
Novel Insights into the Contribution of Cyclic Electron Flow to Cotton Bracts in Response to High Light

Abstract
:1. Introduction
2. Results
2.1. The Difference of Pigment Contents between Cotton Leaves and Bracts
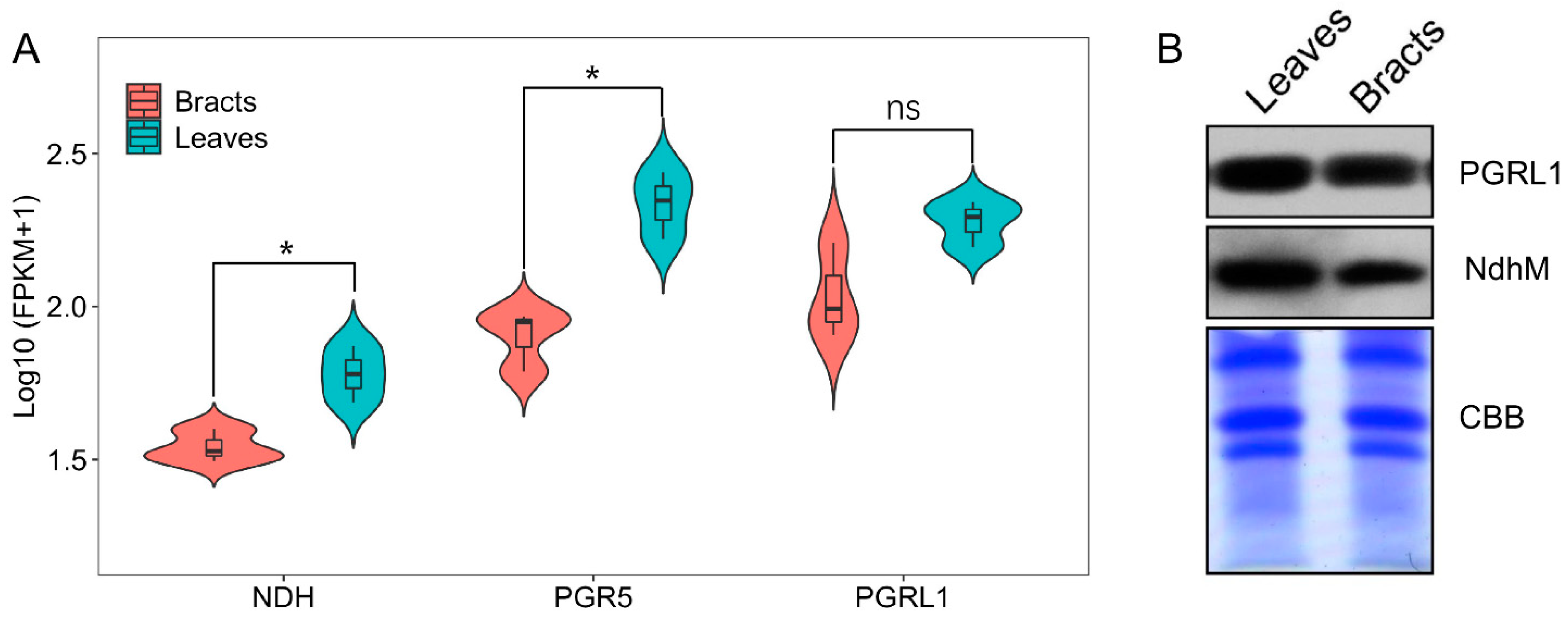
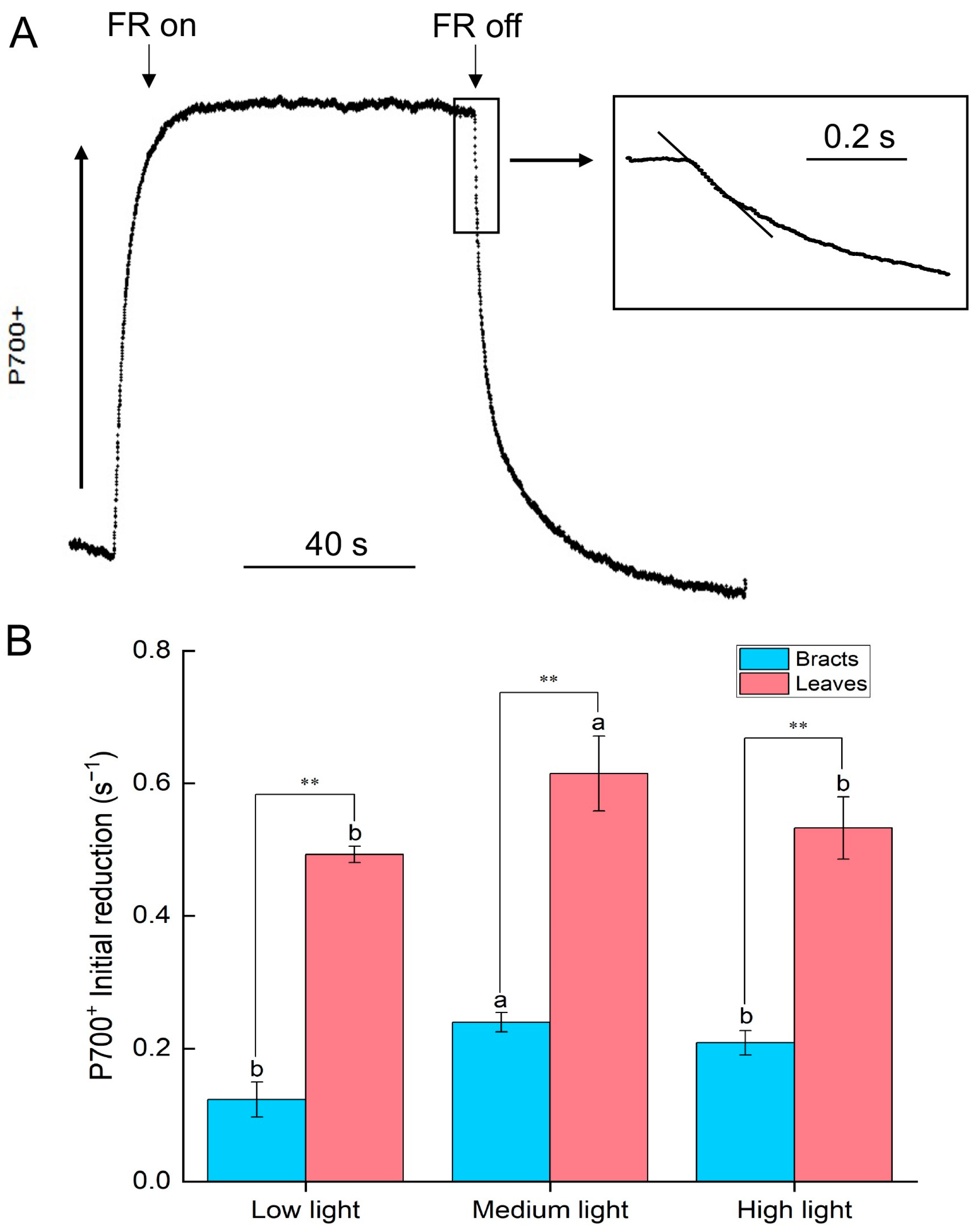
2.2. Cyclic Electron Flow Characteristics of Leaves and Bracts
2.3. Cotton Leaves Have a Higher ATP Synthase Activity Compared with Cotton Bracts
2.4. Heat Dissipation Characteristics of Leaves and Bracts
3. Discussion
3.1. Bracts’ Photoprotection Mechanisms Differ from Those of Leaves, Which Primarily Rely on Establishing a Proton Gradient across the Thylakoid Membrane through Cyclic Electron Flow to Stimulate Heat Dissipation
3.2. The Importance of Heat Dissipation Capacity in Bracts in Response to High Light
4. Materials and Methods
4.1. Plant Material
4.2. Chlorophyll Content
4.3. Transcriptome Sequencing
4.4. Isolation of Chloroplasts
4.5. Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE)
4.6. Western Blotting Analysis of Proteins
4.7. P700 Dark Reduction Curve
4.8. Post-Illumination Chlorophyll Fluorescence Transient
4.9. P515 Signal Changes
4.10. Non-Photochemical Quenching (NPQ)
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hu, L.; Zhang, Y.; Xia, H.; Fan, S.; Song, J.; Lv, X.; Kong, L. Photosynthetic characteristics of non-foliar organs in main C(3) cereals. Physiol. Plant 2019, 166, 226–239. [Google Scholar] [CrossRef]
- Kong, L.A.; Sun, M.Z.; Xie, Y.; Wang, F.H.; Zhao, Z.D. Photochemical and antioxidative responses of the glume and flag leaf to seasonal senescence in wheat. Front. Plant Sci. 2015, 6, 358. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.A.; Xie, Y.; Sun, M.Z.; Si, J.S.; Hu, L. Comparison of the photosynthetic characteristics in the pericarp and flag leaves during wheat (Triticum aestivum L.) caryopsis development. Photosynthetica 2016, 54, 40–46. [Google Scholar] [CrossRef]
- Chang, T.G.; Song, Q.F.; Zhao, H.L.; Chang, S.; Xin, C.; Qu, M.; Zhu, X.G. An in situ approach to characterizing photosynthetic gas exchange of rice panicle. Plant Methods 2020, 16, 92. [Google Scholar] [CrossRef] [PubMed]
- Constable, G.A.; Rawson, H.M. Carbon production and utilization in cotton: Inferences from a carbon budget. Aust. J. Plant Physiol. 1980, 7, 539–553. [Google Scholar] [CrossRef]
- Wullschleger, S.D.; Oosterhuis, D.M.; Hurren, R.G.; Hanson, P.J. Evidence for light-dependent recycling of respired carbon dioxide by the cotton fruit. Plant Physiol. 1991, 97, 574–579. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.Y.; Zhang, Y.L.; Luo, H.H.; Li, W.; Oguchi, R.; Fan, D.Y.; Chow, W.S.; Zhang, W.F. Important photosynthetic contribution from the non-foliar green organs in cotton at the late growth stage. Planta 2012, 235, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.L.; Feng, G.Y.; Hu, Y.Y.; Yao, Y.D.; Zhang, W.F. Photosynthetic activity and its correlation with matter production in non-foliar green organs of cotton. Acta Agron. Sin. 2010, 36, 701–708. [Google Scholar] [CrossRef]
- Yi, X.P.; Zhang, Y.L.; Yao, H.S.; Han, J.M.; Chow, W.S.; Fan, D.Y.; Zhang, W.F. Changes in activities of both photosystems and the regulatory effect of cyclic electron flow in field-grown cotton (Gossypium hirsutum L.) under water deficit. J. Plant Physiol. 2018, 220, 74–82. [Google Scholar] [CrossRef] [Green Version]
- Munekage, Y.; Hojo, M.; Meurer, J.; Endo, T.; Tasaka, M.; Shikanai, T. PGR5 is involved in cyclic electron flow around photosystem I and is essential for photoprotection in Arabidopsis. Cell 2002, 110, 361–371. [Google Scholar] [CrossRef] [Green Version]
- Alric, J. Redox and ATP control of photosynthetic cyclic electron flow in chlamydomonas reinhardtii (II) involvement of the PGR5-PGRL1 pathway under anaerobic conditions. Biochim. Biophys. Acta 2014, 1837, 825–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.X.; Zheng, F.F.; Ma, W.M.; Han, Z.G.; Gu, Q.; Shen, Y.K.; Mi, H.L. Regulation of NAD(P)H dehydrogenase-dependent cyclic electron transport around PSI by NaHSO3 at low concentrations in tobacco chloroplast. Plant Cell Physiol. 2011, 52, 1734–1743. [Google Scholar] [CrossRef] [Green Version]
- Kato, Y.; Sugimoto, K.; Shikanai, T. NDH-PSI supercomplex assembly precedes full assembly of the NDH complex in chloroplast. Plant Physiol. 2017, 176, 1728–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endo, T.; Shikanai, T.; Sato, F.; Asadaet, K. NAD(P)H dehydrogenase-dependent, antimycin a-sensitive electron donation to plastoquinone in tobacco chloroplasts. Plant Cell Physiol. 1998, 39, 1226–1231. [Google Scholar] [CrossRef]
- Wang, P.; Duan, W.; Takabayashi, A.; Endo, T.; Shikanai, T.; Ye, J.Y.; Mi, H.L. Chloroplastic NAD(P)H dehydrogenase in tobacco leaves functions in alleviation of oxidative damage caused by temperature stress. Plant Physiol. 2006, 141, 465–474. [Google Scholar] [CrossRef] [Green Version]
- Munekage, Y.; Hashimoto, M.; Miyake, C.; Tomizawa, K.; Endo, T.; Tasaka, M. Cyclic electron flow around photosystem I is essential for photosynthesis. Nature 2004, 429, 579–582. [Google Scholar] [CrossRef]
- Johnson, G.N. Reprint of: Physiology of PSI cyclic electron transport in higher plants. Biochim. Biophys. Acta-Bioenerg. 2011, 1807, 906–911. [Google Scholar] [CrossRef]
- Huang, W.; Yang, Y.J.; Zhang, S.B.; Liu, T. Cyclic electron flow around photosystem I promotes ATP synthesis possibly helping the rapid repair of photodamaged photosystem II at low light. Front. Plant Sci. 2018, 9, 239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, S.; Milward, S.E.; Fan, D.Y.; Chow, W.S.; Badger, M.R. How does cyclic electron flow alleviate photoinhibition in Arabidopsis? Plant Physiol. 2009, 149, 1560–1567. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Yang, Y.J.; Hu, H.; Zhang, S.B. Different roles of cyclic electron flow around photosystem I under sub-saturating and saturating light intensities in tobacco leaves. Front. Plant Sci. 2015, 6, 923. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.Y.; Oguchi, R.; Yamori, W.; Chow, W.S.; Zhang, W.F. Cotton bracts are adapted to a microenvironment of concentrated CO2 produced by rapid fruit respiration. Ann. Bot. 2013, 112, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhan, D.X.; Luo, H.H.; Zhang, Y.L.; Zhang, W.F. Photorespiration and photoinhibition in the bracts of cotton under water stress. Photosynthetica 2016, 54, 12–18. [Google Scholar] [CrossRef]
- Biswall, B. Carotenoid catabolism during leaf senescence and its control by light. J. Photoch. Photobio. B 1995, 30, 3–13. [Google Scholar] [CrossRef]
- Ma, W.; Mi, H. Expression and activity of type 1 NAD(P)H dehydrogenase at different growth phases of the cyanobacterium, Synechocystis PCC 6803. Physiol. Plantarum 2010, 125, 135–140. [Google Scholar] [CrossRef]
- Rumeau, D.; Becuwe, L.N.; Beyly, A.; Louwagie, M.; Garin, J.; Peltier, G. New subunits NDH-M, -N, and -O, encoded by nuclear genes, are essential for plastid Ndh complex functioning in higher plants. Plant Cell 2005, 17, 219–232. [Google Scholar] [CrossRef] [Green Version]
- Klughammer, C.; Schreiber, U. New accessory for the DUAL-PAM-100: The P515/535 module and examples of its application. PAM Appl. Notes 2008, 1, 1–10. [Google Scholar]
- Bailleul, B.; Cardol, P.; Breyton, C.; Finazzi, G. Electrochromism: A useful probe to study algal photosynthesis. Photosynth. Res. 2010, 106, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Kramer, D.M.; Sacksteder, C.A. A diffused-optics flash kinetic spectrophotometer (DOFS) for measurements of absorbance changes in intact plants in the steady-state. Photosynth. Res. 1998, 56, 103–112. [Google Scholar] [CrossRef]
- Horton, P.; Ruban, A.V. ΔpH-dependent quenching of theFolevel of chlorophyll fluorescence in spinach leaves. BBA-Bioenergetics 1993, 1142, 203–206. [Google Scholar] [CrossRef]
- Miyake, C.; Amako, K.; Shiraishi, N.; Sugimoto, T. Acclimation of tobacco leaves to high light intensity drives the plastoquinone oxidation system--relationship among the fraction of open PSII centers, non-photochemical quenching of Chl fluorescence and the maximum quantum yield of PSII in the dark. Plant Cell Physiol. 2009, 50, 730–743. [Google Scholar] [CrossRef]
- Burlacot, A.; Dao, O.; Auroy, P.; Cuine, S.; Li-Beisson, Y.; Peltier, G. Alternative photosynthesis pathways drive the algal CO(2)-concentrating mechanism. Nature 2022, 605, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Murchie, E.H.; Niyogi, K.K. Manipulation of photoprotection to improve plant photosynthesis. Plant Physiol. 2011, 155, 86–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allahverdiyeva, Y.; Suorsa, M.; Tikkanen, M.; Aro, E.M. Photoprotection of photosystems in fluctuating light intensities. J. Exp. Bot. 2015, 66, 2427–2436. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophyll fluorescence signatures of leaves during the autumnal chlorophyll breakdown. J. Plant Physiol. 1987, 131, 101–110. [Google Scholar] [CrossRef]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectrometry. Biochim. Biophys. Acta 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Bilger, W.; Björkman, O. Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis. Photosynth. Res. 1990, 25, 173–185. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Chla (μmol dm−2) | Chlb (μmol dm−2) | Chla/Chlb | Car (μmol dm−2) | ChlT (μmol dm−2) | Car/ChlT |
|---|---|---|---|---|---|---|
| Leaves | 6.64 ± 0.66 a | 2.33 ± 0.3 a | 2.85 ± 0.07 a | 1.57 ± 0.04 a | 8.98 ± 0.96 a | 0.17 ± 0.02 b |
| Bracts | 0.89 ± 0.08 b | 0.31 ± 0.04 b | 2.92 ± 0.15 a | 0.56 ± 0.56 b | 1.19 ± 0.12 b | 0.47 ± 0.03 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Ma, W.; Zhang, W.; Zhang, Y. Novel Insights into the Contribution of Cyclic Electron Flow to Cotton Bracts in Response to High Light. Int. J. Mol. Sci. 2023, 24, 5589. https://doi.org/10.3390/ijms24065589
Li X, Ma W, Zhang W, Zhang Y. Novel Insights into the Contribution of Cyclic Electron Flow to Cotton Bracts in Response to High Light. International Journal of Molecular Sciences. 2023; 24(6):5589. https://doi.org/10.3390/ijms24065589
Chicago/Turabian StyleLi, Xiafei, Weimin Ma, Wangfeng Zhang, and Yali Zhang. 2023. "Novel Insights into the Contribution of Cyclic Electron Flow to Cotton Bracts in Response to High Light" International Journal of Molecular Sciences 24, no. 6: 5589. https://doi.org/10.3390/ijms24065589