.; Lofgren, S.; Schaffert, S.; Azad, T.D.; Bongen, E.; Haynes, W.; Alsup, M.; Alonso, M.; Davis, M.; et al. Leveraging heterogeneity across multiple datasets increases cell-mixture deconvolution accuracy and reduces biological and technical biases. Nat. Commun. 2018, 9, 4735. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Responders (n = 13) | p-Value T0-T1 | Non-Responders (n = 11) | p-Value T0–T1 | p-Value at T0 or T1 | ||
|---|---|---|---|---|---|---|
| Disease UC/CD | 6/7 | 3/8 | 0.42 | |||
| Gender (female/male) | 4/9 | 3/8 | 1.00 | |||
| Age (years) | 30.2 (16.6) | 37.6 (30.2) | 0.28 | |||
| Disease duration (years) | 10.2 (13.5) | 16.3 (15.2) | 0.26 | |||
| Days since last anti-TNF-α drug † | 77 (1593) | 99 (763) | 0.98 | |||
| Duration last anti-TNF-α drug (days) † | 202 (356) | 267 (903) | 0.10 | |||
| Disease activity UC | T0 | 11 (3) | 10 (5) | 0.52 | ||
| T1 | 6 (6) | 0.02 | 8 (9) | 1.00 | 0.52 | |
| 6 months ‡ | 3 (4) | 0.09 | 8 (15) | 0.79 | 1.00 | |
| Disease activity CD | T0 | 10 (5) | 7 (5) | 0.14 | ||
| T1 | 4 (4) | 0.03 | 8 (6) | 0.55 | 0.28 | |
| 6 months § | 6 (3) | 0.07 | 6 (9) | 0.11 | 0.91 | |
| Clinical remission | T0 | 1 | 2 | 0.57 | ||
| T1 | 4 | 2 | 0.65 | |||
| PGA | T0 | 2 (0) | 2 (0) | 0.69 | ||
| T1 | 1 (1) | 0.04 | 2 (1) | 0.11 | 1.00 | |
| f-Calprotectin (mg/kg feces) | T0 ¶ | 1390 (2112) | 549.5 (877) | 0.11 | ||
| T1 †† | 191 (1417) | 0.06 | 266 (269) | 0.58 | 0.96 | |
| s-CRP (mg/L) | T0 | 9.0 (21.0) | 3.0 (5.0) | 0.02 | ||
| T1 | 13.0 (14.0) | 0.23 | 3.0 (1.0) | 0.83 | 0.01 | |
| b-Leukocyte count (×109/L) | T0 | 9.2 (4.1) | 9.2 (4.0) | 0.86 | ||
| T1 ‡‡ | 8.35 (3.0) | 0.16 | 7.8 (3.4) | 0.32 | 0.98 | |
| b-Hb (g/L) | T0 | 130 (17) | 133 (22) | 0.57 | ||
| T1 §§ | 125 (25) | 0.39 | 140 (25) | 0.79 | 0.35 | |
| s-Alb (g/L) | T0 | 35 (6) | 37 (6) | 0.57 | ||
| T1 | 35 (1) | 0.48 | 36 (4) | 0.62 | 0.46 | |
| Dose VDZ (mg/kg body weight) | ||||||
| 4.1 (0.47) | 3.9 (1.2) | 0.65 | ||||
| p-VDZ at follow-up (T1) (µg/mL) | 10.5 (9.9) | 16.2 (8.1) | 0.19 |
| Pathway Enrichment | Reactome | |
|---|---|---|
| Up | Down | |
| T0 Responders vs. Non-responders | 279 | 46 |
| T1 Responders vs. Non-responders | 33 | 7 |
| Responders T1 vs. T0 | 51 | 221 |
| Non-responders T1 vs. T0 | 1 | 193 |
| Pathways Upregulated in Responders | Size | NES | FDR p-Value |
| Amino acid transport across the plasma membrane | 19 | 2.30 | 2.44 × 10−4 |
| Regulation of actin dynamics for phagocytic cup formation | 61 | 2.27 | 8.12 × 10−5 |
| FCGR3A-mediated phagocytosis | 58 | 2.27 | 4.87 × 10−5 |
| Glycosaminoglucan metabolism | 70 | 2.18 | 3.65 × 10−4 |
| EPH-Ephrin signaling | 66 | 2.13 | 8.33 × 10−4 |
| Pathways Downregulated in Responders | |||
| Mitochondrial translation initiation | 82 | −2.41 | <1.00 × 10−5 |
| Mitochondrial translation termination | 82 | −2.40 | <1.00 × 10−5 |
| Mitochondrial translation | 88 | −2.40 | <1.00 × 10−5 |
| tRNA processing | 93 | −2.26 | 3.10 × 10−5 |
| rRNA processing in the nucleus and cytosol | 171 | 2.05 | <1.00 × 10−5 |
| Pathways Upregulated in Responders | Size | NES | FDR p-Value |
| L13-mediated translational silencing of ceruloplasmin expression | 107 | 2.28 | 2.67 × 10−4 |
| GTP hydrolysis and joining of the 60S ribosomal subunit | 108 | 2.26 | 2.26 × 10−4 |
| Eukaryotic translation elongation | 87 | 2.25 | 2.42 × 10−4 |
| Peptide chain elongation | 85 | 2.24 | 2.04 × 10−4 |
| Viral mRNA translation | 85 | 2.21 | 2.35 × 10−4 |
| Pathways Downregulated in Responders | |||
| Interferon alpha beta signaling | 49 | −2.11 | 1.03 × 10−2 |
| Antigen processing cross-presentation | 94 | −1.98 | 5.31 × 10−2 |
| Interferon gamma signaling | 75 | −1.97 | 4.15 × 10−2 |
| ADP signaling through P2Y purinoceptor | 14 | −1.90 | 7.14 × 10−2 |
| ER-phagosome pathway | 80 | −1.89 | 7.01 × 10−2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haglund, S.; Söderman, J.; Almer, S. Differences in Whole-Blood Transcriptional Profiles in Inflammatory Bowel Disease Patients Responding to Vedolizumab Compared with Non-Responders. Int. J. Mol. Sci. 2023, 24, 5820. https://doi.org/10.3390/ijms24065820
Haglund S, Söderman J, Almer S. Differences in Whole-Blood Transcriptional Profiles in Inflammatory Bowel Disease Patients Responding to Vedolizumab Compared with Non-Responders. International Journal of Molecular Sciences. 2023; 24(6):5820. https://doi.org/10.3390/ijms24065820
Chicago/Turabian StyleHaglund, Sofie, Jan Söderman, and Sven Almer. 2023. "Differences in Whole-Blood Transcriptional Profiles in Inflammatory Bowel Disease Patients Responding to Vedolizumab Compared with Non-Responders" International Journal of Molecular Sciences 24, no. 6: 5820. https://doi.org/10.3390/ijms24065820