

Haslea ostrearia Pigment Marennine Affects Key Actors of Neuroinflammation and Decreases Cell Migration in Murine Neuroglial Cell Model

 , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Marennine Is Not Cytotoxic at 10 µg/mL and 50 µg/mL
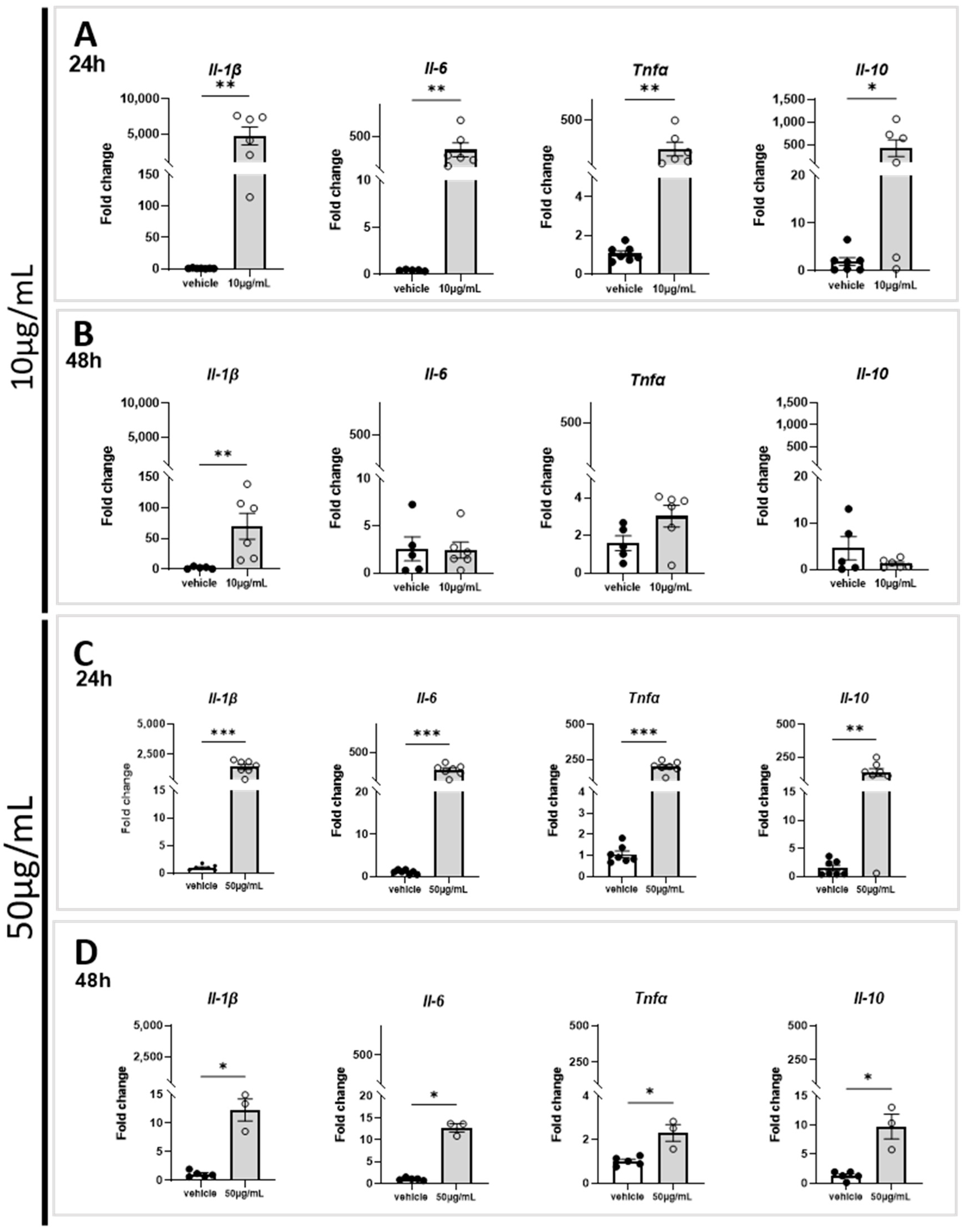
2.2. Marennine Induces an Upregulation of Immunomodulatory Genes
2.3. Marennine Affects the Population of Immunocompetent Cells of the CNS
2.3.1. Marennine Modifies the Inflammatory Status without Affecting the Total Amount of Astrocyte Cells
2.3.2. Marennine Modifies the Inflammatory Status and Modulates the Total Amount of Microglial Cells
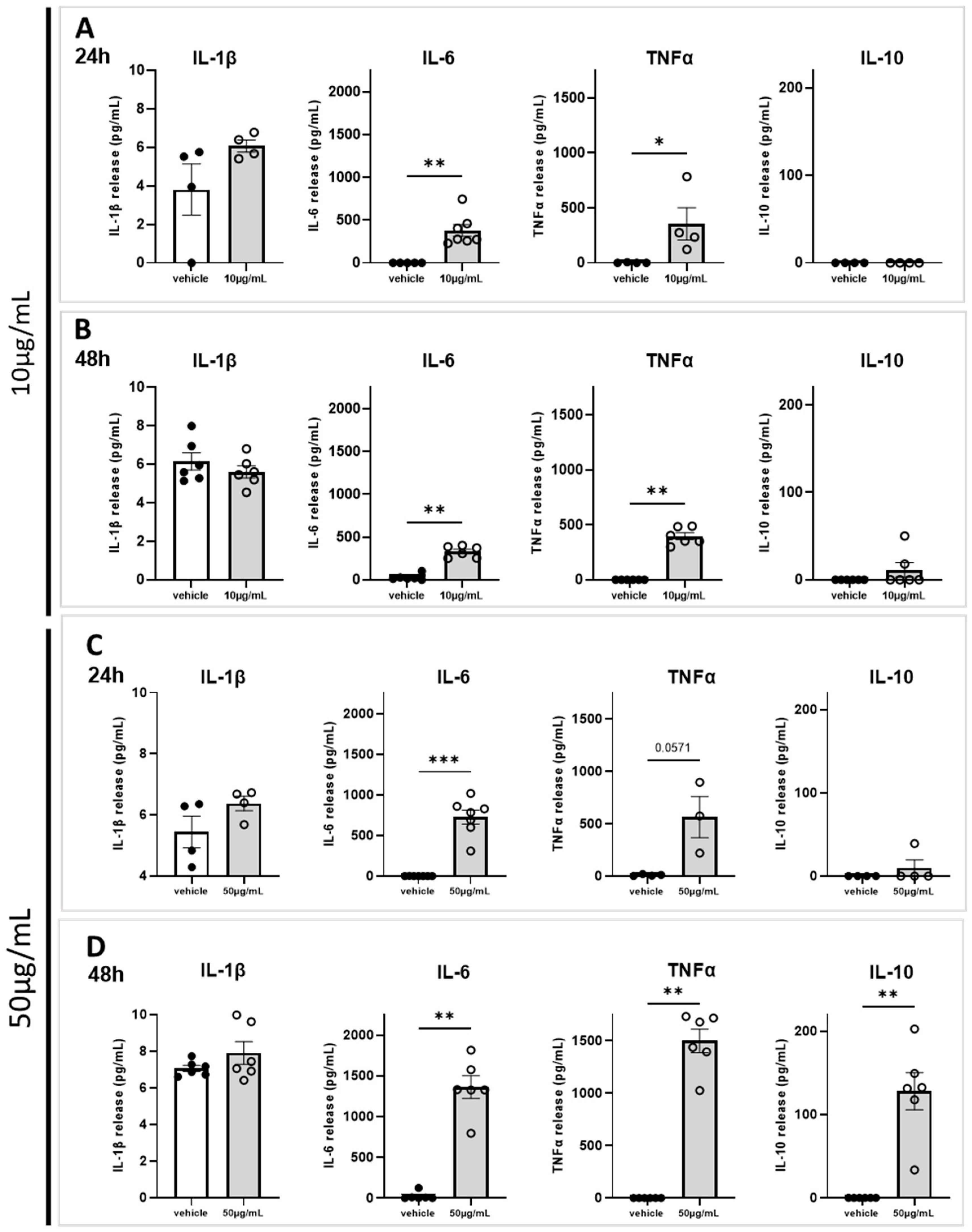
2.4. Marennine Induces a Release of Key Actors of Neuroinflammation
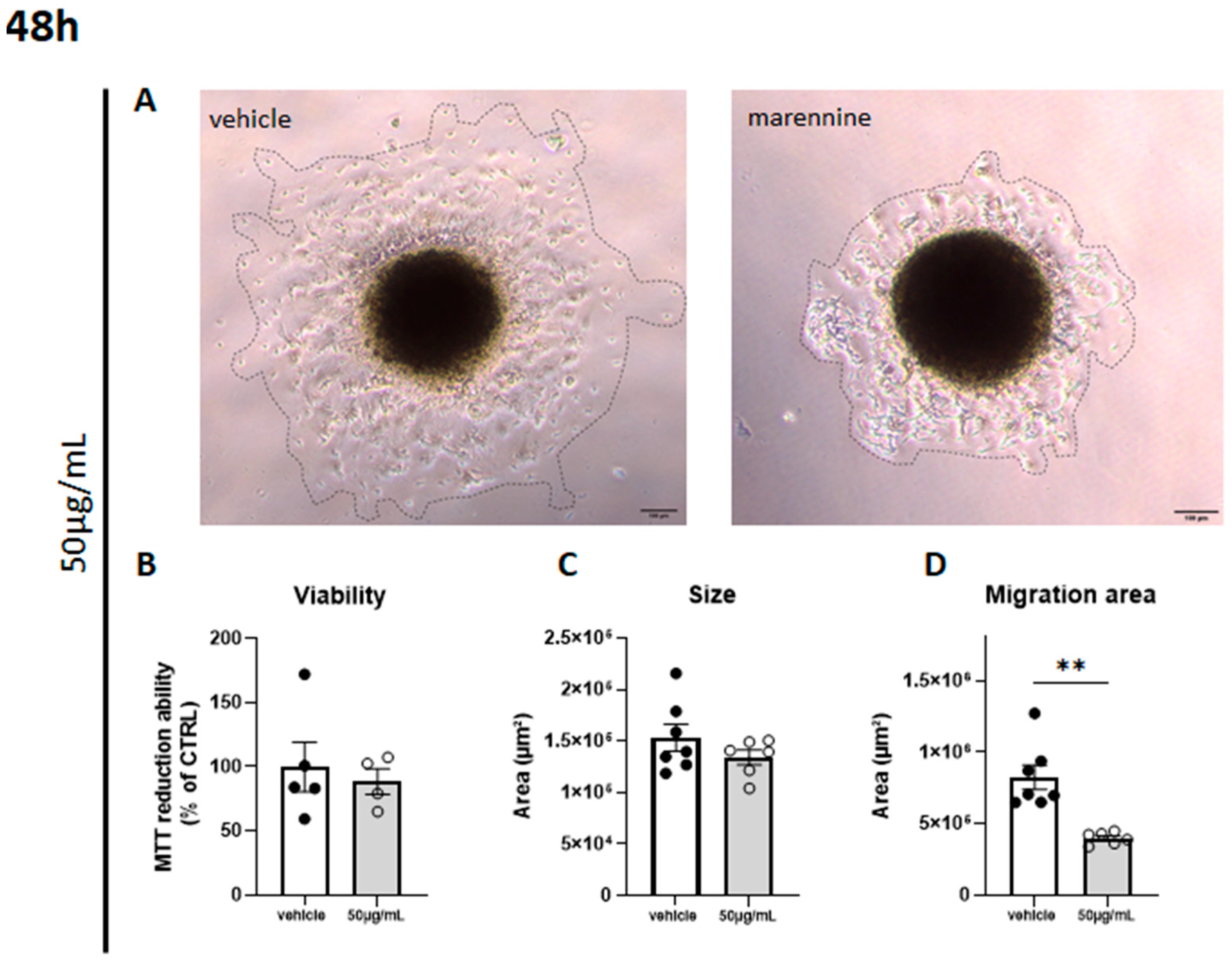
2.5. Marennine Reduces the Migration Surface of Cells from Neurospheres without Inducing Cytotoxicity
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cultivation of Microalgae and Marennine Extraction
4.3. Neuroglial Cell Culture and Marennine Exposure
4.4. Neuroglial Cell Viability Analysis
4.5. Gene Expression Analysis
4.6. Neuroglial Cell Populations and Activation Status Analysis
4.7. Measurement of Cytokine Release
4.8. Neurospheres Migration Assay
4.9. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chapman, R.L. Algae: The World’s Most Important “Plants”—An Introduction. Mitig. Adapt. Strateg. Glob. Chang. 2013, 18, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Barkia, I.; Saari, N.; Manning, S.R. Microalgae for High-Value Products Towards Human Health and Nutrition. Mar. Drugs 2019, 17, 304. [Google Scholar] [CrossRef] [Green Version]
- Qiao, Y.; Yang, F.; Xie, T.; Du, Z.; Zhong, D.; Qi, Y.; Li, Y.; Li, W.; Lu, Z.; Rao, J.; et al. Engineered Algae: A Novel Oxygen-Generating System for Effective Treatment of Hypoxic Cancer. Sci. Adv. 2020, 6, eaba5996. [Google Scholar] [CrossRef]
- Li, G.; Chang, X.; Luo, X.; Zhao, Y.; Wang, W.; Kang, X. Fucoxanthin Induces Prostate Cancer PC-3 Cell Apoptosis by Causing Mitochondria Dysfunction and Oxidative Stress. Nan Fang Yi Ke Da Xue Xue Bao 2021, 41, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Nurcahyanti, A.D.R.; Kusmita, L.; Wink, M. Bixin and Fucoxanthin Sensitize Human Lung Cancer and Cervical Cancer Cell to Cisplatin in Vitro. BMC Res. Notes 2021, 14, 454. [Google Scholar] [CrossRef]
- Zhang, D.; Zhong, D.; Ouyang, J.; He, J.; Qi, Y.; Chen, W.; Zhang, X.; Tao, W.; Zhou, M. Microalgae-Based Oral Microcarriers for Gut Microbiota Homeostasis and Intestinal Protection in Cancer Radiotherapy. Nat. Commun. 2022, 13, 1413. [Google Scholar] [CrossRef]
- Benemann, J.R.; Woertz, I.; Lundquist, T. Autotrophic Microalgae Biomass Production: From Niche Markets to Commodities. Ind. Biotechnol. 2018, 14, 3–10. [Google Scholar] [CrossRef]
- Dolganyuk, V.; Belova, D.; Babich, O.; Prosekov, A.; Ivanova, S.; Katserov, D.; Patyukov, N.; Sukhikh, S. Microalgae: A Promising Source of Valuable Bioproducts. Biomolecules 2020, 10, 1153. [Google Scholar] [CrossRef] [PubMed]
- Coulombier, N.; Jauffrais, T.; Lebouvier, N. Antioxidant Compounds from Microalgae: A Review. Mar. Drugs 2021, 19, 549. [Google Scholar] [CrossRef] [PubMed]
- Sorrenti, V.; Castagna, D.A.; Fortinguerra, S.; Buriani, A.; Scapagnini, G.; Willcox, D.C. Spirulina Microalgae and Brain Health: A Scoping Review of Experimental and Clinical Evidence. Mar. Drugs 2021, 19, 293. [Google Scholar] [CrossRef] [PubMed]
- Mohibbullah, M.; Haque, M.N.; Sohag, A.A.M.; Hossain, M.T.; Zahan, M.S.; Uddin, M.J.; Hannan, M.A.; Moon, I.S.; Choi, J.-S. A Systematic Review on Marine Algae-Derived Fucoxanthin: An Update of Pharmacological Insights. Mar. Drugs 2022, 20, 279. [Google Scholar] [CrossRef]
- Morabito, S.; Silvestro, S.; Faggio, C. How the Marine Biotoxins Affect Human Health. Nat. Prod. Res. 2018, 32, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-Y.; Rasmussen, U.; Rydberg, S. Effect and Function of β-N-Methylamino-L-Alanine in the Diatom Phaeodactylum Tricornutum. Sci. Total Environ. 2022, 830, 154778. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.; Rodríguez, A.; Taglialatela-Scafati, O.; Fusetani, N. Marine Pharmacology in 2009–2011: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis, and Antiviral Activities; Affecting the Immune and Nervous Systems, and Other Miscellaneous Mechanisms of Action. Mar. Drugs 2013, 11, 2510–2573. [Google Scholar] [CrossRef]
- Hannan, M.A.; Dash, R.; Haque, M.N.; Mohibbullah, M.; Sohag, A.A.M.; Rahman, M.A.; Uddin, M.J.; Alam, M.; Moon, I.S. Neuroprotective Potentials of Marine Algae and Their Bioactive Metabolites: Pharmacological Insights and Therapeutic Advances. Mar. Drugs 2020, 18, 347. [Google Scholar] [CrossRef]
- Piovan, A.; Filippini, R.; Corbioli, G.; Costa, V.D.; Giunco, E.M.V.; Burbello, G.; Pagetta, A.; Giusti, P.; Zusso, M. Carotenoid Extract Derived from Euglena Gracilis Overcomes Lipopolysaccharide-Induced Neuroinflammation in Microglia: Role of NF-ΚB and Nrf2 Signaling Pathways. Mol. Neurobiol. 2021, 58, 3515–3528. [Google Scholar] [CrossRef]
- Lee, A.-H.; Shin, H.-Y.; Park, J.-H.; Koo, S.Y.; Kim, S.M.; Yang, S.-H. Fucoxanthin from Microalgae Phaeodactylum Tricornutum Inhibits Pro-Inflammatory Cytokines by Regulating Both NF-ΚB and NLRP3 Inflammasome Activation. Sci. Rep. 2021, 11, 543. [Google Scholar] [CrossRef] [PubMed]
- Zebiri, I.; Jacquette, B.; Francezon, N.; Herbaut, M.; Latigui, A.; Bricaud, S.; Tremblay, R.; Pasetto, P.; Mouget, J.-L.; Dittmer, J. The Polysaccharidic Nature of the Skeleton of Marennine as Determined by NMR Spectroscopy. Mar. Drugs 2023, 21, 42. [Google Scholar] [CrossRef]
- Lankester, E.R. Green Oysters. Nature 1895, 52, 28–29. [Google Scholar] [CrossRef] [Green Version]
- Falaise, C.; Cormier, P.; Tremblay, R.; Audet, C.; Deschênes, J.-S.; Turcotte, F.; François, C.; Seger, A.; Hallegraeff, G.; Lindquist, N.; et al. Harmful or Harmless: Biological Effects of Marennine on Marine Organisms. Aquat. Toxicol. 2019, 209, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Gastineau, R.; Turcotte, F.; Pouvreau, J.-B.; Morançais, M.; Fleurence, J.; Windarto, E.; Prasetiya, F.; Arsad, S.; Jaouen, P.; Babin, M.; et al. Marennine, Promising Blue Pigments from a Widespread Haslea Diatom Species Complex. Mar. Drugs 2014, 12, 3161–3189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schubert, H.; Tremblin, G.; Robert, J.-M.; Sagert, S.; Rincé, Y. In-Vivo Fluorescence Measurement of Photosynthesis of Haslea ostrearia Simonsen in Relation to Marennine Content. Diatom Res. 1995, 10, 341–349. [Google Scholar] [CrossRef]
- Pouvreau, J.-B.; Morançais, M.; Fleury, F.; Rosa, P.; Thion, L.; Cahingt, B.; Zal, F.; Fleurence, J.; Pondaven, P. Preliminary Characterisation of the Blue-Green Pigment “Marennine” from the Marine Tychopelagic Diatom Haslea Ostrearia (Gaillon/Bory) Simonsen. J. Appl. Phycol. 2006, 18, 757–767. [Google Scholar] [CrossRef]
- Pouvreau, J.-B.; Taran, F.; Rosa, P.; Pin, S.; Fleurence, J.; Pondaven, P. Antioxidant and Free Radical Scavenging Properties of Marennine, a Blue-Green Polyphenolic Pigment from the Diatom Haslea ostrearia (Gaillon/Bory) Simonsen Responsible for the Natural Greening of Cultured Oysters. J. Agric. Food Chem 2008, 56, 6278–6286. [Google Scholar] [CrossRef]
- Prasetiya, F.S.; Safitri, I.; Widowati, I.; Cognie, B.; Decottignies, P.; Gastineau, R.; Morançais, M.; Windarto, E.; Tremblay, R.; Mouget, J.-L. Does Allelopathy Affect Co-Culturing Haslea Ostrearia with Other Microalgae Relevant to Aquaculture? J. Appl. Phycol. 2016, 28, 2241–2254. [Google Scholar] [CrossRef]
- Bergé, J.-P.; Bourgougnon, N.; Alban, S.; Pojer, F.; Billaudel, S.; Chermann, J.-C.; Robert, J.M.; Franz, G. Antiviral and Anticoagulant Activities of a Water-Soluble Fraction of the Marine Diatom Haslea Ostrearia. Planta Med. 1999, 65, 604–609. [Google Scholar] [CrossRef]
- Gastineau, R.; Pouvreau, J.-B.; Hellio, C.; Morançais, M.; Fleurence, J.; Gaudin, P.; Bourgougnon, N.; Mouget, J.-L. Biological Activities of Purified Marennine, the Blue Pigment Responsible for the Greening of Oysters. J. Agric. Food Chem. 2012, 60, 3599–3605. [Google Scholar] [CrossRef] [PubMed]
- Carbonnelle, D.; Pondaven, P.; Morancais, M.; Masse, G.; Bosch, S.; Jacquot, C.; Briand, G.; Robert, J.; Roussakis, C. Antitumor and Antiproliferative Effects of an Aqueous Extract from the Marine Diatom Haslea Ostrearia (Simonsen) against Solid Tumors: Lung Carcinoma (NSCLC-N6), Kidney Carcinoma (E39) and Melanoma (M96) Cell Lines. Anticancer Res. 1999, 19, 621–624. [Google Scholar]
- Habanjar, O.; Diab-Assaf, M.; Caldefie-Chezet, F.; Delort, L. 3D Cell Culture Systems: Tumor Application, Advantages, and Disadvantages. Int. J. Mol. Sci. 2021, 22, 12200. [Google Scholar] [CrossRef]
- Masood, M.I.; Hauke, N.T.; Nasim, M.J.; Sarfraz, M.; Naseem, M.; Schäfer, K.H. Neural Stem Cell-Based in Vitro Bioassay for the Assessment of Neurotoxic Potential of Water Samples. J. Environ. Sci. 2021, 101, 72–86. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Zorec, R.; Parpura, V. Stratification of Astrocytes in Healthy and Diseased Brain. Brain Pathol. 2017, 27, 629–644. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Valin, K.L.; Dixon, M.L.; Leavenworth, J.W. The Role of Microglia and Macrophages in CNS Homeostasis, Autoimmunity, and Cancer. J. Immunol. Res. 2017, 2017, 5150678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogdan, C.; Vodovotz, Y.; Nathan, C. Macrophage Deactivation by Interleukin 10. J. Exp. Med. 1991, 174, 1549–1555. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, T.A.; Ohmori, Y.; Tebo, J.M.; Kishore, R. Regulation of Macrophage Gene Expression by Pro- and Anti-Inflammatory Cytokines. Pathobiology 1999, 67, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Locksley, R.M.; Killeen, N.; Lenardo, M.J. The TNF and TNF Receptor Superfamilies. Cell 2001, 104, 487–501. [Google Scholar] [CrossRef] [Green Version]
- Vieira, L.Q.; Goldschmidt, M.; Nashleanas, M.; Pfeffer, K.; Mak, T.; Scott, P. Mice Lacking the TNF Receptor P55 Fail to Resolve Lesions Caused by Infection with Leishmania Major, but Control Parasite Replication. J. Immunol. 1996, 157, 827–835. [Google Scholar] [CrossRef]
- Marino, M.W.; Dunn, A.; Grail, D.; Inglese, M.; Noguchi, Y.; Richards, E.; Jungbluth, A.; Wada, H.; Moore, M.; Williamson, B.; et al. Characterization of Tumor Necrosis Factor-Deficient Mice. Proc. Natl. Acad. Sci. USA 1997, 94, 8093–8098. [Google Scholar] [CrossRef] [Green Version]
- Welser-Alves, J.V.; Crocker, S.J.; Milner, R. A Dual Role for Microglia in Promoting Tissue Inhibitor of Metalloproteinase (TIMP) Expression in Glial Cells in Response to Neuroinflammatory Stimuli. J. Neuroinflamm. 2011, 8, 61. [Google Scholar] [CrossRef] [Green Version]
- Hobbs, S.; Reynoso, M.; Geddis, A.V.; Mitrophanov, A.Y.; Matheny, R.W. LPS-stimulated NF-κB P65 Dynamic Response Marks the Initiation of TNF Expression and Transition to IL-10 Expression in RAW 264.7 Macrophages. Physiol. Rep. 2018, 6, e13914. [Google Scholar] [CrossRef] [Green Version]
- Rhee, S.H. Lipopolysaccharide: Basic Biochemistry, Intracellular Signaling, and Physiological Impacts in the Gut. Intest. Res. 2014, 12, 90. [Google Scholar] [CrossRef] [Green Version]
- Lobo-Silva, D.; Carriche, G.M.; Castro, A.G.; Roque, S.; Saraiva, M. Balancing the Immune Response in the Brain: IL-10 and Its Regulation. J. Neuroinflamm. 2016, 13, 297. [Google Scholar] [CrossRef] [Green Version]
- Kessler, B.; Rinchai, D.; Kewcharoenwong, C.; Nithichanon, A.; Biggart, R.; Hawrylowicz, C.M.; Bancroft, G.J.; Lertmemongkolchai, G. Interleukin 10 Inhibits Pro-Inflammatory Cytokine Responses and Killing of Burkholderia Pseudomallei. Sci. Rep. 2017, 7, 42791. [Google Scholar] [CrossRef] [Green Version]
- dos Santos, S.E.; Medeiros, M.; Porfirio, J.; Tavares, W.; Pessôa, L.; Grinberg, L.; Leite, R.E.P.; Ferretti-Rebustini, R.E.L.; Suemoto, C.K.; Filho, W.J.; et al. Similar Microglial Cell Densities across Brain Structures and Mammalian Species: Implications for Brain Tissue Function. J. Neurosci. 2020, 40, 4622–4643. [Google Scholar] [CrossRef] [PubMed]
- Thornberry, N.A.; Bull, H.G.; Calaycay, J.R.; Chapman, K.T.; Howard, A.D.; Kostura, M.J.; Miller, D.K.; Molineaux, S.M.; Weidner, J.R.; Aunins, J.; et al. A Novel Heterodimeric Cysteine Protease Is Required for Interleukin-1βprocessing in Monocytes. Nature 1992, 356, 768–774. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Liu, J.; Bao, J.; Bai, Q.; Wang, G. Interaction of Microglia and Astrocytes in the Neurovascular Unit. Front. Immunol. 2020, 11, 1024. [Google Scholar] [CrossRef]
- Norden, D.M.; Trojanowski, P.J.; Walker, F.R.; Godbout, J.P. Insensitivity of Astrocytes to Interleukin 10 Signaling Following Peripheral Immune Challenge Results in Prolonged Microglial Activation in the Aged Brain. Neurobiol. Aging 2016, 44, 22–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shemer, A.; Scheyltjens, I.; Frumer, G.R.; Kim, J.-S.; Grozovski, J.; Ayanaw, S.; Dassa, B.; van Hove, H.; Chappell-Maor, L.; Boura-Halfon, S.; et al. Interleukin-10 Prevents Pathological Microglia Hyperactivation Following Peripheral Endotoxin Challenge. Immunity 2020, 53, 1033–1049.e7. [Google Scholar] [CrossRef]
- Qadri, M.; Jay, G.D.; Zhang, L.X.; Schmidt, T.A.; Totonchy, J.; Elsaid, K.A. Proteoglycan-4 Is an Essential Regulator of Synovial Macrophage Polarization and Inflammatory Macrophage Joint Infiltration. Arthritis Res. Ther. 2021, 23, 241. [Google Scholar] [CrossRef]
- Méresse, S.; Larrigaldie, V.; Oummadi, A.; de Concini, V.; Morisset-Lopez, S.; Reverchon, F.; Menuet, A.; Montécot-Dubourg, C.; Mortaud, S. β-N-Methyl-Amino-L-Alanine Cyanotoxin Promotes Modification of Undifferentiated Cells Population and Disrupts the Inflammatory Status in Primary Cultures of Neural Stem Cells. Toxicology 2022, 482, 153358. [Google Scholar] [CrossRef]
- Peng, B.; Xiao, J.; Wang, K.; So, K.-F.; Tipoe, G.L.; Lin, B. Suppression of Microglial Activation Is Neuroprotective in a Mouse Model of Human Retinitis Pigmentosa. J. Neurosci. 2014, 34, 8139–8150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.-O.; Suh, H.-S.; Si, Q.; Terman, B.I.; Lee, S.C. Anti-CD45RO Suppresses Human Immunodeficiency Virus Type 1 Replication in Microglia: Role of Hck Tyrosine Kinase and Implications for AIDS Dementia. J. Virol. 2006, 80, 62–72. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.; Town, T.; Mullan, M. CD45 Inhibits CD40L-Induced Microglial Activation via Negative Regulation of the Src/P44/42 MAPK Pathway. J. Biol. Chem. 2000, 275, 37224–37231. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.; Town, T.; Mori, T.; Wu, Y.; Saxe, M.; Crawford, F.; Mullan, M. CD45 Opposes β-Amyloid Peptide-Induced Microglial Activation via Inhibition of P44/42 Mitogen-Activated Protein Kinase. J. Neurosci. 2000, 20, 7587–7594. [Google Scholar] [CrossRef] [Green Version]
- Tepavčević, V.; Lazarini, F.; Alfaro-Cervello, C.; Kerninon, C.; Yoshikawa, K.; Garcia-Verdugo, J.M.; Lledo, P.-M.; Nait-Oumesmar, B.; Baron-Van Evercooren, A. Inflammation-Induced Subventricular Zone Dysfunction Leads to Olfactory Deficits in a Targeted Mouse Model of Multiple Sclerosis. J. Clin. Investig. 2011, 121, 4722–4734. [Google Scholar] [CrossRef] [PubMed]
- Fritze, J.; Ginisty, A.; McDonald, R.; Quist, E.; Stamp, E.; Monni, E.; Dhapola, P.; Lang, S.; Ahlenius, H. Loss of Cxcr5 Alters Neuroblast Proliferation and Migration in the Aged Brain. Stem Cells 2020, 38, 1175–1187. [Google Scholar] [CrossRef] [PubMed]
- Pantelis, C.; Jayaram, M.; Hannan, A.J.; Wesselingh, R.; Nithianantharajah, J.; Wannan, C.M.; Syeda, W.T.; Choy, K.C.; Zantomio, D.; Christopoulos, A.; et al. Neurological, Neuropsychiatric and Neurodevelopmental Complications of COVID-19. Aust. N. Z. J. Psychiatry 2021, 55, 750–762. [Google Scholar] [CrossRef] [PubMed]
- Herzine, A.; Laugeray, A.; Feat, J.; Menuet, A.; Quesniaux, V.; Richard, O.; Pichon, J.; Montécot-Dubourg, C.; Perche, O.; Mortaud, S. Perinatal Exposure to Glufosinate Ammonium Herbicide Impairs Neurogenesis and Neuroblast Migration through Cytoskeleton Destabilization. Front. Cell Neurosci. 2016, 10, 191. [Google Scholar] [CrossRef] [Green Version]
- Urra, F.; Araya-Maturana, R. Targeting Metastasis with Snake Toxins: Molecular Mechanisms. Toxins 2017, 9, 390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Huang, J.; Liu, Z. Vincristine Impairs Microtubules and Causes Neurotoxicity in Cerebral Organoids. Neuroscience 2019, 404, 530–540. [Google Scholar] [CrossRef]
- Francezon, N.; Herbaut, M.; Bardeau, J.-F.; Cougnon, C.; Bélanger, W.; Tremblay, R.; Jacquette, B.; Dittmer, J.; Pouvreau, J.-B.; Mouget, J.-L.; et al. Electrochromic Properties and Electrochemical Behavior of Marennine, a Bioactive Blue-Green Pigment Produced by the Marine Diatom Haslea Ostrearia. Mar. Drugs 2021, 19, 231. [Google Scholar] [CrossRef]
- Mouget, J.-L.; Gastineau, R.; Davidovich, O.; Gaudin, P.; Davidovich, N.A. Light Is a Key Factor in Triggering Sexual Reproduction in the Pennate Diatom Haslea Ostrearia. FEMS Microbiol. Ecol. 2009, 69, 194–201. [Google Scholar] [CrossRef] [Green Version]
- Schildge, S.; Bohrer, C.; Beck, K.; Schachtrup, C. Isolation and Culture of Mouse Cortical Astrocytes. J. Vis. Exp. 2013, e50079. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.; Geil Nickell, C.R.; Chen, K.Y.; McClain, J.A.; Nixon, K. Increased Expression of M1 and M2 Phenotypic Markers in Isolated Microglia after Four-Day Binge Alcohol Exposure in Male Rats. Alcohol 2017, 62, 29–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgehyr, N.; Meunier, A.; Faucourt, M.; Bosch Grau, M.; Strehl, L.; Janke, C.; Spassky, N. Ependymal Cell Differentiation, from Monociliated to Multiciliated Cells. Methods Cell Biol. 2015, 127, 19–35. [Google Scholar]
- Feat-Vetel, J.; Larrigaldie, V.; Meyer-Dilhet, G.; Herzine, A.; Mougin, C.; Laugeray, A.; Gefflaut, T.; Richard, O.; Quesniaux, V.; Montécot-Dubourg, C.; et al. Multiple Effects of the Herbicide Glufosinate-Ammonium and Its Main Metabolite on Neural Stem Cells from the Subventricular Zone of Newborn Mice. Neurotoxicology 2018, 69, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Bresciani, G.; Hofland, L.J.; Dogan, F.; Giamas, G.; Gagliano, T.; Zatelli, M.C. Evaluation of Spheroid 3D Culture Methods to Study a Pancreatic Neuroendocrine Neoplasm Cell Line. Front. Endocrinol. 2019, 10, 682. [Google Scholar] [CrossRef] [PubMed]
- Baumann, J.; Barenys, M.; Gassmann, K.; Fritsche, E. Comparative Human and Rat “Neurosphere Assay” for Developmental Neurotoxicity Testing. Curr. Protoc. Toxicol. 2014, 59, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Bouhlel, Z.; Arnold, A.A.; Deschênes, J.-S.; Mouget, J.-L.; Warschawski, D.E.; Tremblay, R.; Marcotte, I. Investigating the Action of the Microalgal Pigment Marennine on Vibrio Splendidus by in Vivo 2H and 31P Solid-State NMR. Biochim. Biophys. Acta (BBA)-Biomembr. 2021, 1863, 183642. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Méresse, S.; Gateau, H.; Tirnan, T.; Larrigaldie, V.; Casse, N.; Pasetto, P.; Mouget, J.-L.; Mortaud, S.; Fodil, M. Haslea ostrearia Pigment Marennine Affects Key Actors of Neuroinflammation and Decreases Cell Migration in Murine Neuroglial Cell Model. Int. J. Mol. Sci. 2023, 24, 5388. https://doi.org/10.3390/ijms24065388
Méresse S, Gateau H, Tirnan T, Larrigaldie V, Casse N, Pasetto P, Mouget J-L, Mortaud S, Fodil M. Haslea ostrearia Pigment Marennine Affects Key Actors of Neuroinflammation and Decreases Cell Migration in Murine Neuroglial Cell Model. International Journal of Molecular Sciences. 2023; 24(6):5388. https://doi.org/10.3390/ijms24065388
Chicago/Turabian StyleMéresse, Sarah, Hélène Gateau, Tessa Tirnan, Vanessa Larrigaldie, Nathalie Casse, Pamela Pasetto, Jean-Luc Mouget, Stéphane Mortaud, and Mostefa Fodil. 2023. "Haslea ostrearia Pigment Marennine Affects Key Actors of Neuroinflammation and Decreases Cell Migration in Murine Neuroglial Cell Model" International Journal of Molecular Sciences 24, no. 6: 5388. https://doi.org/10.3390/ijms24065388