
CD73: Friend or Foe in Lung Injury
Abstract
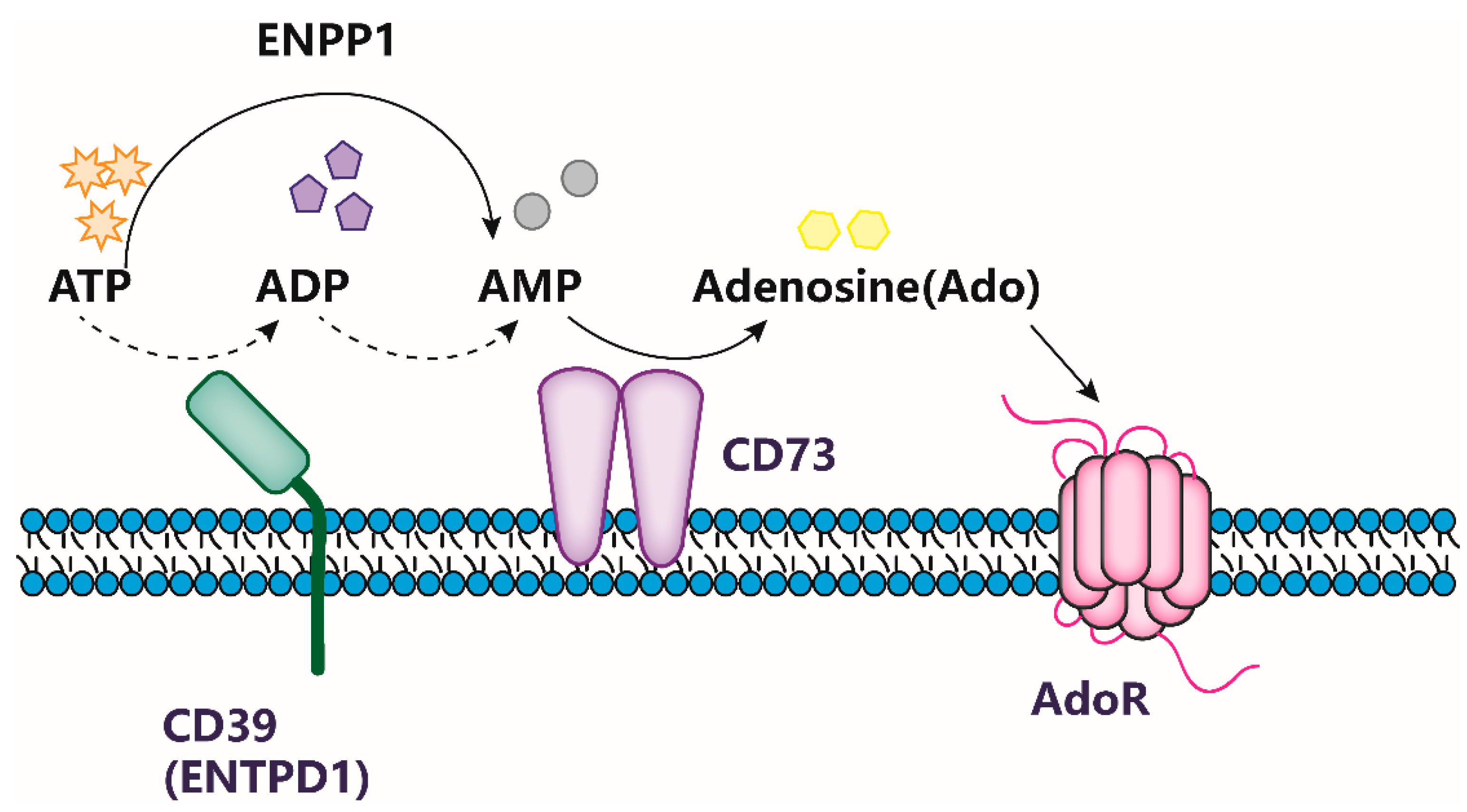
:1. Introduction
2. CD73 Exerts Bidirectional Modulatory Effects on Lung Injury
2.1. CD73 Enhances Lung Injury via Boosting Cell Invasion and Tumor Formation
2.2. CD73 Maintains Lung Tissue-Specific Homeostasis and Respiratory Function
2.3. CD73 Protects against Lung Injury in Hypoxia and Hyperoxia Condition
2.4. CD73 in Inflammation-Driven Lung Injury: A Double-Edged Sword
2.5. Blocking of CD73 Alleviates Carcinoma-Potentiated Lung Injury
3. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yue, N.; Huang, H.; Zhu, X.; Han, Q.; Wang, Y.; Li, B.; Liu, Q.; Wu, G.; Zhang, Y.; Yu, J. Activation of P2X7 receptor and NLRP3 inflammasome assembly in hippocampal glial cells mediates chronic stress-induced depressive-like behaviors. J. Neuroinflamm. 2017, 14, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gulland, J.M.; Jackson, E.M. 5-Nucleotidase. Biochem. J. 1938, 32, 597–601. [Google Scholar] [CrossRef] [Green Version]
- Thompson, L.F.; Ruedi, J.M.; Glass, A.; Low, M.G.; Lucas, A.H. Antibodies to 5′-nucleotidase (CD73), a glycosyl-phosphatidylinositol-anchored protein, cause human peripheral blood T cells to proliferate. J. Immunol. 1989, 143, 1815–1821. [Google Scholar] [CrossRef]
- Minor, M.; Alcedo, K.P.; Battaglia, R.A.; Snider, N.T. Cell type- and tissue-specific functions of ecto-5′-nucleotidase (CD73). Am. J. Physiol. Cell Physiol. 2019, 317, C1079–C1092. [Google Scholar] [CrossRef]
- Sadej, R.; Skladanowski, A.C. Dual, enzymatic and non-enzymatic, function of ecto-5′-nucleotidase (eN, CD73) in migration and invasion of A375 melanoma cells. Acta Biochim. Pol. 2012, 59, 647–652. [Google Scholar] [CrossRef] [Green Version]
- Sträter, N. Ecto-5′-nucleotidase: Structure function relationships. Purinergic Signal. 2006, 2, 343–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heuts, D.P.; Weissenborn, M.J.; Olkhov, R.V.; Shaw, A.M.; Gummadova, J.; Levy, C.; Scrutton, N.S. Crystal structure of a soluble form of human CD73 with ecto-5′-nucleotidase activity. Chembiochem 2012, 13, 2384–2391. [Google Scholar] [CrossRef]
- Yegutkin, G.G. Nucleotide- and nucleoside-converting ectoenzymes: Important modulators of purinergic signalling cascade. Biochim. Biophys. Acta 2008, 1783, 673–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Zhang, Y.; Wu, H.; D’Alessandro, A.; Yegutkin, G.G.; Song, A.; Sun, K.; Li, J.; Cheng, N.Y.; Huang, A.; et al. Beneficial Role of Erythrocyte Adenosine A2B Receptor-Mediated AMP-Activated Protein Kinase Activation in High-Altitude Hypoxia. Circulation 2016, 134, 405–421. [Google Scholar] [CrossRef]
- Antonioli, L.; Pacher, P.; Vizi, E.S.; Haskó, G. CD39 and CD73 in immunity and inflammation. Trends Mol. Med. 2013, 19, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Airas, L.; Niemelä, J.; Salmi, M.; Puurunen, T.; Smith, D.J.; Jalkanen, S. Differential regulation and function of CD73, a glycosyl-phosphatidylinositol-linked 70-kD adhesion molecule, on lymphocytes and endothelial cells. J. Cell Biol. 1997, 136, 421–431. [Google Scholar] [CrossRef] [Green Version]
- Koszalka, P.; Ozüyaman, B.; Huo, Y.; Zernecke, A.; Flögel, U.; Braun, N.; Buchheiser, A.; Decking, U.K.; Smith, M.L.; Sévigny, J.; et al. Targeted disruption of cd73/ecto-5′-nucleotidase alters thromboregulation and augments vascular inflammatory response. Circ. Res. 2004, 95, 814–821. [Google Scholar] [CrossRef] [Green Version]
- Ålgars, A.; Karikoski, M.; Yegutkin, G.G.; Stoitzner, P.; Niemelä, J.; Salmi, M.; Jalkanen, S. Different role of CD73 in leukocyte trafficking via blood and lymph vessels. Blood 2011, 117, 4387–4393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Resta, R.; Yamashita, Y.; Thompson, L.F. Ecto-enzyme and signaling functions of lymphocyte CD73. Immunol. Rev. 1998, 161, 95–109. [Google Scholar] [CrossRef]
- Zimmermann, H. History of ectonucleotidases and their role in purinergic signaling. Biochem. Pharmacol. 2021, 187, 114322. [Google Scholar] [CrossRef]
- Zimmermann, H. Ectonucleoside triphosphate diphosphohydrolases and ecto-5′-nucleotidase in purinergic signaling: How the field developed and where we are now. Purinergic Signal. 2021, 17, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Borea, P.A.; Gessi, S.; Merighi, S.; Vincenzi, F.; Varani, K. Pharmacology of Adenosine Receptors: The State of the Art. Physiol. Rev. 2018, 98, 1591–1625. [Google Scholar] [CrossRef] [Green Version]
- Horenstein, A.L.; Chillemi, A.; Zaccarello, G.; Bruzzone, S.; Quarona, V.; Zito, A.; Serra, S.; Malavasi, F. A CD38/CD203a/CD73 ectoenzymatic pathway independent of CD39 drives a novel adenosinergic loop in human T lymphocytes. Oncoimmunology 2013, 2, e26246. [Google Scholar] [CrossRef] [Green Version]
- Tian, Z.; Dixon, J.; Guo, X.; Deal, B.; Liao, Q.; Zhou, Y.; Cheng, F.; Allen-Gipson, D.S. Co-inhibition of CD73 and ADORA2B Improves Long-Term Cigarette Smoke Induced Lung Injury. Front. Physiol. 2021, 12, 614330. [Google Scholar] [CrossRef] [PubMed]
- Franciosi, M.L.M.; Lima, M.D.M.; Schetinger, M.R.C.; Cardoso, A.M. Possible role of purinergic signaling in COVID-19. Mol. Cell. Biochem. 2021, 476, 2891–2898. [Google Scholar] [CrossRef]
- Allard, B.; Turcotte, M.; Spring, K.; Pommey, S.; Royal, I.; Stagg, J. Anti-CD73 therapy impairs tumor angiogenesis. Int. J. Cancer 2014, 134, 1466–1473. [Google Scholar] [CrossRef]
- Beavis, P.A.; Stagg, J.; Darcy, P.K.; Smyth, M.J. CD73: A potent suppressor of antitumor immune responses. Trends Immunol. 2012, 33, 231–237. [Google Scholar] [CrossRef]
- Yegutkin, G.G.; Marttila-Ichihara, F.; Karikoski, M.; Niemelä, J.; Laurila, J.P.; Elima, K.; Jalkanen, S.; Salmi, M. Altered purinergic signaling in CD73-deficient mice inhibits tumor progression. Eur. J. Immunol. 2011, 41, 1231–1241. [Google Scholar] [CrossRef]
- Stagg, J.; Divisekera, U.; Duret, H.; Sparwasser, T.; Teng, M.W.; Darcy, P.K.; Smyth, M.J. CD73-deficient mice have increased antitumor immunity and are resistant to experimental metastasis. Cancer Res. 2011, 71, 2892–2900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stagg, J.; Divisekera, U.; McLaughlin, N.; Sharkey, J.; Pommey, S.; Denoyer, D.; Dwyer, K.M.; Smyth, M.J. Anti-CD73 antibody therapy inhibits breast tumor growth and metastasis. Proc. Natl. Acad. Sci. USA 2010, 107, 1547–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terp, M.G.; Olesen, K.A.; Arnspang, E.C.; Lund, R.R.; Lagerholm, B.C.; Ditzel, H.J.; Leth-Larsen, R. Anti-human CD73 monoclonal antibody inhibits metastasis formation in human breast cancer by inducing clustering and internalization of CD73 expressed on the surface of cancer cells. J. Immunol. 2013, 191, 4165–4173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattarai, S.; Freundlieb, M.; Pippel, J.; Lund, R.R.; Lagerholm, B.C.; Ditzel, H.J.; Leth-Larsen, R. α,β-Methylene-ADP (AOPCP) Derivatives and Analogues: Development of Potent and Selective ecto-5′-Nucleotidase (CD73) Inhibitors. J. Med. Chem. 2015, 58, 6248–6263. [Google Scholar] [CrossRef]
- Allard, B.; Pommey, S.; Smyth, M.J.; Stagg, J. Targeting CD73 enhances the antitumor activity of anti-PD-1 and anti-CTLA-4 mAbs. Clin. Cancer Res. 2013, 19, 5626–5635. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Min, Y.K.; Jang, J.; Park, H.; Lee, S.; Lee, C.H. Single-cell RNA sequencing reveals distinct cellular factors for response to immunotherapy targeting CD73 and PD-1 in colorectal cancer. J. Immunother. Cancer 2021, 9, e002503. [Google Scholar] [CrossRef] [PubMed]
- Tu, E.; McGlinchey, K.; Wang, J.; Martin, P.; Ching, S.L.; Floc’h, N.; Kurasawa, J.; Starrett, J.H.; Lazdun, Y.; Wetzel, L.; et al. Anti-PD-L1 and anti-CD73 combination therapy promotes T cell response to EGFR-mutated NSCLC. JCI Insight 2022, 7, e142843. [Google Scholar] [CrossRef]
- Colgan, S.P.; Eltzschig, H.K.; Eckle, T.; Thompson, L.F. Physiological roles for ecto-5′-nucleotidase (CD73). Purinergic Signal. 2006, 2, 351–360. [Google Scholar] [CrossRef] [Green Version]
- Eckle, T.; Füllbier, L.; Wehrmann, M.; Khoury, J.; Mittelbronn, M.; Ibla, J.; Rosenberger, P.; Eltzschig, H.K. Identification of ectonucleotidases CD39 and CD73 in innate protection during acute lung injury. J. Immunol. 2007, 178, 8127–8137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picher, M.; Burch, L.H.; Hirsh, A.J.; Spychala, J.; Boucher, R.C. Ecto 5′-nucleotidase and nonspecific alkaline phosphatase. Two AMP-hydrolyzing ectoenzymes with distinct roles in human airways. J. Biol. Chem. 2003, 278, 13468–13479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morse, D.M.; Smullen, J.L.; Davis, C.W. Differential effects of UTP, ATP, and adenosine on ciliary activity of human nasal epithelial cells. Am. J. Physiol. Cell Physiol. 2001, 280, C1485–C1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazarowski, E.R.; Mason, S.J.; Clarke, L.; Harden, T.K.; Boucher, R.C. Adenosine receptors on human airway epithelia and their relationship to chloride secretion. Br. J. Pharmacol. 1992, 106, 774–782. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, L.; Correia, A.; Cristina Costa, A.; Guerra-Gomes, S.; Ferreirinha, F.; Magalhães-Cardoso, M.T.; Vilanova, M.; Correia-de-Sá, P. Deficits in endogenous adenosine formation by ecto-5′-nucleotidase/CD73 impair neuromuscular transmission and immune competence in experimental autoimmune myasthenia gravis. Mediat. Inflamm. 2015, 2015, 460610. [Google Scholar] [CrossRef]
- Hanidziar, D.; Robson, S.C. Hyperoxia and modulation of pulmonary vascular and immune responses in COVID-19. Am. J. Physiol. Lung Cell. Mol. Physiol. 2021, 320, L12–L16. [Google Scholar] [CrossRef] [PubMed]
- Dylag, A.M.; Haak, J.; Yee, M.; O’Reilly, M.A. Pulmonary mechanics and structural lung development after neonatal hyperoxia in mice. Pediatr. Res. 2020, 87, 1201–1210. [Google Scholar] [CrossRef]
- Kanthi, Y.; Knight, J.S.; Zuo, Y.; Pinsky, D.J. New (re)purpose for an old drug: Purinergic modulation may extinguish the COVID-19 thromboinflammatory firestorm. JCI Insight 2020, 5, e140971. [Google Scholar] [CrossRef]
- Afsar, B.; Kanbay, M.; Afsar, R.E. Hypoxia inducible factor-1 protects against COVID-19: A hypothesis. Med. Hypotheses 2020, 143, 109857. [Google Scholar] [CrossRef] [PubMed]
- Keskinidou, C.; Lotsios, N.S.; Vassiliou, A.G.; Dimopoulou, I.; Kotanidou, A.; Orfanos, S.E. The Interplay between Aquaporin-1 and the Hypoxia-Inducible Factor 1α in a Lipopolysaccharide-Induced Lung Injury Model in Human Pulmonary Microvascular Endothelial Cells. Int. J. Mol. Sci. 2022, 23, 10588. [Google Scholar] [CrossRef] [PubMed]
- Singer, M.; Young, P.J.; Laffey, J.G.; Asfar, P.; Taccone, F.S.; Skrifvars, M.B.; Meyhoff, C.S.; Radermacher, P. Dangers of hyperoxia. Crit. Care 2021, 25, 440. [Google Scholar] [CrossRef]
- Amarelle, L.; Quintela, L.; Hurtado, J.; Malacrida, L. Hyperoxia and Lungs: What We Have Learned from Animal Models. Front. Med. 2021, 8, 606678. [Google Scholar] [CrossRef]
- Thompson, L.F.; Eltzschig, H.K.; Ibla, J.C.; Van De Wiele, C.J.; Resta, R.; Morote-Garcia, J.C.; Colgan, S.P. Crucial role for ecto-5′-nucleotidase (CD73) in vascular leakage during hypoxia. J. Exp. Med. 2004, 200, 1395–1405. [Google Scholar] [CrossRef]
- Kiss, J.; Yegutkin, G.G.; Koskinen, K.; Savunen, T.; Jalkanen, S.; Salmi, M. IFN-beta protects from vascular leakage via up-regulation of CD73. Eur. J. Immunol. 2007, 37, 3334–3338. [Google Scholar] [CrossRef]
- Bellingan, G.; Maksimow, M.; Howell, D.C.; Stotz, M.; Beale, R.; Beatty, M.; Walsh, T.; Binning, A.; Davidson, A.; Kuper, M.; et al. The effect of intravenous interferon-beta-1a (FP-1201) on lung CD73 expression and on acute respiratory distress syndrome mortality: An open-label study. Lancet Respir. Med. 2014, 2, 98–107. [Google Scholar] [CrossRef]
- Li, H.; Karmouty-Quintana, H.; Chen, N.Y.; Mills, T.; Molina, J.; Blackburn, M.R.; Davies, J. Loss of CD73-mediated extracellular adenosine production exacerbates inflammation and abnormal alveolar development in newborn mice exposed to prolonged hyperoxia. Pediatr. Res. 2017, 82, 1039–1047. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, B.J.; Faraoni, E.Y.; Strickland, L.N.; Ma, Z.; Mota, V.; Mota, S.; Chen, X.; Mills, T.; Eltzschig, H.K.; DelGiorno, K.E.; et al. CD73-generated extracellular adenosine promotes resolution of neutrophil-mediated tissue injury and restrains metaplasia in pancreatitis. FASEB J. 2023, 37, e22684. [Google Scholar] [CrossRef]
- Xu, S.; Wang, J.; Zhong, J.; Shao, M.; Jiang, J.; Song, J.; Zhu, W.; Zhang, F.; Xu, H.; Xu, G.; et al. CD73 alleviates GSDMD-mediated microglia pyroptosis in spinal cord injury through PI3K/AKT/Foxo1 signaling. Clin. Transl. Med. 2021, 11, e269. [Google Scholar] [CrossRef] [PubMed]
- Aeffner, F.; Woods, P.S.; Davis, I.C. Activation of A1-adenosine receptors promotes leukocyte recruitment to the lung and attenuates acute lung injury in mice infected with influenza A/WSN/33 (H1N1) virus. J. Virol. 2014, 88, 10214–10227. [Google Scholar] [CrossRef] [Green Version]
- Ranieri, V.M.; Pettilä, V.; Karvonen, M.K.; Jalkanen, J.; Nightingale, P.; Brealey, D.; Mancebo, J.; Ferrer, R.; Mercat, A.; Patroniti, N.; et al. Effect of Intravenous Interferon β-1a on Death and Days Free from Mechanical Ventilation among Patients with Moderate to Severe Acute Respiratory Distress Syndrome: A Randomized Clinical Trial. JAMA 2020, 323, 725–733. [Google Scholar] [CrossRef]
- Jalkanen, J.; Pettilä, V.; Huttunen, T.; Hollmén, M.; Jalkanen, S. Glucocorticoids inhibit type I IFN beta signaling and the upregulation of CD73 in human lung. Intensive Care Med. 2020, 46, 1937–1940. [Google Scholar] [CrossRef]
- Knapp, S.; Leemans, J.C.; Florquin, S.; Branger, J.; Maris, N.A.; Pater, J.; van Rooijen, N.; van der Poll, T. Alveolar macrophages have a protective antiinflammatory role during murine pneumococcal pneumonia. Am. J. Respir. Crit. Care Med. 2003, 167, 171–179. [Google Scholar] [CrossRef] [Green Version]
- Rolston, K.V. The spectrum of pulmonary infections in cancer patients. Curr. Opin. Oncol. 2001, 13, 218–223. [Google Scholar] [CrossRef]
- Bou Ghanem, E.N.; Clark, S.; Roggensack, S.E.; McIver, S.R.; Alcaide, P.; Haydon, P.G.; Leong, J.M. Extracellular Adenosine Protects against Streptococcus pneumoniae Lung Infection by Regulating Pulmonary Neutrophil Recruitment. PLoS Pathog. 2015, 11, e1005126. [Google Scholar] [CrossRef] [PubMed]
- Siwapornchai, N.; Lee, J.N.; Tchalla, E.Y.I.; Bhalla, M.; Yeoh, J.H.; Roggensack, S.E.; Leong, J.M.; Bou Ghanem, E.N. Extracellular adenosine enhances the ability of PMNs to kill Streptococcus pneumoniae by inhibiting IL-10 production. J. Leukoc. Biol. 2020, 108, 867–882. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.A.; Spina, D.; Page, C.P. Adenosine receptors and asthma. Br. J. Pharmacol. 2008, 153 (Suppl. 1), S446–S456. [Google Scholar] [CrossRef] [Green Version]
- Pelleg, A. Extracellular adenosine 5′-triphosphate in pulmonary disorders. Biochem. Pharmacol. 2021, 187, 114319. [Google Scholar] [CrossRef] [PubMed]
- Rossi, A.; Caiazzo, E.; Bilancia, R.; Riemma, M.A.; Pagano, E.; Cicala, C.; Ialenti, A.; Zjawiony, J.K.; Izzo, A.A.; Capasso, R.; et al. Salvinorin A Inhibits Airway Hyperreactivity Induced by Ovalbumin Sensitization. Front. Pharmacol. 2017, 7, 525. [Google Scholar] [CrossRef] [Green Version]
- Cicala, C.; Ialenti, A. Adenosine signaling in airways: Toward a promising antiasthmatic approach. Eur. J. Pharmacol. 2013, 714, 522–525. [Google Scholar] [CrossRef] [Green Version]
- Caiazzo, E.; Cerqua, I.; Riemma, M.A.; Turiello, R.; Ialenti, A.; Schrader, J.; Fiume, G.; Caiazza, C.; Roviezzo, F.; Morello, S.; et al. Exacerbation of Allergic Airway Inflammation in Mice Lacking ECTO-5′-Nucleotidase (CD73). Front. Pharmacol. 2020, 11, 589343. [Google Scholar] [CrossRef] [PubMed]
- Montuori-Andrade, A.C.M.; Nolasco, A.E.; Malacco, N.L.S.O.; Vaz, L.G.; Afonso, L.C.C.; Russo, R.C.; Vieira, L.Q.; Dos Santos, L.M. Lactobacillus delbrueckii UFV-H2b20 increases IFN-γ production and CD39+ CD73+ Treg cell numbers in lungs, and protects mice against experimental allergic asthma. Immunobiology 2022, 227, 152284. [Google Scholar] [CrossRef]
- Roviezzo, F.; Bertolino, A.; Sorrentino, R.; Terlizzi, M.; Matteis, M.; Calderone, V.; Mattera, V.; Martelli, A.; Spaziano, G.; Pinto, A.; et al. Hydrogen sulfide inhalation ameliorates allergen induced airway hypereactivity by modulating mast cell activation. Pharmacol. Res. 2015, 100, 85–92. [Google Scholar] [CrossRef]
- Le, T.T.; Berg, N.K.; Harting, M.T.; Li, X.; Eltzschig, H.K.; Yuan, X. Purinergic Signaling in Pulmonary Inflammation. Front. Immunol. 2019, 10, 1633. [Google Scholar] [CrossRef]
- Cronstein, B.N.; Sitkovsky, M. Adenosine and adenosine receptors in the pathogenesis and treatment of rheumatic diseases. Nat. Rev. Rheumatol. 2017, 13, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Garcia, L.; Olle, L.; Martin, M.; Roca-Ferrer, J.; Muñoz-Cano, R. Adenosine Signaling in Mast Cells and Allergic Diseases. Int. J. Mol. Sci. 2021, 22, 5203. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Salehi-Rad, R.; Li, R.; Paul, M.K.; Dubinett, S.M.; Liu, B. The Biology of Lung Cancer: Development of More Effective Methods for Prevention, Diagnosis, and Treatment. Clin. Chest Med. 2020, 41, 25–38. [Google Scholar] [CrossRef]
- Kotas, M.E.; Medzhitov, R. Homeostasis, inflammation, and disease susceptibility. Cell 2015, 160, 816–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnstock, G. Purinergic Signalling: Therapeutic Developments. Front. Pharmacol. 2017, 8, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allard, B.; Allard, D.; Buisseret, L.; Stagg, J. The adenosine pathway in immuno-oncology. Nat. Rev. Clin. Oncol. 2020, 17, 611–629. [Google Scholar] [CrossRef] [PubMed]
- Young, A.; Ngiow, S.F.; Gao, Y.; Patch, A.M.; Barkauskas, D.S.; Messaoudene, M.; Lin, G.; Coudert, J.D.; Stannard, K.A.; Zitvogel, L.; et al. A2AR Adenosine Signaling Suppresses Natural Killer Cell Maturation in the Tumor Microenvironment. Cancer Res. 2018, 78, 1003–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maj, T.; Wang, W.; Crespo, J.; Zhang, H.; Wang, W.; Wei, S.; Zhao, L.; Vatan, L.; Shao, I.; Szeliga, W.; et al. Oxidative stress controls regulatory T cell apoptosis and suppressor activity and PD-L1-blockade resistance in tumor. Nat. Immunol. 2017, 18, 1332–1341. [Google Scholar] [CrossRef] [Green Version]
- Alter, C.; Ding, Z.; Flögel, U.; Scheller, J.; Schrader, J. A2bR-dependent signaling alters immune cell composition and enhances IL-6 formation in the ischemic heart. Am. J. Physiol. Heart Circ. Physiol. 2019, 317, H190–H200. [Google Scholar] [CrossRef]
- Vijayan, D.; Young, A.; Teng, M.W.L.; Smyth, M.J. Targeting immunosuppressive adenosine in cancer. Nat. Rev. Cancer 2017, 17, 709–724. [Google Scholar] [CrossRef]
- Giatromanolaki, A.; Kouroupi, M.; Pouliliou, S.; Mitrakas, A.; Hasan, F.; Pappa, A.; Koukourakis, M.I. Ectonucleotidase CD73 and CD39 expression in non-small cell lung cancer relates to hypoxia and immunosuppressive pathways. Life Sci. 2020, 259, 118389. [Google Scholar] [CrossRef]
- Fortunato, O.; Belisario, D.C.; Compagno, M.; Giovinazzo, F.; Bracci, C.; Pastorino, U.; Horenstein, A.; Malavasi, F.; Ferracini, R.; Scala, S.; et al. CXCR4 Inhibition Counteracts Immunosuppressive Properties of Metastatic NSCLC Stem Cells. Front. Immunol. 2020, 11, 02168. [Google Scholar] [CrossRef]
- O’Connor, R.A.; Chauhan, V.; Mathieson, L.; Titmarsh, H.; Koppensteiner, L.; Young, I.; Tagliavini, G.; Dorward, D.A.; Prost, S.; Dhaliwal, K.; et al. T cells drive negative feedback mechanisms in cancer associated fibroblasts, promoting expression of co-inhibitory ligands, CD73 and IL-27 in non-small cell lung cancer. Oncoimmunology 2021, 10, 1940675. [Google Scholar] [CrossRef]
- Isomoto, K.; Haratani, K.; Hayashi, H.; Shimizu, S.; Tomida, S.; Niwa, T.; Yokoyama, T.; Fukuda, Y.; Chiba, Y.; Kato, R.; et al. Impact of EGFR-TKI Treatment on the Tumor Immune Microenvironment in EGFR Mutation-Positive Non-Small Cell Lung Cancer. Clin. Cancer Res. 2020, 26, 2037–2046. [Google Scholar] [CrossRef] [PubMed]
- Rocha, P.; Salazar, R.; Zhang, J.; Ledesma, D.; Solorzano, J.L.; Mino, B.; Villalobos, P.; Dejima, H.; Douse, D.Y.; Diao, L.; et al. CD73 expression defines immune, molecular, and clinicopathological subgroups of lung adenocarcinoma. Cancer Immunol. Immunother. 2021, 70, 1965–1976. [Google Scholar] [CrossRef]
- Wang, L.; Dorn, P.; Simillion, C.; Froment, L.; Berezowska, S.; Tschanz, S.A.; Haenni, B.; Blank, F.; Wotzkow, C.; Peng, R.W.; et al. EpCAM+CD73+ mark epithelial progenitor cells in postnatal human lung and are associated with pathogenesis of pulmonary disease including lung adenocarcinoma. Am. J. Physiol. Lung Cell. Mol. Physiol. 2020, 319, L794–L809. [Google Scholar] [CrossRef] [PubMed]
- Taromi, S.; Firat, E.; Simonis, A.; Braun, L.M.; Apostolova, P.; Elze, M.; Passlick, B.; Schumacher, A.; Lagies, S.; Frey, A.; et al. Enhanced AC133-specific CAR T cell therapy induces durable remissions in mice with metastatic small cell lung cancer. Cancer Lett. 2022, 538, 215697. [Google Scholar] [CrossRef]
- Umemura, S.; Chen, V.; Chahine, J.J.; Kallakury, B.; Zhao, X.; Lee, H.; Avantaggiati, M.L.; He, Y.; Wang, C.; Giaccone, G. Arginase Pathway Markers of Immune-Microenvironment in Thymic Epithelial Tumors and Small Cell Lung Cancer. Clin. Lung Cancer 2022, 23, e140–e147. [Google Scholar] [CrossRef]
- Neo, S.Y.; Yang, Y.; Record, J.; Ma, R.; Chen, X.; Chen, Z.; Tobin, N.P.; Blake, E.; Seitz, C.; Thomas, R.; et al. CD73 immune checkpoint defines regulatory NK cells within the tumor microenvironment. J. Clin. Investig. 2020, 130, 1185–1198. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Lupo, K.B.; Chambers, A.M.; Matosevic, S. Purinergic targeting enhances immunotherapy of CD73+ solid tumors with piggyBac-engineered chimeric antigen receptor natural killer cells. J. Immunother. Cancer 2018, 6, 136. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.F.; Wu, D.; Wang, J.; Ba, L.; Tian, C.; Liu, Y.T.; Hu, Y.; Liu, L. Development and validation of an individualized immune prognostic model in stage I-III lung squamous cell carcinoma. Sci. Rep. 2021, 11, 12727. [Google Scholar] [CrossRef]
- Jin, H.; Lee, J.S.; Kim, D.C.; Ko, Y.S.; Lee, G.W.; Kim, H.J. Increased Extracellular Adenosine in Radiotherapy-Resistant Breast Cancer Cells Enhances Tumor Progression through A2AR-Akt-β-Catenin Signaling. Cancers 2021, 13, 2105. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Z.; Li, X.; Kang, N.; Yang, Y.; Chen, C.; Wu, T.; Zhao, M.; Liu, Y.; Ji, X. A Novel Specific Anti-CD73 Antibody Inhibits Triple-Negative Breast Cancer Cell Motility by Regulating Autophagy. Int. J. Mol. Sci. 2019, 20, 1057. [Google Scholar] [CrossRef] [Green Version]
- Petruk, N.; Tuominen, S.; Åkerfelt, M.; Mattsson, J.; Sandholm, J.; Nees, M.; Yegutkin, G.G.; Jukkola, A.; Tuomela, J.; Selander, K.S. CD73 facilitates EMT progression and promotes lung metastases in triple-negative breast cancer. Sci. Rep. 2021, 11, 6035. [Google Scholar] [CrossRef]
- Tsukui, H.; Horie, H.; Koinuma, K.; Ohzawa, H.; Sakuma, Y.; Hosoya, Y.; Yamaguchi, H.; Yoshimura, K.; Lefor, A.K.; Sata, N.; et al. CD73 blockade enhances the local and abscopal effects of radiotherapy in a murine rectal cancer model. BMC Cancer 2020, 20, 411. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Akdemir, I.; Fan, J.; Linden, J.; Zhang, B.; Cekic, C. The Expression of Adenosine A2B Receptor on Antigen-Presenting Cells Suppresses CD8+ T-cell Responses and Promotes Tumor Growth. Cancer Immunol. Res. 2020, 8, 1064–1074. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.W.; Liu, C.; Yang, L.; Chen, H.C.; Yang, L.F.; Zhang, H.Z.; Dong, K. CD73 Severed as a Potential Prognostic Marker and Promote Lung Cancer Cells Migration via Enhancing EMT Progression. Front. Genet. 2021, 12, 728200. [Google Scholar] [CrossRef]
- Barker, K.A.; Etesami, N.S.; Shenoy, A.T.; Arafa, E.I.; Lyon de Ana, C.; Smith, N.M.; Martin, I.M.; Goltry, W.N.; Barron, A.M.; Browning, J.L.; et al. Lung-resident memory B cells protect against bacterial pneumonia. J. Clin. Investig. 2021, 131, e141810. [Google Scholar] [CrossRef]
- Shah, T.; Qin, S.; Vashi, M.; Predescu, D.N.; Jeganathan, N.; Bardita, C.; Ganesh, B.; diBartolo, S.; Fogg, L.F.; Balk, R.A.; et al. Alk5/Runx1 signaling mediated by extracellular vesicles promotes vascular repair in acute respiratory distress syndrome. Clin. Transl. Med. 2018, 7, 19. [Google Scholar] [CrossRef] [PubMed]
- Raza, M.Z.; Cadassou, O.; Dumontet, C.; Cros-Perrial, E.; Jordheim, L.P. CD73 and cN-II regulate the cellular response to chemotherapeutic and hypoxic stress in lung adenocarcinoma cells. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129842. [Google Scholar] [CrossRef]
- Caiazzo, E.; Cerqua, I.; Turiello, R.; Riemma, M.A.; De Palma, G.; Ialenti, A.; Roviezzo, F.; Morello, S.; Cicala, C. Lack of Ecto-5′-Nucleotidase Protects Sensitized Mice against Allergen Challenge. Biomolecules 2022, 12, 697. [Google Scholar] [CrossRef]
- Cho, J.W.; Park, K.S.; Bae, J.Y. Effects of Wharton’s jelly-derived mesenchymal stem cells on chronic obstructive pulmonary disease. Regen. Ther. 2019, 11, 207–211. [Google Scholar] [CrossRef]
- Meziane, O.; Salahuddin, S.; Pham, T.N.Q.; Farnos, O.; Pagliuzza, A.; Olivenstein, R.; Thomson, E.; Alexandrova, Y.; Orlova, M.; Schurr, E.; et al. HIV Infection and Persistence in Pulmonary Mucosal Double Negative T Cells In Vivo. J. Virol. 2020, 94, e01788-20. [Google Scholar] [CrossRef]
- Gogusev, J.; Lepelletier, Y.; Khattabi, L.E.; Grigoroiu, M.; Validire, P. Establishment and Characterization of a Stromal Cell Line Derived from a Patient with Thoracic Endometriosis. Reprod. Sci. 2020, 27, 1627–1636. [Google Scholar] [CrossRef]
- Amaral, E.P.; Machado de Salles, É.; Barbosa Bomfim, C.C.; Salgado, R.M.; Almeida, F.M.; de Souza, P.C.; Alvarez, J.M.; Hirata, M.H.; Lasunskaia, E.B.; D’Império-Lima, M.R. Inhibiting Adenosine Receptor Signaling Promotes Accumulation of Effector CD4+ T Cells in the Lung Parenchyma during Severe Tuberculosis. J. Infect. Dis. 2019, 219, 964–974. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Fan, J.; He, Y.; Xiong, A.; Yu, J.; Li, Y.; Zhang, Y.; Zhao, W.; Zhou, F.; Li, W.; et al. Single-cell profiling of tumor heterogeneity and the microenvironment in advanced non-small cell lung cancer. Nat. Commun. 2021, 12, 2540. [Google Scholar] [CrossRef]
- Yoshida, R.; Saigi, M.; Tani, T.; Springer, B.F.; Shibata, H.; Kitajima, S.; Mahadevan, N.R.; Campisi, M.; Kim, W.; Kobayashi, Y.; et al. MET-Induced CD73 Restrains STING-Mediated Immunogenicity of EGFR-Mutant Lung Cancer. Cancer Res. 2022, 82, 4079–4092. [Google Scholar] [CrossRef] [PubMed]
- Lupia, M.; Angiolini, F.; Bertalot, G.; Freddi, S.; Sachsenmeier, K.F.; Chisci, E.; Kutryb-Zajac, B.; Confalonieri, S.; Smolenski, R.T.; Giovannoni, R.; et al. CD73 Regulates Stemness and Epithelial-Mesenchymal Transition in Ovarian Cancer-Initiating Cells. Stem Cell Rep. 2018, 10, 1412–1425. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Gu, C.; Yao, X.; Guo, W.; Wang, H.; Lin, T.; Li, F.; Chen, D.; Wu, J.; Ye, G.; et al. CD73 promotes tumor metastasis by modulating RICS/RhoA signaling and EMT in gastric cancer. Cell Death Dis. 2020, 11, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yarnold, J.; Brotons, M.C. Pathogenetic mechanisms in radiation fibrosis. Radiother. Oncol. 2010, 97, 149–161. [Google Scholar] [CrossRef] [PubMed]
- de Leve, S.; Wirsdörfer, F.; Cappuccini, F.; Schütze, A.; Meyer, A.V.; Röck, K.; Thompson, L.F.; Fischer, J.W.; Stuschke, M.; Jendrossek, V. Loss of CD73 prevents accumulation of alternatively activated macrophages and the formation of prefibrotic macrophage clusters in irradiated lungs. FASEB J. 2017, 31, 2869–2880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Leve, S.; Wirsdörfer, F.; Jendrossek, V. Targeting the Immunomodulatory CD73/Adenosine System to Improve the Therapeutic Gain of Radiotherapy. Front. Immunol. 2019, 10, 698. [Google Scholar] [CrossRef] [Green Version]
- Wirsdörfer, F.; de Leve, S.; Cappuccini, F.; Eldh, T.; Meyer, A.V.; Gau, E.; Thompson, L.F.; Chen, N.Y.; Karmouty-Quintana, H.; Fischer, U.; et al. Extracellular Adenosine Production by ecto-5′-Nucleotidase (CD73) Enhances Radiation-Induced Lung Fibrosis. Cancer Res. 2016, 76, 3045–3056. [Google Scholar] [CrossRef] [Green Version]
- Ullah, I.; Subbarao, R.B.; Rho, G.J. Human mesenchymal stem cells—Current trends and future prospective. Biosci. Rep. 2015, 35, e00191. [Google Scholar] [CrossRef]
- Joolharzadeh, P.; St. Hilaire, C. CD73 (Cluster of Differentiation 73) and the Differences between Mice and Humans. Arter. Thromb. Vasc. Biol. 2019, 39, 339–348. [Google Scholar] [CrossRef] [Green Version]
- Sudo, H.; Tsuji, A.B.; Sugyo, A.; Kurosawa, G.; Kurosawa, Y.; Alexander, D.; Tsuda, H.; Saga, T.; Higashi, T. Radiolabeled Human Monoclonal Antibody 067-213 has the Potential for Noninvasive Quantification of CD73 Expression. Int. J. Mol. Sci. 2020, 21, 2304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmies, C.C.; Rolshoven, G.; Idris, R.M.; Schmies, C.C.; Rolshoven, G.; Idris, R.M.; Losenkova, K.; Renn, C.; Schäkel, L.; Al-Hroub, H.; et al. Fluorescent Probes for Ecto-5′-nucleotidase (CD73). ACS Med. Chem. Lett. 2020, 11, 2253–2260. [Google Scholar] [CrossRef] [PubMed]
- Antonioli, L.; Yegutkin, G.G.; Pacher, P.; Blandizzi, C.; Haskó, G. Anti-CD73 in cancer immunotherapy: Awakening new opportunities. Trends Cancer 2016, 2, 95–109. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Zhuang, Z.; Wu, Q.; Lin, S.; Zhao, N.; Zhang, Q.; Xie, L.; Yu, S. CD39/CD73/A2a Adenosine Metabolic Pathway: Targets for Moxibustion in Treating DSS-Induced Ulcerative Colitis. Am. J. Chin. Med. 2021, 49, 661–676. [Google Scholar] [CrossRef]
- Vieira, C.; Magalhães-Cardoso, M.T.; Ferreirinha, F.; Silva, I.; Dias, A.S.; Pelletier, J.; Sévigny, J.; Correia-de-Sá, P. Feed-forward inhibition of CD73 and upregulation of adenosine deaminase contribute to the loss of adenosine neuromodulation in postinflammatory ileitis. Mediat. Inflamm. 2014, 2014, 254640. [Google Scholar] [CrossRef] [Green Version]
- Meng, F.; Guo, Z.; Hu, Y.; Mai, W.; Zhang, Z.; Zhang, B.; Ge, Q.; Lou, H.; Guo, F.; Chen, J.; et al. CD73-derived adenosine controls inflammation and neurodegeneration by modulating dopamine signalling. Brain 2019, 142, 700–718. [Google Scholar] [CrossRef] [PubMed]
- Moor, A.E.; Harnik, Y.; Ben-Moshe, S.; Massasa, E.E.; Rozenberg, M.; Eilam, R.; Bahar Halpern, K.; Itzkovitz, S. Spatial Reconstruction of Single Enterocytes Uncovers Broad Zonation along the Intestinal Villus Axis. Cell 2018, 175, 1156–1167.e15. [Google Scholar] [CrossRef] [Green Version]
- Piovesan, D.; Tan, J.B.L.; Becker, A.; Banuelos, J.; Narasappa, N.; DiRenzo, D.; Zhang, K.; Chen, A.; Ginn, E.; Udyavar, A.R.; et al. Targeting CD73 with AB680 (Quemliclustat), a Novel and Potent Small-Molecule CD73 Inhibitor, Restores Immune Functionality and Facilitates Antitumor Immunity. Mol. Cancer Ther. 2022, 21, 948–959. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Type of Lung Injury | Classification | CD73 | Reference | ||
|---|---|---|---|---|---|
| Expression | Relevant Mechanism | Positive (P) or Negative (N) | |||
| Cancer | NSCLC | ↑ | LDH5 and HIF1α↑, PD-L1 and LDHA↑ | N | [76] |
| ↓ | IL-10↓, CXCR4↓ | P | [77] | ||
| ↓ | CAF/T cell interactions | P | [78] | ||
| EGFR-mutated NSCLC | ↓ | CD8+ T cells↑, IFN-γ, and TNF-α↑ | P | [30] | |
| ↑ | PD-L1↑ | N | [79] | ||
| LUAD | ↑ | PD-L1↑ | N | [80] | |
| Marker | - | - | [81] | ||
| SCLC | ↓ | Elimination of metastatic chemoresistant SCLC | P | [82] | |
| Marker | - | - | [83] | ||
| NK cell | ↑ | LAG-3, VISTA, PD-1, and PD-L1↑,CD4-positive T cell, and IFN-γ↓ | N | [84] | |
| ↓ | TME↓ | P | [85] | ||
| I–III LUSC | Marker | - | - | [86] | |
| Pulmonary metastases | BC | ↓ | ADO-activated intracellular A2A receptor signaling pathway is linked to the AKT-β catenin pathway to regulate BC cell invasiveness and metastasis to the lung | [87] | |
| Triple-negative BC | ↓ | LC3I/LC3II ratio and p62↑ | P | [88] | |
| ↓ | TNBC cell migration in both normoxia and hypoxia↓ | P | [89] | ||
| LuM-1 | ↓ | IFN-γ and cytotoxicity against LuM-1↑ | P | [90] | |
| Tumor-bearing mice | ↓ | A2B↓ | P | [91] | |
| LUAD | ↑ | Epithelial to mesenchymal transition (EMT) progression↑ | N | [92] | |
| Bacterial infection | MBCs | Marker | Coexpression of at least 2 of these 3 memory markers distinguishes MBCs likely to differentiate into antibody-secreting cells (ASCs) upon reactivation | - | [93] |
| PMNs | ↓ | IL-10↑ | N | [56] | |
| ARDS | EVs with MSC origin | Marker | - | - | [94] |
| Lung organ cultures | ↓ | IFN beta-1a signaling↓ | N | [52] | |
| Hypoxic stress | NCI-H292 | ↓ | apoptosis↑, sensitivity to mitomycin↓, sensitivity to vincristine↑ | P | [95] |
| Allergic airway inflammation | OVA | ↓ | IL-4 and IL-5↑, TGF-β↓, CD4+CD25+Foxp3+ T cells↓ | p | [61] |
| OVA | ↓ | CD23+ B cells and IL4+ T cells↓, mast cells and degranulation↓ | P | [96] | |
| Long-term cigarette smoke | Cigarette smoke | ↓ | Inflammatory cells↓, IL-6↓ | P | [19] |
| COPD | COPD | Marker | - | - | [97] |
| HIV | DN T-cells | Marker | - | - | [98] |
| Thoracic endometriosis | TH-EM1 cell | Marker | - | - | [99] |
| Tuberculosis | MP287/03-infected mice | Marker | - | - | [100] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, X.-M.; Shi, N.-R.; Zhang, J.-Z.; Zuo, Y.-Q.; Wang, X.; Zhao, Y.-F.; Wu, J.-S. CD73: Friend or Foe in Lung Injury. Int. J. Mol. Sci. 2023, 24, 5545. https://doi.org/10.3390/ijms24065545
Hu X-M, Shi N-R, Zhang J-Z, Zuo Y-Q, Wang X, Zhao Y-F, Wu J-S. CD73: Friend or Foe in Lung Injury. International Journal of Molecular Sciences. 2023; 24(6):5545. https://doi.org/10.3390/ijms24065545
Chicago/Turabian StyleHu, Xiu-Min, Nan-Rui Shi, Ji-Zhou Zhang, Yan-Qin Zuo, Xin Wang, Ya-Fei Zhao, and Jia-Si Wu. 2023. "CD73: Friend or Foe in Lung Injury" International Journal of Molecular Sciences 24, no. 6: 5545. https://doi.org/10.3390/ijms24065545