
LOX-1 Activation by oxLDL Induces AR and AR-V7 Expression via NF-κB and STAT3 Signaling Pathways Reducing Enzalutamide Cytotoxic Effects
 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. LOX-1 Activation by oxLDLs Increases ROS Generation in CRPC Cells
2.2. OxLDL/LOX-1 Induces the Activation of NF-κB in C4-2B and 22RV1 Cells
2.3. Activation of LOX-1 by oxLDLs Induces the Secretion of IL-6 and STAT3 Activation in C4-2B and 22Rv1 Cells
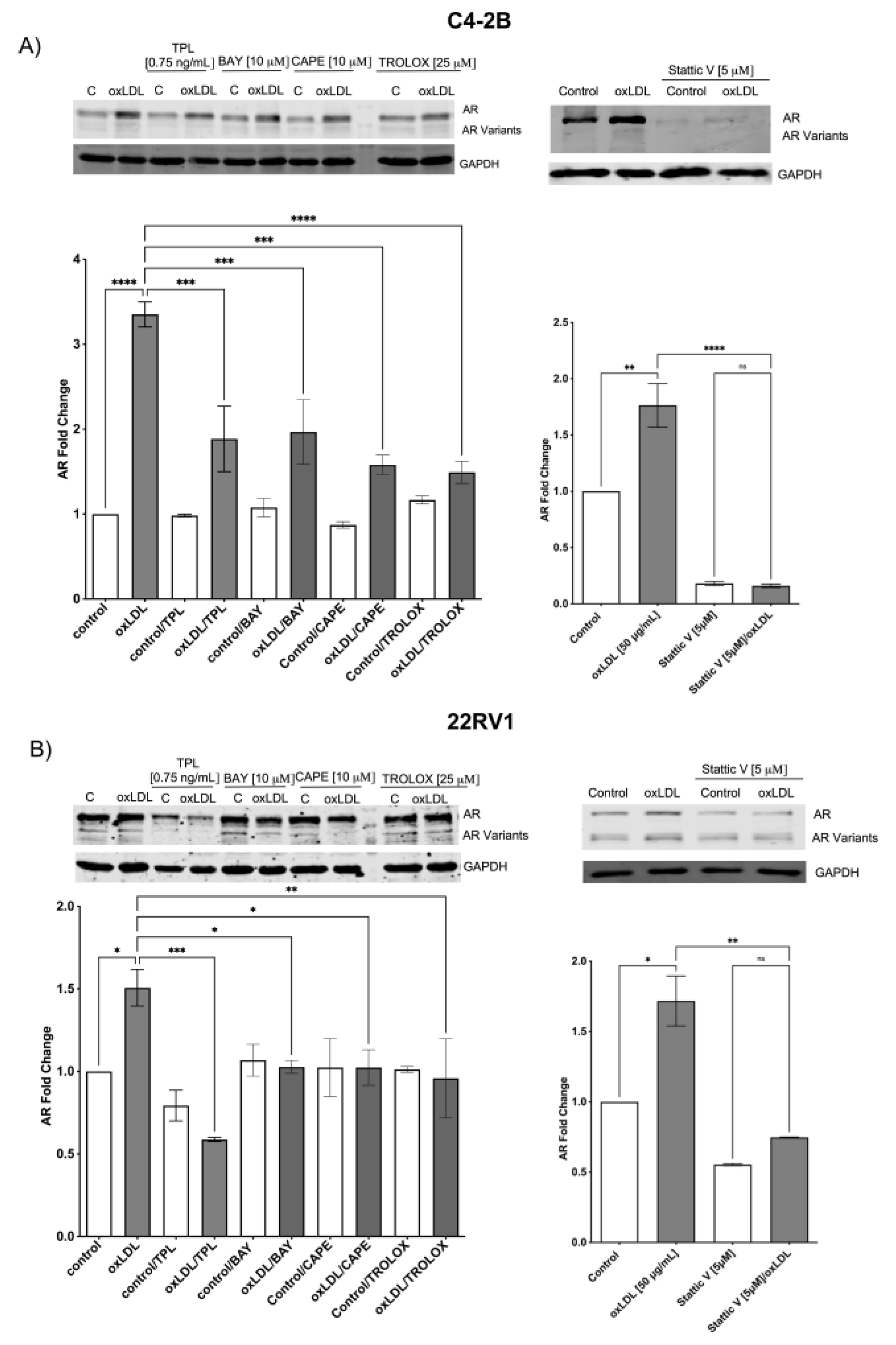
2.4. LOX-1/oxLDL Induces AR and AR-V7 Overexpression in C4-2B and 22RV1 Cells
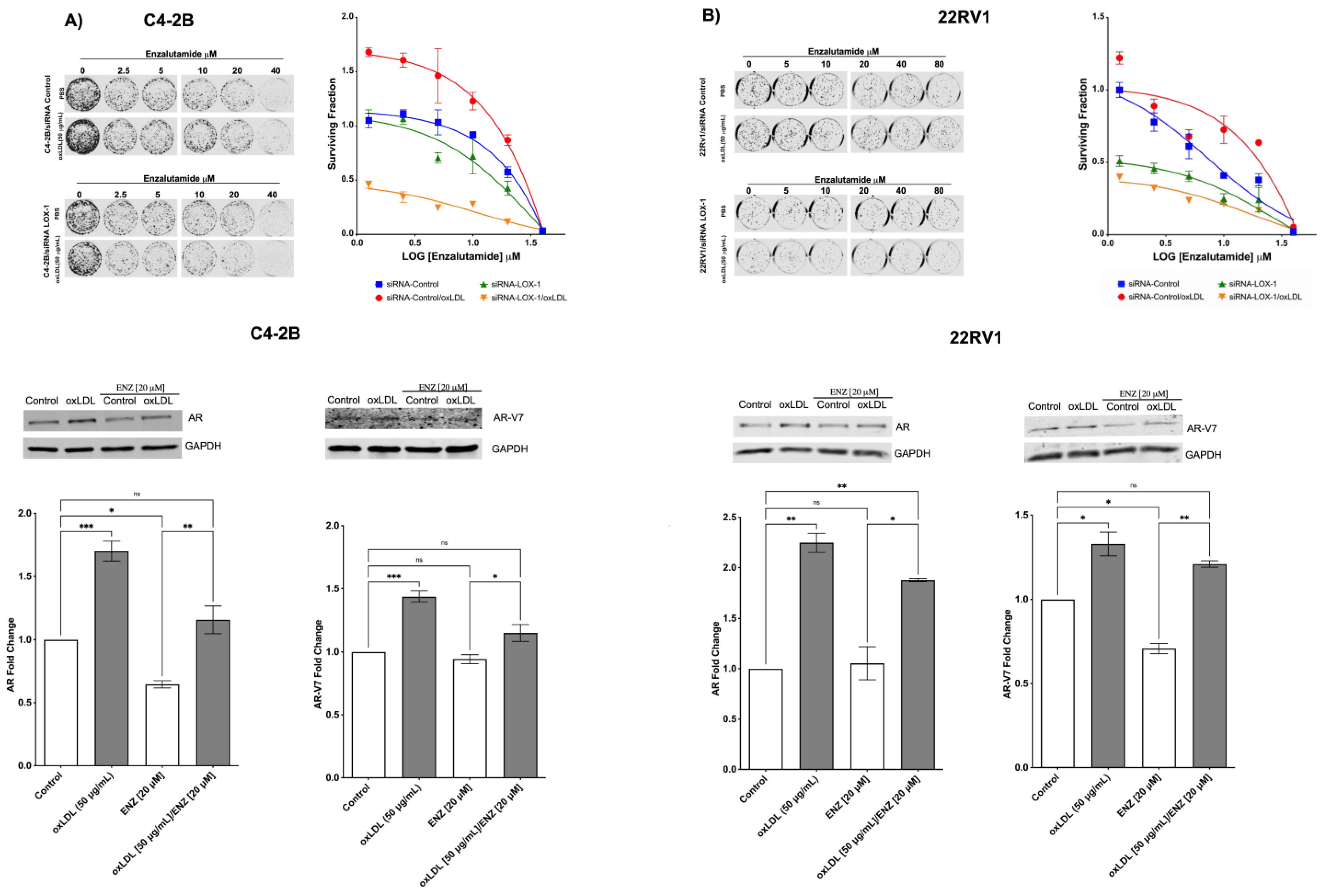
2.5. LOX-1 Activation by oxLDLs Decreases the Cytotoxic Effects of Enzalutamide on C4-2B and 22RV1 Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. OxLDL Preparation
4.3. siRNA Transfection
4.4. Immunodetection
4.5. Real Time PCR
4.6. ROS Determination
4.7. Determination of NF-κB and STAT3 Activation
4.8. Cytotoxic Assay Using Enzalutamide
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hoover-Plow, J.; Huang, M. Lipoprotein(a) metabolism: Potential sites for therapeutic targets. Metabolism 2013, 62, 479–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austin, M.A.; Hokanson, J.; Brunzell, J.D. Characterization of low-density lipoprotein subclasses: Methodologic approaches and clinical relevance. Curr. Opin. Lipidol. 1994, 5, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Elshourbagy, N.A.; Meyers, H.; Abdel-Meguid, S. Cholesterol: The good; the bad, and the ugly—Therapeutic targets for the treatment of dyslipidemia. Med. Princ. Pract. 2014, 23, 99–111. [Google Scholar] [CrossRef]
- Koba, S.; Dyslipidemia, T.H.; Rinsho, N. Dyslipidemia and Atherosclerosis. Nihon Rinsho, 2011; 69, 138–143. [Google Scholar]
- Linton, M.R.F.; Yancey, P.G.; Davies, S.S.; Jerome, W.G.; Linton, E.F.; Song, W.L.; Doran, A.C.; Vickers, K.C. The Role of Lipids and Lipoproteins in Atherosclerosis. In Endotext; Feingold, K.R., Anawalt, B., Blackman, M.R., Eds.; Elsevier Health Sciences: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Chistiakov, D.A.; Melnichenko, A.A.; Myasoedova, V.A.; Grechko, A.V.; Orekhov, A.N. Mechanisms of foam cell formation in atherosclerosis. J. Mol. Med. 2017, 95, 1153–1165. [Google Scholar] [CrossRef]
- Tsai, K.L.; Chen, L.H.; Chiou, S.H.; Chiou, G.Y.; Chen, Y.C.; Chou, H.Y.; Chen, L.K.; Chen, H.Y.; Chiu, T.H.; Tsai, C.S.; et al. Coenzyme Q10 suppresses oxLDL-induced endothelial oxidative injuries by the modulation of LOX-1-mediated ROS generation via the AMPK/PKC/NADPH oxidase signaling pathway. Mol. Nutr. Food Res. 2011, 55 (Suppl. S2), S227–S240. [Google Scholar] [CrossRef] [PubMed]
- Hermonat, P.L.; Zhu, H.; Cao, M.; Mehta, J.L. LOX-1 transcription. Cardiovasc. Drugs Ther. 2011, 25, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Kattoor, A.J.; Goel, A.; Mehta, J. LOX-1: Regulation, Signaling and Its Role in Atherosclerosis. Antioxidants 2019, 8, 218. [Google Scholar] [CrossRef] [Green Version]
- Zingg, J.M.; Vlad, A.; Ricciarelli, R. Oxidized LDLs as Signaling Molecules. Antioxidants 2021, 10, 1184. [Google Scholar] [CrossRef]
- Akhmedov, A.; Sawamura, T.; Chen, C.H.; Kraler, S.; Vdovemko, D.; Lüscher, T.F. Lectin-like oxidized low-density lipoprotein receptor-1 (LOX-1): A crucial driver of atherosclerotic cardiovascular disease. Eur. Heart J. 2021, 42, 1797–1807. [Google Scholar] [CrossRef]
- Barreto, J.; Karathanasis, S.K.; Remaley, A.; Sposito, A.C. Role of LOX-1 (Lectin-Like Oxidized Low-Density Lipoprotein Receptor 1) as a Cardiovascular Risk Predictor: Mechanistic Insight and Potential Clinical Use. Arterioscler. Thromb. Vasc. Biol. 2021, 41, 153–166. [Google Scholar] [CrossRef]
- Jin, P.; Cong, S. LOX-1 and atherosclerotic-related diseases. Clin. Chim. Acta 2019, 491, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Kattoor, A.J.; Kanuri, S.; Mehta, J. Role of Ox-LDL and LOX-1 in Atherogenesis. Curr. Med. Chem. 2019, 26, 1693–1700. [Google Scholar] [CrossRef]
- Pirillo, A.; Norata, G.; Catapano, A. LOX-1, OxLDL. Mediat. Inflamation 2013, 2013, 152786. [Google Scholar]
- Sharma, T.; Romeo, F.; Mehta, J. LOX-1: Implications in atherosclerosis and myocardial ischemia. EXCLI J. 2022, 21, 273–278. [Google Scholar]
- Singh, S.; Gautam, A. Upregulated LOX-1 Receptor: Key Player of the Pathogenesis of Atherosclerosis. Curr. Atheroscler. Rep. 2019, 21, 38. [Google Scholar] [CrossRef] [PubMed]
- Madrigal-Ruiz, P.M.; Navarro, R.E.; Ruíz, S.L.; Corona, F.I.; Vázquez, M.; Gómez, E.; Castro, J.; Sandoval, F.; Fores, L.J.; Martín, B.T. Low CD36 and LOX-1 Levels and CD36 Gene Subexpression Are Associated with Metabolic Dysregulation in Older Individuals with Abdominal Obesity. J. Diabetes Res. 2016, 2016, 5678946. [Google Scholar] [CrossRef] [Green Version]
- Santiago-Fernandez, C.; Martín-Reyes, F.; Tome, M.; Gutierrez-Repiso, C.; Fernandez-Garcia, D.; Ocaña-Wilhelmi, L.; Rivas-Becerra, J.; Tatzber, F.; Pursch, E.; Tinahones, F.J.; et al. Oxidized LDL Increase the Proinflammatory Profile of Human Visceral Adipocytes Produced by Hypoxia. Biomedicines 2021, 9, 1715. [Google Scholar] [CrossRef]
- Balzan, S.; Lubrano, V. LOX-1 receptor: A potential link in atherosclerosis and cancer. Life Sci. 2018, 198, 79–86. [Google Scholar] [CrossRef]
- Duan, Q.; Si, H.; Tian, L.; Zhang, N.; Qiu, J.; Yu, J.; Liu, J.; Zhang, Q. LOX-1 attenuates high glucose-induced autophagy via AMPK/HNF4alpha signaling in HLSECs. Heliyon 2022, 8, e12385. [Google Scholar] [CrossRef]
- Deng, C.F.; Zhu, N.; Zhao, T.J.; Li, H.F.; Gu, J.; Liao, D.F.; Qin, L. Involvement of LDL and ox-LDL in Cancer Development and Its Therapeutical Potential. Front. Oncol. 2022, 12, 803473. [Google Scholar] [CrossRef]
- Hao, F.; Chen, J.; Wu, J.; Ge, X.; Lv, X.; Zhang, D.; Chen, J. Expression of Serum sLOX-1 in Patients with Non-Small-Cell Lung Cancer and Its Correlation with Lipid Metabolism. Can. Respir. J. 2022, 2022, 6619331. [Google Scholar] [CrossRef] [PubMed]
- Kindt, N.; Journé, F.; Carlier, S.; Trelcat, A.; Scalia, A.; Saussez, S. Effect of Oxidized Low-Density Lipoprotein on Head and Neck Squamous Cell Carcinomas. Biomedicines 2021, 9, 513. [Google Scholar] [CrossRef]
- Li, C.; Liu, F.; Yang, X.; Guo, B.; Li, G.; Yin, J.; He, G.; Yang, C.; Xu, L.; Li, S.; et al. Targeting lectin-like oxidized low-density lipoprotein receptor-1 triggers autophagic program in esophageal cancer. Cell Death Differ. 2022, 29, 697–708. [Google Scholar] [CrossRef]
- Murdocca, M.; Torino, F.; Pucci, S.; Costantini, M.; Capuano, R.; Graggi, C.; Polidoro, C.; Somma, G.; Pasqualetti, V.; Ketchanji, Y.; et al. Urine LOX-1 and Volatilome as Promising Tools towards the Early Detection of Renal Cancer. Cancers 2021, 13, 4213. [Google Scholar] [CrossRef]
- Tsumita, T.; Maishi, N.; Annam, D.A.; Towfik, M.A.; Matsuda, A.; Onodera, Y.; Nam, J.M.; Hida, Y.; Hida, K. The oxidized-LDL/LOX-1 axis in tumor endothelial cells enhances metastasis by recruiting neutrophils and cancer cells. Int. J. Cancer 2022, 151, 944–956. [Google Scholar] [CrossRef]
- Scalia, A.; Kindt, N.; Trelcat, A.; Nachtergael, A.; Duez, P.; Journé, F.; Carlier, S. Development of a Method for Producing oxLDL: Characterization of Their Effects on HPV-Positive Head and Neck Cancer Cells. Int. J. Mol. Sci. 2022, 23, 12552. [Google Scholar] [CrossRef]
- Xu, Q.; Wu, Q.; Li, H.; Tian, X.; Zuo, Y.; Zhang, Y.; Zhang, X.; Lin, Y.; Wu, Y.; Wang, Y.; et al. Joint High Level of Oxidized Low-Density Lipoprotein and High-Sensitivity C-Reactive Protein are Associated with Recurrent Stroke and Poor Functional Outcome in Minor Stroke or Transient Ischemic Attack. J. Am. Heart Assoc. 2022, 11, e027665. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.M.; Chen, H.; Sha, W.H.; Chen, X.F.; Yin, J.B.; Zhu, X.B.; Zheng, Z.W.; Ma, J. Oxidized low-density lipoprotein stimulates CD206 positive macrophages upregulating CD44 and CD133 expression in colorectal cancer with high-fat diet. World J. Gastroenterol. 2022, 28, 4993–5006. [Google Scholar] [CrossRef]
- Gonzalez-Chavarria, I.; Cerro, R.P.; Parra, N.P.; Sandoval, F.A.; Zuñiga, F.; Omazábal, V.A.; Lamperti, L.I.; Jiménez, S.P.; Fernandez, E.A.; Gutiérrez, N.A.; et al. Lectin-like oxidized LDL receptor-1 is an enhancer of tumor angiogenesis in human prostate cancer cells. PLoS ONE 2014, 9, e106219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Chavarria, I.; Fernandez, E.; Gutierrez, N.; González-Horta, E.E.; Sandoval, F.; Cifuentes, P.; Castillo, C.; Cerro, C.; Sanchez, O.; Toledo, J.R. LOX-1 activation by oxLDL triggers an epithelial mesenchymal transition and promotes tumorigenic potential in prostate cancer cells. Cancer Lett. 2018, 414, 34–43. [Google Scholar] [CrossRef]
- Jiang, J.; Yan, M.; Metha, J.L.; Hu, C. Angiogenesis is a link between atherosclerosis and tumorigenesis: Role of LOX-1. Cardiovasc. Drugs Ther. 2011, 25, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Mitra, S.; Wang, X.; Khaidakov, M.; Mehta, J.L. Oxidative stress and lectin-like ox-LDL-receptor LOX-1 in atherogenesis and tumorigenesis. Antioxid. Redox Signal. 2011, 15, 2301–2333. [Google Scholar] [CrossRef] [PubMed]
- Murdocca, M.; De Masi, C.; Pucci, S.; Mango, R.; Novelli, G.; Di Natale, C.; Sangiuolo, F. LOX-1 and cancer: An indissoluble liaison. Cancer Gene Ther. 2021, 28, 1088–1098. [Google Scholar] [CrossRef] [PubMed]
- Rizzacasa, B.; Morini, E.; Pucci, S.; Murdocca, M.; Novelli, G.; Amati., F. LOX-1 and Its Splice Variants: A New Challenge for Atherosclerosis and Cancer-Targeted Therapies. Int. J. Mol. Sci. 2017, 18, 290. [Google Scholar] [CrossRef] [Green Version]
- Rawla, P. Epidemiology of Prostate Cancer. World J. Oncol. 2019, 10, 63–89. [Google Scholar] [CrossRef] [Green Version]
- Karantanos, T.; Corn, P.; Thompson, T. Prostate cancer progression after androgen deprivation therapy: Mechanisms of castrate resistance and novel therapeutic approaches. Oncogene 2013, 32, 5501–5511. [Google Scholar] [CrossRef]
- Harris, W.P.; Mostaghel, E.A.; Nelson, P.S.; Montgomery, B. Androgen deprivation therapy: Progress in understanding mechanisms of resistance and optimizing androgen depletion. Nat. Clin. Pract. Urol. 2009, 6, 76–85. [Google Scholar] [CrossRef]
- Saad, F.; Hotte, S. Guidelines for the management of castrate-resistant prostate cancer. Can. Urol. Assoc. J. 2010, 4, 380–384. [Google Scholar] [CrossRef] [Green Version]
- Drake, J.M.; Graham, N.A.; Lee, J.K.; Stoyanova, T.; Faltermeier, C.M.; Sud, S.; Titz, B.; Huang, J.; Pienta, K.J.; Graeber, T.G.; et al. Metastatic castration-resistant prostate cancer reveals intrapatient similarity and interpatient heterogeneity of therapeutic kinase targets. Proc. Natl. Acad. Sci. USA 2013, 110, E4762–E4769. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Lu, J.; Xia, L.; Li, G. Drug Resistance of Enzalutamide in CRPC. Curr. Drug Targets 2018, 19, 613–620. [Google Scholar] [CrossRef]
- Adelaiye-Ogala, R.; Gryder, B.E.; Nguyen, Y.T.M.; Alilin, A.N.; Grayson, A.R.; Bajwa, W.; Jansson, K.H.; Beshiri, M.L.; Agarwal, S.; Rodriguez-Nieves, J.A.; et al. Targeting the PI3K/AKT Pathway Overcomes Enzalutamide Resistance by Inhibiting Induction of the Glucocorticoid Receptor. Mol. Cancer Ther. 2020, 19, 1436–1447. [Google Scholar] [CrossRef] [PubMed]
- Culig, Z. Response to Androgens and Androgen Receptor Antagonists in the Presence of Cytokines in Prostate Cancer. Cancers 2021, 13, 2944. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Peng, G.; Huang, H.; Liu, F.; Kong, D.P.; Dong, K.Q.; Dai, L.H.; Zhou, Z.; Wang, K.J.; Yang, J.; et al. Blocking the Feedback Loop between Neuroendocrine Differentiation and Macrophages Improves the Therapeutic Effects of Enzalutamide (MDV3100) on Prostate Cancer. Clin. Cancer Res. 2018, 24, 708–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tousignant, K.D.; Rockstroh, A.; Poad, B.L.J.; Talebi, A.; Young, R.S.E.; Fard, A.T.; Gupta, R.; Zang, T.; Wang, C.; Lehman, M.L.; et al. Therapy-induced lipid uptake and remodeling underpin ferroptosis hypersensitivity in prostate cancer. Cancer Metab. 2020, 8, 11. [Google Scholar] [CrossRef]
- Canton, J.; Neculai, D.; Grinstein, S. Scavenger receptors in homeostasis and immunity. Nat. Rev. Immunol. 2013, 13, 621–634. [Google Scholar] [CrossRef]
- Burtenshaw, D.; Kitching, M.; Redmond, E.M.; Megson, I.L.; Cahill, P.A. Reactive Oxygen Species (ROS), Intimal Thickening, and Subclinical Atherosclerotic Disease. Front. Cardiovasc. Med. 2019, 6, 89. [Google Scholar] [CrossRef]
- Liu, V.W.S.; Yau, W.L.; Tam, C.W.; Yao, K.M.; Shiu, S.Y.W. Melatonin Inhibits Androgen Receptor Splice Variant-7 (AR-V7)-Induced Nuclear Factor-Kappa B (NF-kappaB) Activation and NF-kappaB Activator-Induced AR-V7 Expression in Prostate Cancer Cells: Potential Implications for the Use of Melatonin in Castration-Resistant Prostate Cancer (CRPC) Therapy. Int. J. Mol. Sci. 2017, 18, 1130. [Google Scholar]
- Xu, L.; Chen, X.; Shen, M.; Dong-Rong, Y.; Fang, L.; Weng, G.; Tsai, Y.; Keng, P.C.; Chen, Y.; Lee, S.O. Inhibition of IL-6-JAK/Stat3 signaling in castration-resistant prostate cancer cells enhances the NK cell-mediated cytotoxicity via alteration of PD-L1/NKG2D ligand levels. Mol. Oncol. 2018, 12, 269–286. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Zhu, Y.; Lou, W.; Cui, Y.; Evans, C.P.; Gao, A.C. Inhibition of constitutively active Stat3 reverses enzalutamide resistance in LNCaP derivative prostate cancer cells. Prostate 2014, 74, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Antonarakis, E.S.; Lu, C.; Wang, H.; Luber, B.; Nakazawa, M.; Roeser, J.A.; Chen, Y.; Mohammad, T.A.; Chen, Y.; Fedor, H.L.; et al. AR-V7 and resistance to enzalutamide and abiraterone in prostate cancer. N. Engl. J. Med. 2014, 371, 1028–1038. [Google Scholar] [CrossRef] [Green Version]
- Sprenger, C. Androgen Receptor Variants and Castration-resistant Prostate Cancer: Looking Back and Looking Forward. Eur. Urol. 2018, 73, 724–726. [Google Scholar]
- Buttigliero, C.; Tucci, M.; Bertaglia, F.; Bironzo, P.; Di Maio, M.; Scagliotti, G.V. Understanding and overcoming the mechanisms of primary and acquired resistance to abiraterone and enzalutamide in castration resistant prostate cancer. Cancer Treat. Rev. 2015, 41, 884–892. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Dalrymple, S.L.; Coleman, I.; Zheng, S.L.; Xu, J.; Hooper, J.E.; Antonarakis, E.S.; De Marzo, A.M.; Meeker, A.K.; Nelson, P.S.; et al. Role of androgen receptor splice variant-7 (AR-V7) in prostate cancer resistance to 2nd-generation androgen receptor signaling inhibitors. Oncogene 2020, 39, 6935–6949. [Google Scholar] [CrossRef]
- Fujita, K.; Nonomura, N. Role of Androgen Receptor in Prostate Cancer: A Review. World J. Mens Health 2019, 37, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Thaper, D.; Vahid, S.; Kaur, R.; Kumar, S.; Nouruzi, S.; Bishop, J.L.; Johansson, M.; Zoubeidi, A. Galiellalactone inhibits the STAT3/AR signaling axis and suppresses Enzalutamide-resistant Prostate Cancer. Sci. Rep. 2018, 8, 17307. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Ogura, S.; Chen, J.; Little, P.J.; Moss, J.; Liu, P. LOX-1 in atherosclerosis: Biological functions and pharmacological modifiers. Cell Mol. Life Sci. 2013, 70, 2859–2872. [Google Scholar] [CrossRef] [Green Version]
- Don-Doncow, N.; Marginean, F.; Coleman, I.; Nelson, P.S.; Ehrnström, R.; Krzyzanowska, A.; Morrissey, C.; Hellsten, R.; Bjartell, A. Expression of STAT3 in Prostate Cancer Metastases. Eur. Urol. 2017, 71, 313–316. [Google Scholar] [CrossRef] [Green Version]
- Lubrano, V.; Balzan, S. LOX-1; ROS, inseparable factors in the process of endothelial damage. Free Radic. Res. 2014, 48, 841–848. [Google Scholar] [CrossRef]
- Sata, M.; Walsh, K. Endothelial cell apoptosis induced by oxidized LDL is associated with the down-regulation of the cellular caspase inhibitor FLIP. J. Biol. Chem. 1998, 273, 33103–33106. [Google Scholar] [CrossRef] [Green Version]
- Heinloth, A.; Heermeier, K.; Raff, U.; Wanner, C.; Galle, J.A.N. Stimulation of NADPH oxidase by oxidized low-density lipoprotein induces proliferation of human vascular endothelial cells. J. Am. Soc. Nephrol. 2000, 11, 1819–1825. [Google Scholar] [CrossRef]
- Hirsch, H.A.; Iliopoulos, D.; Joshi, A.; Zhang, Y.; Jaeger, S.A.; Bulyk, M.; Tsichlis, P.N.; Liu, S.; Strulh, K. A transcriptional signature and common gene networks link cancer with lipid metabolism and diverse human diseases. Cancer Cell 2010, 17, 348–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khaidakov, M.; Mehta, J. Oxidized LDL triggers pro-oncogenic signaling in human breast mammary epithelial cells partly via stimulation of MiR-21. PLoS ONE 2012, 7, e46973. [Google Scholar] [CrossRef] [PubMed]
- Bubici, C.; Papa, S.; Dean, K.; Franzoso, G. Mutual cross-talk between reactive oxygen species and nuclear factor-kappa B: Molecular basis and biological significance. Oncogene 2006, 25, 6731–6748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lingappan, K. NF-kappaB in Oxidative Stress. Curr. Opin. Toxicol. 2018, 7, 81–86. [Google Scholar] [CrossRef]
- Morgan, M.J.; Liu, Z. Crosstalk of reactive oxygen species and NF-kappaB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekar, T.; Yang, J.C.; Gao, A.C.; Evans, C.P. Mechanisms of resistance in castration-resistant prostate cancer (CRPC). Transl. Androl. Urol. 2015, 4, 365–380. [Google Scholar]
- Johnson, D.E.; O’Keefe, R.A.; Grandis, J. Targeting the IL-6/JAK/STAT3 signalling axis in cancer. Nat. Rev. Clin. Oncol. 2018, 15, 234–248. [Google Scholar] [CrossRef]
- Schust, J.; Sperl, B.; Hollis, A.; Mayer, T.U.; Berg, T. Stattic: A small-molecule inhibitor of STAT3 activation and dimerization. Chem. Biol. 2006, 13, 1235–1242. [Google Scholar] [CrossRef] [Green Version]
- Culig, Z.; Puhr, M. Interleukin-6: A multifunctional targetable cytokine in human prostate cancer. Mol. Cell Endocrinol. 2012, 360, 52–58. [Google Scholar] [CrossRef]
- Lou, W.; Ni, Z.; Dyer, K.; Tweardy, D.J.; Gao, A.C. Interleukin-6 induces prostate cancer cell growth accompanied by activation of stat3 signaling pathway. Prostate 2000, 42, 239–242. [Google Scholar] [CrossRef]
- Sreenivasan, L.; Wang, H.; Yap, S.Q.; Leclair, P.; Tam, A.; Lim, C.J. Autocrine IL-6/STAT3 signaling aids development of acquired drug resistance in Group 3 medulloblastoma. Cell Death Dis. 2020, 11, 1035. [Google Scholar] [CrossRef] [PubMed]
- Jin, R.; Yamashita, H.; Yu, X.; Wang, J.; Franco, O.E.; Hayward, S.W.; Matusik, R.J. Inhibition of NF-kappa B signaling restores responsiveness of castrate-resistant prostate cancer cells to anti-androgen treatment by decreasing androgen receptor-variant expression. Oncogene 2015, 34, 3700–3710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austin, D.C.; Strand, D.W.; Love, H.L.; Franco, O.E.; Jang, A.; Grabowska, M.M.; Miller, N.L.; Hameed, O.; Clark, P.E.; Fowke, J.H.; et al. NF-kappaB and androgen receptor variant expression correlate with human BPH progression. Prostate 2016, 76, 491–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiliccioglu, I.; Konac, E.; Dikmen, A.U.; Sozen, S.; Bilen, C.Y. Hsp-27 and NF-kappaB pathway is associated with AR/AR-V7 expression in prostate cancer cells. Gene 2019, 697, 138–143. [Google Scholar] [CrossRef]
- Nadiminty, N.; Tummala, R.; Liu, C.; Yang, J.; Lou, W.; Evans, C.P.; Gao, A.C. NF-kappaB2/p52 induces resistance to enzalutamide in prostate cancer: Role of androgen receptor and its variants. Mol. Cancer Ther. 2013, 12, 1629–1637. [Google Scholar] [CrossRef] [Green Version]
- Kiliccioglu, I.; Bilen, C.Y.; Sozen, S.; Konac, E. Upregulation of potential regulatory signaling molecules correlate with androgen receptor splice variants AR-V7 and AR-V567es in prostate cancer metastasis. Gene 2021, 772, 145377. [Google Scholar] [CrossRef]
- Gales, L.; Forsea, L.; Mitrea, D.; Stefanica, I.; Stanculescu, I.; Mitrica, R.; Georgescu, M.; Trifanescu, O.; Anghel, R.; Serbanescu, L. Antidiabetics, Anthelmintics, Statins, and Beta-Blockers as Co-Adjuvant Drugs in Cancer Therapy. Medicina 2022, 58, 1239. [Google Scholar] [CrossRef]
- Biocca, S.; Iacovelli, F.; Matarazzo, S.; Vindigni, G.; Oteri, F.; Desideri, A.; Falconi, M. Molecular mechanism of statin-mediated LOX-1 inhibition. Cell Cycle 2015, 14, 1583–1595. [Google Scholar] [CrossRef] [Green Version]
- Wadosky, K.M.; Koochekpour, S. Androgen receptor splice variants and prostate cancer: From bench to bedside. Oncotarget 2017, 8, 18550–18576. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Gong, S.; Roy-Burman, P.; Lee, P.; Culig, Z. Current mouse and cell models in prostate cancer research. Endocr. Relat. Cancer 2013, 20, R155–R170. [Google Scholar] [CrossRef] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duprat, F.; Robles, C.; Castillo, M.P.; Rivas, Y.; Mondaca, M.; Jara, N.; Roa, F.; Bertinat, R.; Toledo, J.; Paz, C.; et al. LOX-1 Activation by oxLDL Induces AR and AR-V7 Expression via NF-κB and STAT3 Signaling Pathways Reducing Enzalutamide Cytotoxic Effects. Int. J. Mol. Sci. 2023, 24, 5082. https://doi.org/10.3390/ijms24065082
Duprat F, Robles C, Castillo MP, Rivas Y, Mondaca M, Jara N, Roa F, Bertinat R, Toledo J, Paz C, et al. LOX-1 Activation by oxLDL Induces AR and AR-V7 Expression via NF-κB and STAT3 Signaling Pathways Reducing Enzalutamide Cytotoxic Effects. International Journal of Molecular Sciences. 2023; 24(6):5082. https://doi.org/10.3390/ijms24065082
Chicago/Turabian StyleDuprat, Felix, Catalina Robles, María Paz Castillo, Yerko Rivas, Marcela Mondaca, Nery Jara, Francisco Roa, Romina Bertinat, Jorge Toledo, Cristian Paz, and et al. 2023. "LOX-1 Activation by oxLDL Induces AR and AR-V7 Expression via NF-κB and STAT3 Signaling Pathways Reducing Enzalutamide Cytotoxic Effects" International Journal of Molecular Sciences 24, no. 6: 5082. https://doi.org/10.3390/ijms24065082