
Overexpression of the Wheat TaPsb28 Gene Enhances Drought Tolerance in Transgenic Arabidopsis
Abstract
:1. Introduction
2. Results and Discussion
2.1. Sequence Analysis of the Wheat TaPsb28 Gene
2.2. Expression Pattern under Drought Stress and Subcellular Localization of Wheat TaPsb28
2.3. Generation of TaPsb28-Overexpressing Transgenic Arabidopsis Plants and Subcellular Localization
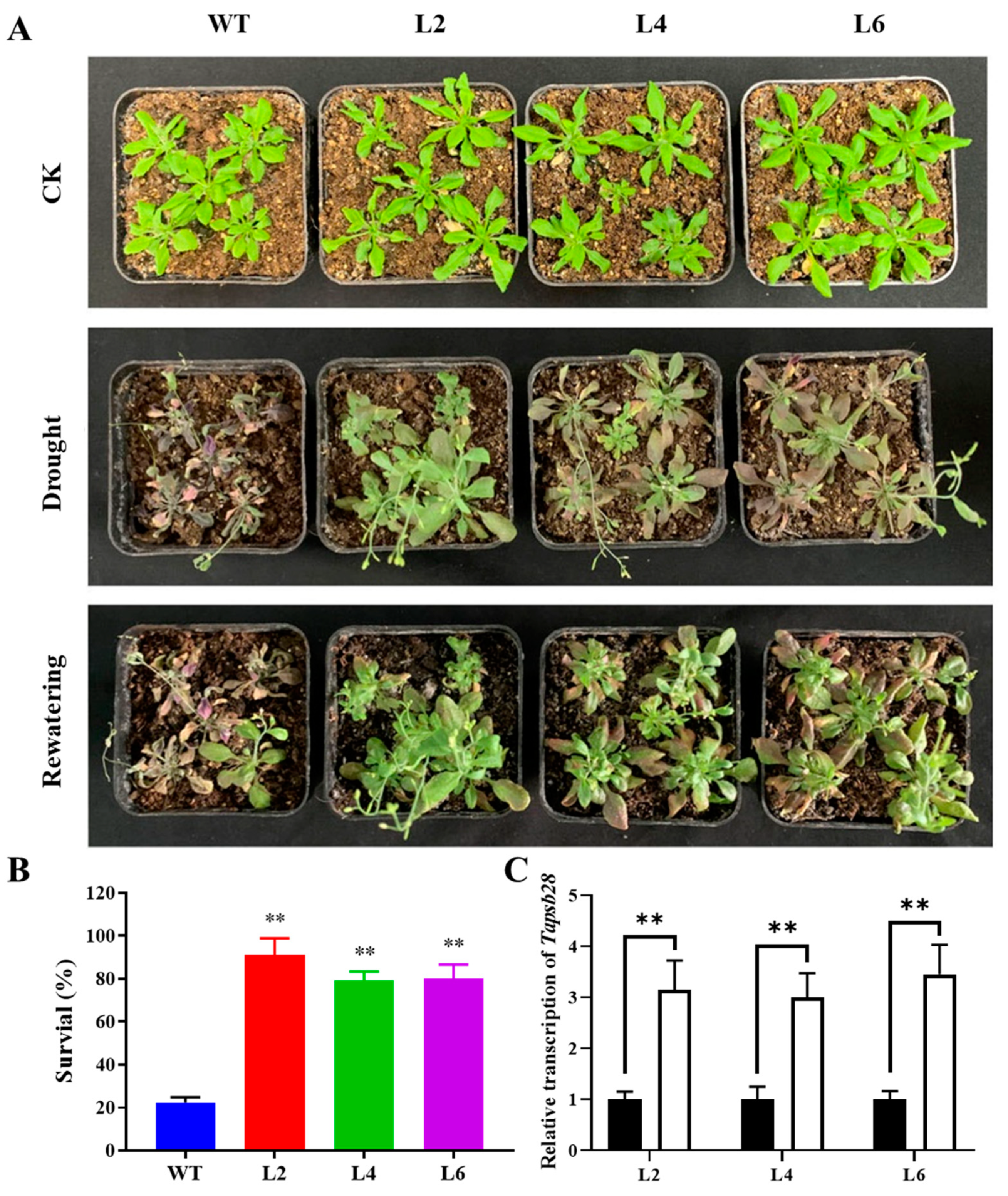
2.4. Overexpression of TaPsb28 Enhanced the Drought Tolerance of Transgenic Arabidopsis Seedlings
2.5. Overexpression of TaPsb28 Decreased the Stomatal Aperture and Water Loss under Drought Stress
2.6. Overexpression of TaPsb28 Increased the Chlorophyll Content and Decreased the MDA Content under Drought Stress
2.7. Overexpression of TaPsb28 Alleviates the Increases in ABA and Zeatin Contents but Improves the Increases in Cyanidin, Delphinidin, and PAs Contents under Drought Stress
2.8. Overexpression of TaPsb28 Improved ChlG, DFR, and ANR Gene Transcription but Alleviated the Induction of RD22 Gene Transcription under Drought Stress
3. Materials and Methods
3.1. Multiple Sequence Alignment and Bioinformatics Analysis of the TaPsb28 Gene
3.2. Wheat Plant Materials, Growth Conditions and Drought Stress Treatment
3.3. Vector Construction
3.4. Transformation of Arabidopsis
3.5. Subcellular Localization of TaPsb28
3.6. Drought Stress in Arabidopsis
3.7. Measurement of Physiological Indices
3.8. Measurement of Stomatal Aperture
3.9. Quantification of Endogenous ABA and Zeatin
3.10. Determination of Endogenous Cyanidin and Delphinidin Contents
3.11. Extraction and Quantification of Proanthocyanidin
3.12. Total RNA Extraction, Reverse Transcription, and Real-Time PCR Quantification
3.13. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviation
| ABA | Abscisic acid |
| ANR | Anthocyanin reductase |
| CDS | Coding sequence |
| ChlG | Chlorophyll synthase |
| CK | Control |
| DFR | Dihydroflavonol 4-reductase |
| LB | Luria-Bertani |
| MDA | Malondialdehyde |
| NCBI | National Center for Biotechnology Information |
| PSII | Photosystem II |
| ROS | Reactive oxygen species |
| SD | Standard deviation |
| WT | Wild-type |
References
- Mohammadi, R. Breeding for increased drought tolerance in wheat: A review. Crop. Pasture Sci. 2018, 69, 223–241. [Google Scholar] [CrossRef]
- Chang, L.; Wang, L.; Peng, C.; Tong, Z.; Wang, D.; Ding, G.; Xiao, J.; Guo, A.; Wang, X. The chloroplast proteome response to drought stress in cassava leaves. Plant Physiol. Biochem. 2019, 142, 351–362. [Google Scholar] [CrossRef]
- Ambastha, V.; Chauhan, G.; Tiwari, B.S.; Tripathy, B.C. Execution of programmed cell death by singlet oxygen generated inside the chloroplasts of Arabidopsis thaliana. Protoplasma 2020, 257, 841–851. [Google Scholar] [CrossRef]
- Schmidt, A.; Mächtel, R.; Ammon, A.; Engelsdorf, T.; Schmitz, J.; Maurino, V.G.; Voll, L.M. Reactive oxygen species dosage in Arabidopsis chloroplasts can improve resistance towards Colletotrichum higginsianum by the induction of WRKY33. New Phytol. 2020, 226, 189–204. [Google Scholar] [CrossRef] [Green Version]
- Sakata, S.; Mizusawa, N.; Kubota-Kawai, H.; Sakurai, I.; Wada, H. Psb28 is involved in recovery of photosystem II at high temperature in Synechocystis sp. PCC 6803. Biochim. Biophys. Acta 2013, 1827, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Yang, M.; Wei, S.; Qin, F.; Zhao, H.; Suo, B. Identification of circular RNAs and their targets in leaves of Triticum aestivum L. under dehydration stress. Front. Plant Sci. 2017, 7, 2024. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.X.; Chen, X.Y.; Li, X.Y.; Song, Y.H.; Wang, J.A.; Huang, J.; Xue, R.L. Exogenous application of 5-aminolevulinic acid alleviated damage to wheat chloroplast ultrastructure under drought stress by transcriptionally regulating genes correlated with photosynthesis and chlorophyll biosynthesis. Acta Physiol. Plant. 2022, 44, 12. [Google Scholar] [CrossRef]
- Li, X.; Ding, M.; Wang, M.; Yang, S.; Ma, X.; Hu, J.; Song, F.; Wang, L.; Liang, W. Proteome profiling reveals changes in energy metabolism, transport and antioxidation during drought stress in Nostoc flagelliforme. BMC Plant Biol. 2022, 22, 162. [Google Scholar] [CrossRef]
- Cao, X.; Hu, Y.; Song, J.; Feng, H.; Wang, J.; Chen, L.; Wang, L.; Diao, X.; Wan, Y.; Liu, S.; et al. Transcriptome sequencing and metabolome analysis reveals the molecular mechanism of drought stress in millet. Int. J. Mol. Sci. 2022, 23, 10792. [Google Scholar] [CrossRef]
- Wang, B.; Liu, C.; Zhang, D.; He, C.; Zhang, J.; Li, Z. Effects of maize organ-specific drought stress response on yields from transcriptome analysis. BMC Plant Biol. 2019, 19, 335. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.H.; Lee, J.; Dardick, C.; Seo, Y.S.; Cao, P.; Canlas, P.; Phetsom, J.; Xu, X.; Ouyang, S.; An, K.; et al. Identification and functional analysis of light-responsive unique genes and gene family members in rice. PLoS Genet. 2008, 4, e1000164. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Chen, X.; Xu, B.; Li, Y.; Ma, Y.; Wang, G. Phenotype and transcriptome analysis reveals chloroplast development and pigment biosynthesis together influenced the leaf color formation in mutants of Anthurium andraeanum ‘Sonate’. Front. Plant Sci. 2015, 6, 139. [Google Scholar] [CrossRef] [Green Version]
- Shalygo, N.; Czarnecki, O.; Peter, E.; Grimm, B. Expression of chlorophyll synthase is also involved in feedback-control of chlorophyll biosynthesis. Plant Mol. Biol. 2009, 71, 425–436. [Google Scholar] [CrossRef]
- Lin, Y.P.; Lee, T.Y.; Tanaka, A.; Charng, Y.Y. Analysis of an Arabidopsis heat-sensitive mutant reveals that chlorophyll synthase is involved in reutilization of chlorophyllide during chlorophyll turnover. Plant J. 2014, 80, 14–26. [Google Scholar] [CrossRef]
- Lin, Y.P.; Shen, Y.Y.; Shiu, Y.B.; Charng, Y.Y.; Grimm, B. Chlorophyll dephytylase 1 and chlorophyll synthase: A chlorophyll salvage pathway for the turnover of photosystems I and II. Plant J. 2022, 111, 979–994. [Google Scholar] [CrossRef]
- Nakabayashi, R.; Yonekura-Sakakibara, K.; Urano, K.; Suzuki, M.; Yamada, Y.; Nishizawa, T.; Matsuda, F.; Kojima, M.; Sakakibara, H.; Shinozaki, K.; et al. Enhancement of oxidative and drought tolerance in Arabidopsis by overaccumulation of antioxidant flavonoids. Plant J. 2014, 77, 367–379. [Google Scholar] [CrossRef] [Green Version]
- Loreti, E.; Povero, G.; Novi, G.; Solfanelli, C.; Alpi, A.; Perata, P. Gibberellins, jasmonate and abscisic acid modulate the sucrose-induced expression of anthocyanin biosynthetic genes in Arabidopsis. New Phytol. 2008, 179, 1004–1016. [Google Scholar] [CrossRef]
- Adamipour, N.; Khosh-Khui, M.; Salehi, H.; Razi, H.; Karami, A.; Moghadam, A. Role of genes and metabolites involved in polyamines synthesis pathways and nitric oxide synthase in stomatal closure on Rosa damascena Mill. under drought stress. Plant Physiol. Biochem. 2020, 148, 53–61. [Google Scholar] [CrossRef]
- Zhang, F.-P.; Sussmilch, F.; Nichols, D.S.; Cardoso, A.A.; Brodribb, T.J.; McAdam, S.A. Leaves, not roots or floral tissue, are the main site of rapid, external pressure-induced ABA biosynthesis in angiosperms. J. Exp. Bot. 2018, 69, 1261–1267. [Google Scholar] [CrossRef]
- Schafer, M.; Brutting, C.; Meza-Canales, I.D.; Grosskinsky, D.K.; Vankova, R.; Baldwin, I.T.; Meldau, S. The role of cis-zeatin-type cytokinins in plant growth regulation and mediating responses to environmental interactions. J. Exp. Bot. 2015, 66, 4873–4884. [Google Scholar] [CrossRef] [Green Version]
- Li, G.G.; Liang, Z.M.; Li, Y.J.; Liao, Y.C.; Liu, Y. Exogenous spermidine regulates starch synthesis and the antioxidant system to promote wheat grain filling under drought stress. Acta Physiol. Plant. 2020, 42, 110. [Google Scholar] [CrossRef]
- Yu, Y.A.; Zhang, L. Overexpression of TaWRKY53 enhances drought tolerance in transgenic Arabidopsis plants. S. Afr. J. Bot. 2022, 148, 605–614. [Google Scholar] [CrossRef]
- Peng, L.N.; Xu, Y.Q.; Wang, X.; Feng, X.; Zhao, Q.Q.; Feng, S.S.; Zhao, Z.Y.; Hu, B.Z.; Li, F.L. Overexpression of paralogues of the wheat expansin gene TaEXPA8 improves low-temperature tolerance in Arabidopsis. Plant Biol. 2019, 21, 1119–1131. [Google Scholar] [CrossRef]
- Chen, Z.; Gallie, D.R. The ascorbic acid redox state controls guard cell signaling and stomatal movement. Plant Cell 2004, 16, 1143–1162. [Google Scholar] [CrossRef] [Green Version]
- Nelson, N.; Yocum, C.F. Structure and function of photosystems I and II. Annu. Rev. Plant Biol. 2006, 57, 521–565. [Google Scholar] [CrossRef] [Green Version]
- Beckova, M.; Gardian, Z.; Yu, J.; Konik, P.; Nixon, P.J.; Komenda, J. Association of Psb28 and Psb27 proteins with PSII-PSI supercomplexes upon exposure of Synechocystis sp. PCC 6803 to high light. Mol. Plant 2017, 10, 62–72. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T.; Mogami, J.; Yamaguchi-Shinozaki, K. ABA-dependent and ABA-independent signaling in response to osmotic stress in plants. Curr. Opin. Plant Biol. 2014, 21, 133–139. [Google Scholar] [CrossRef]
- Zhao, Y.; Du, H.; Wang, Y.; Wang, H.; Yang, S.; Li, C.; Chen, N.; Yang, H.; Zhang, Y.; Zhu, Y.; et al. The calcium-dependent protein kinase ZmCDPK7 functions in heat-stress tolerance in maize. J. Integr. Plant Biol. 2021, 63, 510–527. [Google Scholar] [CrossRef]
- Figueiredo-Lima, K.V.; Falcao, H.M.; Melo-de-Pinna, G.F.; Albacete, A.; Dodd, I.C.; Lima, A.L.; Santos, M.G. Leaf phytohormone levels and stomatal control in an evergreen woody species under semiarid environment in a Brazilian seasonally dry tropical forest. Plant Growth Regul. 2018, 85, 437–445. [Google Scholar] [CrossRef]
- Wang, Y.X.; Wei, S.M.; Wang, J.N.; Su, X.Y.; Suo, B.; Qin, F.J.; Zhao, H.J. Exogenous application of 5-aminolevulinic acid on wheat seedlings under drought stress enhances the transcription of psbA and psbD genes and improves photosynthesis. Braz. J. Bot. 2018, 41, 275–285. [Google Scholar] [CrossRef]
- Jogawat, A.; Yadav, B.; Chhaya; Lakra, N.; Singh, A.K.; Narayan, O.P. Crosstalk between phytohormones and secondary metabolites in the drought stress tolerance of crop plants: A review. Physiol. Plant. 2021, 172, 1106–1132. [Google Scholar] [CrossRef]
- Gao, H.; Yu, W.; Yang, X.; Liang, J.; Sun, X.; Sun, M.; Xiao, Y.; Peng, F. Silicon enhances the drought resistance of peach seedlings by regulating hormone, amino acid, and sugar metabolism. BMC Plant Biol. 2022, 22, 422. [Google Scholar] [CrossRef]
- Wakamatsu, A.; Mori, I.C.; Matsuura, T.; Taniwaki, Y.; Ishii, R.; Yoshida, R. Possible roles for phytohormones in controlling the stomatal behavior of Mesembryanthemum crystallinum during the salt-induced transition from C3 to crassulacean acid metabolism. J. Plant Physiol. 2021, 262, 153448. [Google Scholar] [CrossRef]
- González-Villagra, J.; Cohen, J.D.; Reyes-Díaz, M.M. Abscisic acid is involved in phenolic compounds biosynthesis, mainly anthocyanins, in leaves of Aristotelia chilensis plants (Mol.) subjected to drought stress. Physiol. Plant. 2019, 165, 855–866. [Google Scholar] [CrossRef]
- Gao, H.N.; Jiang, H.; Cui, J.Y.; You, C.X.; Li, Y.Y. The effects of hormones and environmental factors on anthocyanin biosynthesis in apple. Plant Sci. 2021, 312, 111024. [Google Scholar] [CrossRef]
- An, J.P.; Zhang, X.W.; Bi, S.Q.; You, C.X.; Wang, X.F.; Hao, Y.J. The ERF transcription factor MdERF38 promotes drought stress-induced anthocyanin biosynthesis in apple. Plant J. 2020, 101, 573–589. [Google Scholar] [CrossRef]
- Wang, H.; Jin, M.; Xu, L.; Xi, H.; Wang, B.; Du, S.; Liu, H.; Wen, Y. Effects of ketoprofen on rice seedlings: Insights from photosynthesis, antioxidative stress, gene expression patterns, and integrated biomarker response analysis. Environ. Pollut. 2020, 263, 114533. [Google Scholar] [CrossRef]
- Zhao, Y.; Han, Q.; Ding, C.; Huang, Y.; Liao, J.; Chen, T.; Feng, S.; Zhou, L.; Zhang, Z.; Chen, Y.; et al. Effect of low temperature on chlorophyll biosynthesis and chloroplast biogenesis of rice seedlings during greening. Int. J. Mol. Sci. 2020, 21, 1390. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, Y.; Hassan, M.J.; Li, Z.; Peng, Y. Indole-3-acetic acid improves drought tolerance of white clover via activating auxin, abscisic acid and jasmonic acid related genes and inhibiting senescence genes. BMC Plant Biol. 2020, 20, 150. [Google Scholar] [CrossRef] [Green Version]
- Seo, P.J.; Park, C.-M. Cuticular wax biosynthesis as a way of inducing drought resistance. Plant Signal. Behav. 2011, 6, 1043–1045. [Google Scholar] [CrossRef]
- Wade, H.K.; Sohal, A.K.; Jenkins, G.I. Arabidopsis ICX1 is a negative regulator of several pathways regulating flavonoid biosynthesis genes. Plant Physiol. 2003, 131, 707–715. [Google Scholar] [CrossRef] [Green Version]
- Deikman, J.; Hammer, P.E. Induction of anthocyanin accumulation by cytokinins in Arabidopsis thaliana. Plant Physiol. 1995, 108, 47–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Z.H.; Bi, W.L.; Hao, X.Y.; Li, P.M.; Duan, Y.; Walker, M.A.; Xu, Y.; Wang, Q.C. Drought stress enhances up-regulation of anthocyanin biosynthesis in grapevine leafroll-associated virus 3-infected in vitro grapevine (Vitis vinifera) leaves. Plant Dis. 2017, 101, 1606–1615. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Yang, J.; Pak, S.; Zeng, M.; Sun, J.; Yu, S.; He, Y.; Li, C. PuC3H35 confers drought tolerance by enhancing lignin and proanthocyanidin biosynthesis in the roots of Populus ussuriensis. New Phytol. 2022, 233, 390–408. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Wang, Y.; Li, X.; Liu, N.; Wei, S.; Wang, J.; Qin, F.; Suo, B. The iTRAQ-based chloroplast proteomic analysis of Triticum aestivum L. leaves subjected to drought stress and 5-aminolevulinic acid alleviation reveals several proteins involved in the protection of photosynthesis. BMC Plant Biol. 2020, 20, 96. [Google Scholar] [CrossRef] [Green Version]
- Liang, W.; Cui, W.; Ma, X.; Wang, G.; Huang, Z. Function of wheat Ta-UnP gene in enhancing salt tolerance in transgenic Arabidopsis and rice. Biochem. Biophys. Res. Commun. 2014, 450, 794–801. [Google Scholar] [CrossRef]
- Shi, H.; Liu, G.; Wei, Y.; Chan, Z. The zinc-finger transcription factor ZAT6 is essential for hydrogen peroxide induction of anthocyanin synthesis in Arabidopsis. Plant Mol. Biol. 2018, 97, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Li, H.; Lian, H.; Song, P.; Wang, Y.; Duan, J.; Song, Z.; Cao, Y.; Xu, D.; Li, J.; et al. SICKLE represses photomorphogenic development of Arabidopsis seedlings via HY5- and PIF4-mediated signaling. J. Integr. Plant Biol. 2022, 64, 1706–1723. [Google Scholar] [CrossRef]
- Hotto, A.M.; Salesse-Smith, C.; Lin, M.; Busch, F.A.; Simpson, I.; Stern, D.B. Rubisco production in maize mesophyll cells through ectopic expression of subunits and chaperones. J. Exp. Bot. 2021, 72, 4930–4937. [Google Scholar] [CrossRef] [PubMed]
- Jia, T.; Hou, J.; Iqbal, M.Z.; Zhang, Y.; Cheng, B.; Feng, H.; Li, Z.; Liu, L.; Zhou, J.; Feng, G.; et al. Overexpression of the white clover TrSAMDC1 gene enhanced salt and drought resistance in Arabidopsis thaliana. Plant Physiol. Biochem. 2021, 165, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Porra, R.; Thompson, W.; Kriedemann, P. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta (BBA)-Bioenerg. 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Majcherczyk, A.; Rakoczy, L.; Hüttermann, A. Improvements in methods for determination of abscisic acid and indole-3-acetic acid by high-performance liquid chromatography. J. Chromatogr. A 1986, 357, 399–408. [Google Scholar] [CrossRef]
- Yuan, L.; Wang, L.; Han, Z.; Jiang, Y.; Zhao, L.; Liu, H.; Yang, L.; Luo, K. Molecular cloning and characterization of PtrLAR3, a gene encoding leucoanthocyanidin reductase from Populus trichocarpa, and its constitutive expression enhances fungal resistance in transgenic plants. J. Exp. Bot. 2012, 63, 2513–2524. [Google Scholar] [CrossRef]
- Porter, L.J.; Hrstich, L.N.; Chan, B.G. The conversion of procyanidins and prodelphinidins to cyanidin and delphinidin. Phytochemistry 1985, 25, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′-3′) | Purpose | Reference |
|---|---|---|---|
| Psb28-F | ATGCCGGCAGTGATGAAAGC | Clone of TaPsb28 in wheat | This study |
| Psb28-R | GTTCTTGACGAACTGGAGGCC | ||
| Actin-1-F | TGCTATCCTTCGTTTGGACCTT | qRT–PCR in wheat | [47] |
| Actin-1-R | AGCGGTTGTTGTGAGGGAGT | ||
| Actin-2-F | CTTAACCCAAAGGCCAACAGA | qRT–PCR in Arabidopsis | This study |
| Actin-2-R | GCAAGGTCAAGACGGAGGAT | ||
| qPCR-Psb28–1-F | AGCTGAATGCTTGTACCGAGT | qRT–PCR in wheat | This study |
| qPCR-Psb28–1-R | CGGTTTTCCTCATTACGTGCTT | ||
| qPCR-Psb28–2-F | AAAGCCTGCAGTCCTCCTTC | qRT–PCR in Arabidopsis | This study |
| qPCR-Psb28–2-R | GTGCCCTGGATGAACTGGAT | ||
| PYFPLT-TaPsb28-F | acaaatctatctctctcgagATGCCGGCAGTGATGAAAGC | Construction of subcellular vector | This study |
| PYFPLT-TaPsb28-R | ctacgcgtgagctcaagcttGTTCTTGACGAACTGGAGGCC | ||
| 1302-TaPsb28-F | cttgaccatggtagatctgactagtATGCCGGCAGTGATGAA | Construction of expression vector | This study |
| 1302-TaPsb28-R | gaaaagttcttctcctttactagtGTTCTTGACGAACTGGAGGCC | ||
| RD22-F | GCGATTCGTCTTCCTCTGAT | qRT–PCR | This study |
| RD22-R | CTCCGCCTTTACCTACTTGG | ||
| ChlG-F | CTTCCGTCGGTTCTATG | qRT–PCR | This study |
| ChlG-F | CTTCCGTCGGTTCTATG | ||
| DFR-F | CCTTATCACCGCGCTCTCT | qRT–PCR | [48] |
| DFR-R | TGTCCTTGTCTTATGATCGAGTAATGC | ||
| ANR-F | AAGAAAACTGGACTGACGTTGAA | qRT–PCR | [48] |
| ANR-F | AACACCTTCGAGATTGGGTAAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhang, M.; Li, X.; Zhou, R.; Xue, X.; Zhang, J.; Liu, N.; Xue, R.; Qi, X. Overexpression of the Wheat TaPsb28 Gene Enhances Drought Tolerance in Transgenic Arabidopsis. Int. J. Mol. Sci. 2023, 24, 5226. https://doi.org/10.3390/ijms24065226
Wang Y, Zhang M, Li X, Zhou R, Xue X, Zhang J, Liu N, Xue R, Qi X. Overexpression of the Wheat TaPsb28 Gene Enhances Drought Tolerance in Transgenic Arabidopsis. International Journal of Molecular Sciences. 2023; 24(6):5226. https://doi.org/10.3390/ijms24065226
Chicago/Turabian StyleWang, Yuexia, Menghan Zhang, Xiaoyan Li, Ruixiang Zhou, Xinyu Xue, Jing Zhang, Nana Liu, Ruili Xue, and Xueli Qi. 2023. "Overexpression of the Wheat TaPsb28 Gene Enhances Drought Tolerance in Transgenic Arabidopsis" International Journal of Molecular Sciences 24, no. 6: 5226. https://doi.org/10.3390/ijms24065226