
Apelin Enhances the Effects of Fusobacterium nucleatum on Periodontal Ligament Cells In Vitro
 , , , and
, , , and 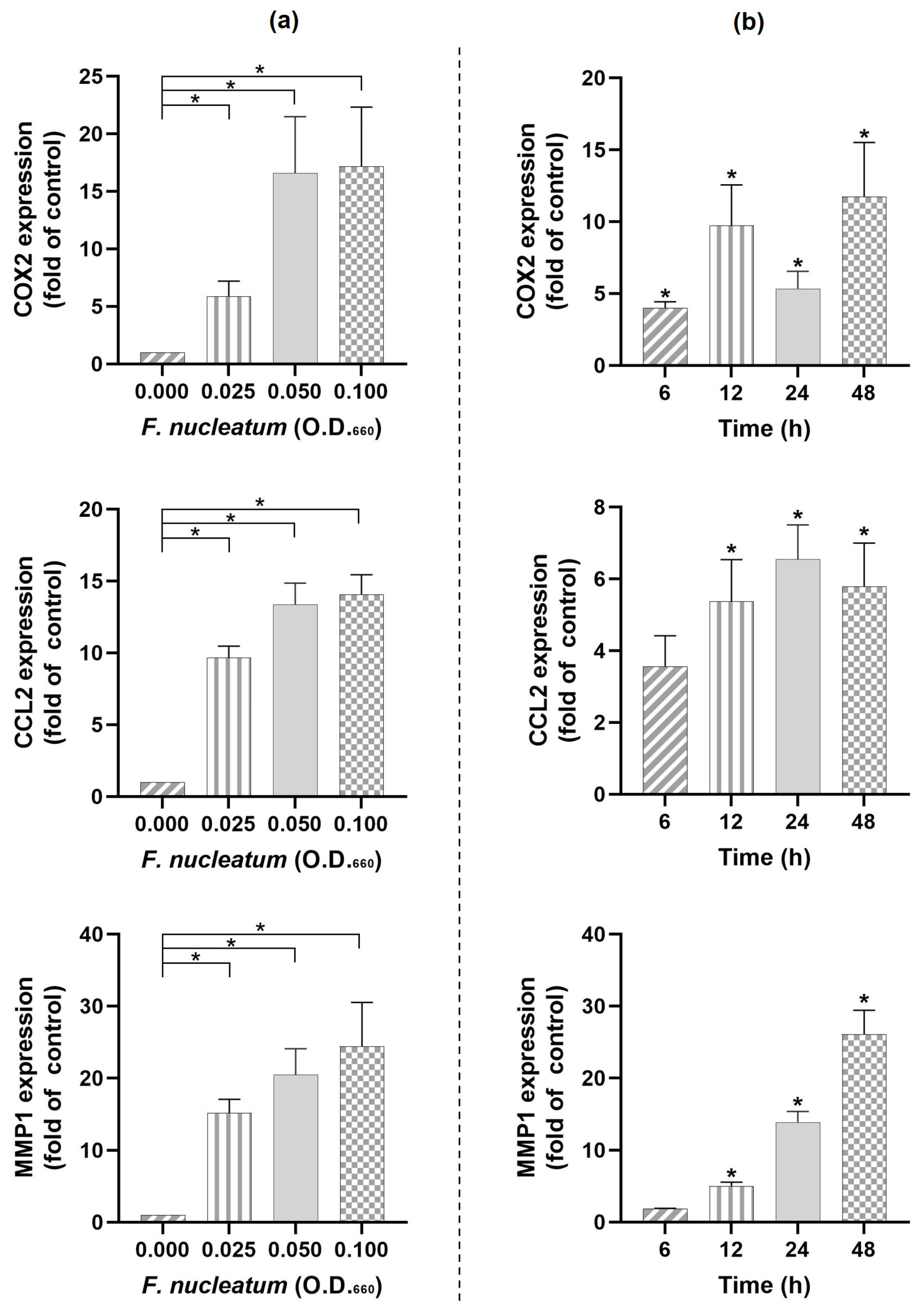{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Regulation of COX2, CCL2, and MMP1 Expressions by F. nucleatum
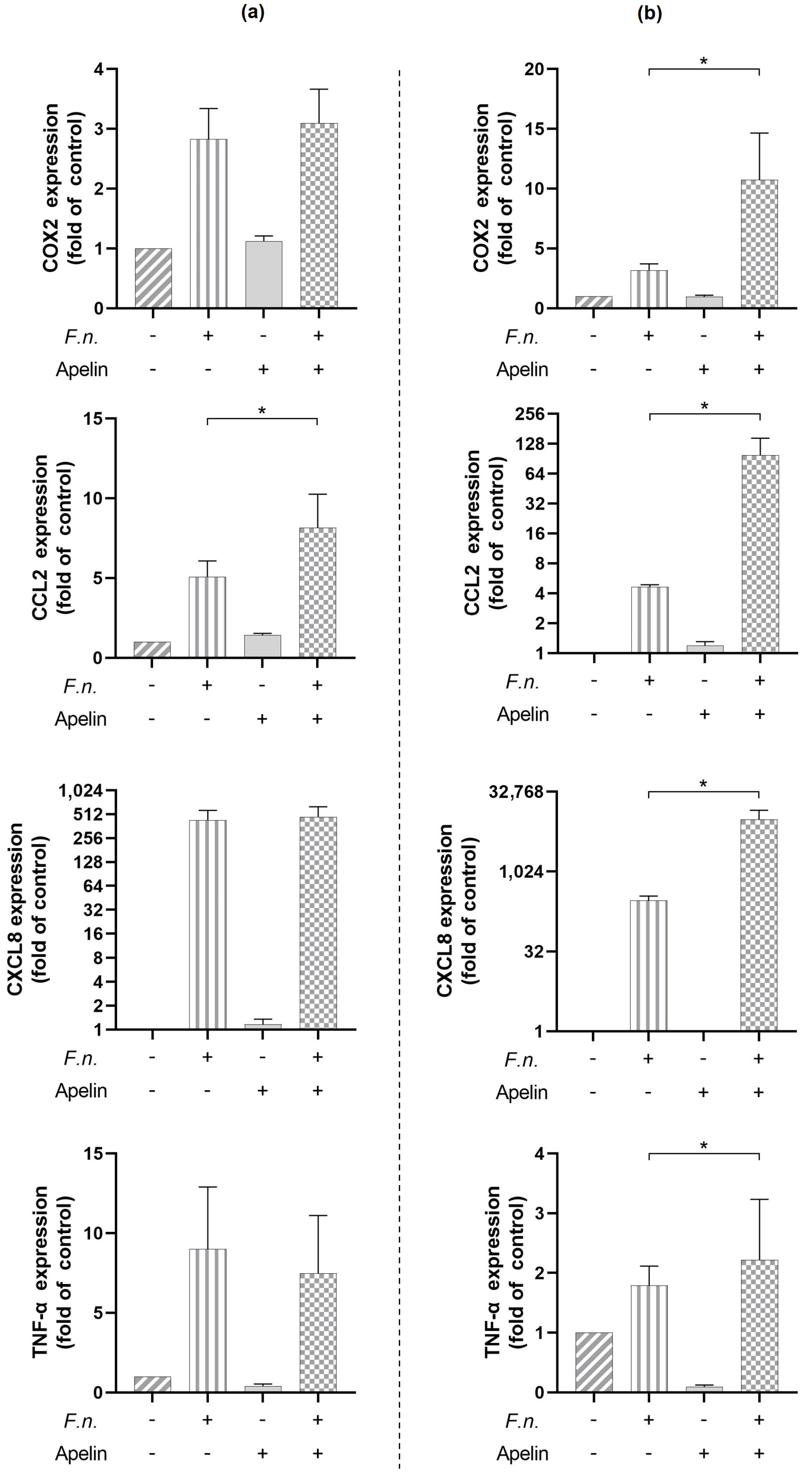
2.2. Modulatory Effects of Apelin on Pro-Inflammatory Actions by F. nucleatum
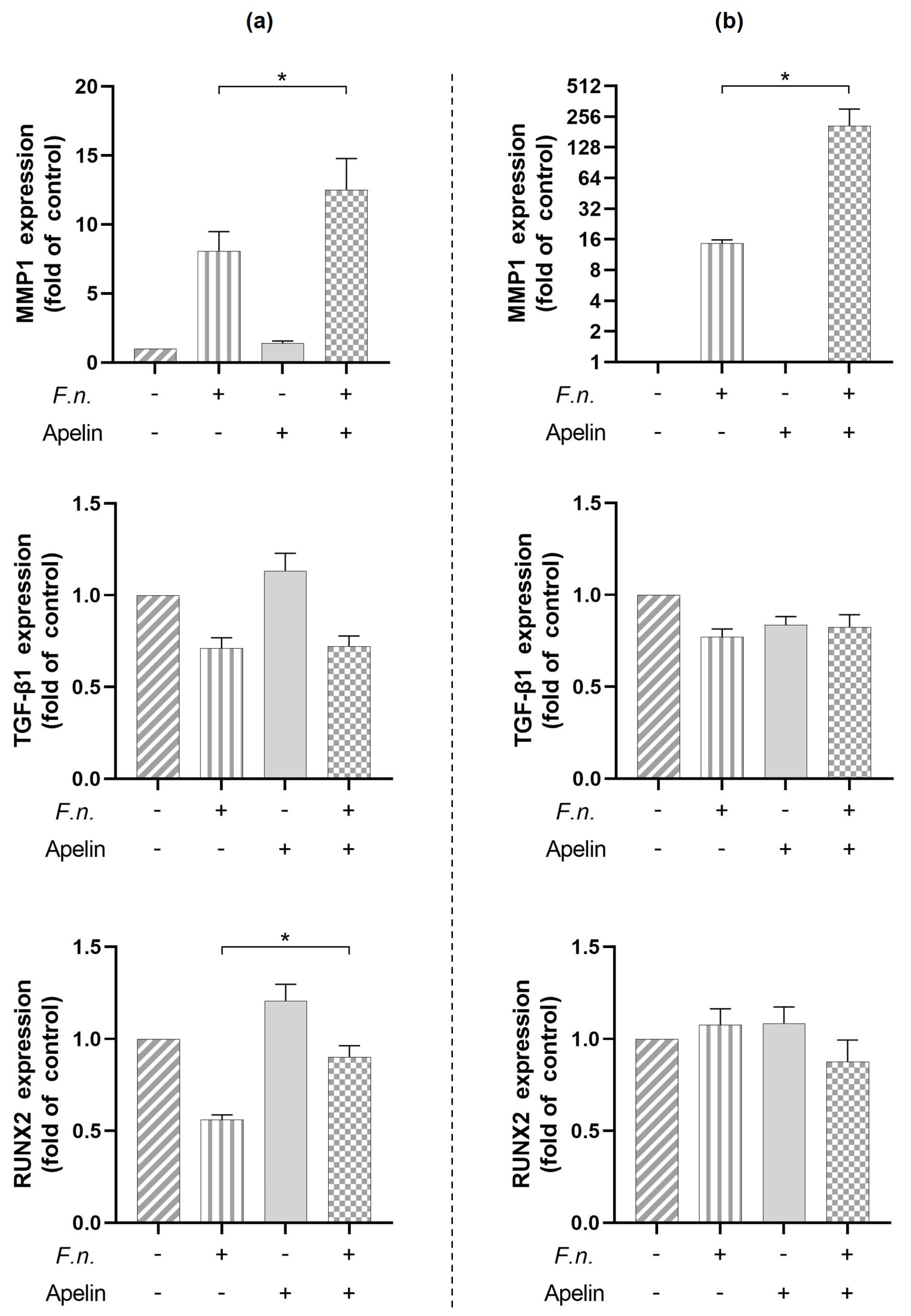
2.3. Modulatory Effects of Apelin on Markers Involved in Soft and Hard Tissue Turnover
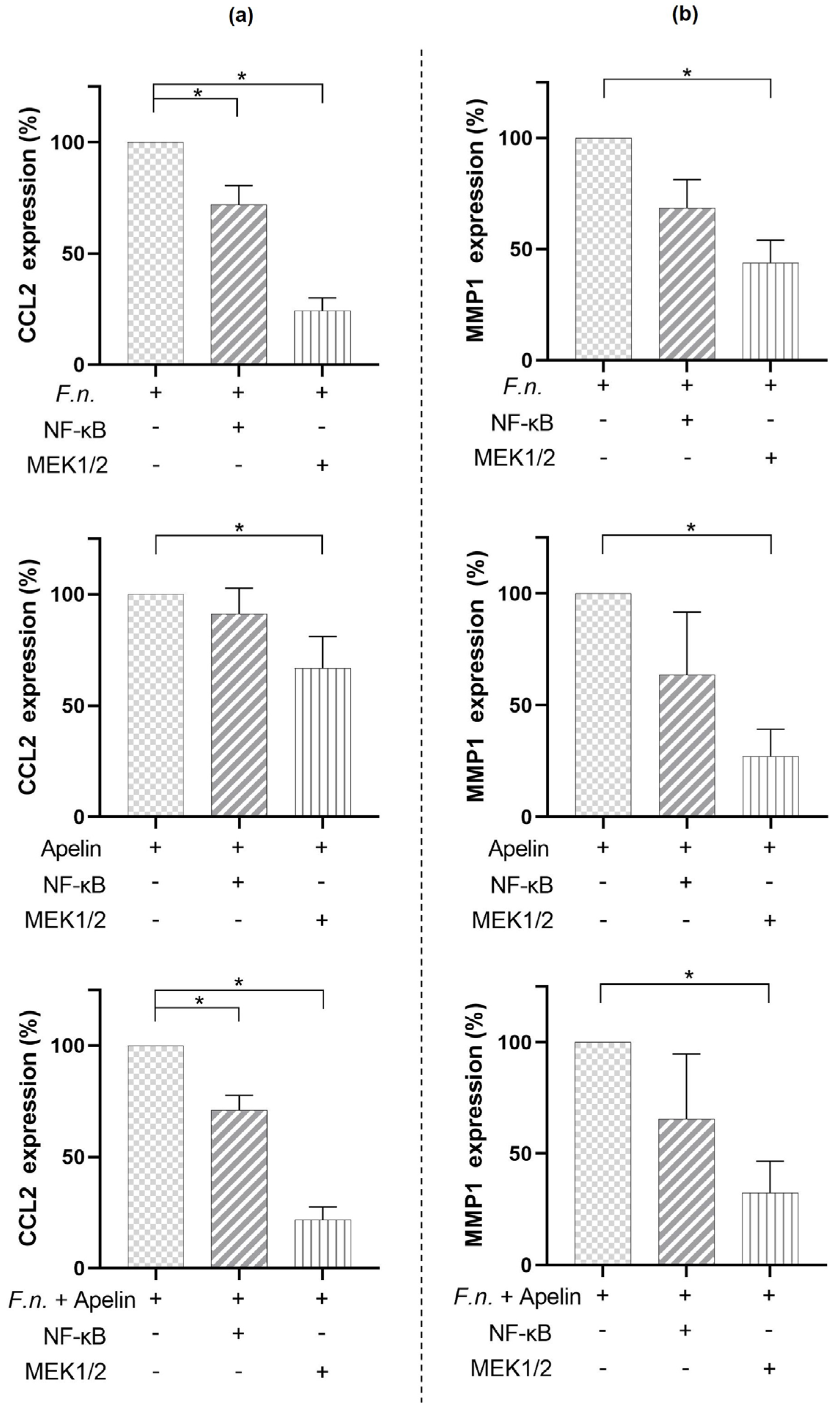
2.4. Involvement of Signaling Pathways in the Modulatory Effects of F. nucleatum and/or Apelin on CCL2 and MMP1 Expressions
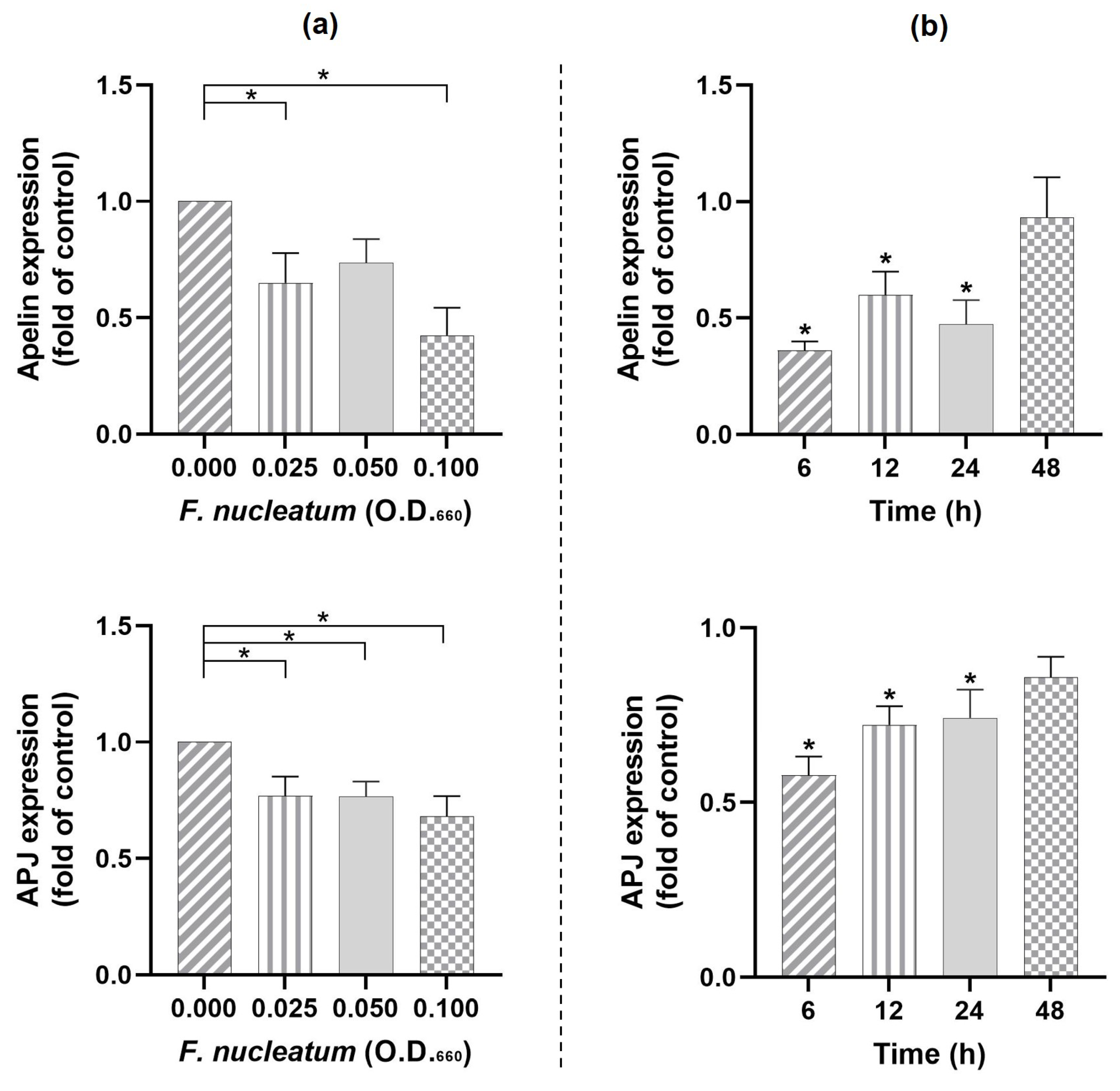
2.5. Effects of F. nucleatum on Apelin and Its Receptor
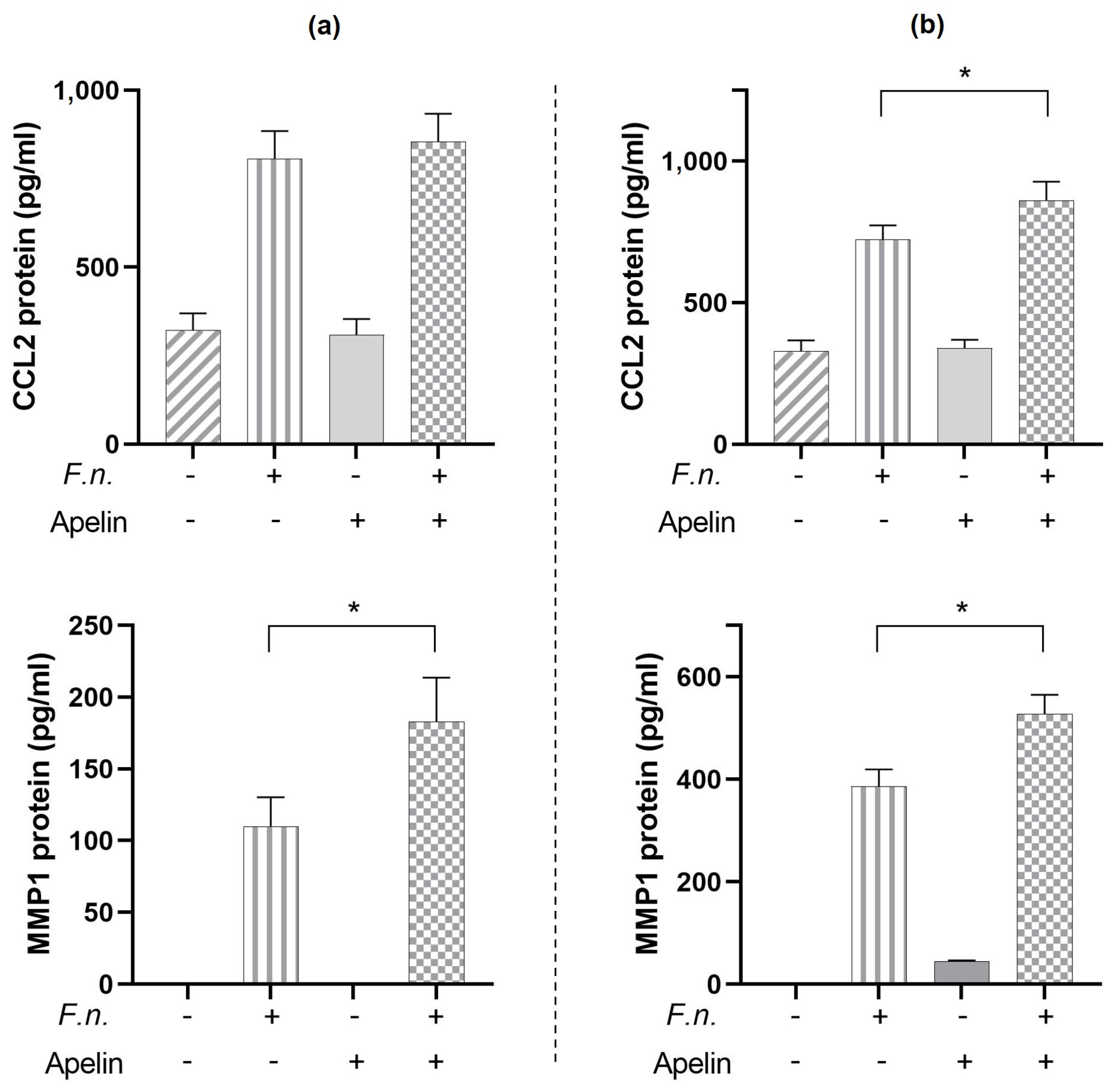
2.6. Modulatory Effects of Apelin on CCL2 and MMP1 Protein Induced by F. nucleatum
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Real-Time PCR
4.3. ELISA
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Papapanou, P.N.; Sanz, M.; Buduneli, N.; Dietrich, T.; Feres, M.; Fine, D.H.; Flemmig, T.F.; Garcia, R.; Giannobile, W.V.; Graziani, F.; et al. Periodontitis: Consensus Report of Workgroup 2 of the 2017 World Workshop on the Classification of Periodontal and Peri-Implant Diseases and Conditions. J. Clin. Periodontol. 2018, 45, S162–S170. [Google Scholar] [CrossRef] [PubMed]
- Tatakis, D.N.; Kumar, P.S. Etiology and Pathogenesis of Periodontal Diseases. Dent. Clin. N. Am. 2005, 49, 491–516. [Google Scholar] [CrossRef] [PubMed]
- Stöhr, J.; Barbaresko, J.; Neuenschwander, M.; Schlesinger, S. Bidirectional Association between Periodontal Disease and Diabetes Mellitus: A Systematic Review and Meta-Analysis of Cohort Studies. Sci. Rep. 2021, 11, 13686. [Google Scholar] [CrossRef] [PubMed]
- Sanz, M.; Marco Del Castillo, A.; Jepsen, S.; Gonzalez-Juanatey, J.R.; D’Aiuto, F.; Bouchard, P.; Chapple, I.; Dietrich, T.; Gotsman, I.; Graziani, F.; et al. Periodontitis and Cardiovascular Diseases: Consensus Report. J. Clin. Periodontol. 2020, 47, 268–288. [Google Scholar] [CrossRef]
- Muñoz Aguilera, E.; Suvan, J.; Buti, J.; Czesnikiewicz-Guzik, M.; Barbosa Ribeiro, A.; Orlandi, M.; Guzik, T.J.; Hingorani, A.D.; Nart, J.; D’Aiuto, F. Periodontitis Is Associated with Hypertension: A Systematic Review and Meta-Analysis. Cardiovasc. Res. 2020, 116, 28–39. [Google Scholar] [CrossRef] [Green Version]
- Suvan, J.; D’Aiuto, F.; Moles, D.R.; Petrie, A.; Donos, N. Association between Overweight/Obesity and Periodontitis in Adults. A Systematic Review. Obes. Rev. 2011, 12, e381–e404. [Google Scholar] [CrossRef]
- Gobin, R.; Tian, D.; Liu, Q.; Wang, J. Periodontal Diseases and the Risk of Metabolic Syndrome: An Updated Systematic Review and Meta-Analysis. Front. Endocrinol. 2020, 11, 336. [Google Scholar] [CrossRef]
- Monsarrat, P.; Blaizot, A.; Kémoun, P.; Ravaud, P.; Nabet, C.; Sixou, M.; Vergnes, J.-N. Clinical Research Activity in Periodontal Medicine: A Systematic Mapping of Trial Registers. J. Clin. Periodontol. 2016, 43, 390–400. [Google Scholar] [CrossRef]
- World Health Organization. Obesity and Overweight. Available online: https://www.WHO.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 30 November 2022).
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in Inflammation and Metabolic Disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef]
- Park, H.S.; Park, J.Y.; Yu, R. Relationship of Obesity and Visceral Adiposity with Serum Concentrations of CRP, TNF-Alpha and IL-6. Diabetes Res. Clin. Pract. 2005, 69, 29–35. [Google Scholar] [CrossRef]
- Jepsen, S.; Suvan, J.; Deschner, J. The Association of Periodontal Diseases with Metabolic Syndrome and Obesity. Periodontology 2020, 83, 125–153. [Google Scholar] [CrossRef] [PubMed]
- Fasshauer, M.; Blüher, M. Adipokines in Health and Disease. Trends Pharm. Sci. 2015, 36, 461–470. [Google Scholar] [CrossRef] [PubMed]
- de Leal, V.O.; Mafra, D. Adipokines in Obesity. Clin. Chim. Acta 2013, 419, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, A.V.B.; Nokhbehsaim, M.; Tekin, S.; de Molon, R.S.; Spolidorio, L.C.; Memmert, S.; Damanaki, A.; Jäger, A.; Eick, S.; Deschner, J.; et al. Resistin Is Increased in Periodontal Cells and Tissues: In Vitro and In Vivo Studies. Mediat. Inflamm. 2020, 2020, 9817095. [Google Scholar] [CrossRef] [Green Version]
- Nogueira, A.V.B.; Nokhbehsaim, M.; Eick, S.; Bourauel, C.; Jäger, A.; Jepsen, S.; Cirelli, J.A.; Deschner, J. Regulation of Visfatin by Microbial and Biomechanical Signals in PDL Cells. Clin. Oral Investig. 2014, 18, 171–178. [Google Scholar] [CrossRef]
- Nokhbehsaim, M.; Keser, S.; Nogueira, A.V.B.; Jäger, A.; Jepsen, S.; Cirelli, J.A.; Bourauel, C.; Eick, S.; Deschner, J. Leptin Effects on the Regenerative Capacity of Human Periodontal Cells. Int. J. Endocrinol. 2014, 2014, 180304. [Google Scholar] [CrossRef] [Green Version]
- Nokhbehsaim, M.; Keser, S.; Nogueira, A.V.B.; Cirelli, J.A.; Jepsen, S.; Jäger, A.; Eick, S.; Deschner, J. Beneficial Effects of Adiponectin on Periodontal Ligament Cells under Normal and Regenerative Conditions. J. Diabetes Res. 2014, 2014, 796565. [Google Scholar] [CrossRef] [Green Version]
- Blüher, M.; Mantzoros, C.S. From Leptin to Other Adipokines in Health and Disease: Facts and Expectations at the Beginning of the 21st Century. Metabolism 2015, 64, 131–145. [Google Scholar] [CrossRef]
- Villarreal-Molina, M.T.; Antuna-Puente, B. Adiponectin: Anti-Inflammatory and Cardioprotective Effects. Biochimie 2012, 94, 2143–2149. [Google Scholar] [CrossRef] [PubMed]
- Al-Suhaimi, E.A.; Shehzad, A. Leptin, Resistin and Visfatin: The Missing Link between Endocrine Metabolic Disorders and Immunity. Eur. J. Med. Res. 2013, 18, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatemoto, K.; Hosoya, M.; Habata, Y.; Fujii, R.; Kakegawa, T.; Zou, M.X.; Kawamata, Y.; Fukusumi, S.; Hinuma, S.; Kitada, C.; et al. Isolation and Characterization of a Novel Endogenous Peptide Ligand for the Human APJ Receptor. Biochem. Biophys. Res. Commun. 1998, 251, 471–476. [Google Scholar] [CrossRef] [PubMed]
- O’Dowd, B.F.; Heiber, M.; Chan, A.; Heng, H.H.; Tsui, L.C.; Kennedy, J.L.; Shi, X.; Petronis, A.; George, S.R.; Nguyen, T. A Human Gene That Shows Identity with the Gene Encoding the Angiotensin Receptor Is Located on Chromosome 11. Gene 1993, 136, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Castan-Laurell, I.; Dray, C.; Attané, C.; Duparc, T.; Knauf, C.; Valet, P. Apelin, Diabetes, and Obesity. Endocrine 2011, 40, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wattanachanya, L.; Lu, W.-D.; Kundu, R.K.; Wang, L.; Abbott, M.J.; O’Carroll, D.; Quertermous, T.; Nissenson, R.A. Increased Bone Mass in Mice Lacking the Adipokine Apelin. Endocrinology 2013, 154, 2069–2080. [Google Scholar] [CrossRef] [Green Version]
- Xie, H.; Tang, S.; Cui, R.; Huang, J.; Ren, X.; Yuan, L.; Lu, Y.; Yang, M.; Zhou, H.; Wu, X.; et al. Apelin and Its Receptor Are Expressed in Human Osteoblasts. Regul. Pept. 2006, 134, 118–125. [Google Scholar] [CrossRef]
- Li, C.; Cheng, H.; Adhikari, B.K.; Wang, S.; Yang, N.; Liu, W.; Sun, J.; Wang, Y. The Role of Apelin-APJ System in Diabetes and Obesity. Front. Endocrinol. 2022, 13, 820002. [Google Scholar] [CrossRef]
- Boucher, J.; Masri, B.; Daviaud, D.; Gesta, S.; Guigné, C.; Mazzucotelli, A.; Castan-Laurell, I.; Tack, I.; Knibiehler, B.; Carpéné, C.; et al. Apelin, a Newly Identified Adipokine Up-Regulated by Insulin and Obesity. Endocrinology 2005, 146, 1764–1771. [Google Scholar] [CrossRef]
- Hirani, T.; Kumar, S.; Patel, V.; Hirani, S.; Mohammed, I.; Shishoo, D. Expression of Apelin among the Individuals of Chronic Periodontitis, with and without Type Ii Diabetes Mellitus: A Study Using Enzyme-Linked Immunosorbent Assay. Adv. Hum. Biol. 2020, 10, 182. [Google Scholar] [CrossRef]
- Sarhat, E.R.; Rmaid, Z.J.; Jabir, T.H. Changes of Salivary Interleukine-17, Apelin, Omentin and Vaspin Levels in Normal Subjects and Diabetic Patients with Chronic Periodontitis. Ann. Trop. Med. Public Health 2020, 23, 135–141. [Google Scholar] [CrossRef]
- de Visser, Y.P.; Walther, F.J.; Laghmani, E.H.; van der Laarse, A.; Wagenaar, G.T.M. Apelin Attenuates Hyperoxic Lung and Heart Injury in Neonatal Rats. Am. J. Respir. Crit. Care Med. 2010, 182, 1239–1250. [Google Scholar] [CrossRef]
- Chen, L.; Tao, Y.; Jiang, Y. Apelin Activates the Expression of Inflammatory Cytokines in Microglial BV2 Cells via PI-3K/Akt and MEK/Erk Pathways. Sci. China Life Sci. 2015, 58, 531–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deschner, J.; Eick, S.; Damanaki, A.; Nokhbehsaim, M. The Role of Adipokines in Periodontal Infection and Healing. Mol. Oral Microbiol. 2014, 29, 258–269. [Google Scholar] [CrossRef] [PubMed]
- Damanaki, A.; Nokhbehsaim, M.; Eick, S.; Götz, W.; Winter, J.; Wahl, G.; Jäger, A.; Jepsen, S.; Deschner, J. Regulation of NAMPT in Human Gingival Fibroblasts and Biopsies. Mediat. Inflamm. 2014, 2014, 912821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nokhbehsaim, M.; Eick, S.; Nogueira, A.V.B.; Hoffmann, P.; Herms, S.; Fröhlich, H.; Jepsen, S.; Jäger, A.; Cirelli, J.A.; Deschner, J. Stimulation of MMP-1 and CCL2 by NAMPT in PDL Cells. Mediat. Inflamm. 2013, 2013, 437123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogueira, A.V.B.; Nokhbehsaim, M.; Damanaki, A.; Eick, S.; Kirschneck, C.; Schröder, A.; Jantsch, J.; Deschner, J. Filifactor Alocis and Tumor Necrosis Factor-Alpha Stimulate Synthesis of Visfatin by Human Macrophages. Int. J. Mol. Sci. 2021, 22, 1235. [Google Scholar] [CrossRef]
- Kraus, D.; Winter, J.; Jepsen, S.; Jäger, A.; Meyer, R.; Deschner, J. Interactions of Adiponectin and Lipopolysaccharide from Porphyromonas Gingivalis on Human Oral Epithelial Cells. PLoS ONE 2012, 7, e30716. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.; Song, W.-H.; Kim, S.-J.; Kim, Y.-G.; Ryu, J.-H. Apelin-APJ Axis Inhibits TNF-Alpha-Mediated Expression of Genes Involved in the Inflammatory Response in Periodontal Ligament Cells. Int. J. Oral Biol. 2019, 44, 182–190. [Google Scholar] [CrossRef]
- Llama-Palacios, A.; Potupa, O.; Sánchez, M.C.; Figuero, E.; Herrera, D.; Sanz, M. Proteomic Analysis of Fusobacterium Nucleatum Growth in Biofilm versus Planktonic State. Mol. Oral Microbiol. 2020, 35, 168–180. [Google Scholar] [CrossRef]
- Patini, R.; Staderini, E.; Lajolo, C.; Lopetuso, L.; Mohammed, H.; Rimondini, L.; Rocchetti, V.; Franceschi, F.; Cordaro, M.; Gallenzi, P. Relationship between Oral Microbiota and Periodontal Disease: A Systematic Review. Eur. Rev. Med. Pharm. Sci. 2018, 22, 5775–5788. [Google Scholar] [CrossRef]
- Chaushu, S.; Wilensky, A.; Gur, C.; Shapira, L.; Elboim, M.; Halftek, G.; Polak, D.; Achdout, H.; Bachrach, G.; Mandelboim, O. Direct Recognition of Fusobacterium Nucleatum by the NK Cell Natural Cytotoxicity Receptor NKp46 Aggravates Periodontal Disease. PLoS Pathog. 2012, 8, e1002601. [Google Scholar] [CrossRef] [Green Version]
- Settem, R.P.; El-Hassan, A.T.; Honma, K.; Stafford, G.P.; Sharma, A. Fusobacterium Nucleatum and Tannerella Forsythia Induce Synergistic Alveolar Bone Loss in a Mouse Periodontitis Model. Infect. Immun. 2012, 80, 2436–2443. [Google Scholar] [CrossRef] [Green Version]
- Polak, D.; Wilensky, A.; Shapira, L.; Halabi, A.; Goldstein, D.; Weiss, E.I.; Houri-Haddad, Y. Mouse Model of Experimental Periodontitis Induced by Porphyromonas Gingivalis / Fusobacterium Nucleatum Infection: Bone Loss and Host Response. J. Clin. Periodontol. 2009, 36, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Marsh, P.D.; Al-Hebshi, N.N. SMDI: An Index for Measuring Subgingival Microbial Dysbiosis. J. Dent. Res. 2022, 101, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Rath-Deschner, B.; Nogueira, A.V.B.; Beisel-Memmert, S.; Nokhbehsaim, M.; Eick, S.; Cirelli, J.A.; Deschner, J.; Jäger, A.; Damanaki, A. Interaction of Periodontitis and Orthodontic Tooth Movement-an in Vitro and in Vivo Study. Clin. Oral Investig. 2022, 26, 171–181. [Google Scholar] [CrossRef]
- Rath-Deschner, B.; Memmert, S.; Damanaki, A.; de Molon, R.S.; Nokhbehsaim, M.; Eick, S.; Kirschneck, C.; Cirelli, J.A.; Deschner, J.; Jäger, A.; et al. CXCL5, CXCL8, and CXCL10 Regulation by Bacteria and Mechanical Forces in Periodontium. Ann. Anat. 2021, 234, 151648. [Google Scholar] [CrossRef] [PubMed]
- Memmert, S.; Damanaki, A.; Nogueira, A.V.B.; Eick, S.; Nokhbehsaim, M.; Papadopoulou, A.K.; Till, A.; Rath, B.; Jepsen, S.; Götz, W.; et al. Role of Cathepsin S in Periodontal Inflammation and Infection. Mediat. Inflamm. 2017, 2017, 4786170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhang, L.; Li, P.; Zheng, Y.; Yang, Y.; Ji, S. Apelin/APJ System in Inflammation. Int. Immunopharmacol. 2022, 109, 108822. [Google Scholar] [CrossRef] [PubMed]
- Antushevich, H.; Wójcik, M. Review: Apelin in Disease. Clin. Chim. Acta 2018, 483, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhu, X.; Liang, G.-X.; Cui, R.-R.; Liu, Y.; Wu, S.-S.; Liang, Q.-H.; Liu, G.-Y.; Jiang, Y.; Liao, X.-B.; et al. Apelin–APJ Induces ICAM-1, VCAM-1 and MCP-1 Expression via NF-ΚB/JNK Signal Pathway in Human Umbilical Vein Endothelial Cells. Amino Acids 2012, 43, 2125–2136. [Google Scholar] [CrossRef]
- Liu, J.; Liu, M.; Chen, L. Novel Pathogenesis: Regulation of Apoptosis by Apelin/APJ System. Acta Biochim. Et Biophys. Sin. (Shanghai) 2017, 49, 471–478. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Cao, J.; Chen, L. Apelin/APJ System: A Novel Therapeutic Target for Oxidative Stress-Related Inflammatory Diseases (Review). Int. J. Mol. Med. 2016, 37, 1159–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Tao, Y.; Feng, J.; Jiang, Y.R. Apelin Protects Primary Rat Retinal Pericytes from Chemical Hypoxia-Induced Apoptosis. J. Ophthalmol. 2015, 2015, 186946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rayalam, S.; Della-Fera, M.A.; Krieg, P.A.; Cox, C.M.; Robins, A.; Baile, C.A. A Putative Role for Apelin in the Etiology of Obesity. Biochem. Biophys. Res. Commun. 2008, 368, 815–819. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.-Y.; Yu, T.-Y.; Wei, J.-N.; Hung, C.-S.; Lin, M.-S.; Liao, Y.-J.; Pei, D.; Su, C.-C.; Lu, K.-C.; Liu, P.-H.; et al. Plasma Apelin: A Novel Biomarker for Predicting Diabetes. Clin. Chim. Acta 2014, 435, 18–23. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cores Ziskoven, P.; Nogueira, A.V.B.; Gutierrez, L.S.; Weusmann, J.; Eick, S.; Buduneli, N.; Deschner, J. Apelin Enhances the Effects of Fusobacterium nucleatum on Periodontal Ligament Cells In Vitro. Int. J. Mol. Sci. 2023, 24, 4733. https://doi.org/10.3390/ijms24054733
Cores Ziskoven P, Nogueira AVB, Gutierrez LS, Weusmann J, Eick S, Buduneli N, Deschner J. Apelin Enhances the Effects of Fusobacterium nucleatum on Periodontal Ligament Cells In Vitro. International Journal of Molecular Sciences. 2023; 24(5):4733. https://doi.org/10.3390/ijms24054733
Chicago/Turabian StyleCores Ziskoven, Pablo, Andressa V. B. Nogueira, Lorena S. Gutierrez, Jens Weusmann, Sigrun Eick, Nurcan Buduneli, and James Deschner. 2023. "Apelin Enhances the Effects of Fusobacterium nucleatum on Periodontal Ligament Cells In Vitro" International Journal of Molecular Sciences 24, no. 5: 4733. https://doi.org/10.3390/ijms24054733