
MiR-146a in ALS: Contribution to Early Peripheral Nerve Degeneration and Relevance as Disease Biomarker
 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Results
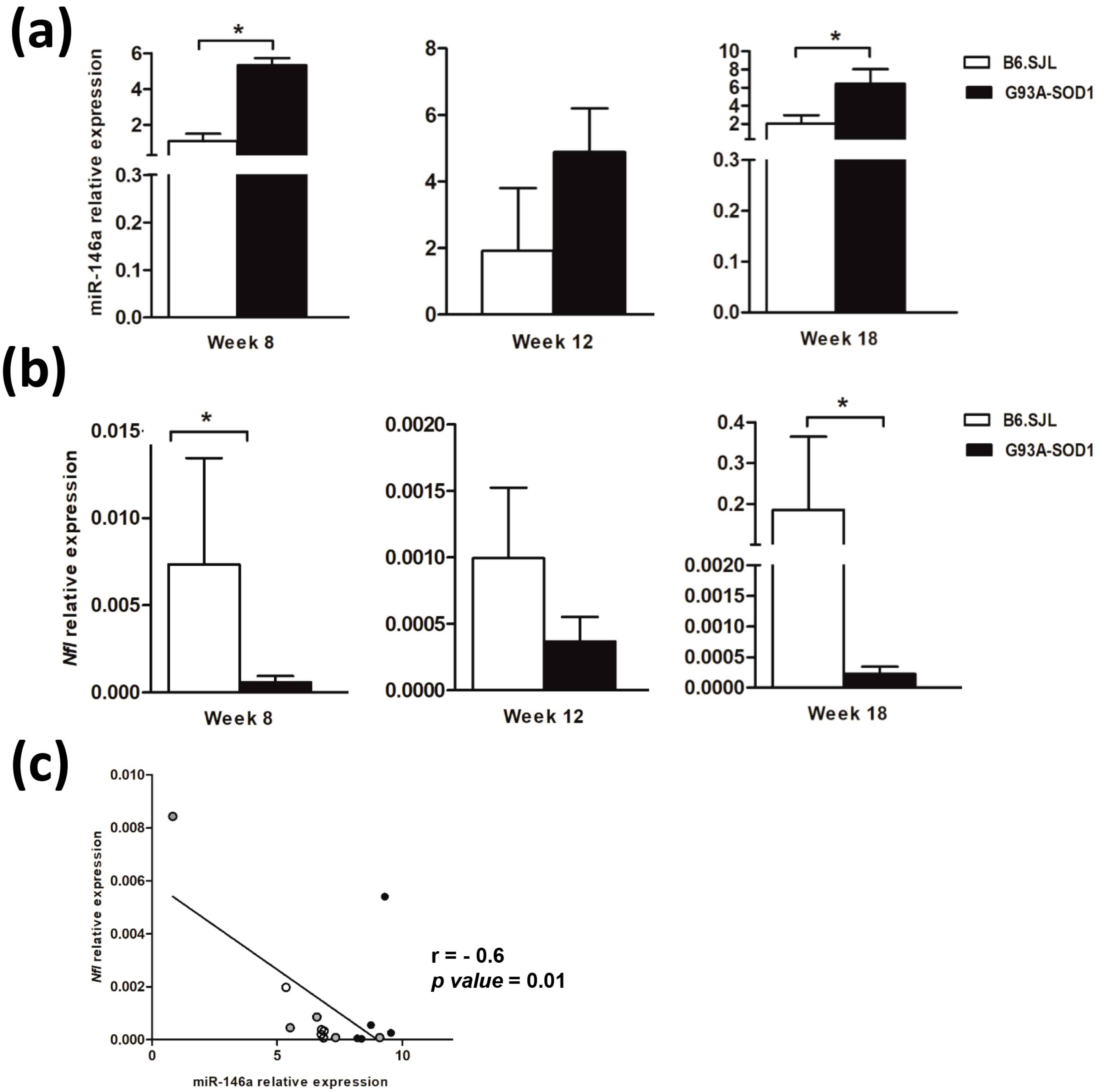
2.1. MiR-146a Increases and Nfl Gene Expression Decreases in Sciatic Nerve of G93A-SOD1 Mice during Disease Progression
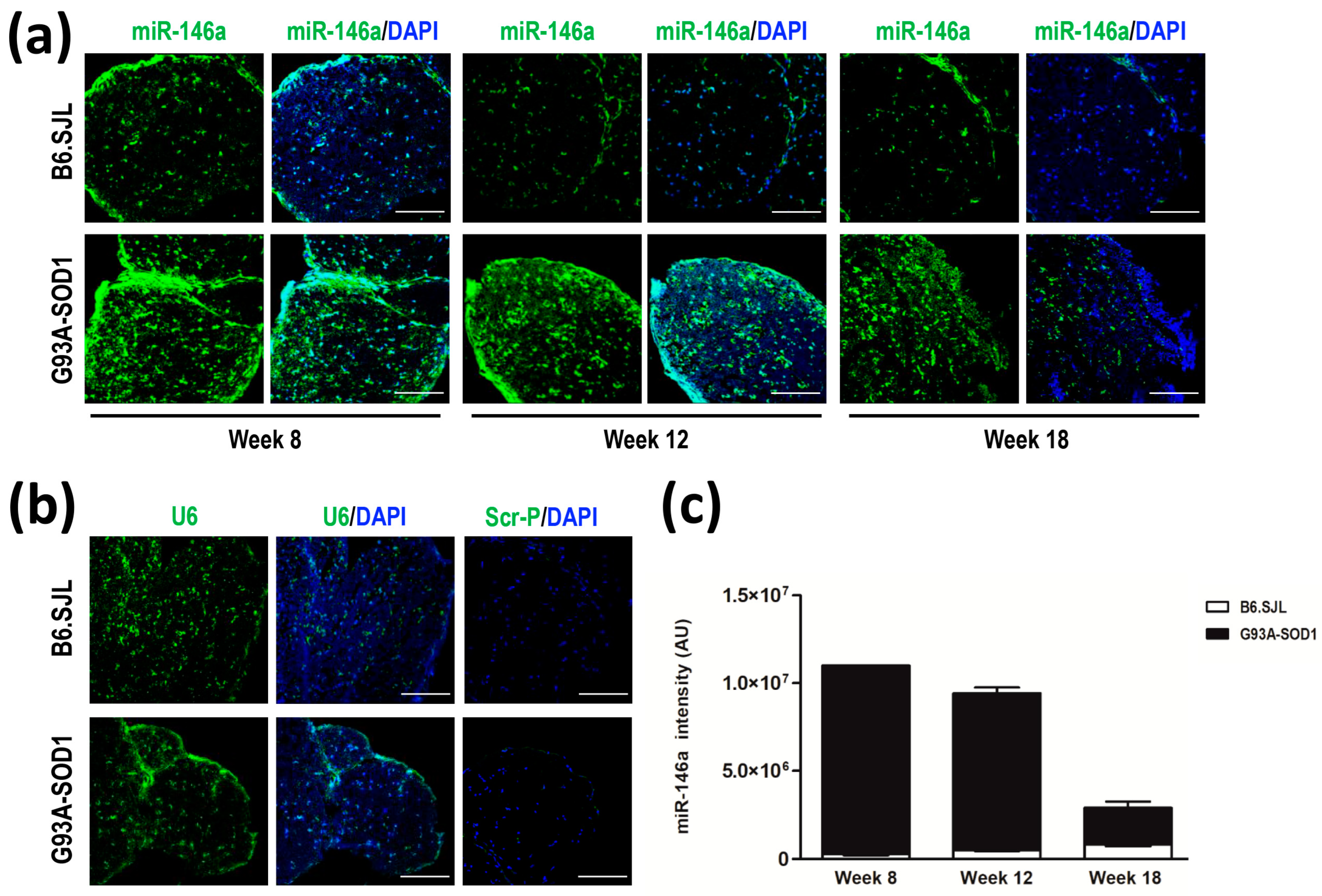
2.2. Increased miR-146a Levels in Sciatic Nerve of G93A-SOD1 Mice by In Situ Hybridization
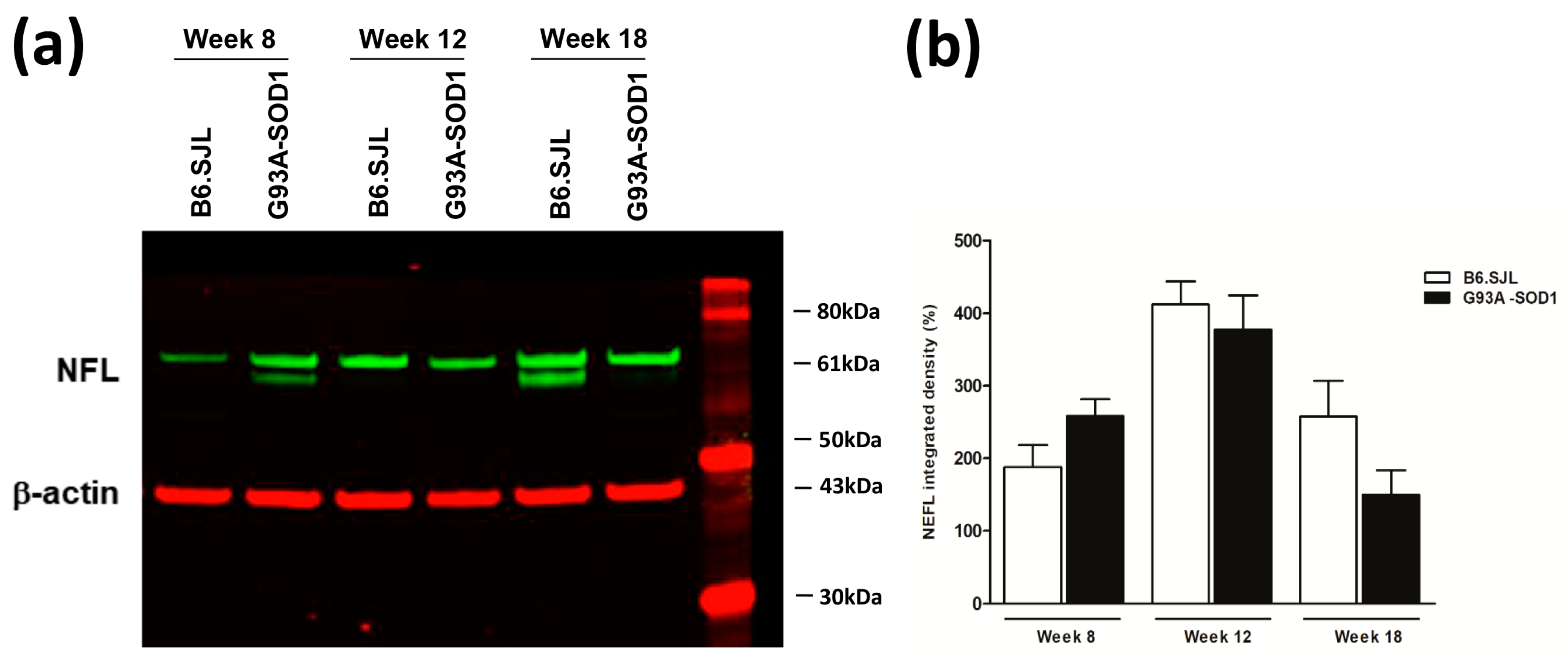
2.3. NFL Protein Decrease in Sciatic Nerve of G93A-SOD1 Mice
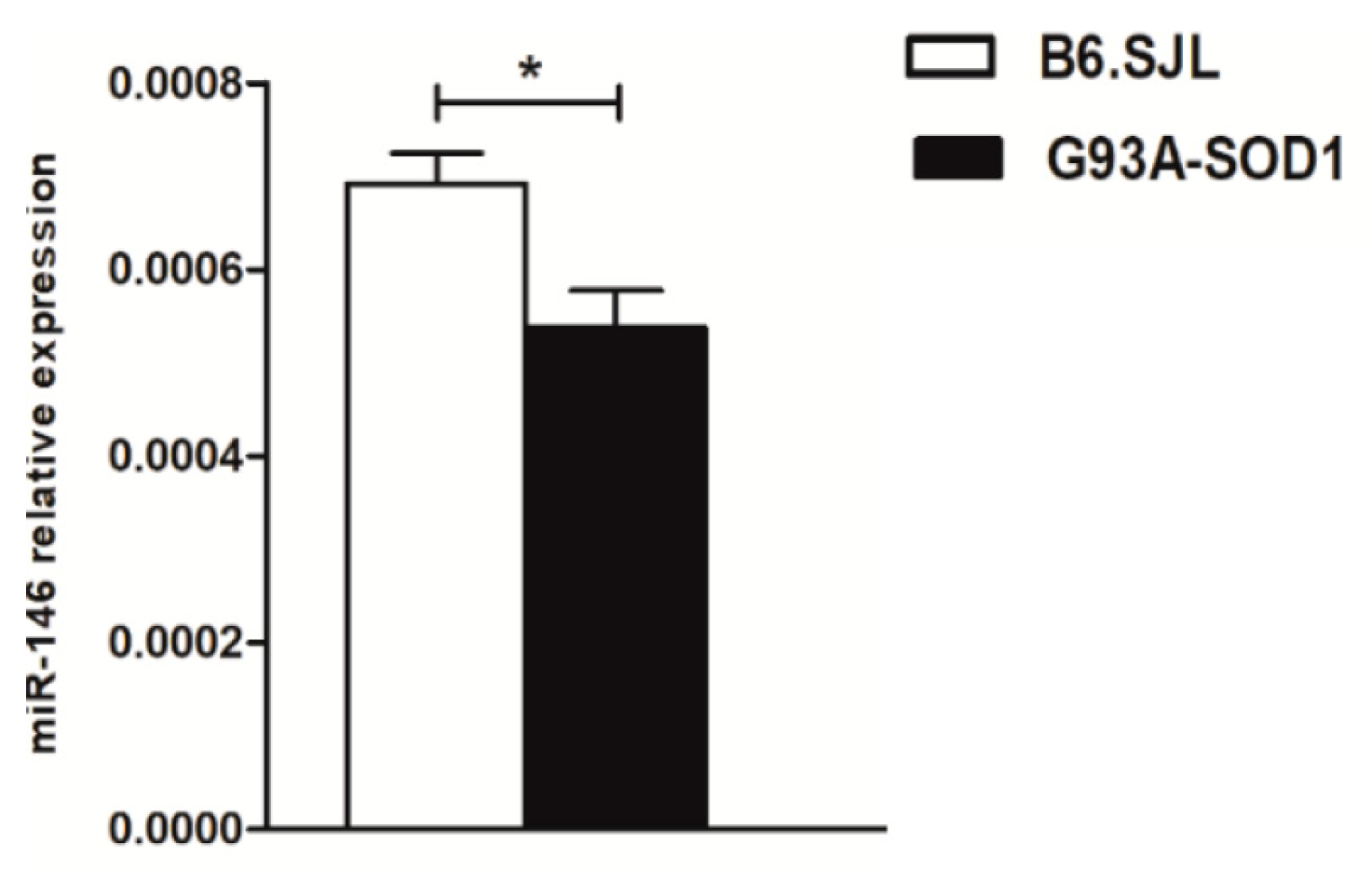
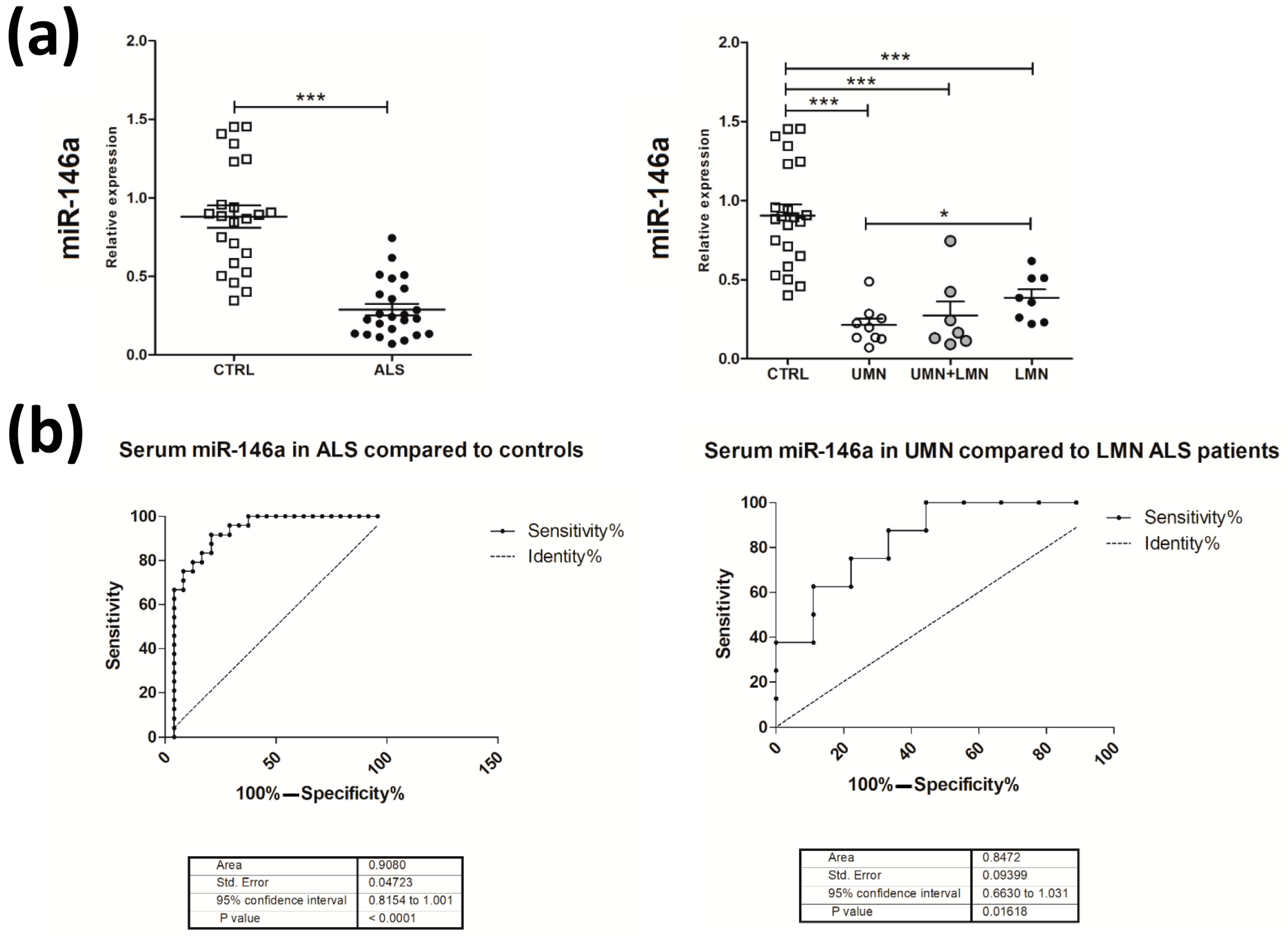
2.4. Decreased Expression of miR-146a in Serum Samples of G93A-SOD1 Animal Model and Human ALS Patients
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. NSC-34 Motor Neuron-like Cell Culture and Differentiation
4.3. Transfection of miR-146-5p Inhibitor
4.4. Quantitative Real-Time PCR to Assess MiR-146a Expression in Mouse Sciatic Nerve Tissue
4.5. Fluorescent In Situ Hybridization to Detect MiR-146a in Mouse Sciatic Nerve
4.6. Quantitative Real-Time PCR to Assess Nfl Gene Expression in Mouse Sciatic Nerve Tissue and in NSC-34 Transfected Cells
4.7. Western Blotting Assay
4.8. Quantitative Real-Time PCR to Assess miR-146a Expression in Mouse Serum
4.9. Patients and Biological Samples
4.10. Quantitative Real-Time PCR to Assess miR-146a Expression in Human Serum
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hardiman, O.; Al-Chalabi, A.; Chio, A.; Corr, E.M.; Logroscino, G.; Robberecht, W.; Shaw, P.J.; Simmons, Z.; Van Den Berg, L.H. Amyotrophic lateral sclerosis. Nat. Rev. Dis. Prim. 2017, 3, 17071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiò, A.; Logroscino, G.; Hardiman, O.; Swingler, R.; Mitchell, D.; Beghi, E.; Traynor, B.G.; Eurals Consortium. Prognostic factors in ALS: A critical review. Amyotroph. Lateral Scler. 2009, 10, 310–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, S.; Khurana, S.; Vats, A.; Sahu, B.; Ganguly, N.K.; Chakraborti, P.; Gourie-Devi, M.; Taneja, V. Neuromuscular junction dysfunction in amyotrophic lateral sclerosis. Mol. Neurobiol. 2022, 59, 1502–1527. [Google Scholar] [CrossRef] [PubMed]
- Riva, N.; Chaabane, L.; Peviani, M.; Ungaro, D.; Domi, T.; Dina, G.; Bianchi, F.; Spano, G.; Cerri, F.; Podini, P.; et al. Defining peripheral nervous system dysfunction in the SOD-1G93A transgenic rat model of amyotrophic lateral sclerosis. J. Neuropathol. Exp. Neurol. 2014, 73, 658–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcuzzo, S.; Bonanno, S.; Figini, M.; Scotti, A.; Zucca, I.; Minati, L.; Riva, N.; Domi, T.; Fossaghi, A.; Quattrini, A.; et al. A longitudinal DTI and histological study of the spinal cord reveals early pathological alterations in G93A-SOD1 mouse model of amyotrophic lateral sclerosis. Exp. Neurol. 2017, 293, 43–52. [Google Scholar] [CrossRef]
- Nardo, G.; Trolese, M.C.; Verderio, M.; Mariani, A.; de Paola, M.; Riva, N.; Dina, G.; Panini, N.; Erba, E.; Quattrini, A.; et al. Counteracting roles of MHCI and CD8+ T cells in the peripheral and central nervous system of ALS SOD1G93A mice. Mol. Neurodegener. 2018, 13, 42. [Google Scholar] [CrossRef]
- Yu, H.; Chen, L.; Zhang, S.; He, J.; Fan, D. Early axonal dysfunction of the peripheral nervous system influences disease progression of ALS: Evidence from clinical neuroelectrophysiology. Front. Neurol. 2021, 12, 574919. [Google Scholar] [CrossRef]
- Boylan, K. Familial amyotrophic lateral sclerosis. Neurol. Clin. 2015, 33, 807–830. [Google Scholar] [CrossRef] [Green Version]
- Brown, R.H.; Al-Chalabi, A. Amyotrophic lateral sclerosis. N. Engl. J. Med. 2017, 377, 162–172. [Google Scholar] [CrossRef] [Green Version]
- Ghasemi, M.; Brown, R.H., Jr. Genetics of amyotrophic lateral sclerosis. Cold Spring Harb Perspect. Med. 2018, 8, a024125. [Google Scholar] [CrossRef]
- Zaepfel, B.L.; Rothstein, J.D. RNA Is a double-Edged sword in ALS pathogenesis. Front. Cell. Neurosci. 2021, 15, 708181. [Google Scholar] [CrossRef]
- Bakkar, N.; Boehringer, A.; Bowser, R. Use of biomarkers in ALS drug development and clinical trials. Brain Res. 2015, 1607, 94–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiernan, M.C.; Vucic, S.; Talbot, K.; McDermott, C.J.; Hardiman, O.; Shefner, J.M.; Al-Chalabi, A.; Huynh, W.; Cudkowicz, M.; Talman, P.; et al. Improving clinical trial outcomes in amyotrophic lateral sclerosis. Nat. Rev. Neurol. 2021, 17, 104–118. [Google Scholar] [CrossRef] [PubMed]
- Gille, B.; De Schaepdryver, M.; Goossens, J.; Dedeene, L.; De Vocht, J.; Oldoni, E.; Goris, A.; Van Den Bosch, L.; Depreitere, B.; Claeys, K.G.; et al. Serum neurofilament light chain levels as a marker of upper motor neuron degeneration in patients with Amyotrophic Lateral Sclerosis. Neuropathol. Appl. Neurobiol. 2019, 45, 291–304. [Google Scholar] [CrossRef]
- Zhang, L.; Ji, T.; Wu, C.; Zhang, S.; Tang, L.; Zhang, N.; Liu, X.; Fan, D. Serum neurofilament light chain levels may be a marker of lower motor neuron damage in amyotrophic lateral sclerosis. Front. Neurol. 2022, 13, 833507. [Google Scholar] [CrossRef] [PubMed]
- Verde, F.; Otto, M.; Silani, V. Neurofilament light chain as biomarker for amyotrophic lateral sclerosis and frontotemporal dementia. Front. Neurosci. 2021, 15, 679199. [Google Scholar] [CrossRef] [PubMed]
- Behzadi, A.; Pujol-Calderón, F.; Tjust, A.E.; Wuolikainen, A.; Höglund, K.; Forsberg, K.; Portelius, E.; Blennow, K.; Zetterberg, H.; Andersen, P.M. Neurofilaments can differentiate ALS subgroups and ALS from common diagnostic mimics. Sci. Rep. 2021, 11, 22128. [Google Scholar] [CrossRef] [PubMed]
- Butti, Z.; Patten, S.A. RNA dysregulation in amyotrophic lateral sclerosis. Front. Genet. 2019, 9, 712. [Google Scholar] [CrossRef] [Green Version]
- Laneve, P.; Tollis, P.; Caffarelli, E. RNA deregulation in amyotrophic lateral sclerosis: The Noncoding Perspective. Int. J. Mol. Sci. 2021, 22, 10285. [Google Scholar] [CrossRef]
- Saliminejad, K.; Khorram Khorshid, H.R.; Soleymani Fard, S.; Ghaffari, S.H. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J. Cell. Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef]
- Im, H.I.; Kenny, P.J. MicroRNAs in neuronal function and dysfunction. Trends Neurosci. 2012, 35, 325–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajman, M.; Schratt, G. MicroRNAs in neural development: From master regulators to fine-tuners. Development 2017, 144, 2310–2322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcuzzo, S.; Kapetis, D.; Mantegazza, R.; Baggi, F.; Bonanno, S.; Barzago, C.; Cavalcante, P.; Kerlero de Rosbo, N.; Bernasconi, P. Altered miRNA expression is associated with neuronal fate in G93A-SOD1 ependymal stem progenitor cells. Exp. Neurol. 2014, 253, 91–101. [Google Scholar] [CrossRef]
- Marcuzzo, S.; Bonanno, S.; Kapetis, D.; Barzago, C.; Cavalcante, P.; D'Alessandro, S.; Mantegazza, R.; Bernasconi, P. Up-regulation of neural and cell cycle-related microRNAs in brain of amyotrophic lateral sclerosis mice at late disease stage. Mol. Brain 2015, 8, 5. [Google Scholar] [CrossRef] [Green Version]
- Bonanno, S.; Marcuzzo, S.; Malacarne, C.; Giagnorio, E.; Masson, R.; Zanin, R.; Arnoldi, M.T.; Andreetta, F.; Simoncini, O.; Venerando, A.; et al. Circulating MyomiRs as potential biomarkers to monitor response to nusinersen in pediatric SMA patients. Biomedicines 2020, 8, 21. [Google Scholar] [CrossRef] [Green Version]
- Giagnorio, E.; Malacarne, C.; Mantegazza, R.; Bonanno, S.; Marcuzzo, S. MyomiRs and their multifaceted regulatory roles in muscle homeostasis and amyotrophic lateral sclerosis. J. Cell Sci. 2021, 134, jcs258349. [Google Scholar] [CrossRef]
- Malacarne, C.; Galbiati, M.; Giagnorio, E.; Cavalcante, P.; Salerno, F.; Andreetta, F.; Cagnoli, C.; Taiana, M.; Nizzardo, M.; Corti, S.; et al. Dysregulation of muscle-specific microRNAs as common pathogenic feature associated with muscle atrophy in ALS, SMA and SBMA: Evidence from animal models and human patients. Int. J. Mol. Sci. 2021, 22, 5673. [Google Scholar] [CrossRef]
- Campos-Melo, D.; Droppelmann, C.A.; He, Z.; Volkening, K.; Strong, M.J. Altered microRNA expression profile in amyotrophic lateral sclerosis: A role in the regulation of NFL mRNA levels. Mol. Brain 2013, 6, 26. [Google Scholar] [CrossRef] [Green Version]
- Pegoraro, V.; Merico, A.; Angelini, C. Micro-RNAs in ALS muscle: Differences in gender, age at onset and disease duration. J. Neurol. Sci. 2017, 380, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Taganov, K.D.; Boldin, M.P.; Chang, K.J.; Baltimore, D. NF-kappaB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc. Natl. Acad. Sci. USA 2006, 103, 12481–12486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, C.; Sequeira, C.; Likhite, S.; Dennys, C.N.; Kolb, S.J.; Shaw, P.J.; Vaz, A.R.; Kaspar, B.K.; Meyer, K.; Brites, D. Neurotoxic astrocytes directly converted from sporadic and familial ALS patient fibroblasts reveal signature diversities and miR-146a theragnostic potential in specific subtypes. Cells 2022, 11, 1186. [Google Scholar] [CrossRef] [PubMed]
- Barry, D.M.; Millecamps, S.; Julien, J.P.; Garcia, M.L. New movements in neurofilament transport, turnover and disease. Exp. Cell Res. 2007, 313, 2110–2120. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Qin, X.; Chang, X.; Wang, H.; Guo, J.; Zhang, W. Neurofilament markers in serum and cerebrospinal fluid of patients with amyotrophic lateral sclerosis. J. Cell. Mol. Med. 2022, 26, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Akiyama, T.; Warita, H.; Aoki, M. Omics approach to axonal dysfunction of motor neurons in amyotrophic lateral sclerosis (ALS). Front Neurosci. 2020, 14, 194. [Google Scholar] [CrossRef] [Green Version]
- Jia, L.; Wang, L.; Chopp, M.; Zhang, Y.; Szalad, A.; Zhang, Z.G. MicroRNA 146a locally mediates distal axonal growth of dorsal root ganglia neurons under high glucose and sildenafil conditions. Neuroscience 2016, 329, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Fan, W.; Liang, C.; Ou, M.; Zou, T.; Sun, F.; Zhou, H.; Cui, L. MicroRNA-146a Is a Wide-Reaching Neuroinflammatory regulator and potential treatment target in neurological diseases. Front. Mol. Neurosci. 2020, 13, 90. [Google Scholar] [CrossRef]
- Yichen, L.; Lee, B.H.; Yip, W.; Chou, P.; Yip, B.S. Neurofilament Proteins as Prognostic Biomarkers in Neurological Disorders. Curr. Pharm. Des. 2019, 25, 4560–4569 . [Google Scholar] [CrossRef]
- Huang, C.-J.; Nguyen, P.N.N.; Choo, K.B.; Sugii, S.; Wee, K.; Cheong, S.K.; Kamarul, T. Frequent Co-Expression of miRNA-5p and -3p Species and Cross-Targeting in Induced Pluripotent Stem Cells. Int. J. Med Sci. 2014, 11, 824–833. [Google Scholar] [CrossRef] [Green Version]
- Yurikova, O.Y.; Aisina, D.E.; Niyazova, R.E.; Atambayeva, S.A.; Labeit, S.; Ivashchenko, A.T. The Interaction of miRNA-5p and miRNA-3p with the mRNAs of Orthologous Genes. Bioinformatics 2019, 53, 692–704. [Google Scholar] [CrossRef]
- Yuan, A.; Nixon, R.A. Neurofilament proteins as biomarkers to monitor neurological diseases and the efficacy of therapies. Front. Neurosci. 2021, 15, 689938. [Google Scholar] [CrossRef]
- Dobrowolny, G.; Martone, J.; Lepore, E.; Casola, I.; Petrucci, A.; Inghilleri, M.; Morlando, M.; Colantoni, A.; Scicchitano, B.M.; Calvo, A.; et al. A longitudinal study defined circulating microRNAs as reliable biomarkers for disease prognosis and progression in ALS human patients. Cell Death Discov. 2021, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Si, Y.; Cui, X.; Crossman, D.K.; Hao, J.; Kazamel, M.; Kwon, Y.; King, P.H. Muscle microRNA signatures as biomarkers of disease progression in amyotrophic lateral sclerosis. Neurobiol. Dis. 2018, 114, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. miRNAs as biomarkers in disease: Latest findings regarding their role in diagnosis and prognosis. Cells 2020, 9, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricci, C.; Marzocchi, C.; Battistini, S. MicroRNAs as biomarkers in amyotrophic lateral sclerosis. Cells 2018, 7, 219. [Google Scholar] [CrossRef] [Green Version]
- Marcuzzo, S.; Zucca, I.; Mastropietro, A.; de Rosbo, N.K.; Cavalcante, P.; Tartari, S.; Bonanno, S.; Preite, L.; Mantegazza, R.; Bernasconi, P. Hind limb muscle atrophy precedes cerebral neuronal degeneration in G93A-SOD1 mouse model of amyotrophic lateral sclerosis: A longitudinal MRI study. Exp. Neurol. 2011, 231, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Nango, H.; Kosuge, Y.; Sato, M.; Shibukawa, Y.; Aono, Y.; Saigusa, T.; Ito, Y.; Ishige, K. Highly Efficient Conversion of Motor Neuron-Like NSC-34 Cells into Functional Motor Neurons by Prostaglandin E2. Cells 2020, 9, 1741. [Google Scholar] [CrossRef] [PubMed]
- Genevini, P.; Colombo, M.N.; Venditti, R.; Marcuzzo, S.; Colombo, S.F.; Bernasconi, P.; De Matteis, M.A.; Borgese, N.; Navone, F. VAPB depletion alters neuritogenesis and phosphoinositide balance in motoneuron-like cells: Relevance to VAPB-linked amyotrophic lateral sclerosis. J. Cell Sci. 2019, 132, jcs220061. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ALS Patients (n = 24) | Healthy Controls (n = 23) | |
|---|---|---|
| Sex | ||
| M% | 11(45.83%) | 11 (47.82%) |
| Age (years, mean ± SEM) | 61 ± 8.19 | 60.17 ± 7.13 |
| Clinical predominance | ||
| UMN | 9 (37.5%) | |
| LMN | 8 (33.33%) | |
| UMN + LMN | 7 (29.16%) | |
| Disease duration—months (mean ± SEM) | ||
| Whole population | 17.93 ± 12.96 | |
| UMN | 19.56 ± 12.29 | |
| LMN | 21.13 ± 18.24 | |
| UMN + LMN | 12.43 ± 9.66 | |
| ALSFRS-R | ||
| Whole population | 40.71 ± 5.18 | |
| UMN | 40.78 ± 6.26 | |
| LMN | 42.25 ± 3.11 | |
| UMN + LMN | 39.57 ± 3.31 | |
| Genotype | ||
| C9orf72 | 1 (4.16%) | |
| SOD1 | 1 (4.16%) | |
| TDP43 | 1 (4.16%) | |
| VUS SOD | 2 (8.33%) | |
| VUS UBQLN2 | 1 (4.16%) | |
| No genetic cause is found | 18 (75%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giagnorio, E.; Malacarne, C.; Cavalcante, P.; Scandiffio, L.; Cattaneo, M.; Pensato, V.; Gellera, C.; Riva, N.; Quattrini, A.; Dalla Bella, E.; et al. MiR-146a in ALS: Contribution to Early Peripheral Nerve Degeneration and Relevance as Disease Biomarker. Int. J. Mol. Sci. 2023, 24, 4610. https://doi.org/10.3390/ijms24054610
Giagnorio E, Malacarne C, Cavalcante P, Scandiffio L, Cattaneo M, Pensato V, Gellera C, Riva N, Quattrini A, Dalla Bella E, et al. MiR-146a in ALS: Contribution to Early Peripheral Nerve Degeneration and Relevance as Disease Biomarker. International Journal of Molecular Sciences. 2023; 24(5):4610. https://doi.org/10.3390/ijms24054610
Chicago/Turabian StyleGiagnorio, Eleonora, Claudia Malacarne, Paola Cavalcante, Letizia Scandiffio, Marco Cattaneo, Viviana Pensato, Cinzia Gellera, Nilo Riva, Angelo Quattrini, Eleonora Dalla Bella, and et al. 2023. "MiR-146a in ALS: Contribution to Early Peripheral Nerve Degeneration and Relevance as Disease Biomarker" International Journal of Molecular Sciences 24, no. 5: 4610. https://doi.org/10.3390/ijms24054610