
Silencing of Pepper CaFtsH1 or CaFtsH8 Genes Alters Normal Leaf Development


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification of FtsH Proteins in Solanaceae
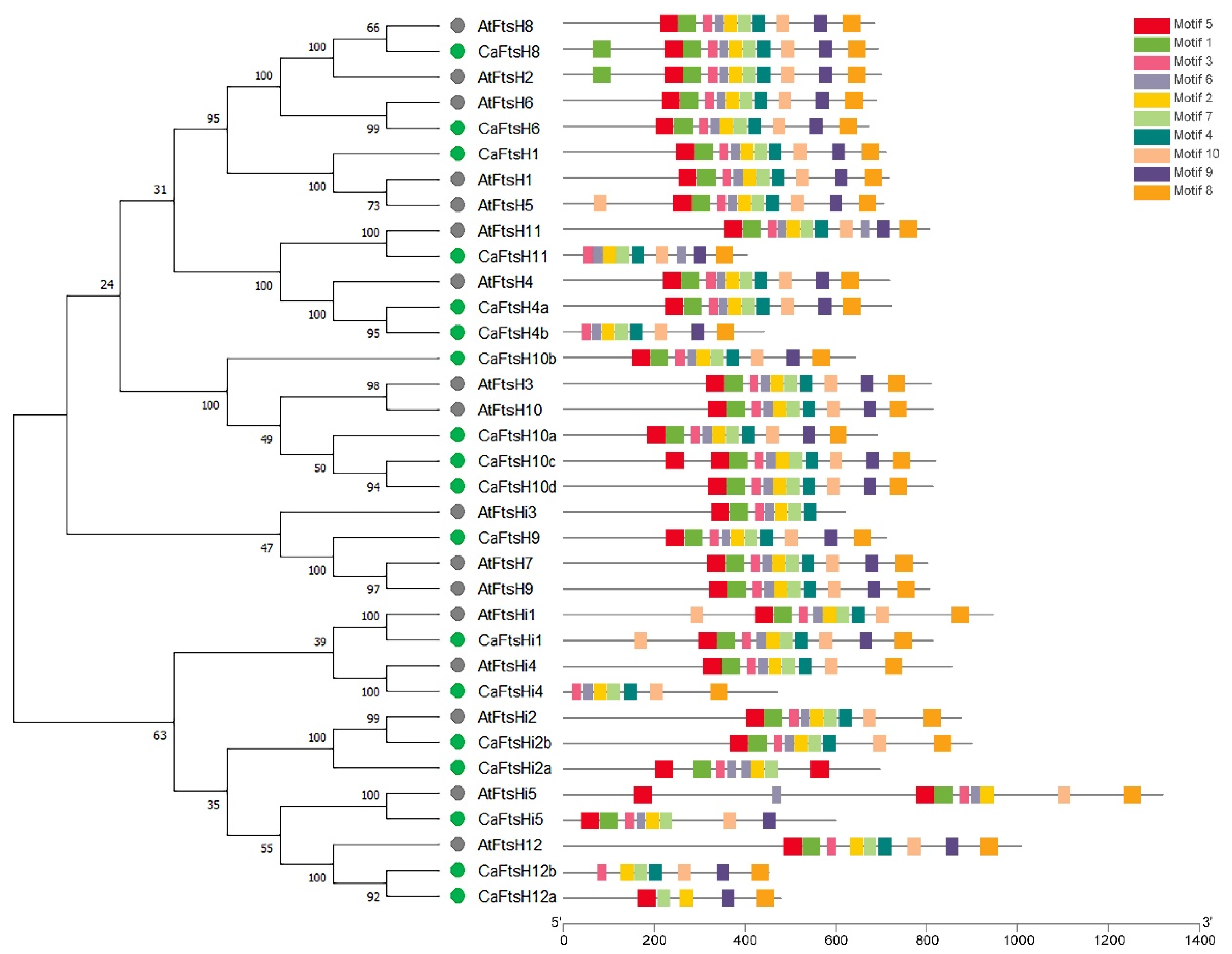
2.2. Phylogenetic and Motif Analysis of FtsH Proteins
2.3. Essential Roles of CaFtsH1 and CaFtsH8 in Pepper
2.4. Expression Pattern and Subcellular Location of CaFtsH1 and CaFtsH8
2.5. Gene Silencing of CaFtsH1 or CaFtsH8 Confers Albino Phenotypes
2.6. Analysis of Differentially Expressed Genes in CaFtsH1-Silenced Plants
2.7. Suppression of Photosynthesis-Related Genes in CaFtsH1-Silenced Plants
2.8. Silencing of CaFtsH1 Expression Influences CaPsbA Expression
3. Discussion
4. Materials and Methods
4.1. Identification of FtsH Proteins in Solanaceae
4.2. Properties, Structure, and Conserved Motifs of FtsH Proteins
4.3. Phylogenetic Analysis of FtsH Proteins
4.4. Expression Pattern and RT-qPCR Analysis
4.5. Subcellular Localization of CaFtsH1 and CaFtsH8
4.6. Gene Function Verification of CaFtsHs by VIGS
4.7. TEM Analysis
4.8. Acquisition and Analysis of Transcriptome Data
4.9. Yeast Two-Hybrid Pairwise Assays
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, Y.E.; Yuan, S.; Schröder, W.P. Comparison of methods for extracting thylakoid membranes of Arabidopsis plants. Physiol. Plant. 2016, 156, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Nelson, N.; Junge, W. Structure and energy transfer in photosystems of oxygenic photosynthesis. Annu. Rev. Biochem. 2015, 84, 659–683. [Google Scholar] [CrossRef] [PubMed]
- Holtzegel, U. The Lhc family of Arabidopsis thaliana. Endocyt. Cell Res. 2016, 27, 71–89. [Google Scholar]
- Su, X.; Ma, J.; Wei, X.; Cao, P.; Zhu, D.; Chang, W.; Liu, Z.; Zhang, X.; Li, M. Structure and assembly mechanism of plant C2S2M2-type PS II-LHC II supercomplex. Science 2017, 357, 815–820. [Google Scholar] [CrossRef] [Green Version]
- Alboresi, A.; Ballottari, M.; Hienerwadel, R.; Giacometti, G.M.; Morosinotto, T. Antenna complexes protect photosystem I from photoinhibition. BMC Plant Biol. 2009, 9, 71. [Google Scholar] [CrossRef] [Green Version]
- Murchie, E.H.; Niyogi, K.K. Manipulation of photoprotection to improve plant photosynthesis. Plant Physiol. 2011, 155, 86–92. [Google Scholar] [CrossRef] [Green Version]
- Abe, A.; Kosugi, S.; Yoshida, K.; Natsume, S.; Takagi, H.; Kanzaki, H.; Matsumura, H.; Yoshida, K.; Mitsuoka, C.; Tamiru, M. Genome sequencing reveals agronomically important loci in rice using MutMap. Nat. Biotechnol. 2012, 30, 174–178. [Google Scholar] [CrossRef] [Green Version]
- Kato, Y.; Miura, E.; Ido, K.; Ifuku, K.; Sakamoto, W. The variegated mutants lacking chloroplastic FtsHs are defective in D1 degradation and accumulate reactive oxygen species. Plant Physiol. 2009, 151, 1790–1801. [Google Scholar] [CrossRef] [Green Version]
- Nixon, P.J.; Michoux, F.; Yu, J.; Boehm, M.; Komenda, J. Recent advances in understanding the assembly and repair of photosystem II. Ann. Bot. 2010, 106, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Kato, Y.; Sun, X.; Zhang, L.; Sakamoto, W. Cooperative D1 degradation in the photosystem II repair mediated by chloroplastic proteases in Arabidopsis. Plant Physiol. 2012, 159, 1428–1439. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Fu, T.; Chen, N.; Guo, J.; Ma, J.; Zou, M.; Lu, C.; Zhang, L. The stromal chloroplast Deg7 protease participates in the repair of photosystem II after photoinhibition in Arabidopsis. Plant Physiol. 2010, 152, 1263–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Peng, L.; Guo, J.; Chi, W.; Ma, J.; Lu, C.; Zhang, L. Formation of DEG5 and DEG8 complexes and their involvement in the degradation of photodamaged photosystem II reaction center D1 protein in Arabidopsis. Plant Cell 2007, 19, 1347–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Han, T.; Yang, L.; Wang, Q.; Zhao, Y.; Jiang, D.; Ruan, X. The essential roles of OsFtsH2 in developing the chloroplast of rice. BMC Plant Biol. 2021, 21, 445. [Google Scholar] [CrossRef] [PubMed]
- Wagner, R.; Aigner, H.; Funk, C. FtsH proteases located in the plant chloroplast. Physiol. Plant 2012, 145, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Gao, M.Q.; Xue, R.L.; Wang, D.; Zhao, H.J. Effect of hydrogen sulfide on D1 protein in wheat under drought stress. Acta Physiol. Plant 2015, 37, 225. [Google Scholar] [CrossRef]
- Yin, Z.; Meng, F.; Song, H.; Wang, X.; Chao, M.; Zhang, G.; Xu, X.; Deng, D.; Yu, D. GmFtsH9 expression correlates with in vivo photosystem II function: Chlorophyll a fluorescence transient analysis and eQTL mapping in soybean. Planta 2011, 234, 815–827. [Google Scholar] [CrossRef]
- Zheng, C.; Kong, X.; Sui, J.; Shu, C. Identification, classification and salt stress expression analysis of metalloproteinase family gene FtsH in Peanut. Jiangsu Agric. Sci. 2016, 44, 74–77. [Google Scholar]
- Sun, A.; Yi, S.; Yang, J.; Zhao, C.; Liu, J. Identification and characterization of a heat-inducible FtsH gene from tomato (Lycopersicon esculentum Mill.). Plant Sci. 2006, 170, 551–562. [Google Scholar] [CrossRef]
- Pu, T.; Mo, Z.; Su, L.; Yang, J.; Wan, K.; Wang, L.; Liu, R.; Liu, Y. Genome-wide identification and expression analysis of the FtsH protein family and its response to abiotic stress in Nicotiana tabacum L. BMC Genom. 2022, 23, 503. [Google Scholar] [CrossRef]
- Guo, Z.; Gao, X.; Cai, H.; Yu, L.; Gu, C.; Zhang, S.L. Genome-wide identification, evolution and expression analysis of the FtsH gene during fruit development in pear (Pyrus bretschneideri). Plant Biotechnol. Rep. 2021, 15, 537–550. [Google Scholar] [CrossRef]
- Xu, K.; Wu, Y.; Song, J.; Hu, K.; Wu, Z.; Wen, J.; Yi, B.; Ma, C.; Shen, J.; Fu, T. Fine mapping and identification of BnaC06.FtsH1, a lethal gene that regulates the PSII repair cycle in Brassica napus. Int. J. Mol. Sci. 2021, 22, 2087. [Google Scholar] [CrossRef] [PubMed]
- Ivashuta, S.; Imai, R.; Uchiyama, K.; Gau, M.; Shimamoto, Y. Changes in chloroplast FtsH-like gene during cold acclimation in alfalfa (Medicago sativa). J. Plant Physiol. 2002, 159, 85–90. [Google Scholar] [CrossRef]
- Sakamoto, W.; Miura, E.; Kaji, Y.; Okuno, T.; Nishizono, M.; Ogura, T. Allelic characterization of the leaf-variegated mutation var2 identifies the conserved amino acid residues of FtsH that are important for ATP hydrolysis and proteolysis. Plant Mol. Biol. 2004, 56, 705–716. [Google Scholar] [CrossRef]
- Boehm, M.; Yu, J.; Krynicka, V.; Barker, M.; Tichy, M.; Komenda, J.; Nixon, P.J.; Nield, J. Subunit organization of a Synechocystis hetero-oligomeric thylakoid FtsH complex involved in photosystem II repair. Plant Cell 2012, 24, 3669–3683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, L.S.; Funk, C. The FtsHi enzymes of Arabidopsis thaliana: Pseudo-proteases with an important function. Int. J. Mol. Sci. 2021, 22, 5917. [Google Scholar] [CrossRef]
- Sakamoto, W.; Zaltsman, A.; Adam, Z.; Takahashi, Y. Coordinated regulation and complex formation of yellow variegated1 and yellow variegated2, chloroplastic FtsH metalloproteases involved in the repair cycle of photosystem II in Arabidopsis thylakoid membranes. Plant Cell 2003, 15, 2843–2855. [Google Scholar] [CrossRef] [Green Version]
- Putarjunan, A.; Liu, X.; Nolan, T.; Yu, F.; Rodermel, S. Understanding chloroplast biogenesis using second-site suppressors of immutans and var2. Photosynth. Res. 2013, 116, 437–453. [Google Scholar] [CrossRef]
- Rodrigues, R.A.; Silva-Filho, M.C.; Cline, K. FtsH2 and FtsH5: Two homologous subunits use different integration mechanisms leading to the same thylakoid multimeric complex. Plant J. 2011, 65, 600–609. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Park, S.; Rodermel, S.R. The Arabidopsis FtsH metalloprotease gene family: Interchangeability of subunits in chloroplast oligomeric complexes. Plant J. 2004, 37, 864–876. [Google Scholar] [CrossRef]
- Zaltsman, A.; Ori, N.; Adam, Z. Two types of FtsH protease subunits are required for chloroplast biogenesis and photosystem II repair in Arabidopsis. Plant Cell 2005, 17, 2782–2790. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.; Song, J.; Wu, Y.; Zhuo, C.; Wen, J.; Yi, B.; Ma, C.; Shen, J.; Fu, T.; Tu, J. Brassica evolution of essential BnaFtsH1 genes involved in the PSII repair cycle and loss of FtsH5. Plant Sci. 2022, 315, 111128. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Nelson, C.J.; Trösch, J.; Castleden, I.; Huang, S.; Millar, A.H. Protein degradation rate in Arabidopsis thaliana leaf growth and development. Plant Cell 2017, 29, 207–228. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Qi, Y.; Malnoë, A.; Choquet, Y.; Wollman, F.-A.; De Vitry, C. The high light response and redox control of thylakoid FtsH protease in Chlamydomonas reinhardtii. Mol. Plant 2017, 10, 99–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, Y.; Hyodo, K.; Sakamoto, W. The photosystem II repair cycle requires FtsH turnover through the EngA GTPase. Plant Physiol. 2018, 178, 596–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, C.; Yu, C.; Shen, Y.; Fang, X.; Chen, L.; Min, J.; Cheng, J.; Zhao, S.; Xu, M.; Luo, Y. Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization. Proc. Natl. Acad. Sci. USA 2014, 111, 5135–5140. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Jiang, H.; Liu, J.; Liu, Y.; Hu, X. Characteristics of TaFtsH6 gene and expression analysis under dry and heat stress in wheat. Triticeae Genom. Genet. 2021, 12, 5376. [Google Scholar] [CrossRef]
- Chen, J.H.; Chen, S.T.; He, N.Y.; Wang, Q.L.; Zhao, Y.; Gao, W.; Guo, F.Q. Nuclear-encoded synthesis of the D1 subunit of photosystem II increases photosynthetic efficiency and crop yield. Nat. Plants 2020, 6, 570–580. [Google Scholar] [CrossRef]
- Lamesch, P.; Berardini, T.Z.; Li, D.; Swarbreck, D.; Wilks, C.; Sasidharan, R.; Muller, R.; Dreher, K.; Alexander, D.L.; Garcia-Hernandez, M. The Arabidopsis Information Resource (TAIR): Improved gene annotation and new tools. Nucleic Acids Res. 2012, 40, D1202–D1210. [Google Scholar] [CrossRef]
- Fernandez-Pozo, N.; Menda, N.; Edwards, J.D.; Saha, S.; Tecle, I.Y.; Strickler, S.R.; Bombarely, A.; Fisher-York, T.; Pujar, A.; Foerster, H. The Sol Genomics Network (SGN)-from genotype to phenotype to breeding. Nucleic Acids Res. 2015, 43, D1036–D1041. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.; Tosatto, S.C.; Paladin, L.; Raj, S.; Richardson, L.J. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Bjellqvist, B.; Basse, B.; Olsen, E.; Celis, J.E. Reference points for comparisons of two-dimensional maps of proteins from different human cell types defined in a pH scale where isoelectric points correlate with polypeptide compositions. Electrophoresis 1994, 15, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Chou, K.; Shen, H. Plant-mPLoc: A top-down strategy to augment the power for predicting plant protein subcellular localization. PloS ONE 2010, 5, e11335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.; Clamp, M.; Barton, G.J. Jalview Version 2-a multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Schneider, S.; Roessli, D.; Excoffier, L. Arlequin: A software for population genetics data analysis. User Man. Ver. 2000, 2, 2496–2497. [Google Scholar]
- Liu, F.; Yu, H.; Deng, Y.; Zheng, J.; Liu, M.; Ou, L.; Yang, B.; Dai, X.; Ma, Y.; Feng, S. PepperHub, an informatics hub for the chili pepper research community. Mol. Plant 2017, 10, 1129–1132. [Google Scholar] [CrossRef]
- Wan, H.; Yuan, W.; Ruan, M.; Ye, Q.; Wang, R.; Li, Z.; Zhou, G.; Yao, Z.; Zhao, J.; Liu, S. Identification of reference genes for reverse transcription quantitative real-time PCR normalization in pepper (Capsicum annuum L.). Biochem. Biophys. Res. Commun. 2011, 416, 24–30. [Google Scholar] [CrossRef]
- Ye, S.; Hua, S.; Ma, T.; Ma, X.; Chen, Y.; Wu, L.; Zhao, L.; Yi, B.; Ma, C.; Tu, J. Genetic and multi-omics analyses reveal BnaA07.PAP2 In−184−317 as the key gene conferring anthocyanin-based color in Brassica napus flowers. J. Exp. Bot. 2022, 73, 6630–6645. [Google Scholar] [CrossRef]
- Liu, J.; Zhu, L.; Wang, B.; Wang, H.; Zhang, S.; Wen, J.; Ma, C.; Dai, C.; Tu, J.; Shen, J. BnA1.CER4 and BnC1.CER4 are redundantly involved in branched primary alcohols in the cuticle wax of Brassica napus. Theor. Appl. Genet. 2021, 134, 3051–3067. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Deng, Y.; Liu, D.; Wang, H.; Zhang, X.; Liu, T.; Wang, J.; Li, Y.; Ou, L.; Liu, F. Promoting virus-induced gene silencing of pepper genes by a heterologous viral silencing suppressor. Plant Biotechnol. J. 2021, 19, 2398–2400. [Google Scholar] [CrossRef] [PubMed]
- Yi, B.; Zeng, F.; Lei, S.; Chen, Y.; Yao, X.; Zhu, Y.; Wen, J.; Shen, J.; Ma, C.; Tu, J.; et al. Two duplicate CYP704B1-homologous genes BnMs1 and BnMs2 are required for pollen exine formation and tapetal development in Brassica napus. Plant J. 2010, 63, 925–938. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, Y.; Shi, C.; Huang, Z.; Zhang, Y.; Li, S.; Li, Y.; Ye, J.; Yu, C.; Li, Z. SOAPnuke: A MapReduce acceleration-supported software for integrated quality control and preprocessing of high-throughput sequencing data. Gigascience 2018, 7, gix120. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Langmead, B. Aligning short sequencing reads with Bowtie. Curr. Protoc. Bioinf. 2010, 32, 11.17.11–11.17.14. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinf. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, K.; Li, N.; Zhang, Y.; Gao, S.; Yin, Y.; Yao, M.; Wang, F. Silencing of Pepper CaFtsH1 or CaFtsH8 Genes Alters Normal Leaf Development. Int. J. Mol. Sci. 2023, 24, 4927. https://doi.org/10.3390/ijms24054927
Xu K, Li N, Zhang Y, Gao S, Yin Y, Yao M, Wang F. Silencing of Pepper CaFtsH1 or CaFtsH8 Genes Alters Normal Leaf Development. International Journal of Molecular Sciences. 2023; 24(5):4927. https://doi.org/10.3390/ijms24054927
Chicago/Turabian StyleXu, Kai, Ning Li, Yiwen Zhang, Shenghua Gao, Yanxu Yin, Minghua Yao, and Fei Wang. 2023. "Silencing of Pepper CaFtsH1 or CaFtsH8 Genes Alters Normal Leaf Development" International Journal of Molecular Sciences 24, no. 5: 4927. https://doi.org/10.3390/ijms24054927