
Comparison of Phenotype and Genotype Virulence and Antimicrobial Factors of Salmonella Typhimurium Isolated from Human Milk
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
2.1. Identification of the Isolated Strain
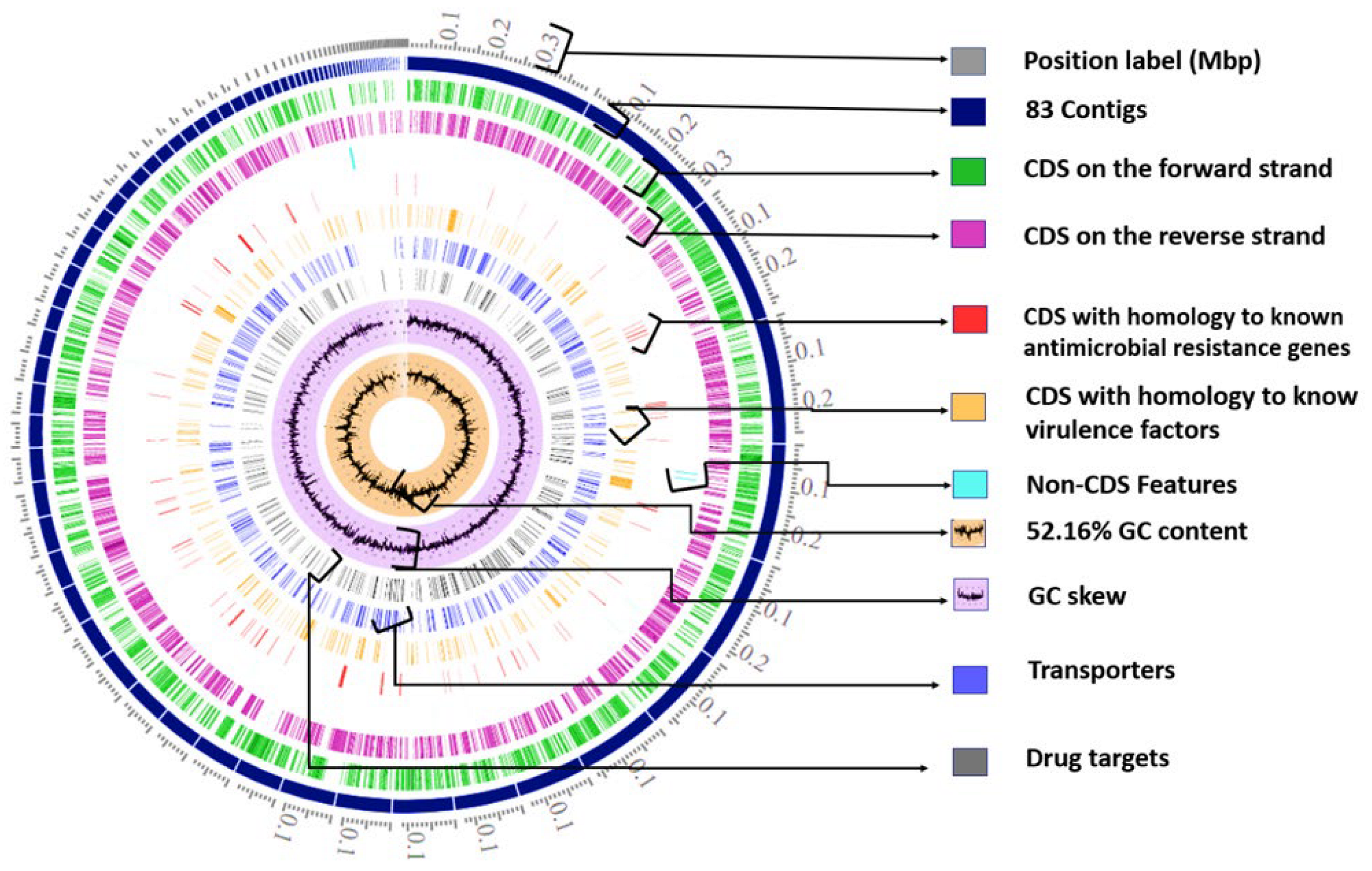
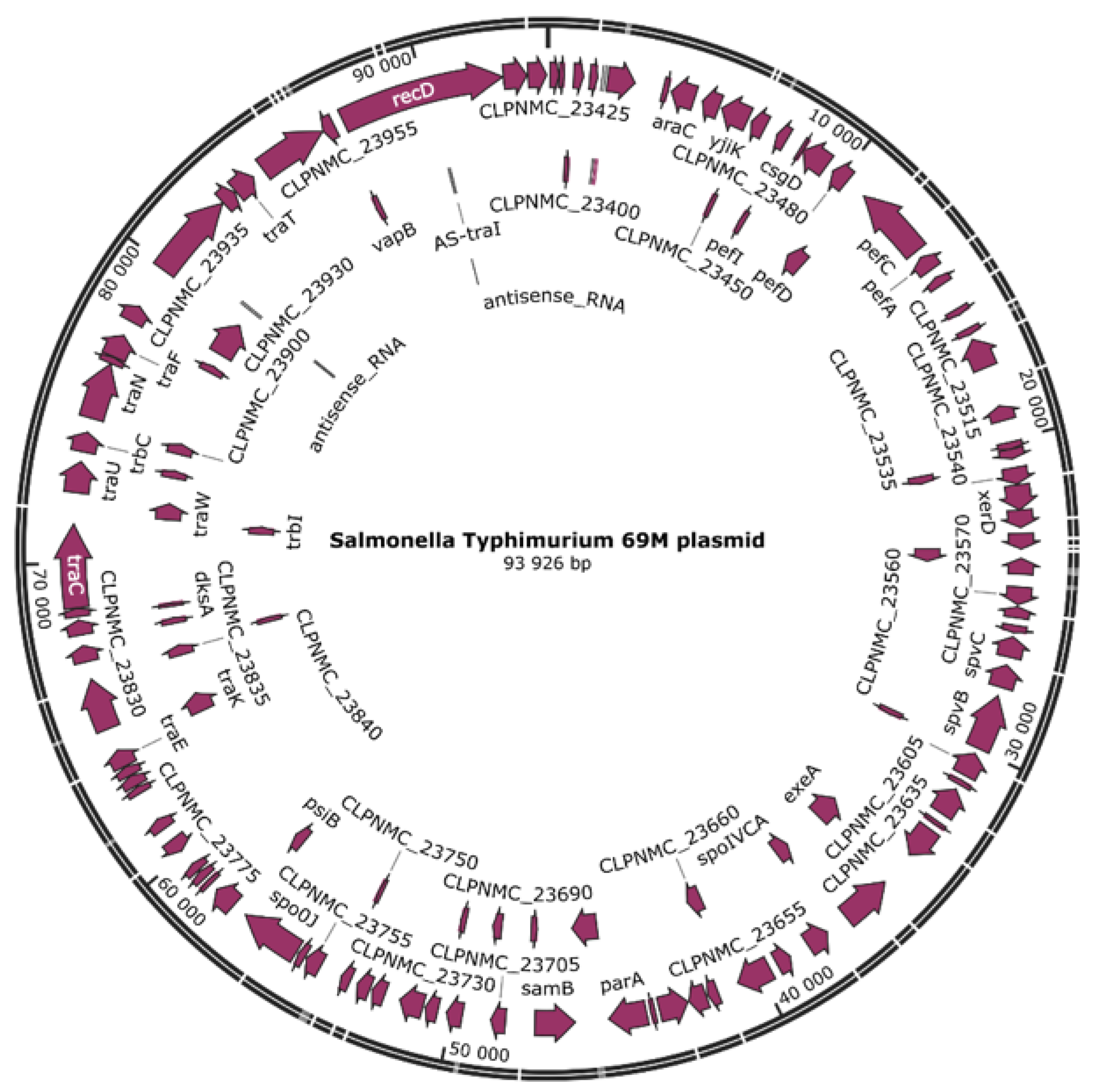
2.1.1. Whole-Genome Sequencing (WGS) and Bioinformatic Analysis
The Genome Assembly and Annotation
2.1.2. Serotype Determination Using Whole-Genome Sequencing Data
2.2. Analysis for Virulence and Antibiotic Resistance Genes
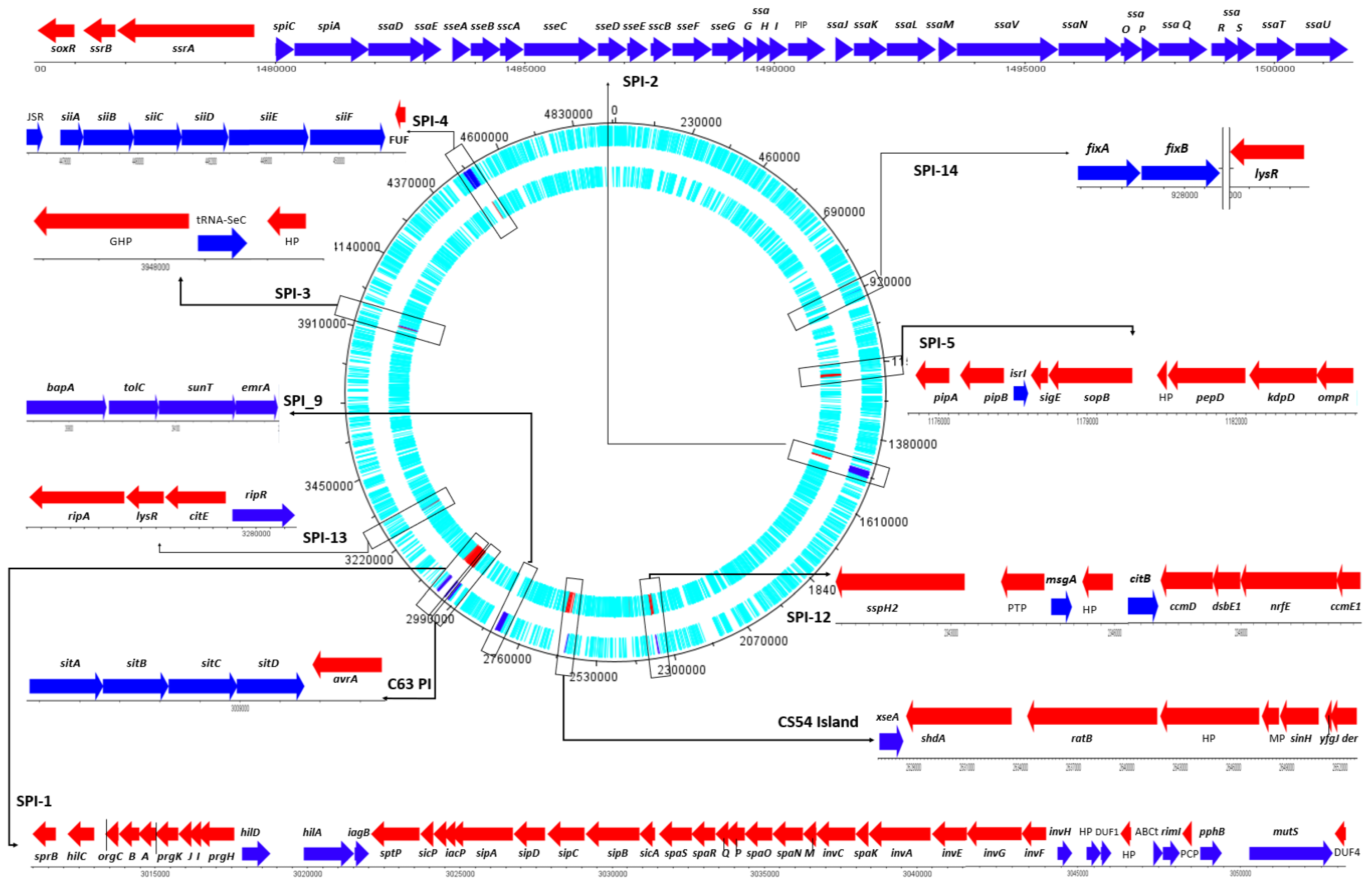
2.2.1. Salmonella Pathogenic Island (SPI)
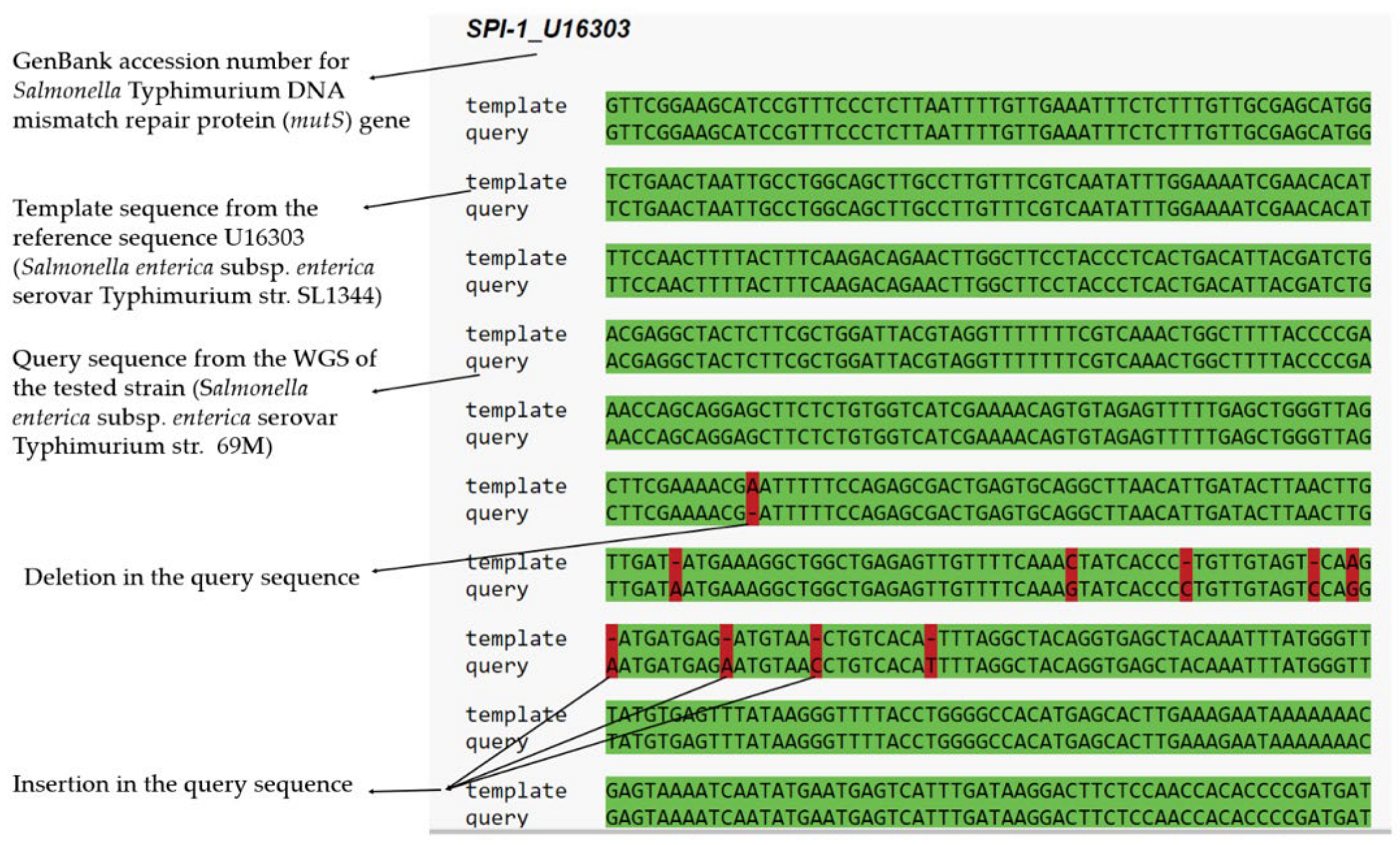
2.2.2. Multiple Sequence Alignment (MSA) and Single-Nucleotide Polymorphism (SNP)
2.2.3. Antimicrobal Resistance Genes
3. Discussion
4. Materials and Methods
4.1. Serological Testing
4.2. Biochemical Strain Identification
4.3. Confirmation of Salmonella Identification with Molecular Biology Methods
4.4. Whole-Genome Sequencing
4.5. Antimicrobial Sensitivity Testing: Phenotypic Antibiotic Resistance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- EFSA (European Food Safety Authority); ECDC (European Centre for Disease Prevention and Control). The European Union Summary Report on Antimicrobial Resistance in zoonotic and indicator bacteria from humans, animals and food in 2019–2020. EFSA J. 2022, 20, 1–197. [Google Scholar]
- Havelaar, A.H.; Kirk, M.D.; Torgerson, P.R.; Gibb, H.J.; Hald, T.; Lake, R.J.; Praet, N.; Bellinger, D.C.; de Silva, N.R.; Gargouri, N.; et al. World Health Organization Global Estimates and Regional Comparisons of the Burden of Foodborne Disease in 2010. PLoS Med. 2015, 12, e1001923. [Google Scholar] [CrossRef] [Green Version]
- Ao, T.T.; Feasey, N.A.; Gordon, M.A.; Keddy, K.H.; Angulo, F.J.; Crump, J.A. Global burden of invasive nontyphoidal Salmonella disease, 2010. Emerg. Infect. Dis. 2015, 21, 941–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gal-Mor, O.; Finlay, B.B. Pathogenicity islands: A molecular toolbox for bacterial virulence. Cell. Microbiol. 2006, 8, 1707–1719. [Google Scholar] [CrossRef]
- Schmidt, H.; Hensel, M. Pathogenicity Islands in Bacterial Pathogenesis. Clin. Microbiol. Rev. 2004, 17, 14–56. [Google Scholar] [CrossRef] [Green Version]
- Deriu, E.; Liu, J.Z.; Pezeshki, M.; Edwards, R.A.; Ochoa, R.J.; Contreras, H.; Libby, S.J.; Fang, F.C.; Raffatellu, M. Probiotic bacteria reduce Salmonella Typhimurium intestinal colonization by competing for iron. Cell Host Microbe 2013, 14, 26–37. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Wang, P.; Song, X.; Zhang, H.; Ma, S.; Wang, J.; Li, W.; Lv, R.; Liu, X.; Ma, S.; et al. Salmonella Typhimurium reprograms macrophage metabolism via T3SS effector SopE2 to promote intracellular replication and virulence. Nat. Commun. 2021, 12, 879. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, G.K.; Ethelberg, S. Investigating Outbreaks of Salmonella Typhimurium Using Case-Control Studies, with a Reference to the One Health Approach, 3rd ed.; Schatten, H., Ed.; Springer: Hatfield, UK, 2021; Volume 2182, ISBN 9781071607909. [Google Scholar]
- Lund, S.; Tahir, M.; Vohra, L.I.; Hamdana, A.H.; Ahmad, S. Outbreak of monophasic Salmonella Typhimurium Sequence Type 34 linked to chocolate products. Ann. Med. Surg. 2022, 82, 104597. [Google Scholar] [CrossRef]
- Ryder, R.W.; Crosby Ritchie, A.; Mcdonough, B.; Hall, W.J. Human Milk Contaminated with Salmonella Kottbus: A Cause of Nosocomial Illness in Infants. JAMA J. Am. Med. Assoc. 1977, 238, 1533–1534. [Google Scholar] [CrossRef]
- Revathi, G.; Mahajan, R.; Faridi, M.M.A.; Kumar, A.; Talwar, V. Transmission of lethal Salmonella Senftenberg from mother’s breast-milk to her baby. Ann. Trop. Paediatr. 1995, 15, 159–161. [Google Scholar] [CrossRef]
- Chen, T.L.; Thien, P.F.; Liaw, S.C.; Fung, C.P.; Siu, L.K. First report of Salmonella enterica serotype Panama meningitis associated with consumption of contaminated breast milk by a neonate. J. Clin. Microbiol. 2005, 43, 5400–5402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drhová, A.; Dobiásová, V.; Stefkovicová, M. Mother’s milk—Unusual factor of infection transmission in a salmonellosis epidemic on a newborn ward. J. Hyg. Epidemiol. Microbiol. Immunol. 1990, 34, 353–355. [Google Scholar] [PubMed]
- Blackshaw, K.; Valtchev, P.; Koolaji, N.; Berry, N.; Schindeler, A.; Dehghani, F.; Banati, R.B. The risk of infectious pathogens in breast-feeding, donated human milk and breast milk substitutes. Public Health Nutr. 2021, 24, 1725–1740. [Google Scholar] [CrossRef]
- Wesołowska, A. Banki mleka w Polsce : Funkcjonowanie w Podmiotach Leczniczych: Idea i Praktyka; Wesołowska, A., Ed.; Fundacja Bank Mleka Kobiecego: Warsaw, Poland, 2017; ISBN 9788393793310. [Google Scholar]
- Qutaishat, S.S.; Stemper, M.E.; Spencer, S.K.; Opitz, J.C.; Monson, T.A.; Anderson, J.L. Transmission of Salmonella enterica Serotype Typhimurium DT104 to Infants Through Mother’s Breast Milk. Pediatrics 2003, 111, 1442–1446. [Google Scholar] [CrossRef]
- Mitchell, H.; Ebbeson, R.; Maclean, M.; Hajek, J. A mother with Salmonella mastitis and a baby with Salmonella bacteremia. Can. Fam. Physician 2021, 67, 747–749. [Google Scholar] [CrossRef]
- Seah, X.F.V.; Ngeow, J.H.A.; Thoon, K.C.; Tee, W.S.N.; Maiwald, M.; Chong, C.Y.; Tan, N.W.H. Infant with Invasive Nontyphoidal Salmonellosis and Mastitis. Glob. Pediatr. Health 2015, 2, 2333794X1559156. [Google Scholar] [CrossRef]
- Wang, X.; Biswas, S.; Paudyal, N.; Pan, H.; Li, X.; Fang, W.; Yue, M. Antibiotic resistance in Salmonella Typhimurium isolates recovered from the food chain through national antimicrobial resistance monitoring system between 1996 and 2016. Front. Microbiol. 2019, 10, 985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karabay, M.; Şehmusoğlu, Z.; Terzi, H.A.; Caner, İ.; Köroğlu, M. Neonatal Sepsis due to Salmonella enteritidis from Colonized Breast Milk; A Case Report and Literature Review of Breast Milk-induced Neonatal Sepsis. Flora J. Infect. Dis. Clin. Microbiol. 2021, 26, 323–327. [Google Scholar] [CrossRef]
- Maroszyńska, I.; Fortecka-Piestrzeniewicz, K.; Niedźwiecka, M. Salmonella Typhimurium—A threat for a newborn. Open J. Obstet. Gynecol. 2013, 3, 628–630. [Google Scholar] [CrossRef] [Green Version]
- Pławińska-Czarnak, J.; Wódz, K.; Kizerwetter-świda, M.; Nowak, T.; Bogdan, J.; Kwieciński, P.; Kwieciński, A.; Anusz, K. Citrobacter braakii yield false-positive identification as salmonella, a note of caution. Foods 2021, 10, 2177. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the all-bacterial bioinformatics database and analysis resource center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef]
- Olson, R.D.; Assaf, R.; Brettin, T.; Conrad, N.; Cucinell, C.; Davis, J.J.; Dempsey, D.M.; Dickerman, A.; Dietrich, E.M.; Kenyon, R.W.; et al. Introducing the Bacterial and Viral Bioinformatics Resource Center (BV-BRC): A resource combining PATRIC, IRD and ViPR. Nucleic Acids Res. 2022, 51, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Schomburg, I.; Chang, A.; Ebeling, C.; Gremse, M.; Heldt, C.; Huhn, G.; Schomburg, D. BRENDA, the enzyme database: Updates and major new developments. Nucleic Acids Res. 2004, 32, D431–D433. [Google Scholar] [CrossRef] [Green Version]
- Finke, W.; Rachimow, C.; Pfützner, B. Untersuchungen zu Wasserdargebot und Wasserverfügbarkeit im Ballungsraum Berlin im Rahmen des Verbundprojektes GLOWA Elbe. Hydrol. Wasserbewirtsch. 2004, 48, 2–11. [Google Scholar]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, J.J.; Gerdes, S.; Olsen, G.J.; Olson, R.; Pusch, G.D.; Shukla, M.; Vonstein, V.; Wattam, A.R.; Yoo, H. PATtyFams: Protein families for the microbial genomes in the PATRIC database. Front. Microbiol. 2016, 7, 118. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; den Bakker, H.C.; Li, S.; Chen, J.; Dinsmore, B.A.; Lane, C.; Lauer, A.C.; Fields, P.I.; Deng, X. SeqSero2: Rapid and improved Salmonella serotype determination using whole-genome sequencing data. Appl. Environ. Microbiol. 2019, 85, e01746-19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yin, Y.; Jones, M.B.; Zhang, Z.; Kaiser, B.L.D.; Dinsmore, B.A.; Fitzgerald, C.; Fields, P.I.; Deng, X. Salmonella serotype determination utilizing high-throughput genome sequencing data. J. Clin. Microbiol. 2015, 53, 1685–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roer, L.; Hendriksen, R.S.; Leekitcharoenphon, P.; Lukjancenko, O.; Kaas, S.; Hasman, H. Is the Evolution of Salmonella enterica subsp. enterica Linked to Restriction-Modification Systems? mSystems 2016, 1, e00009-16. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Genetics 2000, 156, 1997–2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carver, T.; Thomson, N.; Bleasby, A.; Berriman, M.; Parkhill, J. DNAPlotter: Circular and linear interactive genome visualization. Bioinformatics 2009, 25, 119–120. [Google Scholar] [CrossRef] [Green Version]
- Vihinen, M. When a Synonymous Variant Is Nonsynonymous. Genes 2022, 13, 1485. [Google Scholar] [CrossRef] [PubMed]
- Boutet, E.; Lieberherr, D.; Tognolli, M.; Schneider, M.; Bansal, P.; Bridge, A.J.; Poux, S.; Bougueleret, L.; Xenarios, I. UniProtKB/Swiss-Prot, the Manually Annotated Section of the UniProt KnowledgeBase: How to Use the Entry View. In Plant Bioinformatics, Methods and Protocols, Methods in Molecular Biology; Edwards, D., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; Volume 1374, pp. 23–54. ISBN 9781493931675. [Google Scholar]
- Wang, L.; Liu, D.; Reeves, P.R. C-terminal half of Salmonella enterica WbaP (RfbP) is the galactosyl-1-phosphate transferase domain catalyzing the first step of O-antigen synthesis. J. Bacteriol. 1996, 178, 2598–2604. [Google Scholar] [CrossRef] [Green Version]
- Mcquiston, J.R.; Parrenas, R.; Gheesling, L.; Brenner, F.; Fields, P.I. Sequencing and Comparative Analysis of Flagellin Genes fliC, fljB, and flpA from Salmonella. J. Clin. Microbiol. 2004, 42, 1923–1932. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Knirel, Y.A.; Feng, L.; Perepelov, A.V.; Senchenkova, S.N.; Reeves, P.R.; Wang, L. Structural diversity in Salmonella O antigens and its genetic basis. Fed. Eur. Microbiol. Soc. Microbiol Rev. 2014, 38, 56–89. [Google Scholar]
- European, T.; One, U.; Report, Z. The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, e06971. [Google Scholar]
- Campos, J.; Mourão, J.; Peixe, L.; Antunes, P. Non-typhoidal salmonella in the pig production chain: A comprehensive analysis of its impact on human health. Pathogens 2019, 8, 19. [Google Scholar] [CrossRef] [Green Version]
- Public Health England. UK Standards for Microbiology Investigations. Bacteriology 2015, B55, 1–21. [Google Scholar]
- Patel, K.B.; Furlong, S.E.; Valvano, M.A. Functional analysis of the C-terminal domain of the WbaP protein that mediates initiation of O antigen synthesis in Salmonella enterica. Glycobiology 2010, 20, 1389–1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, Q.; Yang, J.; Liu, Q.; Alamuri, P.; Roland, K.L.; Curtiss, R. Effect of deletion of genes involved in lipopolysaccharide core and O-antigen synthesis on virulence and immunogenicity of Salmonella enterica serovar Typhimurium. Infect. Immun. 2011, 79, 4227–4239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, E.; Azriel, S.; Austeri, O.; Gal, A.; Zitronblat, C.; Mikhlin, S.; Scharte, F.; Hensei, M.; Rahav, G.; Mor, O.G. Pathoadaptation of the passerine-associated Salmonella enterica serovar Typhimurium lineage to the avian host. PLoS Pathog. 2021, 17, e1009451. [Google Scholar] [CrossRef] [PubMed]
- Barak, R.; Eisenbach, M. Fumarate or a fumarate metabolite restores switching ability to rotating flagella of bacterial envelopes. J. Bacteriol. 1992, 174, 643–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Zhao, H.; Sun, J.; Liu, Y.; Zhou, X.; Beier, R.C.; Wu, G.; Hou, X. Characterization of Multidrug-Resistant Salmonella enterica Serovars Indiana and Enteritidis from Chickens in Eastern China. PLoS ONE 2014, 9, e96050. [Google Scholar] [CrossRef] [PubMed]
- Hong, R.W.; Shchepetov, M.; Weiser, J.N.; Axelsen, P.H. Transcriptional profile of the Escherichia coli response to the antimicrobial insect peptide cecropin A. Antimicrob. Agents Chemother. 2003, 47, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, M.; Goto, R.; Haneda, T.; Okada, N.; Miki, T. Salmonella enterica Serovar Typhimurium CpxRA Two-Component System Contributes to Gut Colonization in Salmonella-Induced Colitis. Infect. Immun. 2018, 86, e00280-18. [Google Scholar] [CrossRef] [Green Version]
- Kingsley, R.A.; Humphries, A.D.; Weening, E.H.; De Zoete, M.R.; Winter, S.; Papaconstantinopoulou, A.; Dougan, G.; Ba, A.J. Molecular and Phenotypic Analysis of the CS54 Island of. Infect. Immun. 2003, 71, 629–640. [Google Scholar] [CrossRef] [Green Version]
- Urrutia, I.M.; Fuentes, J.A.; Valenzuela, L.M.; Ortega, A.P.; Hidalgo, A.A.; Mora, G.C. Salmonella Typhi shdA: Pseudogene or allelic variant? Infect. Genet. Evol. 2014, 26, 146–152. [Google Scholar] [CrossRef]
- Gonzalez, C.F.; Proudfoot, M.; Brown, G.; Korniyenko, Y.; Mori, H.; Savchenko, A.V.; Yakunin, A.F. Molecular basis of formaldehyde detoxification: Characterization of two S-formylglutathione hydrolases from Escherichia coli, FrmB and YeiG. J. Biol. Chem. 2006, 281, 14514–14522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, N.H.; Djoko, K.Y.; Veyrier, F.J.; McEwan, A.G. Formaldehyde stress responses in bacterial pathogens. Front. Microbiol. 2016, 7, 257. [Google Scholar] [CrossRef] [Green Version]
- Kanehara, K.; Ito, K.; Akiyama, Y. YaeL (EcfE) activates the σE pathway of stress response through a site-2 cleavage of anti-σE, RseA. Genes Dev. 2002, 16, 2147–2155. [Google Scholar] [CrossRef] [Green Version]
- Mandal, R.K.; Kwon, Y.M. Global screening of Salmonella enterica serovar Typhimurium genes for desiccation survival. Front. Microbiol. 2017, 8, 1723. [Google Scholar] [CrossRef] [PubMed]
- Guiney, D.G.; Fierer, J. The role of the spv genes in Salmonella pathogenesis. Front. Microbiol. 2011, 2, 129. [Google Scholar] [CrossRef] [Green Version]
- Mansour, M.N.; Yaghi, J.; El Khoury, A.; Felten, A.; Mistou, M.Y.; Atoui, A.; Radomski, N. Prediction of Salmonella serovars isolated from clinical and food matrices in Lebanon and genomic-based investigation focusing on Enteritidis serovar. Int. J. Food Microbiol. 2020, 333, 108831. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Singh, P.; Ranieri, M.L.; Wiedmann, M.; Moreno Switt, A.I. Molecular methods for serovar determination of Salmonella. Crit. Rev. Microbiol. 2015, 41, 309–325. [Google Scholar] [CrossRef] [PubMed]
- EN ISO 6579-1:2017; Microbiology of the food chain—Horizontal method for the detection, enumeration and serotyping of Salmonella—Part 1: Detection of Salmonella spp. International Organization for Standardization: Geneva, Switzerland, 2017.
- Pławińska-Czarnak, J.; Wódz, K.; Kizerwetter-Świda, M.; Bogdan, J.; Kwieciński, P.; Nowak, T.; Strzałkowska, Z.; Anusz, K. Multi-Drug Resistance to Salmonella spp. When Isolated from Raw Meat Products. Antibiotics 2022, 11, 876. [Google Scholar] [CrossRef] [PubMed]
- Grimont, P.A.D.; Weill, F.-X. Antigenic formulae of the Salmonella serovars. WHO Collab. Cent. Ref. Res. Salmonella 2007, 9, 1–167. [Google Scholar]
- Issenhuth-Jeanjean, S.; Roggentin, P.; Mikoleit, M.; Guibourdenche, M.; De Pinna, E.; Nair, S.; Fields, P.I.; Weill, F.X. Supplement 2008–2010 (no. 48) to the White-Kauffmann-Le Minor scheme. Res. Microbiol. 2014, 165, 526–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wingett, S.W.; Andrews, S. Fastq screen: A tool for multi-genome mapping and quality control. [version 2; peer review: 4 approved]. F1000Research 2018, 7, 1338. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Schwengers, O.; Jelonek, L.; Dieckmann, M.A.; Beyvers, S.; Blom, J.; Goesmann, A. Bakta: Rapid and standardized annotation of bacterial genomes via alignment-free sequence identification. Microb. Genom. 2021, 7, 000685. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [Green Version]
- Ando, H.; Kondo, Y.; Suetake, T.; Toyota, E.; Kato, S.; Mori, T.; Kirikae, T. Identification of katG mutations associated with high-level isoniazid resistance in Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2010, 54, 1793–1799. [Google Scholar] [CrossRef] [Green Version]
- Loewen, P.C.; De Silva, P.M.; Donald, L.J.; Switala, J.; Villanueva, J.; Fita, I.; Kumar, A. KatG-Mediated Oxidation Leading to Reduced Susceptibility of Bacteria to Kanamycin. ACS Omega 2018, 3, 4213–4219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holden, E.R.; Webber, M.A. MarA, RamA, and SoxS as Mediators of the Stress Response: Survival at a Cost. Front. Microbiol. 2020, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Haycocks, J.R.J.; Middlemiss, A.D.; Kettles, R.A.; Sellars, L.E.; Ricci, V.; Piddock, L.J.V.; Grainger, D.C. The multiple antibiotic resistance operon of enteric bacteria controls DNA repair and outer membrane integrity. Nat. Commun. 2017, 8, 1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eaves, D.J.; Ricci, V.; Piddock, L.J. V soxS in Salmonella enterica Serovar Typhimurium: Role in Multiple Antibiotic Resistance Serovar Typhimurium: Role in Multiple Antibiotic Resistance. Antimicrob. Agent Chemother. 2004, 48, 1145–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galakatos, N.G.; Daub, E.; Botstein, D.; Walsh, C.T. Biosynthetic alr alanine racemase from Salmonella typhimurium: DNA and protein sequence determination. Biochemistry. 1986, 25, 3255–3260. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.; Das, S.; Panda, P.K.; Suar, M. Identification of a new alanine racemase in Salmonella Enteritidis and its contribution to pathogenesis. Gut Pathog. 2018, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Zawadzke, L.E.; Bugg, T.D.H.; Walsh, C.T. Existence of Two D-Alanine:D-Alanine Ligases in Escherichia coli: Cloning and Sequencing of the ddlA Gene and Purification and Characterization of the DdlA and DdlB Enzymes. Biochemistry 1991, 30, 1673–1682. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.C.; Parish, T. Dxr is essential in Mycobacterium tuberculosis and fosmidomycin resistance is due to a lack of uptake. BMC Microbiol. 2008, 8, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macvanin, M.; Björkman, J.; Eriksson, S.; Rhen, M.; Andersson, D.I.; Hughes, D. Fusidic Acid-Resistant Mutants of Salmonella enterica Serovar Typhimurium with Low Fitness In Vivo are Defective in RpoS Induction. Antimicrob. Agents Chemother. 2003, 47, 3743–3749. [Google Scholar] [CrossRef] [Green Version]
- Harvey, K.L.; Jarocki, V.M.; Charles, I.G.; Djordjevic, S.P. The diverse functional roles of elongation factor tu (Eftu) in microbial pathogenesis. Front. Microbiol. 2019, 10, 2351. [Google Scholar] [CrossRef] [PubMed]
- Manna, M.S.; Tamer, Y.T.; Gaszek, I.; Poulides, N.; Ahmed, A.; Wang, X.; Toprak, F.C.R.; Woodard, D.N.R.; Koh, Y.; Williams, N.S.; et al. A trimethoprim derivative impedes antibiotic resistance evolution. Nat. Commun. 2021, 12, 2949. [Google Scholar] [CrossRef] [PubMed]
- Vedantam, G.; Guay, G.G.; Austria, N.E.; Doktor, S.Z.; Nichols, B.P. Characterization of mutations contributing to sulfathiazole resistance in Escherichia coli. Antimicrob. Agents Chemother. 1998, 42, 88–93. [Google Scholar] [CrossRef] [Green Version]
- Weigel, L.M.; Steward, C.D.; Tenover, F.C. gyrA mutations associated with fluoroquinolone resistance in eight species of Enterobacteriaceae. Antimicrob. Agents Chemother. 1998, 42, 2661–2667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.; Zhang, Z.; Li, X.; Song, Y.; Kang, J.; Yin, D.; Gao, Y.; Shi, N.; Duan, J. Mutations in gyrB play an important role in ciprofloxacin-resistant pseudomonas aeruginosa. Infect. Drug Resist. 2019, 12, 261–272. [Google Scholar] [CrossRef] [Green Version]
- Webber, M.A.; Randall, L.P.; Cooles, S.; Woodward, M.J.; Piddock, L.J.V. Triclosan resistance in Salmonella enterica serovar Typhimurium. J. Antimicrob. Chemother. 2008, 62, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Takahata, S.; Ida, T.; Hiraishi, T.; Sakakibara, S.; Maebashi, K.; Terada, S.; Muratani, T.; Matsumoto, T.; Nakahama, C.; Tomono, K. Molecular mechanisms of fosfomycin resistance in clinical isolates of Escherichia coli. Int. J. Antimicrob. Agents 2010, 35, 333–337. [Google Scholar] [CrossRef]
- Hafeezunnisa, M.; Sen, R. The Rho-Dependent Transcription Termination Is Involved in Broad-Spectrum Antibiotic Susceptibility in Escherichia coli. Front. Microbiol. 2020, 11, 605305. [Google Scholar] [CrossRef]
- Siu, G.K.H.; Zhang, Y.; Lau, T.C.K.; Lau, R.W.T.; Ho, P.L.; Yew, W.W.; Tsui, S.K.W.; Cheng, V.C.C.; Yuen, K.Y.; Yam, W.C. Mutations outside the rifampicin resistance-determining region associated with rifampicin resistance in Mycobacterium tuberculosis. J. Antimicrob. Chemother. 2011, 66, 730–733. [Google Scholar] [CrossRef]
- De Vos, M.; Müller, B.; Borrell, S.; Black, P.A.; Van Helden, P.D.; Warren, R.M.; Gagneux, S.; Victor, T.C. Putative compensatory mutations in the rpoc gene of rifampin-resistant mycobacterium tuberculosis are associated with ongoing transmission. Antimicrob. Agents Chemother. 2013, 57, 827–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radeck, J.; Gebhard, S.; Orchard, P.S.; Kirchner, M.; Bauer, S.; Mascher, T.; Fritz, G. Anatomy of the bacitracin resistance network in Bacillus subtilis. Mol. Microbiol. 2016, 100, 607–620. [Google Scholar] [CrossRef] [Green Version]
- Du, D.; Wang, Z.; James, N.R.; Voss, J.E.; Klimont, E.; Ohene-agyei, T.; Venter, H.; Chiu, W.; Luisi, B.F. Structure of the AcrAB—TolC multidrug efflux pump. Nature 2014, 509, 512–515. [Google Scholar] [CrossRef] [Green Version]
- Atac, N.; Kurt-Azap, O.; Dolapci, I.; Yesilkaya, A.; Ergonul, O.; Gonen, M.; Can, F. The Role of AcrAB–TolC Efflux Pumps on Quinolone Resistance of E. coli ST131. Curr. Microbiol. 2018, 75, 1661–1666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Wen, Y.; Hu, G.; Liu, Y.; Beier, R.C.; Hou, X. Genomic sequence analysis of the multidrug-resistance region of avian Salmonella enterica serovar Indiana strain MHYL. Microorganisms 2019, 7, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alav, I.; Bavro, V.N.; Blair, J.M.A. Interchangeability of periplasmic adaptor proteins AcrA and AcrE in forming functional efflux pumps with AcrD in Salmonella enterica serovar Typhimurium. J. Antimicrob. Chemother. 2021, 76, 2558–2564. [Google Scholar] [CrossRef] [PubMed]
- Alenazy, R. Antibiotic resistance in Salmonella: Targeting multidrug resistance by understanding efflux pumps, regulators and the inhibitors. J. King Saud Univ.-Sci. 2022, 34, 102275. [Google Scholar] [CrossRef]
- Yardeni, E.H.; Zomot, E.; Bibi, E. The fascinating but mysterious mechanistic aspects of multidrug transport by MdfA from Escherichia coli. Res. Microbiol. 2018, 169, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Nilsen, I.W.; Bakke, I.; Vader, A.; Olsvik, Ø.; El-Gewely, M.R. Isolation of cmr, a novel Escherichia coli chloramphenicol resistance gene encoding a putative efflux pump. J. Bacteriol. 1996, 178, 3188–3193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayhan, D.H.; Tamer, Y.T.; Akbar, M.; Bailey, S.M.; Wong, M.; Daly, S.M.; Greenberg, D.E.; Toprak, E. Sequence-Specific Targeting of Bacterial Resistance Genes Increases Antibiotic Efficacy. PLoS Biol. 2016, 14, e1002552. [Google Scholar] [CrossRef] [Green Version]
- Nishino, K.; Nikaido, E.; Yamaguchi, A. Regulation of multidrug efflux systems involved in multidrug and metal resistance of Salmonella enterica serovar typhimurium. J. Bacteriol. 2007, 189, 9066–9075. [Google Scholar] [CrossRef] [Green Version]
- Jibril, A.H.; Okeke, I.N.; Dalsgaard, A.; Menéndez, V.G.; Olsen, J.E. Genomic analysis of antimicrobial resistance and resistance plasmids in salmonella serovars from poultry in Nigeria. Antibiotics 2021, 10, 99. [Google Scholar] [CrossRef]
- Holdsworth, S.R.; Law, C.J. Functional and biochemical characterisation of the Escherichia coli major facilitator superfamily multidrug transporter MdtM. Biochimie 2012, 94, 1334–1346. [Google Scholar] [CrossRef] [PubMed]
- Spierings, G.; Elders, R.; van Lith, B.; Hofstra, H.; Tommassen, J. Characterization of the Salmonella typhimurium phoE gene and development of Salmonella-specific DNA probes. Gene 1992, 122, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Nishino, K.; Latifi, T.; Groisman, E.A. Virulence and drug resistance roles of multidrug efflux systems of Salmonella enterica serovar Typhimurium. Mol. Microbiol. 2006, 59, 126–141. [Google Scholar] [CrossRef]
- Kapach, G.; Nuri, R.; Schmidt, C.; Danin, A.; Ferrera, S.; Savidor, A.; Gerlach, R.G.; Shai, Y. Loss of the Periplasmic Chaperone Skp and Mutations in the Efflux Pump AcrAB-TolC Play a Role in Acquired Resistance to Antimicrobial Peptides in Salmonella typhimurium. Front. Microbiol. 2020, 11, 189. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-García, Á.; Mares-Alejandre, R.E.; Muñoz-Muñoz, P.L.A.; Ruvalcaba-Ruiz, S.; González-Sánchez, R.A.; Bernáldez-Sarabia, J.; Meléndez-López, S.G.; Licea-Navarro, A.F.; Ramos-Ibarra, M.A. Molecular analysis of streptomycin resistance genes in clinical strains of mycobacterium tuberculosis and biocomputational analysis of the mtgidb l101f variant. Antibiotics 2021, 10, 807. [Google Scholar] [CrossRef]
- Sulaiman, J.E.; Long, L.; Wu, L.; Qian, P.Y.; Lam, H. Comparative proteomic investigation of multiple methicillin-resistant Staphylococcus aureus strains generated through adaptive laboratory evolution. iScience 2021, 24, 102950. [Google Scholar] [CrossRef]
- Yang, B.; Yao, H.; Li, D.; Liu, Z. The phosphatidylglycerol phosphate synthase PgsA utilizes a trifurcated amphipathic cavity for catalysis at the membrane-cytosol interface. Curr. Res. Struct. Biol. 2021, 3, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.T.; Mishra, N.N.; Seepersaud, R.; Diaz, L.; Rios, R.; Dinh, A.Q.; Garcia-de-la-Maria, C.; Rybak, M.J.; Miro, J.M.; Shelburne, S.A.; et al. Mutations in cdsA and pgsA Correlate with Daptomycin Resistance in Streptococcus mitis and S. oralis. Antimicrob. Agents Chemother. 2019, 63, e01531-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalafatis, M.; Slauch, J.M. Long-distance effects of H-NS binding in the control of hilD expression in the salmonella SPI1 locus. J. Bacteriol. 2021, 203, e0030821. [Google Scholar] [CrossRef]
- Johnson, J.R.; Clabots, C.; Rosen, H. Effect of inactivation of the global oxidative stress regulator oxyR on the colonization ability of Escherichia coli O1:K1:H7 in a mouse model of ascending urinary tract infection. Infect. Immun. 2006, 74, 461–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anand, A.; Chen, K.; Catoiu, E.; Sastry, A.V.; Olson, C.A.; Sandberg, T.E.; Seif, Y.; Xu, S.; Szubin, R.; Yang, L.; et al. OxyR Is a Convergent Target for Mutations Acquired during Adaptation to Oxidative Stress-Prone Metabolic States. Mol. Biol. Evol. 2020, 37, 660–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robicsek, A.; Strahilevitz, J.; Jacoby, G.A.; Macielag, M.; Abbanat, D.; Chi, H.P.; Bush, K.; Hooper, D.C. Fluoroquinolone-modifying enzyme: A new adaptation of a common aminoglycoside acetyltransferase. Nat. Med. 2006, 12, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.H.Y.; Chan, E.W.C.; Liu, L.Z.; Chen, S. PMQR genes oqxAB and aac(6′)Ib-cr accelerate the development of fluoroquinolone resistance in Salmonella Typhimurium. Front. Microbiol. 2014, 5, 521. [Google Scholar] [CrossRef]
- Nishino, K.; Yamaguchi, A. Role of Histone-Like Protein H-NS in Multidrug Resistance of Escherichia coli. J. Bacteriol. 2004, 186, 1423–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SPI | Identity % | Query/Template Length | Position in Contig | Genes | Organism | Accession Number |
|---|---|---|---|---|---|---|
| C63PI | 100 | 4000/4000 | 3,006,066..3,010,065 | sitA, sitB, sitC, sitD | Salmonella-enterica-Typhimurium-SL1344 | AF128999 |
| CS54_island | 99.92 | 25255/25252 | 2,627,012..2,652,263 | xse, shdA, ratC, ratB, ratA, sinI, sinH, yfgK | Salmonella-enterica-Typhimurium-ATCC_14028 | AF140550 |
| SPI-1 | 99.96 | 2705/2705 | 3,010,641..3,013,345 | sprA, sprB | Salmonella-enterica-Typhimurium-SL1344 | AF148689 |
| SPI-1 | 99.77 | 440/440 | 3,015,995..3,016,434 | prgH, prgI, prgJ | Salmonella-enterica-Gallinarum-SGB_1 | AY956822 |
| SPI-1 | 100 | 430/430 | 3,018,105..3,018,534 | hilD | Salmonella-enterica-Gallinarum-SGE_2 | AY956823 |
| SPI-1 | 100 | 470/470 | 3,026,813..3,027,282 | sipD | Salmonella-enterica-Gallinarum-SGB_4 | AY956824 |
| SPI-1 | 99.28 | 415/415 | 3,038,600..3,039,014 | invA | Salmonella-enterica-Gallinarum-SGB_8 | AY956825 |
| SPI-1 | 98.84 | 259/257 | 3,039,894..3,040,152 | invA | Salmonella-enterica-Typhimurium-J4STEHO | JN982040 |
| SPI-1 | 99.43 | 3155/3141 | 3,049,734..3,052,886 | mutS | Salmonella-enterica-Typhimurium-SL1344 | U16303 |
| SPI-12 | 97.64 | 9345/11075 | 2,342,056..2,351,393 | sspH2, isrG, citB, ccmD, dsbE1, nrfE, ccmE1 | Salmonella-enterica-Choleraesuis-SC_B67 | NC_006905 |
| SPI-13 | 99.41 | 338/338 | 3,279,563..3,279,900 | gacD | Salmonella-enterica-Gallinarum-SGD_3 | AY956832 |
| SPI-13 | 100 | 404/404 | 3,278,852..3,279,255 | gtrA | Salmonella-enterica-Gallinarum-SGG_1 | AY956833 |
| SPI-13 | 100 | 341/341 | 3,277,144..3,277,484 | gtrB | Salmonella-enterica-Gallinarum-SGA_10 | AY956834 |
| SPI-14 | 100 | 501/501 | 926,728..927,228 | gpiA | Salmonella-enterica-Gallinarum-SGA_8 | AY956835 |
| SPI-14 | 99.55 | 441/441 | 32,304..932,744 | gpiB | Salmonella-enterica-Gallinarum-SGC_8 | AY956836 |
| SPI-2 | 98.90 | 637/637 | 1,497,019..1,497,655 | ssaO, ssaP | Salmonella-enterica-Gallinarum-SGB_10 | AY956826 |
| SPI-2 | 100 | 642/642 | 1,489,709…1,490,350 | ssaG, ssaH, ssaI, ssaJ | Salmonella-enterica-Gallinarum-SGC_2 | AY956827 |
| SPI-2 | 100 | 395/396 | 1,494342..1,494,736 | ssaV | Salmonella-enterica-Gallinarum-SGC_9 | AY956828 |
| SPI-2 | 99.76 | 425/425 | 1,487,684..1,488,108 | sscB, sseF | Salmonella-enterica-Gallinarum-SGH_1 | AY956829 |
| SPI-2 | 99.82 | 547/547 | 1,477,547..1,478,093 | ssrA | Salmonella-enterica-Gallinarum-SGD_8 | AY956830 |
| SPI-2 | 100 | 384/384 | 1,480,008..1,480,391 | spiC | Salmonella-enterica-Typhimurium-St11 | JN673272 |
| SPI-2 | 100 | 1252/1252 | 1,475,967..1,477,218 | ssrA, ssrB | Salmonella-enterica-Typhimurium | Z95891 |
| SPI-3 | 100 | 738/738 | 3,965,704..3,966,441 | mgtC | Salmonella-enterica-Typhimurium-14028s | AJ000509 |
| SPI-3 | 100 | 1514/1514 | 3,948,169..3,949,682 | selC | Salmonella-enterica-Typhimurium-14028s | Y13864 |
| SPI-4 | 100 | 24660/24660 | 4,476,929..4,501,588 | siiABCDEF | Salmonella-enterica-Typhimurium-ST4/74 | AJ576316 |
| SPI-5 | 100 | 9069/9069 | 1,175,309..1,184,377 | Ser_trna, pipA, pipB, isrI, sigE, sopB, pepD, kdpD, ompR | Salmonella-Typhimurium-LT2 | NC_003197 |
| SPI-9 | 98.58 | 12647/15696 | 2,831,233..2,843,879 | bapA, isrL, tolC, sunT, emrA | Salmonella-Typhi-CT18 | NC_003198 |
| Pos | Ref | Var | Type | Ref_nt | Var_nt | Ref_nt_pos_change | Ref_aa_pos_change | Locus Tag | Gene Name | Function | snpEff Type | snpEff Impact |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
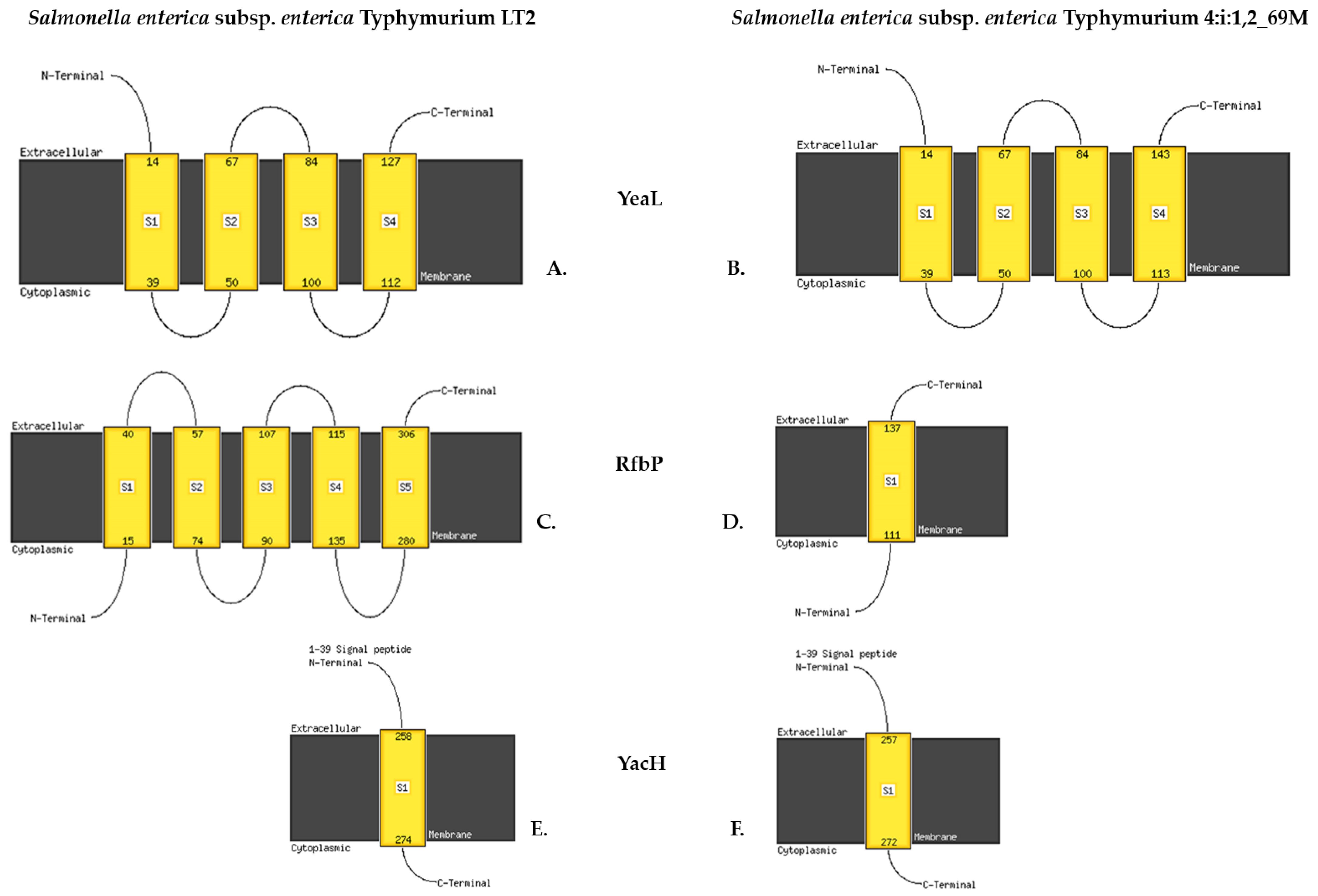
| 1357016 | CGC | CGGC | Insertion | gtgcgg | gtGCCGg | 390_391insC | Val130_Arg131fs | STM1280 | yeaL | UPF0756 membrane protein YeaL | Frameshift variant | HIGH |
| 1543816 | GGC | GCGC | Insertion | ggcgct | GCGCgct | 1631_1632insC | Gly544_Ala545fs | STM1468 | fumA | Fumarate hydratase class I, aerobic (EC 4.2.1.2) | Frameshift variant | HIGH |
| 1794104 | A | G | Nonsyn | tga | tgG | 981A>G | Ter327Trpext *? | STM1701 | yciW | Uncharacterized protein YciW | stop_lost | HIGH |
| 184611 | GTGATGATCG | GG | Deletion | gacgatcatcac | gaCC | 850_857delGATCATCA | Asp284fs | STM0157 | yacH | Uncharacterized protein YacH | Frameshift variant | HIGH |
| 2162286 | TAAAAAAAATCAA | TAAAAAAAAATCAA | Insertion | ttgattttttttaat | TTGATTTTTTTTTAat | 67_68insT | Ile23_Phe24fs | STM2082 | rfbP | Undecaprenyl-phosphate galactosephosphotransferase (EC 2.7.8.6) | Frameshift variant | HIGH |
| 2292159 | CGC | CGGC | Insertion | cgcaaa | CGGCaaa | 720_721insG | Arg240_Lys241fs | STM2194 | yeiG | S-formylglutathione hydrolase (EC 3.1.2.12) | Frameshift variant | HIGH |
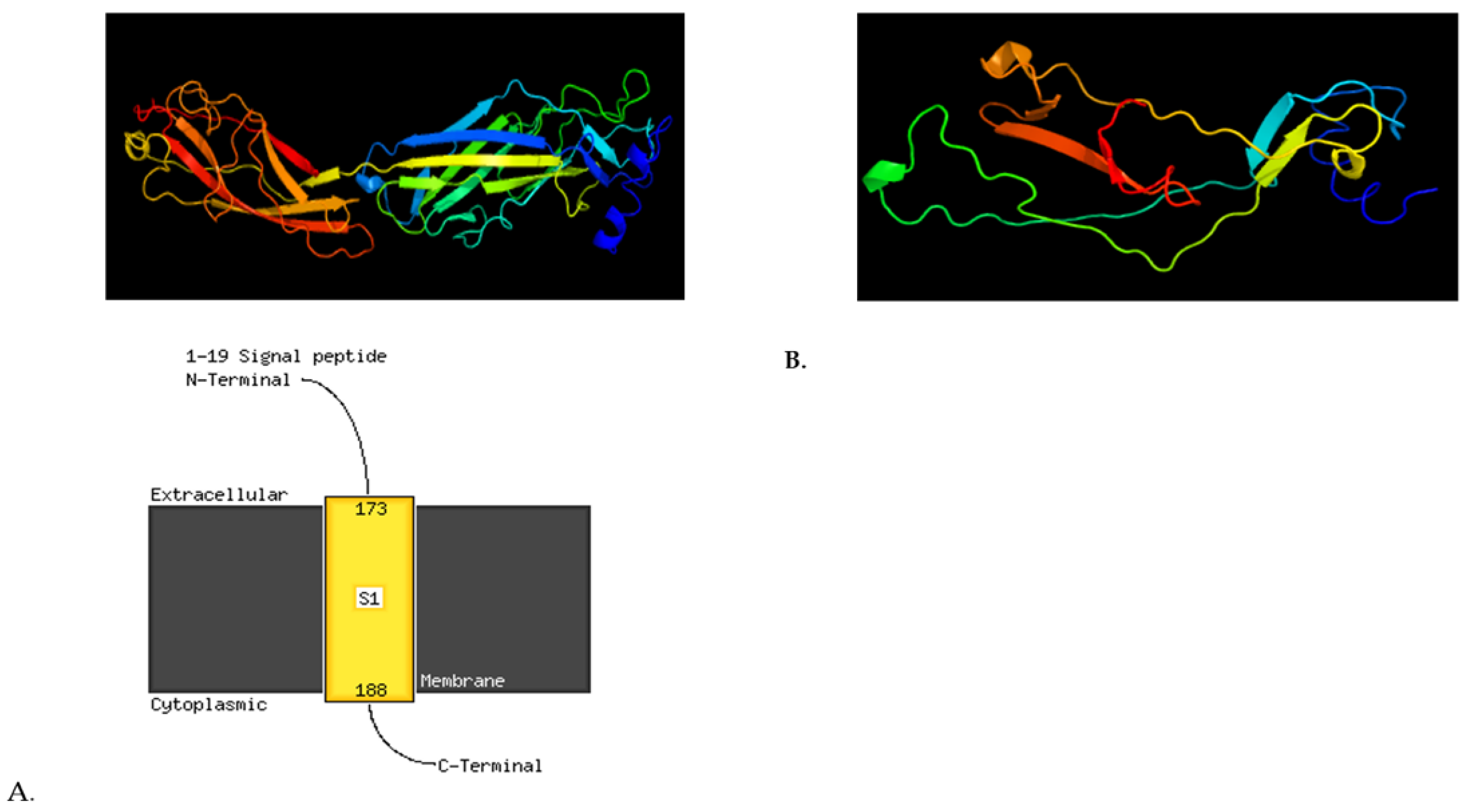
| 2633672 | G | A | Nonsyn | cga | Tga | 28C>T | Arg10 * | STM2513 | shdA | AIDA autotransporter-like protein | stop_gained | HIGH |
| 2635902 | CAT | CT | Deletion | atg | AG | 5807delT | Met1936fs | STM2514 | ratB | Putative outer membrane protein | Frameshift variant | HIGH |
| 3010007 | GTTTCA | GTTCA | Deletion | aatgaaacg | aaTGAACg | 755delA | Glu252fs | STM2865 | avrA | Type III secretion injected virulence protein (YopP, YopJ, induces apoptosis, prevents cytokine induction, inhibits NFkb activation) | Frameshift variant | HIGH |
| 3823687 | TACATTCTCAAA | TA | Deletion | tttgagaatgta | TA | 400_409delTTTGAGAATG | Phe134fs | STM3637 | lpfD | Protein LpfD | Frameshift variant | HIGH |
| 720479 | TGGGGGAT | TGGGGGGAT | Insertion | tgggggatt | TGGGGGGATt | 139_140insG | Ile47_Asp48fs | STM0657 | ybeU | hypothetical protein | Frameshift variant | HIGH |
| 757823 | G | A | Nonsyn | cag | Tag | 280C>T | Gln94 * | STM0695 | ybfE | Uncharacterized protein YbfE | stop_gained | HIGH |
| AMR Mechanism | Genes |
|---|---|
| Antibiotic activation enzyme | KatG |
| Antibiotic inactivation enzyme | AAC(6’)-Ic,f,g,h,j,k,l,r-z |
| Antibiotic resistance gene cluster, cassette, or operon | MarA, MarB, MarR |
| Antibiotic target in susceptible species | Alr, Ddl, dxr, EF-G, EF-Tu, folA, Dfr, folP, gyrA, gyrB, inhA, fabI, Iso-tRNA, kasA, MurA, rho, rpoB, rpoC, S10p, S12p |
| Antibiotic target protection protein | BcrC |
| Efflux pump conferring antibiotic resistance | AcrAB-TolC, AcrAD-TolC, AcrEF-TolC, AcrZ, EmrAB-TolC, MacA, MacB, MdfA/Cmr, MdtABC-TolC, MdtL, MdtM, MexPQ-OpmE, OprM/OprM family, SugE, TolC/OpmH |
| Gene conferring resistance via absence | gidB |
| Protein altering cell wall charge conferring antibiotic resistance | GdpD, PgsA |
| Regulator modulating expression of antibiotic resistance genes | AcrAB-TolC, EmrAB-TolC, H-NS, OxyR |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pławińska-Czarnak, J.; Wódz, K.; Guzowska, M.; Rosiak, E.; Nowak, T.; Strzałkowska, Z.; Kwieciński, A.; Kwieciński, P.; Anusz, K. Comparison of Phenotype and Genotype Virulence and Antimicrobial Factors of Salmonella Typhimurium Isolated from Human Milk. Int. J. Mol. Sci. 2023, 24, 5135. https://doi.org/10.3390/ijms24065135
Pławińska-Czarnak J, Wódz K, Guzowska M, Rosiak E, Nowak T, Strzałkowska Z, Kwieciński A, Kwieciński P, Anusz K. Comparison of Phenotype and Genotype Virulence and Antimicrobial Factors of Salmonella Typhimurium Isolated from Human Milk. International Journal of Molecular Sciences. 2023; 24(6):5135. https://doi.org/10.3390/ijms24065135
Chicago/Turabian StylePławińska-Czarnak, Joanna, Karolina Wódz, Magdalena Guzowska, Elżbieta Rosiak, Tomasz Nowak, Zuzanna Strzałkowska, Adam Kwieciński, Piotr Kwieciński, and Krzysztof Anusz. 2023. "Comparison of Phenotype and Genotype Virulence and Antimicrobial Factors of Salmonella Typhimurium Isolated from Human Milk" International Journal of Molecular Sciences 24, no. 6: 5135. https://doi.org/10.3390/ijms24065135