
miRNome and Proteome Profiling of Human Keratinocytes and Adipose Derived Stem Cells Proposed miRNA-Mediated Regulations of Epidermal Growth Factor and Interleukin 1-Alpha
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
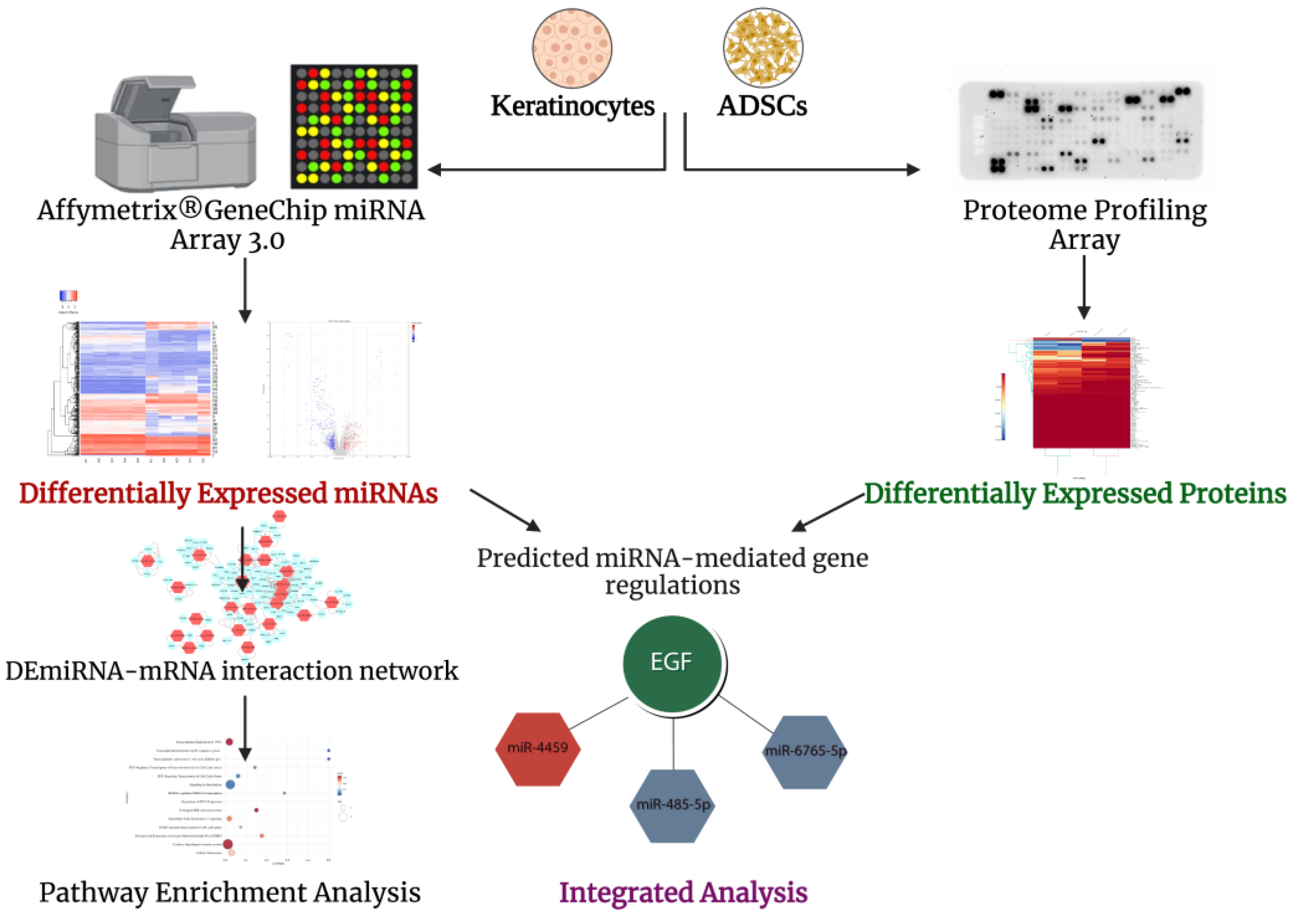
2. Results
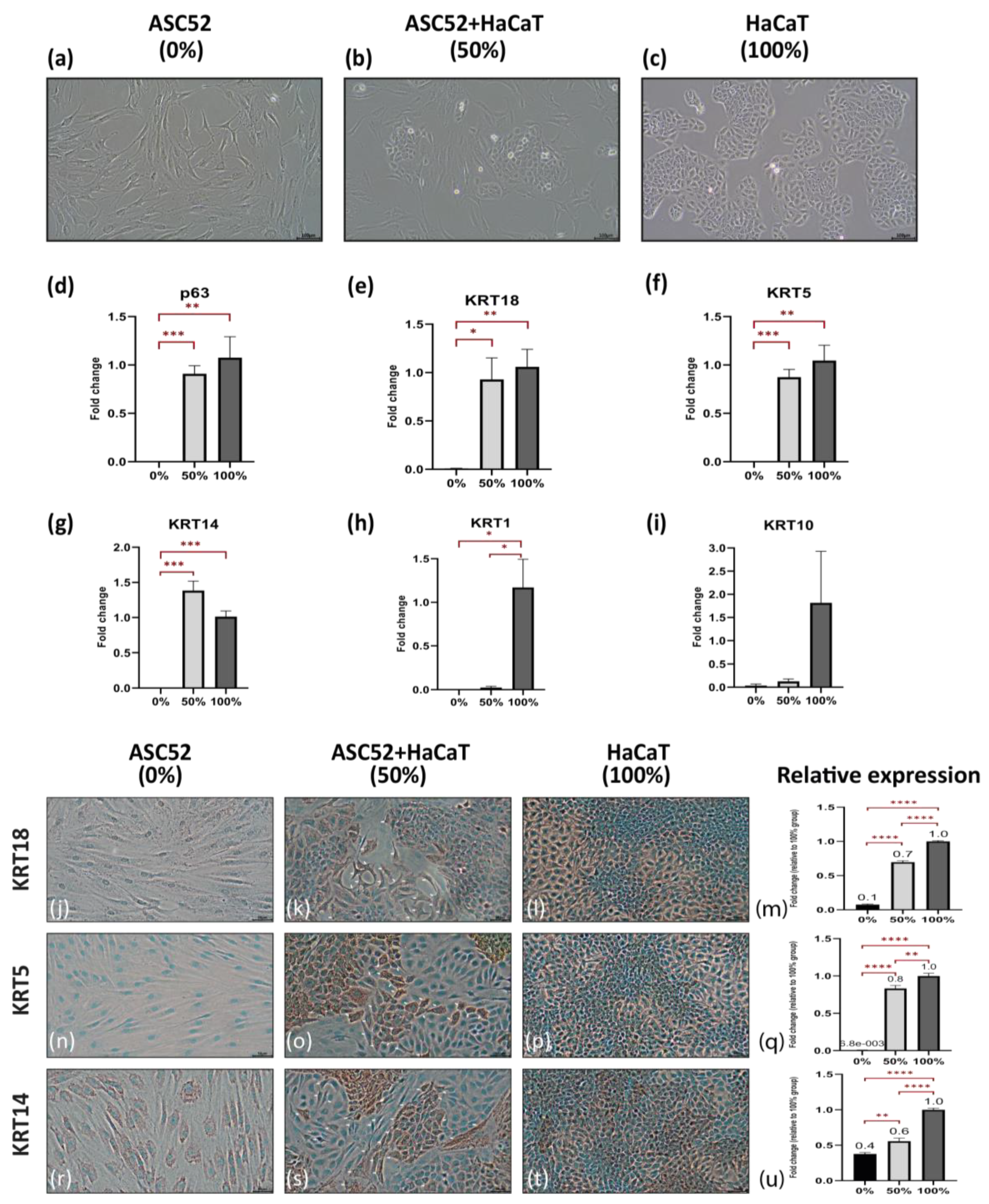
2.1. Direct Co-Culture Can Enhance the Commitment of ADSCs towards the Epidermal Lineage
2.2. 378 miRNAs Differentially Expressed between Primary Keratinocytes and ADSCs
2.3. Differentially Expressed miRNAs–mRNAs Interactome and Enrichment Analysis of the Upregulated miRNA Targets in Keratinocytes Reveals 14 Pathways
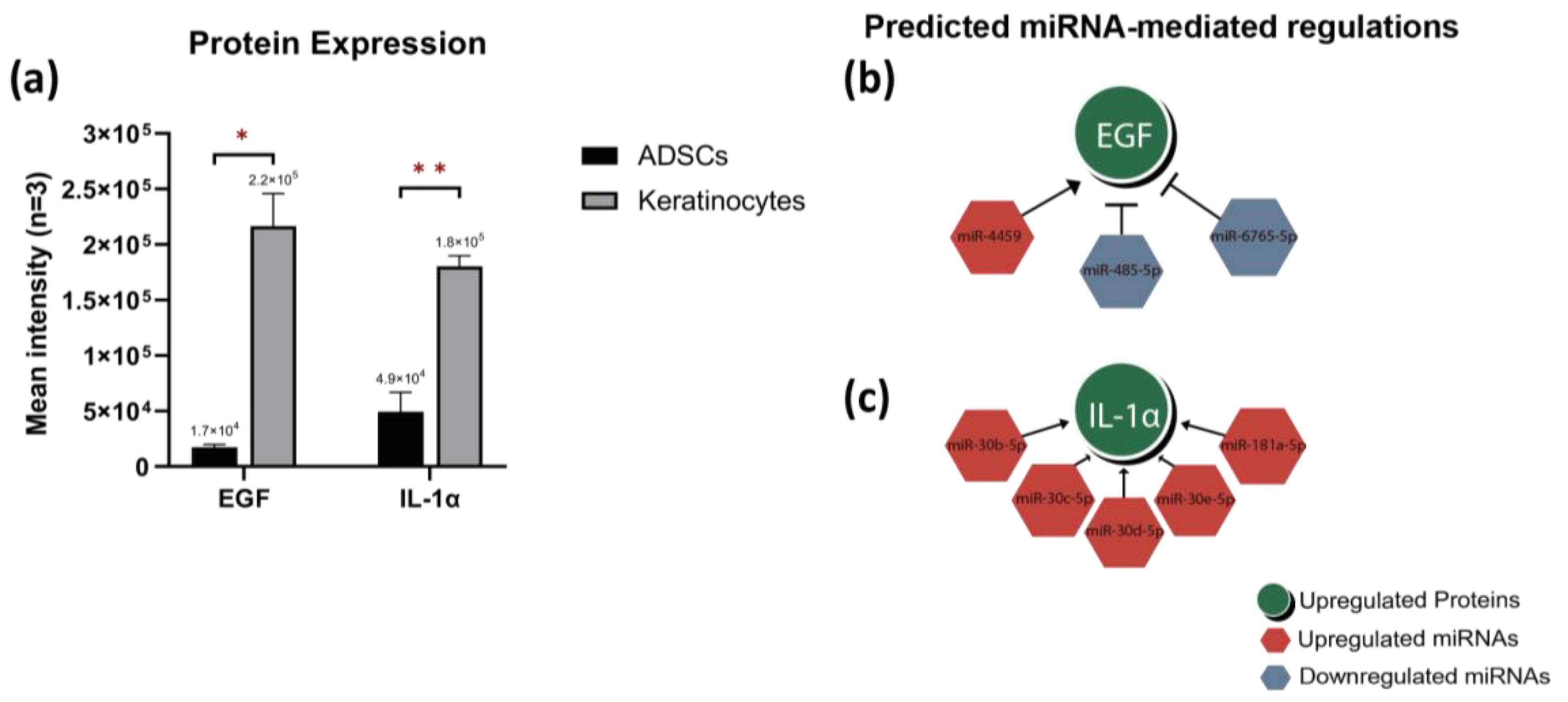
2.4. Proteome Profiling and Integrated Analysis with Differentially Expressed miRNAs Highlights miRNA-Mediated Regulations
3. Discussion
4. Materials and Methods
4.1. Epidermal Differentiation Induction in Direct Co-Culture Model
4.2. Primary Cell Isolation
4.3. Human miRNome Profiling with Microarrays
4.4. Gene and miRNA Expression
4.5. DEmiRNAs-mRNA Interaction Network and Pathway Enrichment Analysis
4.6. Immunocytochemistry Staining
4.7. Proteome Profiling
4.8. miRNA-Mediated Gene and Protein Regulations (Integrated Analysis)
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hassan, W.U.; Greiser, U.; Wang, W. Role of adipose-derived stem cells in wound healing. Wound Repair Regen. 2014, 22, 313–325. [Google Scholar] [CrossRef]
- Soliman, A.M.; Das, S.; Ghafar, N.A.; Teoh, S.L. Role of MicroRNA in Proliferation Phase of Wound Healing. Front Genet. Front. Genet. 2018, 9, 38. [Google Scholar] [CrossRef] [Green Version]
- Reinke, J.; Sorg, H. Wound Repair and Regeneration. Eur. Surg. Res. 2012, 49, 35–43. [Google Scholar] [CrossRef]
- Roh, C.; Lyle, S. Cutaneous Stem Cells and Wound Healing. Pediatr. Res. 2006, 59, 100R–103R. [Google Scholar] [CrossRef] [Green Version]
- Pastar, I.; Stojadinovic, O.; Yin, N.C.; Ramirez, H.; Nusbaum, A.G.; Sawaya, A.; Patel, S.B.; Khalid, L.; Isseroff, R.R.; Tomic-Canic, M. Epithelialization in Wound Healing: A Comprehensive Review. Adv. Wound Care 2014, 3, 445–464. [Google Scholar] [CrossRef] [Green Version]
- Kapur, S.; Katz, A.J. Characterization of Adipose-Derived Stem Cells: An Update. Curr. Stem Cell Res. Ther. 2010, 5, 95–102. [Google Scholar]
- Shi, J.-G.; Fu, W.-J.; Wang, X.-X.; Xu, Y.-D.; Li, G.; Hong, B.-F.; Hu, K.; Cui, F.-Z.; Wang, Y.; Zhang, X. Transdifferentiation of human adipose-derived stem cells into urothelial cells: Potential for urinary tract tissue engineering. Cell Tissue Res. 2012, 347, 737–746. [Google Scholar] [CrossRef]
- Alghfeli, L.; Parambath, D.; Manzoor, S.; Roach, H.I.; Oreffo, R.O.; El-Serafi, A.T. Synthesis of scaffold-free, three dimensional, osteogenic constructs following culture of skeletal osteoprogenitor cells on glass surfaces. Bone Rep. 2021, 15, 101143. [Google Scholar] [CrossRef]
- Elsharkawi, I.; Parambath, D.; Saber-Ayad, M.; Khan, A.A.; El-Serafi, A.T. Exploring the effect of epigenetic modifiers on developing insulin-secreting cells. Hum. Cell 2019, 33, 1–9. [Google Scholar] [CrossRef]
- Karlsson, M.; Steinvall, I.; Olofsson, P.; Thorfinn, J.; Sjöberg, F.; Åstrand, L.; Fayiz, S.; Khalaf, A.; Divyasree, P.; El-Serafi, A.T.; et al. Sprayed cultured autologous keratinocytes in the treatment of severe burns: A retrospective matched cohort study. Ann. Burn. Fire Disasters 2020, 33, 134–142. [Google Scholar]
- Sierra-Sánchez, A.; Ordonez-Luque, A.; Ibanez, O.E.; Ruiz-Garcia, A.; Santiago, S.A. Epithelial In vitro Differentiation of Mesenchymal Stem Cells. Curr. Stem Cell Res. Ther. 2018, 13, 409–422. [Google Scholar] [CrossRef]
- Enzo, E.; Seconetti, A.S.; Forcato, M.; Tenedini, E.; Polito, M.P.; Sala, I.; Carulli, S.; Contin, R.; Peano, C.; Tagliafico, E.; et al. Single-keratinocyte transcriptomic analyses identify different clonal types and proliferative potential mediated by FOXM1 in human epidermal stem cells. Nat. Commun. 2021, 12, 2505. [Google Scholar] [CrossRef]
- El-Serafi, A.T.; El-Serafi, I.; Elmasry, M.; Steinvall, I.; Sjöberg, F. Skin regeneration in three dimensions, current status, challenges and opportunities. Differentiation 2017, 96, 26–29. [Google Scholar] [CrossRef]
- Roxburgh, J.; Metcalfe, A.D.; Martin, Y.H. The effect of medium selection on adipose-derived stem cell expansion and differentiation: Implications for application in regenerative medicine. Cytotechnology 2015, 68, 957–967. [Google Scholar] [CrossRef] [Green Version]
- Petry, L.; Kippenberger, S.; Meissner, M.; Kleemann, J.; Kaufmann, R.; Rieger, U.M.; Wellenbrock, S.; Reichenbach, G.; Zöller, N.; Valesky, E. Directing adipose-derived stem cells into keratinocyte-like cells: Impact of medium composition and culture condition. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 2010–2019. [Google Scholar] [CrossRef]
- Lau, K.; Paus, R.; Tiede, S.; Day, P.; Bayat, A. Exploring the role of stem cells in cutaneous wound healing. Exp. Dermatol. 2009, 18, 921–933. [Google Scholar] [CrossRef]
- Segalla, L.; Chirumbolo, S.; Sbarbati, A. Dermal white adipose tissue: Much more than a metabolic, lipid-storage organ? Tissue Cell 2021, 71, 101583. [Google Scholar] [CrossRef]
- Merrick, D.; Seale, P. Skinny Fat Cells Stimulate Wound Healing. Cell Stem Cell 2020, 26, 801–803. [Google Scholar] [CrossRef]
- Shook, B.A.; Wasko, R.R.; Mano, O.; Rutenberg-Schoenberg, M.; Rudolph, M.C.; Zirak, B.; Rivera-Gonzalez, G.C.; López-Giráldez, F.; Zarini, S.; Rezza, A.; et al. Dermal Adipocyte Lipolysis and Myofibroblast Conversion Are Required for Efficient Skin Repair. Cell Stem Cell 2020, 26, 880–895.e6. [Google Scholar] [CrossRef]
- Hu, L.; Wang, J.; Zhou, X.; Xiong, Z.; Zhao, J.; Yu, R.; Huang, F.; Zhang, H.; Chen, L. Exosomes derived from human adipose mensenchymal stem cells accelerates cutaneous wound healing via optimizing the characteristics of fibroblasts. Sci. Rep. 2016, 6, 32993. [Google Scholar] [CrossRef] [Green Version]
- Van Dongen, J.A.; Harmsen, M.C.; Van der Lei, B.; Stevens, H.P. Augmentation of Dermal Wound Healing by Adipose Tissue-Derived Stromal Cells (ASC). Bioengineering 2018, 5, 91. [Google Scholar] [CrossRef] [Green Version]
- Gebert, L.F.R.; Macrae, I.J. Regulation of microRNA function in animals. Nat. Rev. Mol. Cell Biol. 2018, 20, 21–37. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Makarova, J.A.; Shkurnikov, M.U.; Wicklein, D.; Lange, T.; Samatov, T.R.; Turchinovich, A.A.; Tonevitsky, A.G. Intracellular and extracellular microRNA: An update on localization and biological role. Prog. Histochem. Cytochem. 2016, 51, 33–49. [Google Scholar] [CrossRef]
- El-Serafi, A.T. Epigenetic Modifiers and Stem Cell Differentiation, in Stem Cells and Cancer Stem Cells; Hayat, M., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 147–154. [Google Scholar]
- Yi, R.; Pasolli, H.A.; Landthaler, M.; Hafner, M.; Ojo, T.; Sheridan, R.; Sander, C.; O’Carroll, D.; Stoffel, M.; Tuschl, T.; et al. DGCR8-dependent microRNA biogenesis is essential for skin development. Proc. Natl. Acad. Sci. USA 2009, 106, 498–502. [Google Scholar] [CrossRef] [Green Version]
- Andl, T.; Murchison, E.P.; Liu, F.; Zhang, Y.; Yunta-Gonzalez, M.; Tobias, J.W.; Andl, C.D.; Seykora, J.T.; Hannon, G.J.; Millar, S.E. The miRNA-processing enzyme dicer is essential for the morphogenesis and maintenance of hair follicles. Curr. Biology 2006, 16, 1041–1049. [Google Scholar] [CrossRef] [Green Version]
- Lerman, G.; Sharon, M.; Leibowitz-Amit, R.; Sidi, Y.; Avni, D. The crosstalk between IL-22 signaling and miR-197 in human keratinocytes. PLoS ONE 2014, 9, e107467. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.; Kaplan, N.; Hamanaka, R.B.; Katsnelson, J.; Blatt, H.; Yang, W.; Hao, L.; Bryar, P.J.; Johnson, R.S.; Getsios, S.; et al. microRNA-31/factor-inhibiting hypoxia-inducible factor 1 nexus regulates keratinocyte differentiation. Proc. Natl. Acad. Sci. USA 2012, 109, 14030–14034. [Google Scholar] [CrossRef] [Green Version]
- Yi, R.; Poy, M.N.; Stoffel, M.; Fuchs, E. A skin microRNA promotes differentiation by repressing ‘stemness’. Nature 2008, 452, 225–229. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, J.; Chan, Y.C.; Sen, C.K. MicroRNAs in skin and wound healing. Physiol. Genom. 2011, 43, 543–556. [Google Scholar] [CrossRef]
- Chen, J.; Deng, S.; Zhang, S.; Chen, Z.; Wu, S.; Cai, X.; Yang, X.; Guo, B.; Peng, Q. The role of miRNAs in the differentiation of adipose-derived stem cells. Curr. Stem Cell Res. Ther. 2014, 9, 268–279. [Google Scholar] [CrossRef]
- Yang, C.; Luo, M.; Chen, Y.; You, M.; Chen, Q. MicroRNAs as Important Regulators Mediate the Multiple Differentiation of Mesenchymal Stromal Cells. Front. Cell Dev. Biol. 2021, 9, 619842. [Google Scholar] [CrossRef]
- Alghfeli, L.; Parambath, D.; Eldeen, L.A.T.; El-Serafi, I.; El-Serafi, A.T. Non-additive effect of the DNA methylation inhibitor, 5-Aza-dC, and glass as a culture surface on osteogenic differentiation. Heliyon 2022, 8, e12433. [Google Scholar] [CrossRef]
- Lizio, M.; The FANTOM Consortium; Harshbarger, J.; Shimoji, H.; Severin, J.; Kasukawa, T.; Sahin, S.; Abugessaisa, I.; Fukuda, S.; Hori, F.; et al. Gateways to the FANTOM5 promoter level mammalian expression atlas. Genome Biol. 2015, 16, 22. [Google Scholar] [CrossRef] [Green Version]
- Jin, Q.; Li, P.; Yuan, K.; Zhao, F.; Zhu, X.; Zhang, P.; Huang, Z. Extracellular vesicles derived from human dental pulp stem cells promote osteogenesis of adipose-derived stem cells via the MAPK pathway. J. Tissue Eng. 2020, 11, 2041731420975569. [Google Scholar] [CrossRef]
- Zhu, Z.; Guo, L.; Yeltai, N.; Xu, H.; Zhang, Y. Chemokine (C-C motif) ligand 2-enhanced adipogenesis and angiogenesis of human adipose-derived stem cell and human umbilical vein endothelial cell co-culture system in adipose tissue engineering. J. Tissue Eng. Regen. Med. 2022, 16, 163–176. [Google Scholar] [CrossRef]
- Chavez-Munoz, C.; Nguyen, K.T.; Xu, W.; Hong, S.-J.; Mustoe, T.A.; Galiano, R.D. Transdifferentiation of adipose-derived stem cells into keratinocyte-like cells: Engineering a stratified epidermis. PLoS ONE 2013, 8, e80587. [Google Scholar] [CrossRef] [Green Version]
- Pellegrini, G.; Dellambra, E.; Golisano, O.; Martinelli, E.; Fantozzi, I.; Bondanza, S.; Ponzin, D.; McKeon, F.; De Luca, M. p63 identifies keratinocyte stem cells. Proc. Natl. Acad. Sci. USA 2001, 98, 3156–3161. [Google Scholar] [CrossRef] [Green Version]
- Koster, M.I.; Roop, D.R. Roop, The role of p63 in development and differentiation of the epidermis. J. Dermatol. Sci. 2004, 34, 3–9. [Google Scholar] [CrossRef]
- Truong, A.B.; Kretz, M.; Ridky, T.W.; Kimmel, R.; Khavari, P.A. p63 regulates proliferation and differentiation of developmentally mature keratinocytes. Genes Dev. 2006, 20, 3185–3197. [Google Scholar] [CrossRef] [Green Version]
- dos Santos, J.F.; Borçari, N.R.; Araújo, M.D.S.; Nunes, V.A. Mesenchymal stem cells differentiate into keratinocytes and express epidermal kallikreins: Towards an in vitro model of human epidermis. J. Cell. Biochem. 2019, 120, 13141–13155. [Google Scholar] [CrossRef]
- Cheng, Y.; Qin, K.; Huang, N.; Zhou, Z.; Xiong, H.; Zhao, J.; Zhang, Y.; Yu, S. Cytokeratin 18 regulates the transcription and alternative splicing of apoptoticrelated genes and pathways in HeLa cells. Oncol. Rep. 2019, 42, 301–312. [Google Scholar]
- Păunescu, V.; Deak, E.; Herman, D.; Siska, I.R.; T˘anasie, G.; Bunu, C.; Anghel, S.; Tatu, C.A.; Oprea, T.; Henschler, R.; et al. In vitro differentiation of human mesenchymal stem cells to epithelial lineage. J. Cell. Mol. Med. 2007, 11, 502–508. [Google Scholar] [CrossRef] [Green Version]
- Alam, H.; Sehgal, L.; Kundu, S.T.; Dalal, S.N.; Vaidya, M.M. Novel function of keratins 5 and 14 in proliferation and differentiation of stratified epithelial cells. Mol. Biol. Cell 2011, 22, 4068–4078. [Google Scholar] [CrossRef]
- Maeda, Y.; Hasegawa, T.; Wada, A.; Fukai, T.; Iida, H.; Sakamoto, A.; Ikeda, S. Adipose-derived stem cells express higher levels of type VII collagen under specific culture conditions. Arch. Dermatol. Res. 2017, 309, 843–849. [Google Scholar] [CrossRef]
- Xu, X.; Li, L.; Wang, C.; Liu, Y.; Chen, C.; Yan, J.; Ding, H.; Tang, S.-Y. The expansion of autologous adipose-derived stem cells in vitro for the functional reconstruction of nasal mucosal tissue. Cell Biosci. 2015, 5, 54. [Google Scholar] [CrossRef] [Green Version]
- Bayraktar, R.; Van Roosbroeck, K.; Calin, G.A. Cell-to-cell communication: MicroRNAs as hormones. Mol. Oncol. 2017, 11, 1673–1686. [Google Scholar] [CrossRef] [Green Version]
- Conti, I.; Varano, G.; Simioni, C.; Laface, I.; Milani, D.; Rimondi, E.; Neri, L.M. miRNAs as Influencers of Cell-Cell Communication in Tumor Microenvironment. Cells 2020, 9, 220. [Google Scholar] [CrossRef] [Green Version]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Veith, A.P.; Henderson, K.; Spencer, A.; Sligar, A.D.; Baker, A.B. Therapeutic strategies for enhancing angiogenesis in wound healing. Adv. Drug Deliv. Rev. 2018, 146, 97–125. [Google Scholar] [CrossRef]
- Ma, X.; Li, L.; Jia, T.; Chen, M.; Liu, G.; Li, C.; Li, N.; Yang, D. miR-203a controls keratinocyte proliferation and differentiation via targeting the stemness-associated factor DeltaNp63 and establishing a regulatory circuit with SNAI2. Biochem. Biophys. Res. Commun. 2017, 491, 241–249. [Google Scholar] [CrossRef]
- Hildebrand, J.; Rütze, M.; Walz, N.; Gallinat, S.; Wenck, H.; Deppert, W.; Grundhoff, A.; Knott, A. A comprehensive analysis of microRNA expression during human keratinocyte differentiation in vitro and in vivo. J. Investig. Dermatol. 2011, 131, 20–29. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, P.; Ceccoli, J. Advances in the Application and Impact of MicroRNAs as Therapies for Skin Disease. BioDrugs 2017, 31, 423–438. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Li, X.; Li, D.; Ren, X.; Li, Y.; Herter, E.K.; Qian, M.; Toma, M.-A.; Wintler, A.-M.; Sérézal, I.G.; et al. MicroRNA-34 Family Enhances Wound Inflammation by Targeting LGR4. J. Investig. Dermatol. 2020, 140, 465–476.e11. [Google Scholar] [CrossRef]
- Deverman, B.E.; Patterson, P.H. Cytokines and CNS Development. Neuron 2009, 64, 61–78. [Google Scholar] [CrossRef] [Green Version]
- Aurora, A.B.; Olson, E.N. Immune Modulation of Stem Cells and Regeneration. Cell Stem Cell 2014, 15, 14–25. [Google Scholar] [CrossRef] [Green Version]
- El-Serafi, A.T.; El-Serafi, I.; Steinvall, I.; Sjöberg, F.; Elmasry, M. A Systematic Review of Keratinocyte Secretions: A Regenerative Perspective. Int. J. Mol. Sci. 2022, 23, 7934. [Google Scholar] [CrossRef]
- Santos, J.; Dalla, P.V.; Milthorpe, B.K. A Molecular Analysis of Cytokine Content across Extracellular Vesicles, Secretions, and Intracellular Space from Different Site-Specific Adipose-Derived Stem Cells. Int. J. Mol. Sci. 2021, 23, 397. [Google Scholar] [CrossRef]
- Blumenberg, M. Profiling and metaanalysis of epidermal keratinocytes responses to epidermal growth factor. BMC Genom. 2013, 14, 85. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.Y.; Lee, Y.J.; Kim, J.M.; Kang, H.J.; Cho, S.H.; Chang, S.E. Epidermal Growth Factor Relieves Inflammatory Signals in Staphylococcus aureus-Treated Human Epidermal Keratinocytes and Atopic Dermatitis-Like Skin Lesions in Nc/Nga Mice. BioMed Res. Int. 2018, 2018, 9439182. [Google Scholar] [CrossRef] [Green Version]
- Barrientos, S.; Stojadinovic, O.; Golinko, M.S.; Brem, H.; Tomic-Canic, M. Growth factors and cytokines in wound healing. Wound Repair Regen. 2008, 16, 585–601. [Google Scholar] [CrossRef]
- Tomic-Canic, M.; Komine, M.; Freedberg, I.M.; Blumenberg, M. Epidermal signal transduction and transcription factor activation in activated keratinocytes. J. Dermatol. Sci. 1998, 17, 167–181. [Google Scholar] [CrossRef]
- Mao, Y.; Ma, J.; Xia, Y.; Xie, X. The Overexpression of Epidermal Growth Factor (EGF) in HaCaT Cells Promotes the Proliferation, Migration, Invasion and Transdifferentiation to Epidermal Stem Cell Immunophenotyping of Adipose-Derived Stem Cells (ADSCs). Int. J. Stem Cells 2020, 13, 93–103. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.D.; Kim, J.P.; Zhang, K.; Sarret, Y.; Wynn, K.C.; Kramer, R.H.; Woodley, D.T. Epidermal growth factor (EGF) promotes human keratinocyte locomotion on collagen by increasing the alpha 2 integrin subunit. Exp. Cell Res. 1993, 209, 216–223. [Google Scholar] [CrossRef]
- Cavalli, G.; Colafrancesco, S.; Emmi, G.; Imazio, M.; Lopalco, G.; Maggio, M.C.; Sota, J.; Dinarello, C.A. Interleukin 1alpha: A comprehensive review on the role of IL-1alpha in the pathogenesis and treatment of autoimmune and inflammatory diseases. Autoimmun. Rev. 2021, 20, 102763. [Google Scholar] [CrossRef]
- Xie, H.; Lim, B.; Lodish, H.F. MicroRNAs induced during adipogenesis that accelerate fat cell development are downregulated in obesity. Diabetes 2009, 58, 1050–1057. [Google Scholar] [CrossRef] [Green Version]
- Mao, L.; Liu, S.; Hu, L.; Jia, L.; Wang, H.; Guo, M.; Chen, C.; Liu, Y.; Xu, L. miR-30 Family: A Promising Regulator in Development and Disease. BioMed Res. Int. 2018, 2018, 9623412. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Xu, C.; Lei, F.; Liao, M.; Wang, W.; Xu, N.; Zhang, Y.; Xie, W. MiR-30a targets IL-1alpha and regulates islet functions as an inflammation buffer and response factor. Sci. Rep. 2017, 7, 5270. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Shen, Y.; Wang, W.; Li, X. MiR-30a-5p ameliorates spinal cord injury-induced inflammatory responses and oxidative stress by targeting Neurod 1 through MAPK/ERK signalling. Clin. Exp. Pharmacol. Physiol. 2017, 45, 68–74. [Google Scholar] [CrossRef]
- Valinezhad Orang, A.; Safaralizadeh, R.; Kazemzadeh-Bavili, M. Mechanisms of miRNA-Mediated Gene Regulation from Common Downregulation to mRNA-Specific Upregulation. Int. J. Genom. 2014, 2014, 970607. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.-Y. The Role of MicroRNAs in Epidermal Barrier. Int. J. Mol. Sci. 2020, 21, 5781. [Google Scholar] [CrossRef]
- Neu, J.; Dziunycz, P.J.; Dzung, A.; Lefort, K.; Falke, M.; Denzler, R.; Freiberger, S.N.; Iotzova-Weiss, G.; Kuzmanov, A.; Levesque, M.P.; et al. miR-181a decelerates proliferation in cutaneous squamous cell carcinoma by targeting the proto-oncogene KRAS. PLoS ONE 2017, 12, e0185028. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.; Han, L.; Su, L.; Zhao, J.; Zhang, Y.; Zhang, S.; Zhao, B.; Miao, J. A 3′UTR-associated RNA, FLJ11812 maintains stemness of human embryonic stem cells by targeting miR-4459. Stem Cells Dev. 2015, 24, 1133–1140. [Google Scholar] [CrossRef] [Green Version]
- Jang, T.H.; Huang, W.C.; Tung, S.L.; Lin, S.C.; Chen, P.M.; Cho, C.Y.; Yang, Y.Y.; Yen, T.C.; Lo, G.H.; Chuang, S.E.; et al. MicroRNA-485-5p targets keratin 17 to regulate oral cancer stemness and chemoresistance via the integrin/FAK/Src/ERK/beta-catenin pathway. J. Biomed. Sci. 2022, 29, 42. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Sticht, C.; De La Torre, C.; Parveen, A.; Gretz, N. miRWalk: An online resource for prediction of microRNA binding sites. PLoS ONE 2018, 13, e0206239. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Wong, N.; Wang, X. miRDB: An online resource for microRNA target prediction and functional annotations. Nucleic Acids Res. 2014, 43, D146–D152. [Google Scholar] [CrossRef] [Green Version]
- Hsu, S.D.; Lin, F.M.; Wu, W.Y.; Liang, C.; Huang, W.C.; Chan, W.L.; Tsai, W.T.; Chen, G.Z.; Lee, C.J.; Chiu, C.M.; et al. miRTarBase: A database curates experimentally validated microRNA-target interactions. Nucleic Acids Res. 2011, 39, D163–D169. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Yu, G.; He, Q.-Y. ReactomePA: An R/Bioconductor package for reactome pathway analysis and visualization. Mol. Biosyst. 2015, 12, 477–479. [Google Scholar] [CrossRef]
- Zhang, Y.; Roos, M.; Himburg, H.; Termini, C.M.; Quarmyne, M.; Li, M.; Zhao, L.; Kan, J.; Fang, T.; Yan, X.; et al. PTPsigma inhibitors promote hematopoietic stem cell regeneration. Nat. Commun. 2019, 10, 3667. [Google Scholar] [CrossRef] [Green Version]
- Omori-Miyake, M.; Yamashita, M.; Tsunemi, Y.; Kawashima, M.; Yagi, J. In vitro assessment of IL-4- or IL-13-mediated changes in the structural components of keratinocytes in mice and humans. J. Investig. Dermatol. 2014, 134, 1342–1350. [Google Scholar] [CrossRef] [Green Version]
- Herfs, M.; Hubert, P.; Suarez-Carmona, M.; Reschner, A.; Saussez, S.; Berx, G.; Savagner, P.; Boniver, J.; Delvenne, P. Regulation of p63 isoforms by snail and slug transcription factors in human squamous cell carcinoma. Am. J. Pathol. 2010, 176, 1941–1949. [Google Scholar] [CrossRef]
- Lin, K.Y.; Zhang, X.J.; Feng, D.D.; Zhang, H.; Zeng, C.W.; Han, B.W.; Zhou, A.D.; Qu, L.H.; Xu, L.; Chen, Y.Q. miR-125b, a target of CDX2, regulates cell differentiation through repression of the core binding factor in hematopoietic malignancies. J. Biol. Chem. 2011, 286, 38253–38263. [Google Scholar] [CrossRef] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shahin, H.; Abdallah, S.; Das, J.; He, W.; El-Serafi, I.; Steinvall, I.; Sjöberg, F.; Elmasry, M.; El-Serafi, A.T. miRNome and Proteome Profiling of Human Keratinocytes and Adipose Derived Stem Cells Proposed miRNA-Mediated Regulations of Epidermal Growth Factor and Interleukin 1-Alpha. Int. J. Mol. Sci. 2023, 24, 4956. https://doi.org/10.3390/ijms24054956
Shahin H, Abdallah S, Das J, He W, El-Serafi I, Steinvall I, Sjöberg F, Elmasry M, El-Serafi AT. miRNome and Proteome Profiling of Human Keratinocytes and Adipose Derived Stem Cells Proposed miRNA-Mediated Regulations of Epidermal Growth Factor and Interleukin 1-Alpha. International Journal of Molecular Sciences. 2023; 24(5):4956. https://doi.org/10.3390/ijms24054956
Chicago/Turabian StyleShahin, Hady, Sallam Abdallah, Jyotirmoy Das, Weihai He, Ibrahim El-Serafi, Ingrid Steinvall, Folke Sjöberg, Moustafa Elmasry, and Ahmed T. El-Serafi. 2023. "miRNome and Proteome Profiling of Human Keratinocytes and Adipose Derived Stem Cells Proposed miRNA-Mediated Regulations of Epidermal Growth Factor and Interleukin 1-Alpha" International Journal of Molecular Sciences 24, no. 5: 4956. https://doi.org/10.3390/ijms24054956