
Generation of a Triple-Shuttling Vector and the Application in Plant Plus-Strand RNA Virus Infectious cDNA Clone Construction


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
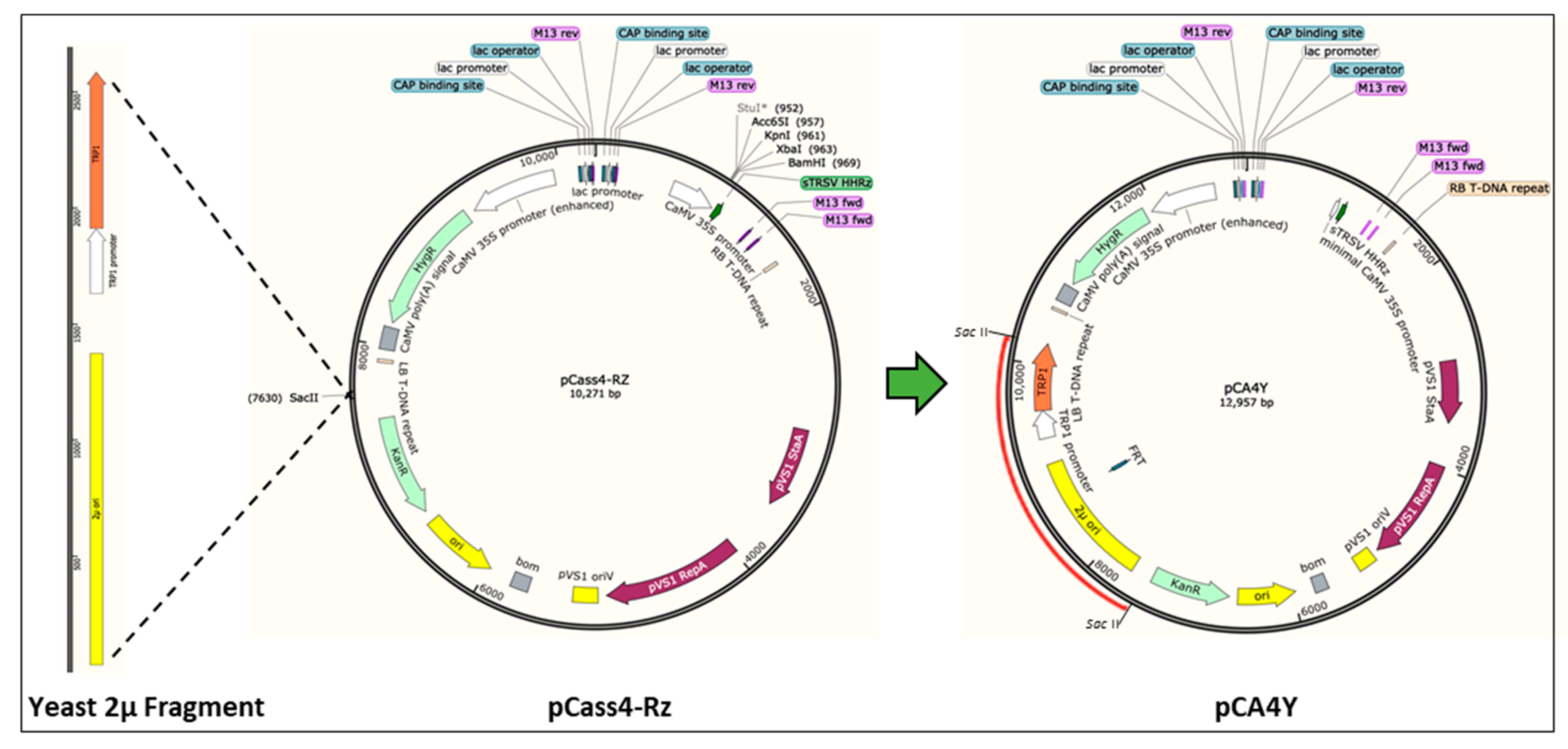
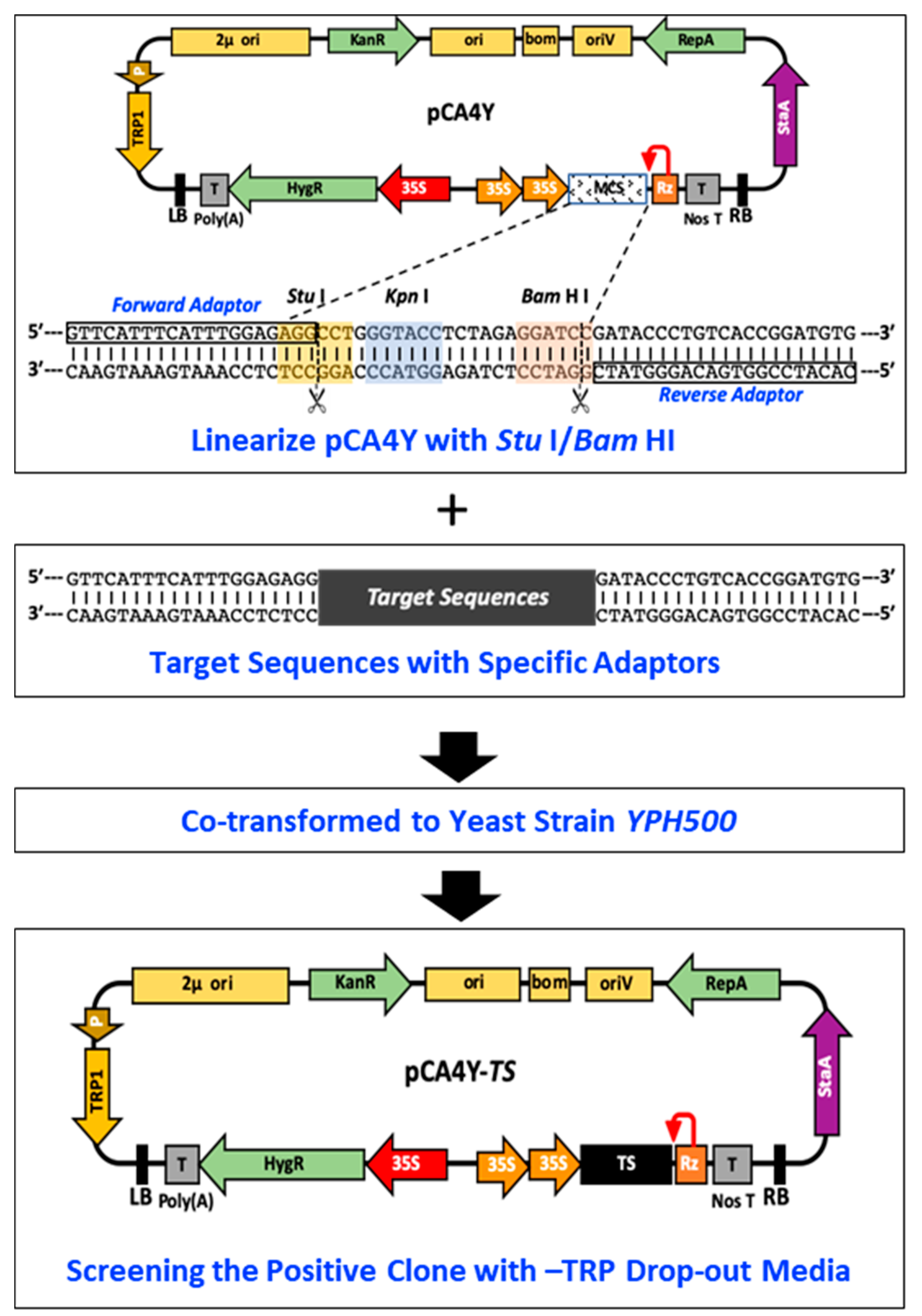
2.1. Construction of the S. cerevisiae–E. coli–Agrobacterium Shuttle Vectors and Development of HR-Based Cloning in Yeast
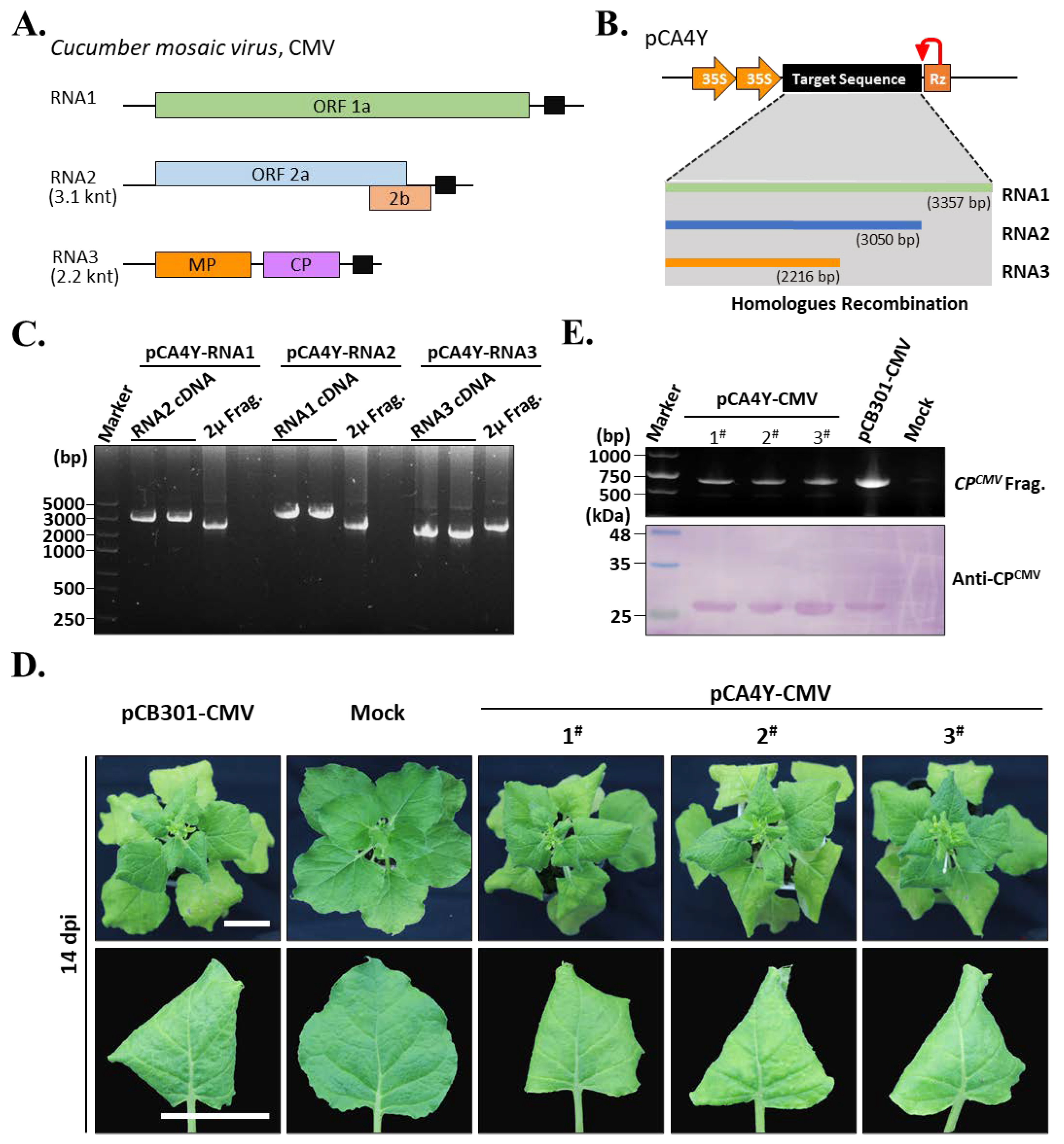
2.2. Construction of the CMV-Fny cDNA Infectious Clones Using the Triple-Shuttle Vector through Yeast Homologous Recombination
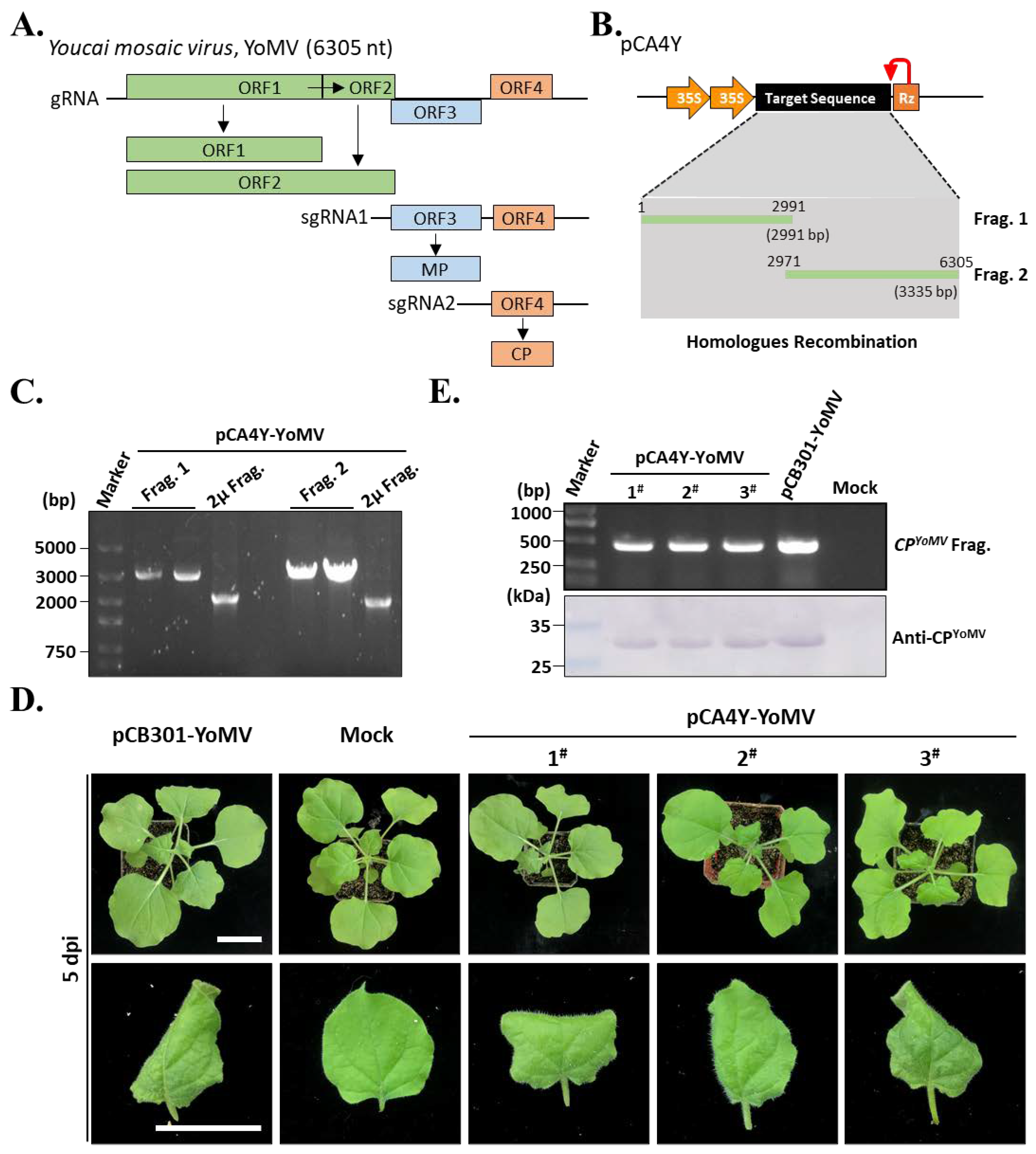
2.3. Construction of the YoMV Full-Length cDNA Infectious Clones
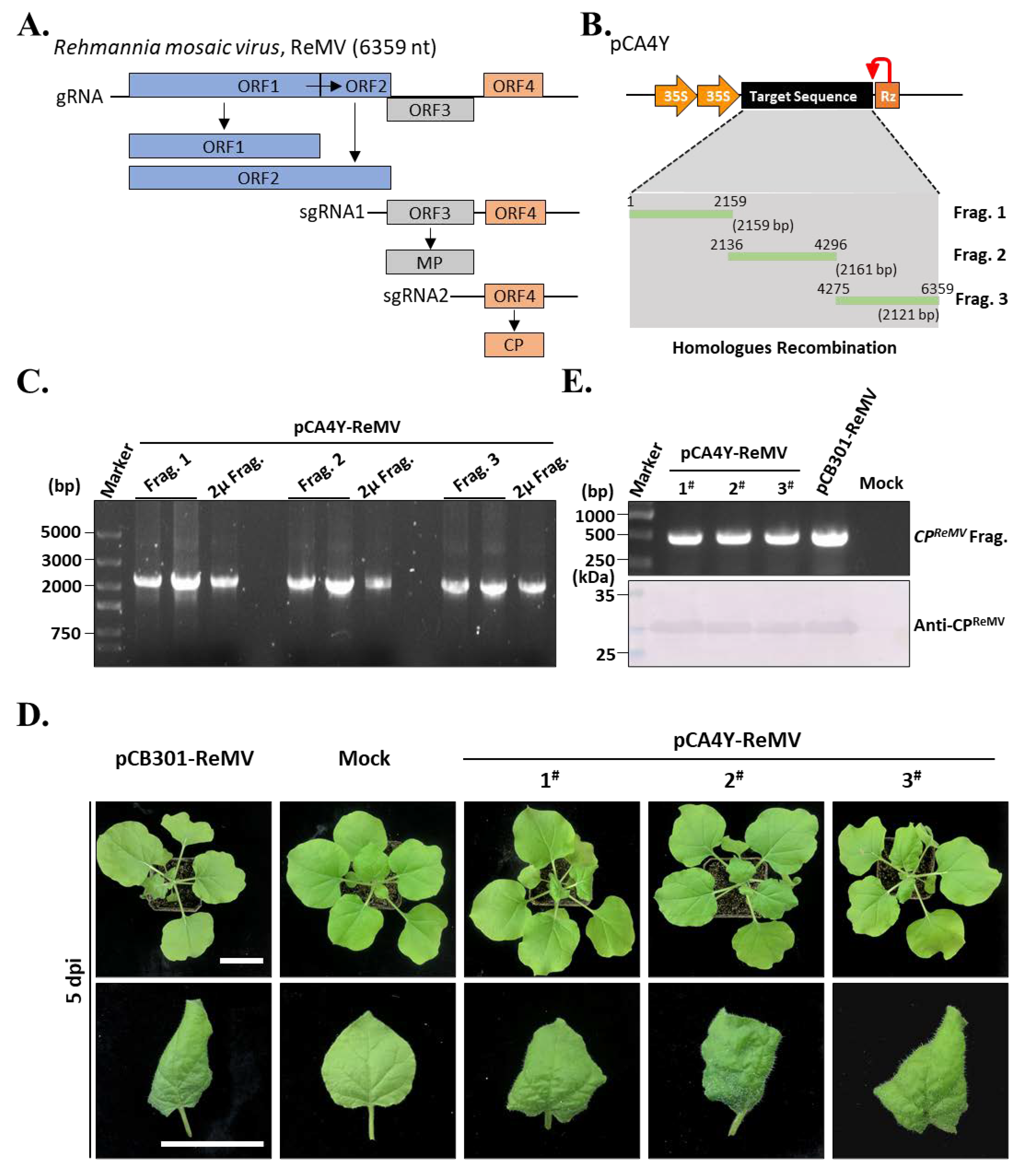
2.4. Construction of the ReMV Full-Length cDNA Infectious Clones
3. Discussion
4. Method and Materials
4.1. Vector Information and Plant Culture Condition
4.2. Candidate Virus and Viral RNA
4.3. Construction of the Triple-Shuttling Vector in S. cerevisiae, E. coli, and Agrobacterium
4.4. Assembly of CMV-Fny, YoMV, and ReMV cDNA Clones by Yeast Homologous Recombination
4.5. Plasmid Purification from Yeast and Propagation
4.6. Agrobacterium-Mediated Infiltration of N. benthamiana Leaves
4.7. RT-PCR and Western Blot Detection of the Target Virus
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boyer, J.C.; Haenni, A.L. Infectious Transcripts and cDNA Clones of RNA Viruses. Virology 1994, 198, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Ahlquist, P.; French, R.; Janda, M.; Loesch-Fries, L.S. Multicomponent RNA Plant Virus Infection Derived from Cloned Viral cDNA. Proc. Natl. Acad. Sci. USA 1984, 81, 7066–7070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, M.; Mise, K.; Kobayashi, K.; Okuno, T.; Furusawa, I. Infectivity of Plasmids Containing Brome Mosaic Virus cDNA Linked to the Cauliflower Mosaic Virus 35S RNA Promoter. J. Gen. Virol. 1991, 72, 243–246. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Zhao, D.; Liu, Y.; Huang, C.; Zhang, W.; Li, Z. Rapid Construction of Complex Plant RNA Virus Infectious cDNA Clones for Agroinfection Using a Yeast-E. Coli-Agrobacterium Shuttle Vector. Viruses 2017, 9, 332. [Google Scholar] [CrossRef] [Green Version]
- Jakab, G.; Droz, E.; Baulcombe, D.; Malnoe, P. Infectious in vivo and in vitro Transcripts from a Full-Length cDNA Clone of PVY-N605, a Swiss Necrotic Isolate of Potato Virus Y. J. Gen. Virol. 1997, 78, 3141–3145. [Google Scholar] [CrossRef]
- Johansen, I.E. Intron Insertion Facilitates Amplification of Cloned Virus cDNA in Escherichia coli While Biological Activity Is Reestablished after Transcription in vivo. Proc. Natl. Acad. Sci. USA 1996, 93, 12400–12405. [Google Scholar] [CrossRef] [Green Version]
- Tuo, D.; Shen, W.; Yan, P.; Li, X.; Zhou, P. Rapid Construction of Stable Infectious Full-Length cDNA Clone of Papaya Leaf Distortion Mosaic Virus Using In-Fusion Cloning. Viruses 2015, 7, 6241–6250. [Google Scholar] [CrossRef] [Green Version]
- Peng, Q.; Yang, D.; Yang, T.; Cheng, Y.; Yang, Y.; Xi, D. Construction of Full-length cDNA Infectious Clones of Chilli Veinal Mottle Virus. Virus Res. 2022, 322, 198948. [Google Scholar] [CrossRef]
- Bao, W.; Yan, T.; Deng, X.; Wuriyanghan, H. Synthesis of Full-Length cDNA Infectious Clones of Soybean Mosaic Virus and Functional Identification of a Key Amino Acid in the Silencing Suppressor Hc-Pro. Viruses 2020, 12, 886. [Google Scholar] [CrossRef]
- Wang, D.; Cui, L.; Zhang, L.; Ma, Z.; Niu, Y. Complete Genome Sequencing and Infectious cDNA Clone Construction of Soybean Mosaic Virus Isolated from Shanxi. J. Plant Pathol. 2021, 37, 162–172. [Google Scholar] [CrossRef]
- Jacquemond, M. Cucumber Mosaic Virus. Adv. Virus Res. 2021, 84, 439–504. [Google Scholar]
- Peter, P.; Fernando, G.-A. Cucumoviruses. Adv. Virus Res. 2003, 62, 241–323. [Google Scholar]
- Peter, P.; Fernando, G.-A. Cucumber Mosaic Virus; The American Phytopathological Society Publications: St. Paul, MN, USA, 2019. [Google Scholar]
- Creager, A.N.; Scholthof, K.B.; Citovsky, V.; Scholthof, H.B. Tobacco Mosaic Virus. Pioneering Research for A Century. Plant Cell 1999, 11, 301–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Feng, C.; Guo, X.; Zhou, Y.; Gu, T.; Zhuang, X.; Cheng, L.; Zhang, K. Development of Polyclonal Antibodies-Based Serological Methods for Detection of the Rehmannia Mosaic Virus in Field Plants. Front. Sustain. Food Syst. 2022, 6, 1013470. [Google Scholar] [CrossRef]
- Li, Z.; Tang, Y.; She, X.; Yu, L.; Lan, G.; He, Z. Molecular Characteristics and Pathogenicity Analysis of Youcai Mosaic Virus Guangdong Isolate Infecting Radish. Sci. Agric. Sin. 2022, 55, 2752–2761. [Google Scholar]
- Kenyon, L.; Kumar, S.; Tsai, W.-S.; Hughes, J.D. Virus Diseases of Peppers (Capsicum spp.) and Their Control. Adv. Virus Res. 2014, 90, 297–354, ISBN 978-0-12-801246-8. [Google Scholar]
- Yao, M.; Zhang, T.Q.; Tian, Z.C.; Wang, Y.C.; Tao, X.R. Construction of Agrobacterium-mediated Cucumber Mosaic Virus Infectious cDNA Clones and 2b Deletion Viral Vector. Sci. Agric. Sin. 2011, 44, 3060–3068. [Google Scholar]
- Zhang, K.; Zhuang, X.; Guo, X.; Xu, H.; He, Z.; Chen, J. Cucurbit Chlorotic Yellows Virus Infecting Rehmannia glutinosa Was Detected in China. Plant Dis. 2021, 105, 3310. [Google Scholar] [CrossRef]
- Chapman, S.N. Construction of Infectious Clones for RNA Viruses: TMV. Methods Mol. Biol. 2008, 451, 477–490. [Google Scholar]
- Blawid, R.; Nagata, T. Construction of an Infectious Clone of a Plant RNA Virus in a Binary Vector Using One-Step Gibson Assembly. J. Virol. Methods 2015, 222, 11–15. [Google Scholar] [CrossRef]
- Bordat, A.; Houvenaghel, M.-C.; German-Retana, S. Gibson Assembly: An Easy Way to Clone Potyviral Full-Length Infectious cDNA Clones Expressing an Ectopic VPg. Virol. J. 2015, 12, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuo, D.; Fu, L.; Shen, W.; Li, X.; Zhou, P.; Yan, P. Generation of Stable Infectious Clones of Plant Viruses by Using Rhizobium Radiobacter for Both Cloning and Inoculation. Virology 2017, 510, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Desbiez, C.; Chandeysson, C.; Lecoq, H.; Moury, B. A Simple, Rapid and Efficient Way to Obtain Infectious Clones of Potyviruses. J. Virol. Methods 2012, 183, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Delmond, I.; Pierrugues, O.; de Wispelaere, M.; Guilbaud, L.; Gaubert, S.; Divéki, Z.; Godon, C.; Tepfer, M.; Jacquemond, M. A Novel Strategy for Creating Recombinant Infectious RNA Virus Genomes. J. Virol. Methods 2004, 121, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Polo, S.; Ketner, G.; Levis, R.; Falgout, B. Infectious RNA Transcripts from Full-Length Dengue Virus Type 2 cDNA Clones Made in Yeast. J. Virol. 1997, 71, 5366–5374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youssef, F.; Marais, A.; Faure, C.; Gentit, P.; Candresse, T. Strategies to Facilitate the Development of Uncloned or Cloned Infectious Full-Length Viral cDNAs: Apple Chlorotic Leaf Spot Virus as a Case Study. Virol. J. 2011, 8, 488. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Liu, T.; Zhang, Z.; Chen, M.; Rong, L.; Ma, L.; Yu, B.; Wu, D.; Zhang, P.; Zhu, X.; et al. Novel Genetically Stable Infectious Clone for a Zika Virus Clinical Isolate and Identification of RNA Elements Essential for Virus Production. Virus Res. 2018, 257, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-Y.; Yu, J.-Y.; Huang, X.-Y.; Fan, H.; Li, X.-F.; Deng, Y.-Q.; Ji, X.; Cheng, M.-L.; Ye, Q.; Zhao, H.; et al. Characterization of Cis -Acting RNA Elements of Zika Virus by Using a Self-Splicing Ribozyme-Dependent Infectious Clone. J Virol. 2017, 91, e00484-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.J.; Revers, F.; Souche, S.; Lot, H.; Le Gall, O.; Candresse, T.; Dunez, J. Construction of Full-Length cDNA Clones of Lettuce Mosaic Virus (LMV) and the Effects of Intron-Insertion on Their Viability in Escherichia coli and on Their Infectivity to Plants. Arch. Virol. 1998, 143, 2443–2451. [Google Scholar] [CrossRef]
- Lai, C.J.; Zhao, B.T.; Hori, H.; Bray, M. Infectious RNA Transcribed from Stably Cloned Full-Length cDNA of Dengue Type 4 Virus. Proc. Natl. Acad. Sci. USA 1991, 88, 5139–5143. [Google Scholar] [CrossRef] [Green Version]
- Quillet, L.; Guilley, H.; Jonard, G.; Richards, K. In Vitro Synthesis of Biologically Active Beet Necrotic Yellow Vein Virus RNA. Virology 1989, 172, 293–301. [Google Scholar] [CrossRef] [PubMed]
- MacFarlane, S.A.; Wallis, C.V.; Taylor, S.C.; Goulden, M.G.; Wood, K.R.; Davies, J.W. Construction and Analysis of Infectious Transcripts Synthesized from Full-Length cDNA Clones of Both Genomic RNAs of Pea Early Browning Virus. Virology 1991, 182, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Skotnicki, M.L.; Ding, S.-W.; Mackenzie, A.M.; Gibbs, A.J. Infectious Eggplant Mosaic Tymovirus and Ononis Yellow Mosaic Tymovirus from Cloned cDNA. Arch. Virol. 1993, 131, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Tian, Y.-P.; Wang, J.; Yin, X.; Li, X.-D.; Valkonen, J.P.T. Construction of an Infectious cDNA Clone and Gene Expression Vector of Tobacco Vein Banding Mosaic Virus (Genus potyvirus). Virus Res. 2012, 169, 276–281. [Google Scholar] [CrossRef]
- López-Moya, J.J.; García, J.A. Construction of a Stable and Highly Infectious Intron-Containing cDNA Clone of Plum Pox Potyvirus and Its Use to Infect Plants by Particle Bombardment. Virus Res. 2000, 68, 99–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moury, B.; Morel, C.; Johansen, E.; Guilbaud, L.; Souche, S.; Ayme, V.; Caranta, C.; Palloix, A.; Jacquemond, M. Mutations in Potato Virus Y Genome-Linked Protein Determine Virulence Toward Recessive Resistances in Capsicum annuum and Lycopersicon hirsutum. Mol. Plant Microbe Interact. 2004, 17, 322–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, B.S.; Johansen, I.E. Nucleotide Sequence and Infectious cDNA Clone of the L1 Isolate of Pea Seed-Borne Mosaic Potyvirus. Arch. Virol. 2001, 146, 15–25. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, J.; Liu, H.; Wang, M.; Dong, Y.; Zhou, Y.; Wong, S.-M.; Xu, K.; Xu, Q. A Plant Virus Hijacks Phosphatidylinositol-3,5-Bisphosphate to Escape Autophagic Degradation in Its Insect Vector. Autophagy 2022, 1–16. [Google Scholar] [CrossRef]
- Xu, Y.; Fu, S.; Tao, X.; Zhou, X. Rice Stripe Virus: Exploring Molecular Weapons in the Arsenal of A Negative-sense RNA virus. Annu. Rev. Phytopathol. 2021, 59, 351–371. [Google Scholar] [CrossRef]
- Matsumoto, M.; Shoyama, Y.; Nishioka, I.; Iwai, H.; Wakimoto, S. Identification of Viruses Infected in Rehmannia Glutinosa Libosch. Var Purpurea Makino and Effect of Virus Infection on Root Yield and Iridoid Glycoside Contents. Plant Cell Rep. 1989, 7, 636–638. [Google Scholar] [CrossRef]
- Qin, Y.H.; Wang, F.; Lu, C.T.; Wang, F.L.; Wen, Y.; Liu, Y.X.; Gao, S.X.; Qi, W.P.; Li, X.M.; Yang, J. First Report of Tobacco Mild Green Mosaic Virus Infecting Rehmannia Glutinosa in China. Plant Dis. 2022, 106, 3004. [Google Scholar] [CrossRef] [PubMed]
- Hofgen, R.; Willmitzer, L. Storage of Competent Cells for Agrobacterium Transformation. Nucleic Acids Res. 1988, 16, 9877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Zhang, Y.; Yang, M.; Liu, S.; Li, Z.; Wang, X.; Han, C.; Yu, J.; Li, D. The Barley Stripe Mosaic Virus Gammab Protein Promotes Chloroplast-targeted Replication by Enhancing Unwinding of RNA Duplexes. PLoS Pathog. 2017, 13, e1006319. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, C.; Guo, X.; Gu, T.; Hua, Y.; Zhuang, X.; Zhang, K. Generation of a Triple-Shuttling Vector and the Application in Plant Plus-Strand RNA Virus Infectious cDNA Clone Construction. Int. J. Mol. Sci. 2023, 24, 5477. https://doi.org/10.3390/ijms24065477
Feng C, Guo X, Gu T, Hua Y, Zhuang X, Zhang K. Generation of a Triple-Shuttling Vector and the Application in Plant Plus-Strand RNA Virus Infectious cDNA Clone Construction. International Journal of Molecular Sciences. 2023; 24(6):5477. https://doi.org/10.3390/ijms24065477
Chicago/Turabian StyleFeng, Chenwei, Xiao Guo, Tianxiao Gu, Yanhong Hua, Xinjian Zhuang, and Kun Zhang. 2023. "Generation of a Triple-Shuttling Vector and the Application in Plant Plus-Strand RNA Virus Infectious cDNA Clone Construction" International Journal of Molecular Sciences 24, no. 6: 5477. https://doi.org/10.3390/ijms24065477