
Key Stratification of Microbiota Taxa and Metabolites in the Host Metabolic Health–Disease Balance
 , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Differential Microbiota Taxa Composition and Stratification According to Their Representation in Metabolic Diseases
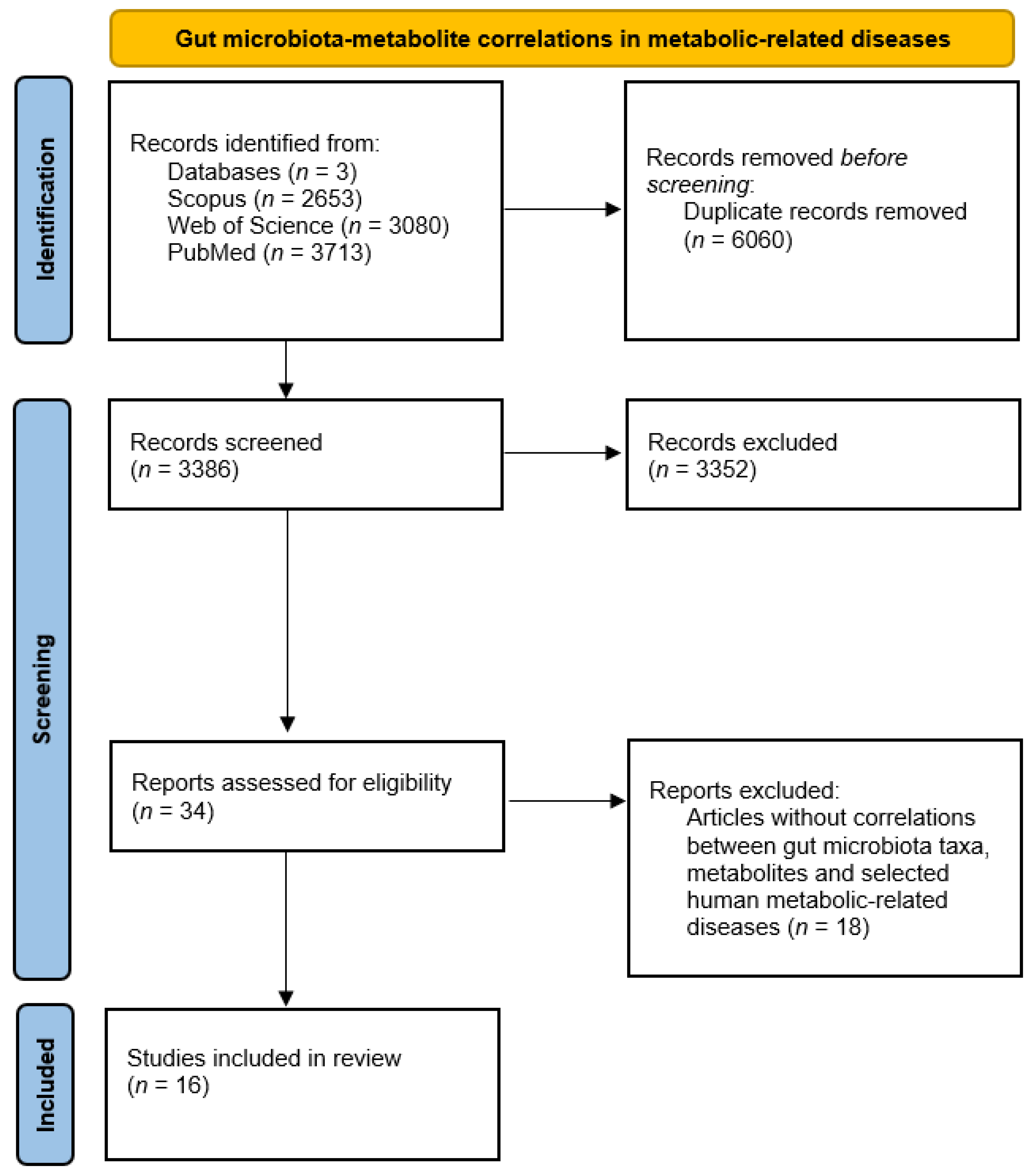
2.1.1. PRISMA Analysis
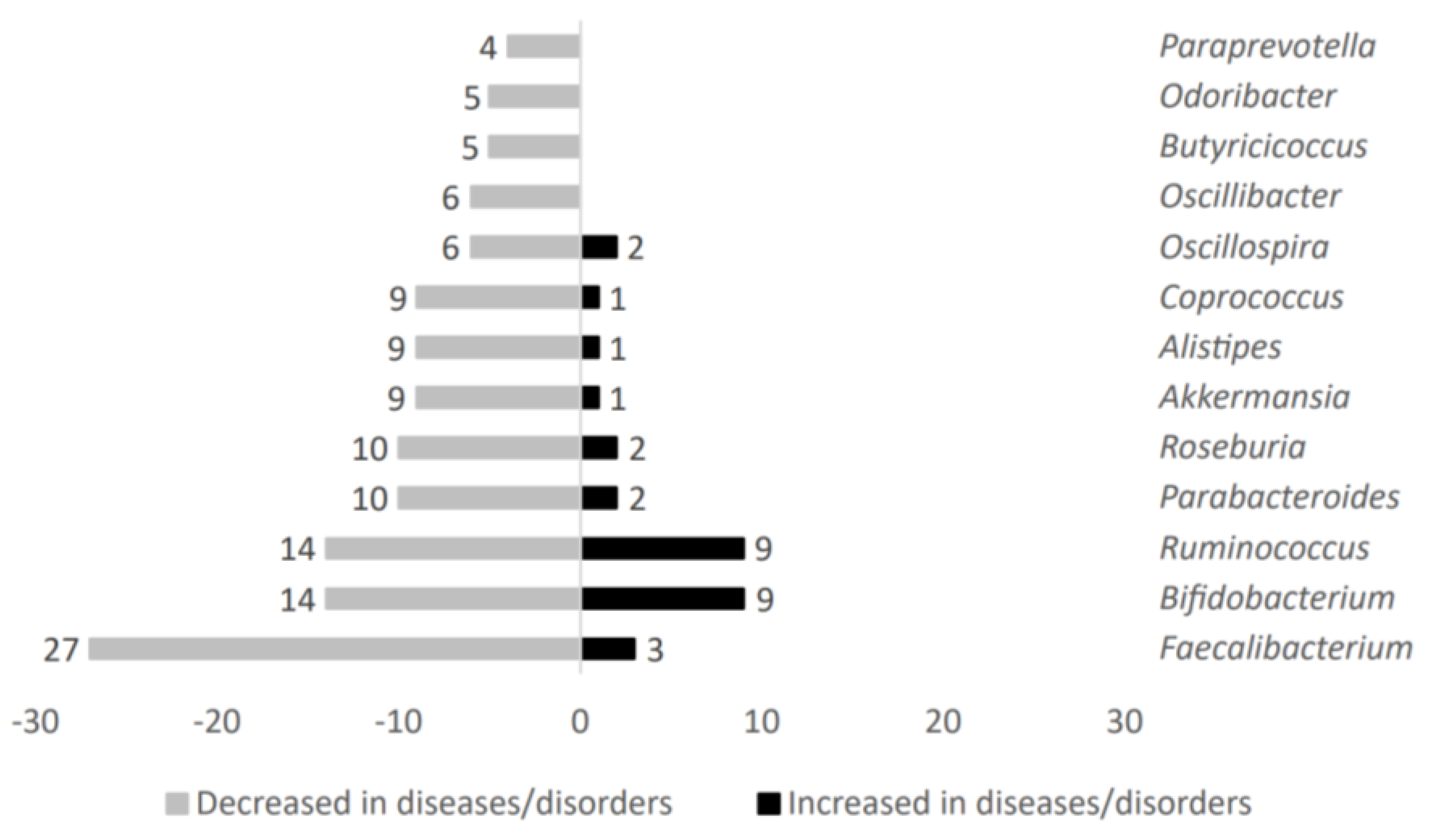
2.1.2. Microbial Taxa Decreased in Patients Suffering from Metabolic-Related Diseases
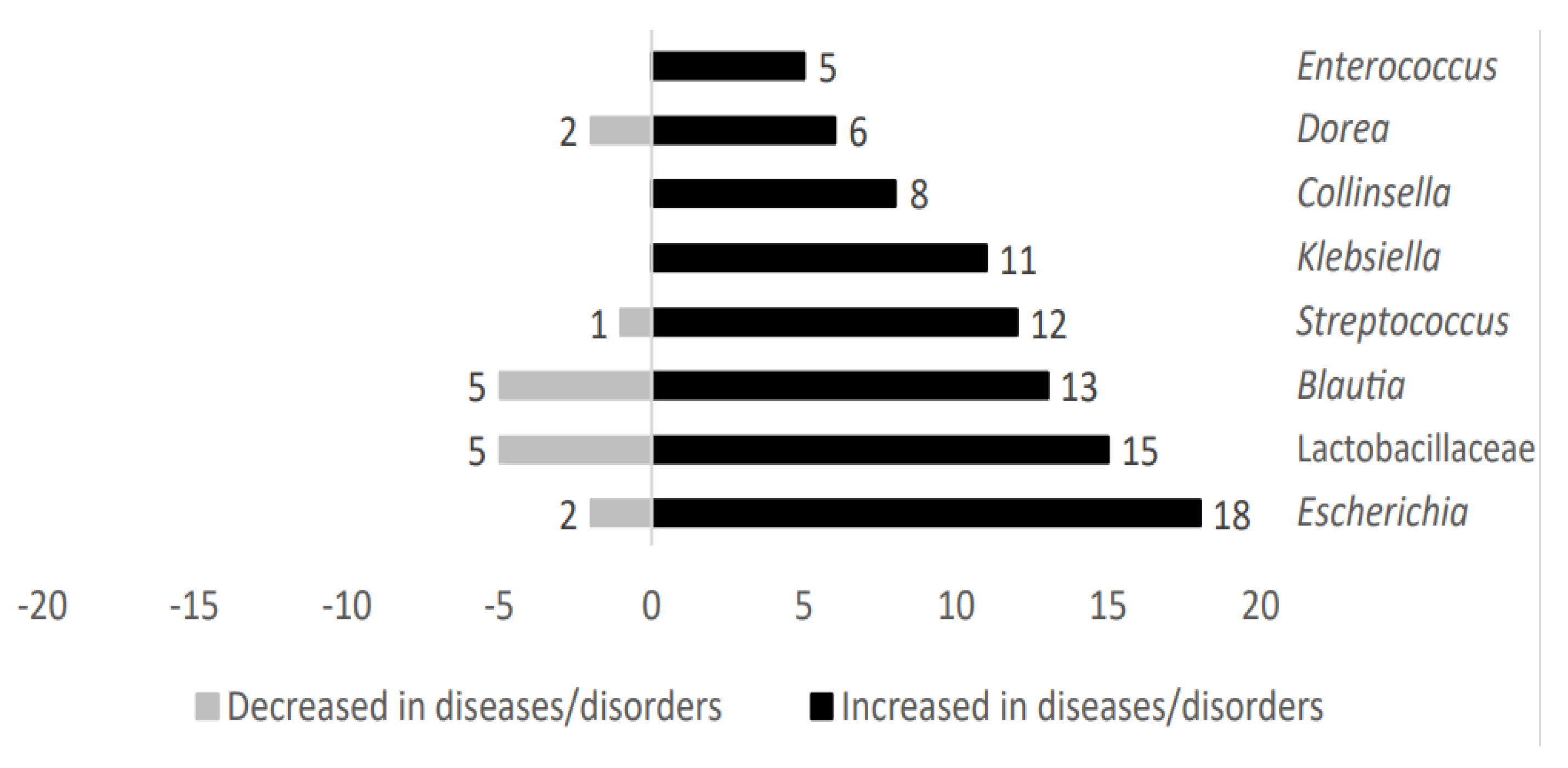
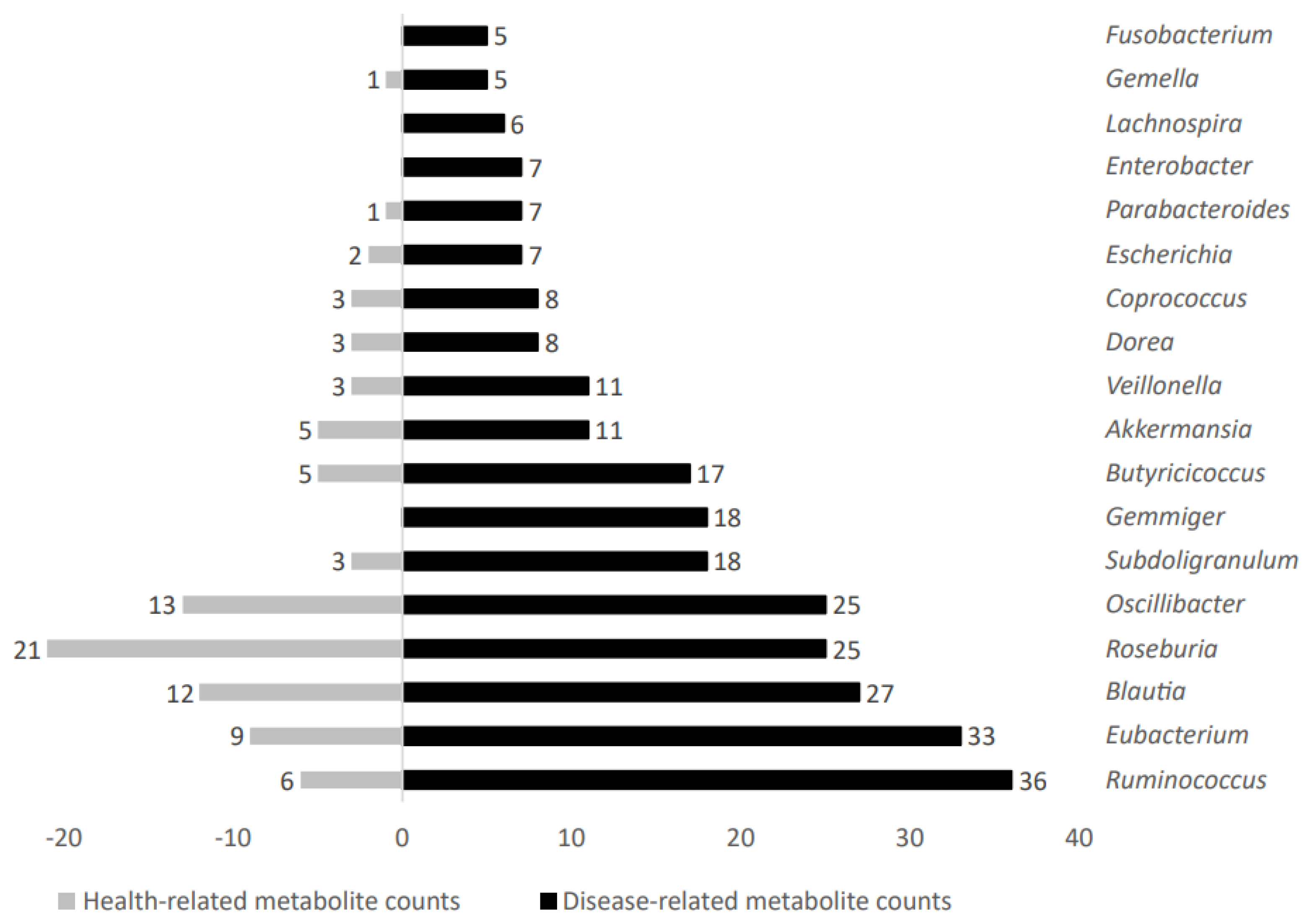
2.1.3. Microbial Taxa Increased in Patients Suffering Metabolic-Related Diseases
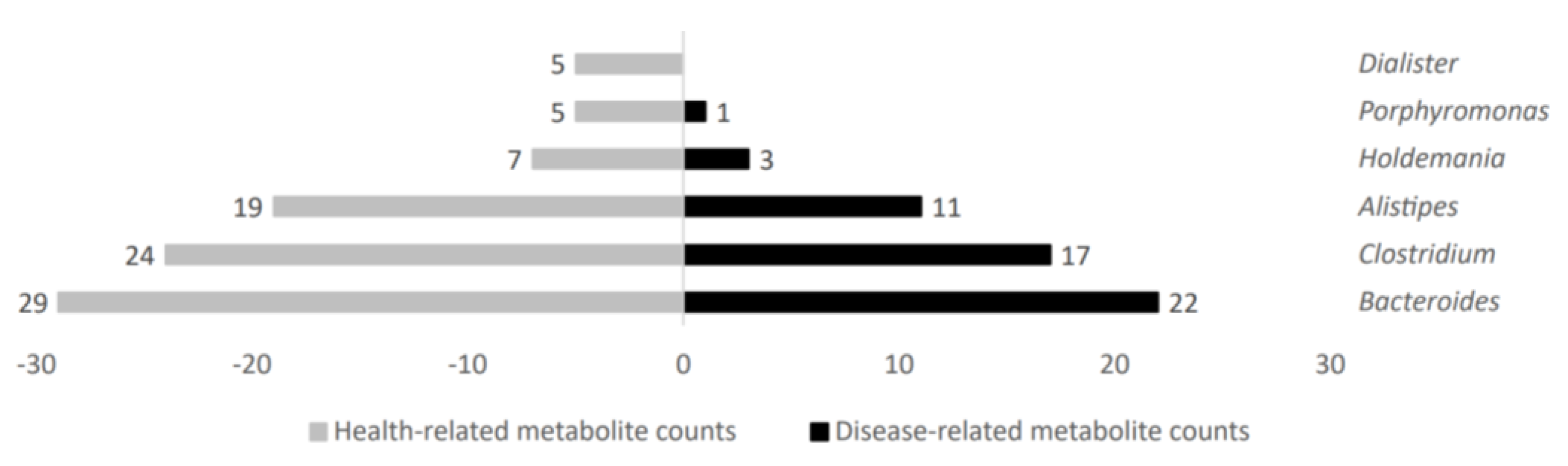
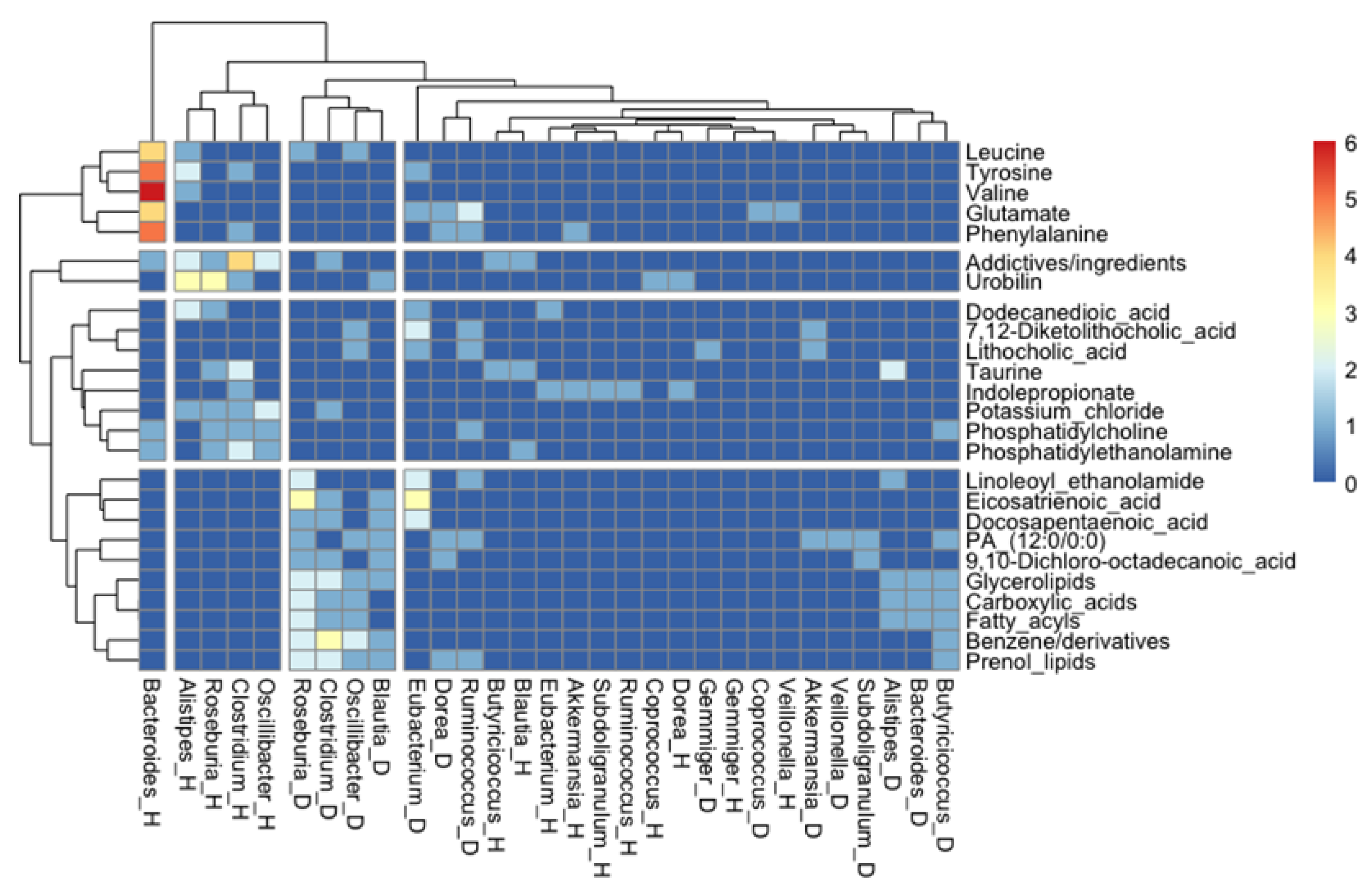
2.2. Differential Microbial Metabolites and Stratification According to Their Representation in Metabolic Diseases
3. Materials and Methods
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duan, M.; Wang, Y.; Zhang, Q.; Zou, R.; Guo, M.; Zheng, H. Characteristics of Gut Microbiota in People with Obesity. PLoS ONE 2021, 16, e0255446. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, C.C.; Monteil, M.A.; Davis, E.M. Overweight and Obesity in Children Are Associated with an Abundance of Firmicutes and Reduction of Bifidobacterium in Their Gastrointestinal Microbiota. Child Obes 2020, 16, 204–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Sun, H.; Jiang, F.; Shen, Y.; Li, X.; Hu, X.; Shen, X.; Wei, P. Alteration of the Gut Microbiota Associated with Childhood Obesity by 16S RRNA Gene Sequencing. PeerJ 2020, 8, e8317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, R.; Zhu, C.; Li, H.; Yin, M.; Pan, C.; Huang, L.; Kong, C.; Wang, X.; Zhang, Y.; Qu, S.; et al. Dysbiosis Signatures of Gut Microbiota Along the Sequence from Healthy, Young Patients to Those with Overweight and Obesity. Obesity 2018, 26, 351–361. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.-P.; He, Q.-Q.; Ouyang, H.-M.; Peng, H.-S.; Wang, Q.; Li, J.; Lv, X.-F.; Zheng, Y.-N.; Li, S.-C.; Liu, H.-L.; et al. Human Gut Microbiota Associated with Obesity in Chinese Children and Adolescents. BioMed Res. Int. 2017, 2017, 7585989. [Google Scholar] [CrossRef] [PubMed]
- Riva, A.; Borgo, F.; Lassandro, C.; Verduci, E.; Morace, G.; Borghi, E.; Berry, D. Pediatric Obesity Is Associated with an Altered Gut Microbiota and Discordant Shifts in Firmicutes Populations. Environ. Microbiol. 2017, 19, 95–105. [Google Scholar] [CrossRef]
- Kashtanova, D.A.; Tkacheva, O.N.; Doudinskaya, E.N.; Strazhesko, I.D.; Kotovskaya, Y.V.; Popenko, A.S.; Tyakht, A.V.; Alexeev, D.G. Gut Microbiota in Patients with Different Metabolic Statuses: Moscow Study. Microorganisms 2018, 6, E98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Zhou, J.; Liu, J.; Wang, Z.; Chen, M.; Zhou, S. Metagenome of Gut Microbiota of Children with Nonalcoholic Fatty Liver Disease. Front. Pediatr. 2019, 7, 518. [Google Scholar] [CrossRef]
- Nistal, E.; Sáenz de Miera, L.E.; Ballesteros Pomar, M.; Sánchez-Campos, S.; García-Mediavilla, M.V.; Álvarez-Cuenllas, B.; Linares, P.; Olcoz, J.L.; Arias-Loste, M.T.; García-Lobo, J.M.; et al. An Altered Fecal Microbiota Profile in Patients with Non-Alcoholic Fatty Liver Disease (NAFLD) Associated with Obesity. Rev. Esp. Enferm. Dig. 2019, 111, 275–282. [Google Scholar] [CrossRef]
- Del Chierico, F.; Nobili, V.; Vernocchi, P.; Russo, A.; De Stefanis, C.; Gnani, D.; Furlanello, C.; Zandonà, A.; Paci, P.; Capuani, G.; et al. Gut Microbiota Profiling of Pediatric Nonalcoholic Fatty Liver Disease and Obese Patients Unveiled by an Integrated Meta-Omics-Based Approach. Hepatology 2017, 65, 451–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, S.M.B.; Stefano, J.T.; Miele, L.; Ponziani, F.R.; Souza-Basqueira, M.; Okada, L.S.R.R.; de Barros Costa, F.G.; Toda, K.; Mazo, D.F.C.; Sabino, E.C.; et al. Gut Microbiome Composition in Lean Patients with NASH Is Associated with Liver Damage Independent of Caloric Intake: A Prospective Pilot Study. Nutr. Metab. Cardiovasc. Dis. 2018, 28, 369–384. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Martín, A.; Iglesias-Aguirre, C.E.; Meoro, A.; Selma, M.V.; Espín, J.C. There Is No Distinctive Gut Microbiota Signature in the Metabolic Syndrome: Contribution of Cardiovascular Disease Risk Factors and Associated Medication. Microorganisms 2020, 8, 416. [Google Scholar] [CrossRef] [Green Version]
- Lippert, K.; Kedenko, L.; Antonielli, L.; Kedenko, I.; Gemeier, C.; Leitner, M.; Kautzky-Willer, A.; Paulweber, B.; Hackl, E. Gut Microbiota Dysbiosis Associated with Glucose Metabolism Disorders and the Metabolic Syndrome in Older Adults. Benef. Microbes 2017, 8, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Thingholm, L.B.; Rühlemann, M.C.; Koch, M.; Fuqua, B.; Laucke, G.; Boehm, R.; Bang, C.; Franzosa, E.A.; Hübenthal, M.; Rahnavard, A.; et al. Obese Individuals with and without Type 2 Diabetes Show Different Gut Microbial Functional Capacity and Composition. Cell Host Microbe 2019, 26, 252–264.e10. [Google Scholar] [CrossRef]
- Zhou, Z.; Zheng, Z.; Xiong, X.; Chen, X.; Peng, J.; Yao, H.; Pu, J.; Chen, Q.; Zheng, M. Gut Microbiota Composition and Fecal Metabolic Profiling in Patients with Diabetic Retinopathy. Front. Cell Dev. Biol. 2021, 9, 2684. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, S.; Yang, Y.; Wang, Z.; Wang, B.; Zhang, B.; Yu, J.; Lu, W.; Pan, M.; Zhao, J.; et al. The Diversity of Gut Microbiota in Type 2 Diabetes with or without Cognitive Impairment. Aging Clin. Exp. Res. 2021, 33, 589–601. [Google Scholar] [CrossRef]
- Li, Q.; Chang, Y.; Zhang, K.; Chen, H.; Tao, S.; Zhang, Z. Implication of the Gut Microbiome Composition of Type 2 Diabetic Patients from Northern China. Sci. Rep. 2020, 10, 5450. [Google Scholar] [CrossRef] [Green Version]
- Navab-Moghadam, F.; Sedighi, M.; Khamseh, M.E.; Alaei-Shahmiri, F.; Talebi, M.; Razavi, S.; Amirmozafari, N. The Association of Type II Diabetes with Gut Microbiota Composition. Microb. Pathog. 2017, 110, 630–636. [Google Scholar] [CrossRef]
- Sedighi, M.; Razavi, S.; Navab-Moghadam, F.; Khamseh, M.E.; Alaei-Shahmiri, F.; Mehrtash, A.; Amirmozafari, N. Comparison of Gut Microbiota in Adult Patients with Type 2 Diabetes and Healthy Individuals. Microb. Pathog. 2017, 111, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Ejtahed, H.; Hoseini-Tavassol, Z.; Khatami, S.; Zangeneh, M.; Behrouzi, A.; Badi, S.A.; Moshiri, A.; Hasani-Ranjbar, S.; Soroush, A.; Vaziri, F.; et al. Main Gut Bacterial Composition Differs between Patients with Type 1 and Type 2 Diabetes and Non-Diabetic Adults. J. Diabetes Metab. Disord. 2020, 19, 265–271. [Google Scholar] [CrossRef]
- Leiva-Gea, I.; Sánchez-Alcoholado, L.; Martín-Tejedor, B.; Castellano-Castillo, D.; Moreno-Indias, I.; Urda-Cardona, A.; Tinahones, F.J.; Fernández-García, J.C.; Queipo-Ortuño, M.I. Gut Microbiota Differs in Composition and Functionality between Children with Type 1 Diabetes and MODY2 and Healthy Control Subjects: A Case-Control Study. Diabetes Care 2018, 41, 2385–2395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, N.; Wittouck, S.; Mattarelli, P.; Zheng, J.; Lebeer, S.; Felis, G.E.; Gänzle, M.G. After the Storm—Perspectives on the Taxonomy of Lactobacillaceae. JDS Commun. 2022, 3, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Sroka-Oleksiak, A.; Młodzińska, A.; Bulanda, M.; Salamon, D.; Major, P.; Stanek, M.; Gosiewski, T. Metagenomic Analysis of Duodenal Microbiota Reveals a Potential Biomarker of Dysbiosis in the Course of Obesity and Type 2 Diabetes: A Pilot Study. J. Clin. Med. 2020, 9, 369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaike, A.H.; Paul, D.; Bhute, S.; Dhotre, D.P.; Pande, P.; Upadhyaya, S.; Reddy, Y.; Sampath, R.; Ghosh, D.; Chandraprabha, D.; et al. The Gut Microbial Diversity of Newly Diagnosed Diabetics but Not of Prediabetics Is Significantly Different from That of Healthy Nondiabetics. mSystems 2020, 5, e00578-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Lou, H.; Peng, Y.; Chen, S.; Zhang, Y.; Li, X. Comprehensive Relationships between Gut Microbiome and Faecal Metabolome in Individuals with Type 2 Diabetes and Its Complications. Endocrine 2019, 66, 526–537. [Google Scholar] [CrossRef]
- Chen, P.-C.; Chien, Y.-W.; Yang, S.-C. The Alteration of Gut Microbiota in Newly Diagnosed Type 2 Diabetic Patients. Nutrition 2019, 63–64, 51–56. [Google Scholar] [CrossRef]
- Liu, X.; Cheng, Y.-W.; Shao, L.; Sun, S.-H.; Wu, J.; Song, Q.-H.; Zou, H.-S.; Ling, Z.-X. Gut Microbiota Dysbiosis in Chinese Children with Type 1 Diabetes Mellitus: An Observational Study. World J. Gastroenterol. 2021, 27, 2394–2414. [Google Scholar] [CrossRef]
- Ahmed, E.A.; Ahmed, S.M.; Zakaria, N.H.; Baddour, N.M.; Header, D.A. Estudio del microbioma intestinal en pacientes egipcios con colitis ulcerosa crónica idiopática. Rev. Gastroenterol. México 2022, 843, 1–10. [Google Scholar] [CrossRef]
- Zakerska-Banaszak, O.; Tomczak, H.; Gabryel, M.; Baturo, A.; Wolko, L.; Michalak, M.; Malinska, N.; Mankowska-Wierzbicka, D.; Eder, P.; Dobrowolska, A.; et al. Dysbiosis of Gut Microbiota in Polish Patients with Ulcerative Colitis: A Pilot Study. Sci. Rep. 2021, 11, 2166. [Google Scholar] [CrossRef]
- Dai, L.; Tang, Y.; Zhou, W.; Dang, Y.; Sun, Q.; Tang, Z.; Zhu, M.; Ji, G. Gut Microbiota and Related Metabolites Were Disturbed in Ulcerative Colitis and Partly Restored after Mesalamine Treatment. Front. Pharmacol. 2021, 11, 620724. [Google Scholar] [CrossRef]
- Knoll, R.L.; Forslund, K.; Kultima, J.R.; Meyer, C.U.; Kullmer, U.; Sunagawa, S.; Bork, P.; Gehring, S. Gut Microbiota Differs between Children with Inflammatory Bowel Disease and Healthy Siblings in Taxonomic and Functional Composition: A Metagenomic Analysis. Am. J. Physiol.-Gastrointest. Liver Physiol. 2017, 312, G327–G339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heidarian, F.; Alebouyeh, M.; Shahrokh, S.; Balaii, H.; Zali, M.R. Altered Fecal Bacterial Composition Correlates with Disease Activity in Inflammatory Bowel Disease and the Extent of IL8 Induction. Curr. Res. Transl. Med. 2019, 67, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.-Q.; Yu, T.-T.; Zhao, X.-J.; Zhang, Y.; Zhang, H.-J. Fecal Microbial Dysbiosis in Chinese Patients with Inflammatory Bowel Disease. World J. Gastroenterol. 2018, 24, 1464–1477. [Google Scholar] [CrossRef] [Green Version]
- Franzosa, E.A.; Sirota-Madi, A.; Avila-Pacheco, J.; Fornelos, N.; Haiser, H.J.; Reinker, S.; Vatanen, T.; Hall, A.B.; Mallick, H.; McIver, L.J.; et al. Gut Microbiome Structure and Metabolic Activity in Inflammatory Bowel Disease. Nat. Microbiol. 2019, 4, 293–305. [Google Scholar] [CrossRef]
- Chang, T.-E.; Luo, J.-C.; Yang, U.-C.; Huang, Y.-H.; Hou, M.-C.; Lee, F.-Y. Fecal Microbiota Profile in Patients with Inflammatory Bowel Disease in Taiwan. J. Chin. Med. Assoc. 2021, 84, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Lopetuso, L.R.; Petito, V.; Graziani, C.; Schiavoni, E.; Paroni Sterbini, F.; Poscia, A.; Gaetani, E.; Franceschi, F.; Cammarota, G.; Sanguinetti, M.; et al. Gut Microbiota in Health, Diverticular Disease, Irritable Bowel Syndrome, and Inflammatory Bowel Diseases: Time for Microbial Marker of Gastrointestinal Disorders. Dig. Dis. 2018, 36, 56–65. [Google Scholar] [CrossRef]
- Budinska, E.; Gojda, J.; Heczkova, M.; Bratova, M.; Dankova, H.; Wohl, P.; Bastova, H.; Lanska, V.; Kostovcik, M.; Dastych, M.; et al. Microbiome and Metabolome Profiles Associated with Different Types of Short Bowel Syndrome: Implications for Treatment. J. Parenter. Enter. Nutr. 2020, 44, 105–118. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, T.; Nishida, A.; Yamano, M.; Kimura, I. Short-Chain Fatty Acid Receptors and Gut Microbiota as Therapeutic Targets in Metabolic, Immune, and Neurological Diseases. Pharmacol. Ther. 2022, 239, 108273. [Google Scholar] [CrossRef]
- Su, X.; Gao, Y.; Yang, R. Gut Microbiota-Derived Tryptophan Metabolites Maintain Gut and Systemic Homeostasis. Cells 2022, 11, 2296. [Google Scholar] [CrossRef]
- Qian, B.; Zhang, K.; Li, Y.; Sun, K. Update on Gut Microbiota in Cardiovascular Diseases. Front. Cell. Infect. Microbiol. 2022, 12, 1694. [Google Scholar] [CrossRef]
- Cai, J.; Rimal, B.; Jiang, C.; Chiang, J.Y.L.; Patterson, A.D. Bile Acid Metabolism and Signaling, the Microbiota, and Metabolic Disease. Pharmacol. Ther. 2022, 237, 108238. [Google Scholar] [CrossRef]
- Ohtani, N.; Kawada, N. Role of the Gut–Liver Axis in Liver Inflammation, Fibrosis, and Cancer: A Special Focus on the Gut Microbiota Relationship. Hepatol. Commun. 2019, 3, 456–470. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.-L.; Li, Z.-J.; Gou, H.-Z.; Song, X.-J.; Zhang, L. The Gut Microbiota–Bile Acid Axis: A Potential Therapeutic Target for Liver Fibrosis. Front. Cell. Infect. Microbiol. 2022, 12, 1416. [Google Scholar] [CrossRef] [PubMed]
- Koppel, N.; Maini Rekdal, V.; Balskus, E.P. Chemical Transformation of Xenobiotics by the Human Gut Microbiota. Science 2017, 356, eaag2770. [Google Scholar] [CrossRef]
- Abdelsalam, N.A.; Ramadan, A.T.; ElRakaiby, M.T.; Aziz, R.K. Toxicomicrobiomics: The Human Microbiome vs. Pharmaceutical, Dietary, and Environmental Xenobiotics. Front. Pharmacol. 2020, 11, 390. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.; Zhai, Q.; Chen, W. Investigations of Bacteroides Spp. towards next-Generation Probiotics. Food Res. Int. 2019, 116, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations; World Health Organization (Eds.) Probiotics in Food: Health and Nutritional Properties and Guidelines for Evaluation; FAO food and nutrition paper; Food and Agriculture Organization of the United Nations; World Health Organization: Rome, Italy, 2006; ISBN 978-92-5-105513-7. [Google Scholar]
- Zhao, S.; Liu, W.; Wang, J.; Shi, J.; Sun, Y.; Wang, W.; Ning, G.; Liu, R.; Hong, J. Akkermansia muciniphila Improves Metabolic Profiles by Reducing Inflammation in Chow Diet-Fed Mice. J. Mol. Endocrinol. 2017, 58, 1–14. [Google Scholar] [CrossRef]
- López-Almela, I.; Romaní-Pérez, M.; Bullich-Vilarrubias, C.; Benítez-Páez, A.; Gómez Del Pulgar, E.M.; Francés, R.; Liebisch, G.; Sanz, Y. Bacteroides uniformis Combined with Fiber Amplifies Metabolic and Immune Benefits in Obese Mice. Gut Microbes 2021, 13, 1–20. [Google Scholar] [CrossRef]
- Yang, J.-Y.; Lee, Y.-S.; Kim, Y.; Lee, S.-H.; Ryu, S.; Fukuda, S.; Hase, K.; Yang, C.-S.; Lim, H.S.; Kim, M.-S.; et al. Gut Commensal Bacteroides Acidifaciens Prevents Obesity and Improves Insulin Sensitivity in Mice. Mucosal Immunol. 2017, 10, 104–116. [Google Scholar] [CrossRef] [Green Version]
- Plovier, H.; Everard, A.; Druart, C.; Depommier, C.; Van Hul, M.; Geurts, L.; Chilloux, J.; Ottman, N.; Duparc, T.; Lichtenstein, L.; et al. A Purified Membrane Protein from Akkermansia muciniphila or the Pasteurized Bacterium Improves Metabolism in Obese and Diabetic Mice. Nat. Med. 2017, 23, 107–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, F.; Guo, X.; Zhang, M.; Ou, Z.; Wu, D.; Deng, L.; Lu, Z.; Zhang, J.; Deng, G.; Chen, S.; et al. An Akkermansia muciniphila Subtype Alleviates High-Fat Diet-Induced Metabolic Disorders and Inhibits the Neurodegenerative Process in Mice. Anaerobe 2020, 61, 102138. [Google Scholar] [CrossRef] [PubMed]
- Fabersani, E.; Portune, K.; Campillo, I.; López-Almela, I.; la Paz, S.M.; Romaní-Pérez, M.; Benítez-Páez, A.; Sanz, Y. Bacteroides Uniformis CECT 7771 Alleviates Inflammation within the Gut-Adipose Tissue Axis Involving TLR5 Signaling in Obese Mice. Sci. Rep. 2021, 11, 11788. [Google Scholar] [CrossRef]
- Munukka, E.; Rintala, A.; Toivonen, R.; Nylund, M.; Yang, B.; Takanen, A.; Hänninen, A.; Vuopio, J.; Huovinen, P.; Jalkanen, S.; et al. Faecalibacterium prausnitzii Treatment Improves Hepatic Health and Reduces Adipose Tissue Inflammation in High-Fat Fed Mice. ISME J. 2017, 11, 1667–1679. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Lee, Y.; Kim, Y.; Seo, Y.; Lee, H.; Ha, J.; Lee, J.; Choi, Y.; Oh, H.; Yoon, Y. Akkermansia muciniphila Prevents Fatty Liver Disease, Decreases Serum Triglycerides, and Maintains Gut Homeostasis. Appl. Environ. Microbiol. 2020, 86, e03004-19. [Google Scholar] [CrossRef] [PubMed]
- Grander, C.; Adolph, T.E.; Wieser, V.; Lowe, P.; Wrzosek, L.; Gyongyosi, B.; Ward, D.V.; Grabherr, F.; Gerner, R.R.; Pfister, A.; et al. Recovery of Ethanol-Induced Akkermansia muciniphila Depletion Ameliorates Alcoholic Liver Disease. Gut 2018, 67, 891–901. [Google Scholar] [CrossRef]
- Zhang, L.; Qin, Q.; Liu, M.; Zhang, X.; He, F.; Wang, G. Akkermansia muciniphila Can Reduce the Damage of Gluco/Lipotoxicity, Oxidative Stress and Inflammation, and Normalize Intestine Microbiota in Streptozotocin-induced Diabetic Rats. Pathog. Dis. 2018, 76, fty028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ou, Z.; Deng, L.; Lu, Z.; Wu, F.; Liu, W.; Huang, D.; Peng, Y. Protective Effects of Akkermansia muciniphila on Cognitive Deficits and Amyloid Pathology in a Mouse Model of Alzheimer’s Disease. Nutr. Diabetes 2020, 10, 12. [Google Scholar] [CrossRef] [Green Version]
- Gómez del Pulgar, E.M.; Benítez-Páez, A.; Sanz, Y. Safety Assessment of Bacteroides uniformis CECT 7771, a Symbiont of the Gut Microbiota in Infants. Nutrients 2020, 12, 551. [Google Scholar] [CrossRef] [Green Version]
- Jia, L.; Shan, K.; Pan, L.-L.; Feng, N.; Lv, Z.; Sun, Y.; Li, J.; Wu, C.; Zhang, H.; Chen, W.; et al. Clostridium butyricum CGMCC0313.1 Protects against Autoimmune Diabetes by Modulating Intestinal Immune Homeostasis and Inducing Pancreatic Regulatory T Cells. Front. Immunol. 2017, 8, 1345. [Google Scholar] [CrossRef] [Green Version]
- Péan, N.; Le Lay, A.; Brial, F.; Wasserscheid, J.; Rouch, C.; Vincent, M.; Myridakis, A.; Hedjazi, L.; Dumas, M.-E.; Grundberg, E.; et al. Dominant Gut Prevotella copri in Gastrectomised Non-Obese Diabetic Goto–Kakizaki Rats Improves Glucose Homeostasis through Enhanced FXR Signalling. Diabetologia 2020, 63, 1223–1235. [Google Scholar] [CrossRef] [Green Version]
- Zhai, R.; Xue, X.; Zhang, L.; Yang, X.; Zhao, L.; Zhang, C. Strain-Specific Anti-Inflammatory Properties of Two Akkermansia muciniphila Strains on Chronic Colitis in Mice. Front. Cell. Infect. Microbiol. 2019, 9, 239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres-Sánchez, A.; Ruiz-Rodríguez, A.; Ortiz, P.; Moreno, M.A.; Ampatzoglou, A.; Gruszecka-Kosowska, A.; Monteoliva-Sánchez, M.; Aguilera, M. Exploring Next Generation Probiotics for Metabolic and Microbiota Dysbiosis Linked to Xenobiotic Exposure: Holistic Approach. Int. J. Mol. Sci. 2022, 23, 12917. [Google Scholar] [CrossRef] [PubMed]
- Del Chierico, F.; Manco, M.; Gardini, S.; Guarrasi, V.; Russo, A.; Bianchi, M.; Tortosa, V.; Quagliariello, A.; Shashaj, B.; Fintini, D.; et al. Fecal Microbiota Signatures of Insulin Resistance, Inflammation, and Metabolic Syndrome in Youth with Obesity: A Pilot Study. Acta Diabetol. 2021, 58, 1009–1022. [Google Scholar] [CrossRef]
- Ahmad, A.; Yang, W.; Chen, G.; Shafiq, M.; Javed, S.; Zaidi, S.S.A.; Shahid, R.; Liu, C.; Bokhari, H. Analysis of Gut Microbiota of Obese Individuals with Type 2 Diabetes and Healthy Individuals. PLoS ONE 2019, 14, e0226372. [Google Scholar] [CrossRef] [Green Version]
- Doumatey, A.P.; Adeyemo, A.; Zhou, J.; Lei, L.; Adebamowo, S.N.; Adebamowo, C.; Rotimi, C.N. Gut Microbiome Profiles Are Associated with Type 2 Diabetes in Urban Africans. Front. Cell. Infect. Microbiol. 2020, 10, 63. [Google Scholar] [CrossRef] [Green Version]
- Adachi, K.; Sugiyama, T.; Yamaguchi, Y.; Tamura, Y.; Izawa, S.; Hijikata, Y.; Ebi, M.; Funaki, Y.; Ogasawara, N.; Goto, C.; et al. Gut Microbiota Disorders Cause Type 2 Diabetes Mellitus and Homeostatic Disturbances in Gut-Related Metabolism in Japanese Subjects. J. Clin. Biochem. Nutr. 2019, 64, 231–238. [Google Scholar] [CrossRef] [Green Version]
- Takagi, T.; Naito, Y.; Kashiwagi, S.; Uchiyama, K.; Mizushima, K.; Kamada, K.; Ishikawa, T.; Inoue, R.; Okuda, K.; Tsujimoto, Y.; et al. Changes in the Gut Microbiota Are Associated with Hypertension, Hyperlipidemia, and Type 2 Diabetes Mellitus in Japanese Subjects. Nutrients 2020, 12, 2996. [Google Scholar] [CrossRef] [PubMed]
- Mejía-León, M.E.; Petrosino, J.F.; Ajami, N.J.; Domínguez-Bello, M.G.; de la Barca, A.M.C. Fecal Microbiota Imbalance in Mexican Children with Type 1 Diabetes. Sci. Rep. 2014, 4, 3814. [Google Scholar] [CrossRef] [Green Version]
- Radwan, S.; Gilfillan, D.; Eklund, B.; Radwan, H.M.; Menofy, N.G.E.; Lee, J.; Kapuscinski, M.; Abdo, Z. A Comparative Study of the Gut Microbiome in Egyptian Patients with Type I and Type II Diabetes. PLoS ONE 2020, 15, e0238764. [Google Scholar] [CrossRef]
- Gradisteanu Pircalabioru, G.; Ilie, I.; Oprea, L.; Picu, A.; Petcu, L.M.; Burlibasa, L.; Chifiriuc, M.-C.; Musat, M. Microbiome, Mycobiome and Related Metabolites Alterations in Patients with Metabolic Syndrome—A Pilot Study. Metabolites 2022, 12, 218. [Google Scholar] [CrossRef]
- Lim, M.Y.; You, H.J.; Yoon, H.S.; Kwon, B.; Lee, J.Y.; Lee, S.; Song, Y.-M.; Lee, K.; Sung, J.; Ko, G. The Effect of Heritability and Host Genetics on the Gut Microbiota and Metabolic Syndrome. Gut 2017, 66, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Cortez, R.V.; Moreira, L.N.; Padilha, M.; Bibas, M.D.; Toma, R.K.; Porta, G.; Taddei, C.R. Gut Microbiome of Children and Adolescents with Primary Sclerosing Cholangitis in Association with Ulcerative Colitis. Front. Immunol. 2021, 11, 598152. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhang, Y.; Jiang, H.; Xiang, S.; Zhao, Y.; Xiao, M.; Du, F.; Ji, H.; Kaboli, P.J.; Wu, X.; et al. Metagenome Analysis of Intestinal Bacteria in Healthy People, Patients with Inflammatory Bowel Disease and Colorectal Cancer. Front. Cell. Infect. Microbiol. 2021, 11, 599734. [Google Scholar] [CrossRef] [PubMed]
- Jee, J.J.; Lim, J.; Park, S.; Koh, H.; Lee, H.W. Gut Microbial Community Differentially Characterizes Patients with Nonalcoholic Fatty Liver Disease. J. Gastroenterol. Hepatol. 2022, 37, 1822–1832. [Google Scholar] [CrossRef]
- Yu, J.; Zhang, H.; Chen, L.; Ruan, Y.; Chen, Y.; Liu, Q. Disease-Associated Gut Microbiota Reduces the Profile of Secondary Bile Acids in Pediatric Nonalcoholic Fatty Liver Disease. Front. Cell. Infect. Microbiol. 2021, 11, 698852. [Google Scholar] [CrossRef] [PubMed]
- Iino, C.; Endo, T.; Mikami, K.; Hasegawa, T.; Kimura, M.; Sawada, N.; Nakaji, S.; Fukuda, S. Significant Decrease in Faecalibacterium among Gut Microbiota in Nonalcoholic Fatty Liver Disease: A Large BMI- and Sex-Matched Population Study. Hepatol. Int. 2019, 13, 748–756. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-N.; Joo, E.-J.; Cheong, H.S.; Kim, Y.; Kim, H.-L.; Shin, H.; Chang, Y.; Ryu, S. Gut Microbiota and Risk of Persistent Nonalcoholic Fatty Liver Diseases. J. Clin. Med. 2019, 8, 1089. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Sun, G.; Wang, Z.; Wu, W.; Guo, H.; Peng, L.; Wu, L.; Guo, X.; Yang, Y. Characteristics of Fecal Microbiota in Non-Alcoholic Fatty Liver Disease Patients. Sci. China Life Sci. 2018, 61, 770–778. [Google Scholar] [CrossRef]
- Shen, F.; Zheng, R.-D.; Sun, X.-Q.; Ding, W.-J.; Wang, X.-Y.; Fan, J.-G. Gut Microbiota Dysbiosis in Patients with Non-Alcoholic Fatty Liver Disease. Hepatobiliary Pancreat. Dis. Int. 2017, 16, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; You, H.J.; Bajaj, J.S.; Joo, S.K.; Yu, J.; Park, S.; Kang, H.; Park, J.H.; Kim, J.H.; Lee, D.H.; et al. Distinct Signatures of Gut Microbiome and Metabolites Associated with Significant Fibrosis in Non-Obese NAFLD. Nat. Commun. 2020, 11, 4982. [Google Scholar] [CrossRef]
- Wang, B.; Jiang, X.; Cao, M.; Ge, J.; Bao, Q.; Tang, L.; Chen, Y.; Li, L. Altered Fecal Microbiota Correlates with Liver Biochemistry in Nonobese Patients with Non-Alcoholic Fatty Liver Disease. Sci. Rep. 2016, 6, 32002. [Google Scholar] [CrossRef]
- Tsai, M.-C.; Liu, Y.-Y.; Lin, C.-C.; Wang, C.-C.; Wu, Y.-J.; Yong, C.-C.; Chen, K.-D.; Chuah, S.-K.; Yao, C.-C.; Huang, P.-Y.; et al. Gut Microbiota Dysbiosis in Patients with Biopsy-Proven Nonalcoholic Fatty Liver Disease: A Cross-Sectional Study in Taiwan. Nutrients 2020, 12, 820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loomba, R.; Seguritan, V.; Li, W.; Long, T.; Klitgord, N.; Bhatt, A.; Dulai, P.S.; Caussy, C.; Bettencourt, R.; Highlander, S.K.; et al. Gut Microbiome-Based Metagenomic Signature for Non-Invasive Detection of Advanced Fibrosis in Human Nonalcoholic Fatty Liver Disease. Cell Metab. 2017, 25, 1054–1062.e5. [Google Scholar] [CrossRef] [PubMed]
- Sobhonslidsuk, A.; Chanprasertyothin, S.; Pongrujikorn, T.; Kaewduang, P.; Promson, K.; Petraksa, S.; Ongphiphadhanakul, B. The Association of Gut Microbiota with Nonalcoholic Steatohepatitis in Thais. BioMed Res. Int. 2018, 2018, e9340316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, H.E.; Teterina, A.; Comelli, E.M.; Taibi, A.; Arendt, B.M.; Fischer, S.E.; Lou, W.; Allard, J.P. Nonalcoholic Fatty Liver Disease Is Associated with Dysbiosis Independent of Body Mass Index and Insulin Resistance. Sci. Rep. 2018, 8, 1466. [Google Scholar] [CrossRef] [Green Version]
- Mouzaki, M.; Comelli, E.M.; Arendt, B.M.; Bonengel, J.; Fung, S.K.; Fischer, S.E.; McGilvray, I.D.; Allard, J.P. Intestinal Microbiota in Patients with Nonalcoholic Fatty Liver Disease. Hepatology 2013, 58, 120–127. [Google Scholar] [CrossRef] [Green Version]
- Bastian, W.P.; Hasan, I.; Lesmana, C.R.A.; Rinaldi, I.; Gani, R.A. Gut Microbiota Profiles in Nonalcoholic Fatty Liver Disease and Its Possible Impact on Disease Progression Evaluated with Transient Elastography: Lesson Learnt from 60 Cases. Case Rep. Gastroenterol. 2019, 13, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Dong, Y.; Zuo, K.; Han, C.; Jiao, J.; Yang, X.; Li, J. Characteristics and Variation of Fecal Bacterial Communities and Functions in Isolated Systolic and Diastolic Hypertensive Patients. BMC Microbiol. 2021, 21, 128. [Google Scholar] [CrossRef]
- Nakai, M.; Ribeiro, R.V.; Stevens, B.R.; Gill, P.; Muralitharan, R.R.; Yiallourou, S.; Muir, J.; Carrington, M.; Head, G.A.; Kaye, D.M.; et al. Essential Hypertension Is Associated with Changes in Gut Microbial Metabolic Pathways: A Multisite Analysis of Ambulatory Blood Pressure. Hypertension 2021, 78, 804–815. [Google Scholar] [CrossRef] [PubMed]
- Silveira-Nunes, G.; Durso, D.F.; de Oliveira, L.R.A., Jr.; Cunha, E.H.M.; Maioli, T.U.; Vieira, A.T.; Speziali, E.; Corrêa-Oliveira, R.; Martins-Filho, O.A.; Teixeira-Carvalho, A.; et al. Hypertension Is Associated With Intestinal Microbiota Dysbiosis and Inflammation in a Brazilian Population. Front. Pharmacol. 2020, 11, 258. [Google Scholar] [CrossRef] [Green Version]
- Yan, Q.; Gu, Y.; Li, X.; Yang, W.; Jia, L.; Chen, C.; Han, X.; Huang, Y.; Zhao, L.; Li, P.; et al. Alterations of the Gut Microbiome in Hypertension. Front. Cell. Infect. Microbiol. 2017, 7, 381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Zhao, F.; Wang, Y.; Chen, J.; Tao, J.; Tian, G.; Wu, S.; Liu, W.; Cui, Q.; Geng, B.; et al. Gut Microbiota Dysbiosis Contributes to the Development of Hypertension. Microbiome 2017, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Wan, C.; Zhu, C.; Jin, G.; Zhu, M.; Hua, J.; He, Y. Analysis of Gut Microbiota in Patients with Coronary Artery Disease and Hypertension. Evid.-Based Complement. Altern. Med. 2021, 2021, 7195082. [Google Scholar] [CrossRef]
- Li, H.; Liu, B.; Song, J.; An, Z.; Zeng, X.; Li, J.; Jiang, J.; Xie, L.; Wu, W. Characteristics of Gut Microbiota in Patients with Hypertension and/or Hyperlipidemia: A Cross-Sectional Study on Rural Residents in Xinxiang County, Henan Province. Microorganisms 2019, 7, 399. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Wang, J.; Zhao, L.-L.; Yao, T.-T.; Chen, Y.; Ma, J.; Zhang, X.; Wang, J.-X.; Wang, Y.; Cui, Z.; et al. Gut Lactobacillus Level Is a Predictive Marker for Coronary Atherosclerotic Lesions Progress and Prognosis in Patients with Acute Coronary Syndrome. Front. Cell. Infect. Microbiol. 2021, 11, 687827. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Chen, S.; Gu, G.; Zhou, J.; Wang, W.; Ren, J.; Wu, J.; Yang, D.; Zheng, Y. Exploration of Crucial Mediators for Carotid Atherosclerosis Pathogenesis through Integration of Microbiome, Metabolome, and Transcriptome. Front. Physiol. 2021, 12, 645212. [Google Scholar] [CrossRef] [PubMed]
- Baragetti, A.; Severgnini, M.; Olmastroni, E.; Dioguardi, C.C.; Mattavelli, E.; Angius, A.; Rotta, L.; Cibella, J.; Caredda, G.; Consolandi, C.; et al. Gut Microbiota Functional Dysbiosis Relates to Individual Diet in Subclinical Carotid Atherosclerosis. Nutrients 2021, 13, 304. [Google Scholar] [CrossRef]
- Liu, S.; Zhao, W.; Liu, X.; Cheng, L. Metagenomic Analysis of the Gut Microbiome in Atherosclerosis Patients Identify Cross-Cohort Microbial Signatures and Potential Therapeutic Target. FASEB J. 2020, 34, 14166–14181. [Google Scholar] [CrossRef]
- Toya, T.; Corban, M.T.; Marrietta, E.; Horwath, I.E.; Lerman, L.O.; Murray, J.A.; Lerman, A. Coronary Artery Disease Is Associated with an Altered Gut Microbiome Composition. PLoS ONE 2020, 15, e0227147. [Google Scholar] [CrossRef]
- Liu, H.; Chen, X.; Hu, X.; Niu, H.; Tian, R.; Wang, H.; Pang, H.; Jiang, L.; Qiu, B.; Chen, X.; et al. Alterations in the Gut Microbiome and Metabolism with Coronary Artery Disease Severity. Microbiome 2019, 7, 68. [Google Scholar] [CrossRef] [Green Version]
- Jie, Z.; Xia, H.; Zhong, S.-L.; Feng, Q.; Li, S.; Liang, S.; Zhong, H.; Liu, Z.; Gao, Y.; Zhao, H.; et al. The Gut Microbiome in Atherosclerotic Cardiovascular Disease. Nat. Commun. 2017, 8, 845. [Google Scholar] [CrossRef] [Green Version]
- Loftfield, E.; Herzig, K.-H.; Caporaso, J.G.; Derkach, A.; Wan, Y.; Byrd, D.A.; Vogtmann, E.; Männikkö, M.; Karhunen, V.; Knight, R.; et al. Association of Body Mass Index with Fecal Microbial Diversity and Metabolites in the Northern Finland Birth Cohort. Cancer Epidemiol. Biomark. Prev. 2020, 29, 2289–2299. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Ni, Z.; Yu, J.; Cheng, W.; Cai, Z.; Yu, C. Correlation Between Fecal Metabolomics and Gut Microbiota in Obesity and Polycystic Ovary Syndrome. Front. Endocrinol. 2020, 11, 628. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Hong, J.; Xu, X.; Feng, Q.; Zhang, D.; Gu, Y.; Shi, J.; Zhao, S.; Liu, W.; Wang, X.; et al. Gut Microbiome and Serum Metabolome Alterations in Obesity and after Weight-Loss Intervention. Nat. Med. 2017, 23, 859–868. [Google Scholar] [CrossRef]
- Nogacka, A.M.; de los Reyes-Gavilán, C.G.; Martínez-Faedo, C.; Ruas-Madiedo, P.; Suarez, A.; Mancabelli, L.; Ventura, M.; Cifuentes, A.; León, C.; Gueimonde, M.; et al. Impact of Extreme Obesity and Diet-Induced Weight Loss on the Fecal Metabolome and Gut Microbiota. Mol. Nutr. Food Res. 2021, 65, 2000030. [Google Scholar] [CrossRef] [PubMed]
- Nuli, R.; Azhati, J.; Cai, J.; Kadeer, A.; Zhang, B.; Mohemaiti, P. Metagenomics and Faecal Metabolomics Integrative Analysis towards the Impaired Glucose Regulation and Type 2 Diabetes in Uyghur-Related Omics. J. Diabetes Res. 2019, 2019, e2893041. [Google Scholar] [CrossRef] [Green Version]
- Qi, Q.; Li, J.; Yu, B.; Moon, J.-Y.; Chai, J.C.; Merino, J.; Hu, J.; Ruiz-Canela, M.; Rebholz, C.; Wang, Z.; et al. Host and Gut Microbial Tryptophan Metabolism and Type 2 Diabetes: An Integrative Analysis of Host Genetics, Diet, Gut Microbiome and Circulating Metabolites in Cohort Studies. Gut 2021, 71, 1095–1105. [Google Scholar] [CrossRef]
- Zhu, S.; Liu, S.; Li, H.; Zhang, Z.; Zhang, Q.; Chen, L.; Zhao, Y.; Chen, Y.; Gu, J.; Min, L.; et al. Identification of Gut Microbiota and Metabolites Signature in Patients With Irritable Bowel Syndrome. Front. Cell. Infect. Microbiol. 2019, 9, 346. [Google Scholar] [CrossRef] [Green Version]
- Alferink, L.J.M.; Radjabzadeh, D.; Erler, N.S.; Vojinovic, D.; Medina-Gomez, C.; Uitterlinden, A.G.; de Knegt, R.J.; Amin, N.; Ikram, M.A.; Janssen, H.L.A.; et al. Microbiomics, Metabolomics, Predicted Metagenomics, and Hepatic Steatosis in a Population-Based Study of 1,355 Adults. Hepatology 2021, 73, 968–982. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Kolde, R. Pheatmap: Pretty Heatmaps 2019. Pheatmap: Pretty Heatmaps. R Package Version 1.0.12. Available online: https://CRAN.R-project.org/package=pheatmap (accessed on 28 January 2023).
- Chen, M.X.; Wang, S.-Y.; Kuo, C.-H.; Tsai, I.-L. Metabolome Analysis for Investigating Host-Gut Microbiota Interactions. J. Formos. Med. Assoc. 2019, 118 (Suppl. S1), S10–S22. [Google Scholar] [CrossRef]
- Visconti, A.; Le Roy, C.I.; Rosa, F.; Rossi, N.; Martin, T.C.; Mohney, R.P.; Li, W.; de Rinaldis, E.; Bell, J.T.; Venter, J.C.; et al. Interplay between the Human Gut Microbiome and Host Metabolism. Nat. Commun. 2019, 10, 4505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Wang, N.; Tan, H.-Y.; Li, S.; Zhang, C.; Feng, Y. Function of Akkermansia muciniphila in Obesity: Interactions with Lipid Metabolism, Immune Response and Gut Systems. Front. Microbiol. 2020, 11, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maioli, T.U.; Borras-Nogues, E.; Torres, L.; Barbosa, S.C.; Martins, V.D.; Langella, P.; Azevedo, V.A.; Chatel, J.-M. Possible Benefits of Faecalibacterium prausnitzii for Obesity-Associated Gut Disorders. Front. Pharmacol. 2021, 12, 740636. [Google Scholar] [CrossRef]
- López-Moreno, A.; Suárez, A.; Avanzi, C.; Monteoliva-Sánchez, M.; Aguilera, M. Probiotic Strains and Intervention Total Doses for Modulating Obesity-Related Microbiota Dysbiosis: A Systematic Review and Meta-Analysis. Nutrients 2020, 12, 1921. [Google Scholar] [CrossRef] [PubMed]
- Gao, P. The Exposome in the Era of One Health. Environ. Sci. Technol. 2021, 55, 2790–2799. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ref. | Sample Size and Clinical Traits | Gut Microbiota Taxa Modification |
|---|---|---|
| [1] | n = 42; HC n = 21; OB n = 21 | ↑ Prevotella, Megamonas, Blautia, and Fusobacterium, ↓ Alistipes, Faecalibacterium, Oscillibacter, Clostridium IV, XIVa, Barnesiella, Gemmiger, Parabacteroides, Coprococcus, Ruminococcus, and Bifidobacterium in OB |
| [2] | n = 51; HC n = 30; OB/OW n = 21 | ↑ Lactobacillus *, ↓ Bifidobacterium in OB/OW |
| [3] | n = 51; HC n = 23; OB/OW n = 28 | ↑ Faecalibacterium, Phascolarctobacterium, Lachnospira, Megamonas, and Haemophilus, ↓ Oscillospira, and Dialister in OB |
| [4] | n = 192; HC n = 25; OW n = 22; OB n = 145 | ↑ Escherichia coli, Pseudomonas, Fusobacterium, ↓ Bifidobacterium in OW/OB |
| [5] | n = 143; HC n = 56; OB n = 87 | ↑ Enterococcus, Blautia, Sutterella, Klebsiella, and Collinsella, ↓ Bacteroides, Parabacteroides, Anaerotruncus, and Coprobacillus in OB |
| [6] | n = 78; HC n = 36; OB n = 42 | ↓ Bacteroides in OB |
| [23] | n = 66; HC n = 27; OB n = 17; OBT2D n = 22 | ↑ Staphylococcus in OB; ↑ Lactobacillus * and Escherichia in T2D |
| [7] | OW n = 34; OB n = 23; AbOB n = 53; Dys n = 78; IFG n = 21; IGT n = 3; T2D n = 21; HT n = 34 | ↑ Serratia and Prevotella, ↓ Oscillospira in OW, OB, AbOB group; ↑ Blautia in T2D; ↑ Prevotella in HT |
| [8] | n = 58; HC n = 15; OB n = 18; OB NAFLD n = 25 | ↑ Phascolarctobacterium, Phascolarctobacterium succinatutens, Klebsiella, Klebsiella pneumoniae, Kluyvera, and Kluyvera ascorbata, ↓ Lactobacillus *, Oscillibacter, Ruminiclostridium, and Parabacteroides johnsonii in OB NAFLD; ↓ Alistipes, Paraprevotella, Bacteroides clarus, and Odoribacter splanchnicus in OB and OB NAFLD; ↓ Helicobacter, Helicobacter pylori in OB |
| [9] | n = 73; HC n = 20; OB NAFLD n = 36; OB Non-NAFLD n = 17 | ↑ Megasphaera, Lactobacillus *, and Acidaminococcus, ↓ Oscillospira, Eubacterium, and Akkermansia in OB NAFLD and OB Non-NAFLD; ↑ Streptococcus, ↓ Blautia, Alkaliphilus, and Flavobacterium in OB NAFLD |
| [10] | n = 115; HC n = 54; OB n = 8; NAFLD n = 27; NASH n = 26 | ↑ Bradyrhizobium, Anaerococcus, Peptoniphilus, Propionibacterium acnes, Dorea, and Ruminococcus, ↓ Oscillospira in NAFLD, NASH and OB vs. HC |
| [11] | n = 23; HC n = 10; NASH n = 13 | ↑ Lactobacillus * in (OB-NASH vs. LN-HC), (OB-NASH vs. OB-HC) and (OB-NASH vs. OW-NASH); ↑ Lachnospira in (OB-NASH vs. OB-HC); ↓ Roseburia in (OB-NASH vs. LN-HC) and (OB-NASH vs. OB-HC); ↓ Bifidobacterium in (OW-NASH vs. LN-HC); ↓ Faecalibacterium and Ruminococcus in (LN-NASH vs. LN-HC) and (LN-NASH vs. OB-HC); ↓ Ruminococcus in (LN-NASH vs. OB-NASH) and (LN-NASH vs. OW-NASH) |
| [64] | n = 106; HC n = 38; OB n = 68 | ↑ Clostridium in HT; ↑ Bacteroides in IGT |
| [12] | n = 119; OB n = 69; Mets n = 50 | ↑ Intestinibacter, Saccharibacteria genera incertae sedis, Clostridium sensu stricto, Romboutsia, Terrisporobacter, and Eggerthia, ↓ Rothia, Adlercreutzia, Parabacteroides, Paraprevotella, Alistipes, Bacteroides, Bilophila, Escherichia-Shigella, Lactobacillus *, Clostridium XIVa, Clostridium XIVb, Anaerotruncus, and Phascolarctobacterium in OB vs. Mets |
| [65] | n = 60; HC n = 20; OB T2D n = 40 | ↑ Eubacterium coprostanoligenes group, Dialister, and Allisonella, ↓ Ruminococcus 2, Prevotella 9, and Escherichia-Shigella 9 in OB T2D |
| [14] | n = 1280; LN-NonT2D n = 633; OB-NonT2D n = 494; OBT2D n = 153 | ↓ Akkermansia, Faecalibacterium, Oscillibacter, and Alistipes in OB- NonT2D and OBT2D |
| [15] | n = 50; HC n = 15; T2D n = 14; DR n = 21 | ↑ Klebsiella and Enterococcus, ↓ Faecalibacterium and Lachnospira in T2D |
| [16] | n = 154; CN n = 73; T2DCI n = 81 | ↑ Peptococcus, ↓ Bifidobacterium, Veillonella, and Pediococcus in T2DCI |
| [66] | n = 291; HC n = 193; T2D n = 98 | ↑ Peptostreptococcus, Eubacterium, and Prevotella, ↓ Anaerostipes, Ruminococcus, Clostridium, Epulopiscium, Cellulosilyticum ruminicola, Clostridium paraputrificum, and Clostridium butyricum in T2D |
| [17] | n = 60; HC n = 40; T2D n = 20 | ↑ Streptococcus, Fusobacterium, and Dorea, ↓ Parabacteroides, Bifidobacterium, Faecalibacterium, and Akkermansia in T2D |
| [24] | n = 102; HC n = 35; pT2D n = 17; NewT2D n = 11; KnownT2D n = 39 | ↑ Escherichia and Acidaminococcus, ↓ Sutterella in KnownT2D; ↑ Megasphaera and Lactobacillus *, ↓ Akkermansia, Blautia, and Ruminococcus in NewT2D |
| [67] | n = 118; HC n = 59; T2D n = 59 | ↑ Bifidobacterium spp., ↓ Bacteroides spp. in T2D |
| [25] | n = 100; HC n = 35; T2D+ n = 49; T2D− n = 16 | ↑ Coprococcus 1, ↓ Bacteroides and Prevotella in T2D+ and T2D- vs. HC; ↑ Parasutterella in T2D+ vs. HC; ↑ Blautia and Eubacterium hallii group in T2D−vs. HC |
| [26] | n = 100; HC n = 50; T2D n = 50 | ↑ Lactobacillus *, ↓ Clostridium leptum and Clostridium coccoides in T2D |
| [18] | n = 36; HC n = 18; T2D n = 18 | ↓ Faecalibacterium prausnitzii in T2D |
| [19] | n = 36; HC n = 18; T2D n = 18 | ↑ Lactobacillus *, ↓ Bifidobacterium in T2D |
| [68] | n = 239; HC n = 54; HT n = 97; HL n = 96; T2D n = 162 | ↑ Bifidobacterium in HL, T2D, RISK1, and RISK2; ↑ Collinsella in HT, HL, T2D, RISK2, and RISK3; ↑ Escherichia in RISK3; ↓ Alistipes in HL |
| [27] | n = 98; HC n = 47; T1D n = 51 | ↑ Blautia, Anaerostipes, Eubacterium hallii group, Dorea, Collinsella, and Klebsiella, ↓ Parabacteroides and Flavonifractor in T1D |
| [69] | n = 29; HC n = 8; T1D at onset n = 8; T1D two years treatment n = 13 | ↑ Bacteroides, ↓ Prevotella, Megamonas, and Acidaminococcus in T1D at onset |
| [70] | n = 47; HC n = 7; T1D n = 22; T2D n = 18 | ↑ Pseudomonas and Prevotella in T1D and T2D vs. HC |
| [20] | n = 110; HC n = 40; T1D n = 21; T2D n = 49 | ↑ Escherichia, Prevotella, and Lactobacillus *, ↓ Bacteroides, Roseburia, and Bifidobacterium in T1D and T2D; ↓ Faecalibacterium in T1D vs. T2D and HC |
| [21] | n = 43; HC n = 13; T1D n = 15; MODY2 n = 15 | ↑ Bacteroides, Ruminococcus, Blautia, Veillonella, Streptococcus, Sutterella, and Enterobacter, ↓ Bifidobacterium in T1D; ↑ Prevotella ↓ Lachnospira, Roseburia, Anaerostipes, and Faecalibacterium in T1D and MODY2 |
| [71] | n = 60; HC n = 30; Metsyn patients n = 30 | ↑ Clostridium leptum, Clostridium coccoides group, and Turicibacter sp., ↓ Butyricicoccus sp., Faecalibacterium prausnitzii, and Akkermansia muciniphila in Mets |
| [72] | n = 655; MZ n = 306; DZ n = 74, Nontwin n = 275 | ↑ Lactobacillus *, Sutterella, Dorea, and Methanobrevibacter, ↓ Parabacteroides, Bifidobacterium, Odoribacter, Akkermansia, and Paraprevotella in Mets |
| [13] | n = 20; No Mets + NGT n = 4; No Mets + IFG n = 3; No Mets + IFG + IGT n = 1; Mets + IFG n = 4; Mets + IFG + IGT n = 4; Mets + T2D n = 4 | ↑ Ruminococcus, Dorea, Blautia, and Oscillospira in OB, Mets, IFG, IFG + IGT, and T2D |
| [28] | n = 41; HC n = 20; UC n = 21 | ↓ Ruminococcus and Faecalibacterium prausnitzii in UC |
| [29] | n = 20; HC n = 10; UC n = 10 | ↑ Escherichia-Shigella, Peptostreptococcus, Bacillus, and Veillonella, ↓ Akkermansia, Faecalibacterium, and Bifdobacterium in UC |
| [30] | n = 42; HC n = 14; UC n = 28 | ↑ Streptococcus, Escherichia-Shigella, Romboutsia, Clostridium sensu stricto, Enterococcus, and Citrobacter, ↓ Faecalibacterium, Agathobacter, Dorea, Ruminococcus, Prevotella, Alistipes, Parabacteroides, and Butyricicoccus in UC |
| [73] | n = 53; HC n = 23; UC n = 12; PSC n = 11; PSC + UC n = 7 | ↑ Bifidobacterium in UC |
| [31] | n = 24; HC n = 12; CD n = 6; UC n = 6 | ↑ Clostridium ramosum, Escherichia coli, Fusobacterium nucleatum, and Ruminococcus gnavus, ↓ Eubacterium rectale, and Faecalibacterium prausnitzii in UC |
| [32] | n = 58; HC n = 29; UC n = 22; CD n = 7 | ↓ Bacteroides, Faecalibacterium prausnitzii, Prevotella spp., and Methanobrevibacterium spp. in IBD |
| [33] | n = 42; HC n = 13; CD n = 15; UC n = 14 | ↑ Abiotrophia, Pseudoramibacter, Eubacterium, and Escherichia, ↓ Butyricicoccus, Mitsuokella, Haemophilus, and Victivallis in CD; ↑ Granulicatella, Peptostreptococcus, Schwartzia, Capnocytophaga, Escherichia, Janthinobacterium, Campylobacter, Actinomyces, Eggerthella, and Corynebacterium, ↓ Holdemania, Lachnobacterium, Megamonas, Mitsuokella, Alistipes, Butyricimonas, Prevotella, Desulfovibrio, Oxalobacter, Pyramidobacter, and Victivallis in UC; ↑ Pseudoramibacter Eubacterium, Desulfovibrio, and Slackia, ↓ Butyricicoccus, Moryella, Staphylococcus, Capnocytophaga, Haemophilus, Janthinobacterium, Cardiobacterium, Lautropia, Lupinus, Shewanella, and Corynebacterium in CD/UC |
| [34] | n = 155; Non-IBD n = 34; CD n = 68; UC n = 53 | ↑ Unclassified Roseburia species in CD and UC; ↑ Bifidobacterium breve and Clostridium symbiosum in UC; ↑ Blautia producta, Lactobacillus gasseri, Enterococcus faecium, Clostridium clostridioforme, Ruminococcus gnavus, and Escherichia coli in CD |
| [74] | n = 1087; HC n = 290; IBD n = 512; CRC n = 285 | ↑ Bacteroides in IBD |
| [35] | n = 68; HC n = 48; IBD n = 20 | ↑Bifidobacterium, Ruminococcus gnavus group, Streptococcus, and Blautia, ↓ Faecalibacterium, Subdoligranulum, Parabacteroides, and Paraprevotella in IBD |
| [36] | n = 30; HC n = 8; DD n = 4; IBS n = 3; UC n = 5; CD n = 10 | ↑ Dialister spp. And Faecalibacterium prausnitzii in IBS; ↑ Bacteroides fragilis, Dialister spp., and Roseburia spp. ↓ Clostridium difficile in UC vs. HC; ↑ Parabacteroides distasonis ↓ Faecalibacterium prausnitzii, and Bacteroides fragilis in CD |
| [37] | n = 69; HC n = 40; Non-PN SBS n = 5; SBS I n = 10; SBS II n = 14 | ↑ Lactobacillus * and Klebsiella, ↓ Coprococcus, Faecalibacterium, Lachnospira, and Ruminococcus in SBS patients; ↓ Blautia, Bacteroides, Odoribacter, Oscillospira, Prevotella, Roseburia, and Sutterella in SBS I and SBS II; ↑ Streptococcus and Staphylococcus in SBS I |
| [75] | n = 16 NAFLD | ↑ Prevotella copri and Prevotella stercorea in NAFLD |
| [76] | n = 68; HC n = 36; NAFLD n = 32 | ↑ Escherichia coli, Klebsiella pneumoniae, and Enterobacter cloacae, ↓ Akkermansia muciniphila, Alistipes putredinis, Bacteroides uniformis, Bacteroides fragilis, Oscillibacter sp., Ruminococcus bromii, Eubacterium ventriosum, and Gemmiger formicilis in NAFLD |
| [77] | n = 874; Non-NAFLD n = 669; NAFLD n = 205 | ↓ Faecalibacterium and Bacteroides in NAFLD |
| [78] | n = 766; Control n = 453; Developed NAFLD n = 40; Regressed NAFLD n = 35; Persistent NAFLD n = 238 | ↓ Oscillospira, Odoribacter, and Coprococcus in persistent NAFLD vs. Control; ↓ Coprococcus eutactus in regressed NAFLD and persistent NAFLD vs. Control |
| [79] | n = 67; HC n = 37; NAFLD n = 30 | ↑ Porphyromonas, Succinivibrio, Clostridium, Blautia, Dorea, Peptococcus, Mitsuokella, and Slackia, ↓ Odoribacter, Proteus, and Coprococcus in NAFLD |
| [80] | n = 47; HC n = 22; NAFLD n = 25 | ↑ Escherichia-Shigella, Blautia, Clostridium XVIII, and Streptococcus, ↓ Prevotella and Faecalibacterium in NAFLD |
| [81] | n = 202; no-NAFLD n = 31; NAFLD n = 171 | ↑ Citrobacter, ↓ Coprococcus and Lachnospira in significant fibrosis |
| [82] | n = 126; no-NAFLD n = 83; NAFLD n = 43 | ↓ Coprococcus, Pseudobutyrivibrio, Moryella, Roseburia, Anaerosporobacter, Anaerotruncus, Ruminococcus, Lactobacillus * in NAFLD |
| [83] | n = 75; HC n = 25; NAFLD n = 25; NASH n = 25 | ↑ Bacteroides and Prevotella, ↓ Faecalibacterium in NAFLD and NASH |
| [84] | n = 86; Mild/moderate NAFLD n = 72; Fibrosis n = 14 | ↑ Eubacterium rectale in mild/moderate NAFLD; ↑ Bacteroides vulgatus and Escherichia coli, ↓ Ruminococcus obeum, and Eubacterium rectale in fibrosis |
| [85] | n = 24; HC n = 8; NASH n = 16 | ↑ Phascolarctobacterium in NASH |
| [86] | n = 67; HC n = 28; NASH n = 24; SS n = 15 | ↓ Ruminococcus, Faecalibacterium prausnitzii, and Coprococcus in NAFLD and SS vs. HC |
| [87] | n = 50; HC n = 17; NASH n = 22; SS n = 11 | ↓ Clostridium coccoides in NASH |
| [88] | n = 60; Non significant fibrosis n = 35; Significant fibrosis n = 25 | ↑ Bacteroides and Lactobacillus *, ↓ Bifidobacterium in significant fibrosis |
| [89] | n = 40; NT n = 15; HT n = 25 | ↑ Rothia ↓ Faecalicoccus, Morganella, Acetohalobium, and Phaeodactylibacter in HT |
| [90] | n = 70; NT n = 47; HT n = 23 | ↑ Acidaminococcus, Eubacterium, and Alistipes in HT |
| [91] | n = 80; NT n = 32; HT n = 48 | ↑ Ligilactobacillus salivarius, Bacteroides plebeius, and Eggerthella, ↓ Roseburia faecis, Faecalibacterium prausnitzii, Parabacteroides distasonis, Unclassified Fusobacterium, and Coprobacillus in HT |
| [92] | n = 120; HC n = 60; HT n = 60 | ↑ Klebsiella, Clostridium, Streptococcus, Parabacteroides, Eggerthella, and Salmonella, ↓ Faecalibacterium, and Roseburia in HT |
| [93] | n = 196; HC n = 41; pHT n = 56; HT n = 99 | ↑ Prevotella and Klebsiella in pHT or HT; ↑ Porphyromonas and Actinomyces in HT; ↓ Faecalibacterium, Oscillibacter, Roseburia, Subdoligranulum, Blautia, Bifidobacterium, Coprococcus, Butyrivibrio, Eggerthella, Streptococcus, and Akkermansia in pHT and HT |
| [94] | n = 900; HC n = 300; HT n = 300; CAD n = 300 | ↑ Escherichia in HT |
| [95] | n = 235; HC n = 42; NH n = 63; AH n = 104; HLD n = 26 | ↑ Blautia, Bacteroides, and Faecalibacterium in NH; ↑ Bacteroides and Faecalibacterium in HLD and HC |
| [96] | n = 502; HC n = 100; ACS n = 402 | ↑ Escherichia coli and Streptococcus, ↓ Lactobacillus * in ACS |
| [97] | n = 64; HC n = 32; CAS n = 32 | ↑ Acidaminococcus, Christensenella, and Lactobacillus *, ↓ Anaerostipes, Fusobacterium, Gemella, Parvimonas, Romboutsia, and Clostridium XVIII/XlVa/XlVb in CAS |
| [98] | n = 345; No SCA n = 201; SCA n = 144 | ↑ Escherichia and Oscillospira in SCA |
| [99] | Sweden cohort n = 25; Control 1 n = 13; Atherosclerosis 1 n = 12; China cohort n = 385; Control 2 n = 171; Atherosclerosis 2 n = 214 | ↑ Bifidobacterium adolescentis, Collinsella aerofaciens, Blautia hydrogenotrophica, and Anaerotruncus colihominis in atherosclerosis 1; ↑ Bacteroides fragilis, Streptococcus salivarius, Clostridium nexile, Ruminococcus gnavus, Ruminococcus torques, coli, Klebsiella pneumoniae, and Akkermansia muciniphila in atherosclerosis 2 |
| [100] | n = 106; Control n = 53; CAD n = 53 | ↑ Porphyromonas, Prevotella, Agathobacter, Ruminococcus gnavus, Catenibacterium, and Succiniclasticum, ↓ Anaerosporobacter, Coprococcus, Eisenbergiella, Fusocatenibacter, Eubacterium hallii, Ruminococcus gauvreauii, Fournierella, and Veillonella in CAD |
| [101] | n = 201; HC n = 40; CAD n = 161 | ↑ Actinomyces, Haemophilus, Granulicatella, Weissella, Veillonella, Streptococcus, Klebsiella, Rothia, Enterococcus (CAG17); ↓ Faecalibacterium, Roseburia, Oscilibacter (CAG4); Ruminococcus 2, Dorea, Blautia, Clostridium XVIII (CAG14); Anaerostipes, Blautia, Lactobacillus *, Fusocatenibacter, Clostridium XIVa, Gemella, Bifidobacterium, Saccharibacteria genera incertae sedis (CAG15); Roseburia, Clostridium XIVb, Parasutterella, Butyricicoccus (CAG16) in CAD |
| [102] | n = 405; HC n = 187; ACVD n = 218 | ↑ Escherichia coli, Klebsiella spp., Enterobacter aerogenes, Streptococcus spp., Ligilactobacillus salivarius, Solobacterium moorei, Atopobium parvulum, Ruminococcus gnavus, and Eggerthella lenta, ↓ Roseburia intestinalis, Faecalibacterium prausnitzii, Bacteroides spp., Prevotella copri, and Alistipes shahii in ACVD |
| Health-Related Metabolites | Disease-Related Metabolites |
|---|---|
| Fatty Acid Pathways—Metabolites and conjugates | |
| 10-Heptadecenoate (17:1n7) | (+)-Cucurbic acid |
| 2-Hydroxyhexadecanoate | 12,13-Dihydroxy-11-methoxy-9-octadecenoic acid |
| Acetate | 17-Oxo-octadecanoic acid |
| Azelaic acid | 2-Hydroxyadipate |
| Caproic acid | 2-Methyl-tridecanedioic acid |
| Caprylic acid | 3-Keto stearic acid |
| Isovalerate | 8,11,14-Eicosatrienoic acid |
| Undecanedionate | 8Z-Decen-4,6-diynoic acid |
| 9,10-Dichloro-octadecanoic acid | |
| Adrenic acid | |
| Arachidonic acid | |
| Diamino-pimelic acid | |
| Dihomo-linolenate (20:3n3 or n6) | |
| Docosahexaenoic acid | |
| Docosanedioic acid | |
| Eicosatrienoic acid | |
| Linolenic acid | |
| Amino Acid Pathways—Metabolites and derivatives | |
| Glycylvaline | Asymmetric dimethylarginine (ADMA) |
| Isoleucine | Carnosine |
| N6,N6,N6-Trimethyllysine | Cinnamoylglycine |
| N-Acetylalanine | Citrulline |
| S-Carboxymethyl-L-cysteine | Ɣ-Glutamylglutamine |
| Valine | Glycine |
| Homocitrulline | |
| Homocysteine | |
| L-Lysine | |
| N6-Carboxymethyllysine | |
| Nɑ-Acetyl-L-arginine | |
| Propionylglutamine | |
| Biliary Acid Pathways—Metabolites and derivatives | |
| Chenodeoxyglycocholate | 12-Dehydrocholic acid |
| Glycoursodeoxycholic acid | 3-Dehydrocholic acid |
| 3β-Cholic acid | |
| 6,7-Diketolithocholic acid | |
| 6-Keto-Lithocholic acid | |
| 7,12-Diketolithocholic acid | |
| 7-Dehydrocholic acid | |
| 7-Ketolithocholic acid Allocholic acid | |
| Chenodeoxycholic acid | |
| Chenodeoxycholic acid-3Gln Cholate sulfate | |
| Dehydrocholic acid | |
| Glycochenodeoxycholic acid | |
| Glycodeoxycholic acid | |
| Glycolithocholic acid | |
| Hyodeoxycholic acid | |
| Lithocholic acid | |
| Murocholic acid | |
| Nordeoxycholic acid | |
| Taurocholic acid | |
| Taurohyocholic acid | |
| Taurolithocholic acid | |
| Tauroursodeoxycholic acid | |
| αMuricholic acid | |
| βDeoxycholic acid | |
| βMuricholic acid | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres-Sánchez, A.; Ruiz-Rodríguez, A.; Ortiz, P.; Aguilera, M. Key Stratification of Microbiota Taxa and Metabolites in the Host Metabolic Health–Disease Balance. Int. J. Mol. Sci. 2023, 24, 4519. https://doi.org/10.3390/ijms24054519
Torres-Sánchez A, Ruiz-Rodríguez A, Ortiz P, Aguilera M. Key Stratification of Microbiota Taxa and Metabolites in the Host Metabolic Health–Disease Balance. International Journal of Molecular Sciences. 2023; 24(5):4519. https://doi.org/10.3390/ijms24054519
Chicago/Turabian StyleTorres-Sánchez, Alfonso, Alicia Ruiz-Rodríguez, Pilar Ortiz, and Margarita Aguilera. 2023. "Key Stratification of Microbiota Taxa and Metabolites in the Host Metabolic Health–Disease Balance" International Journal of Molecular Sciences 24, no. 5: 4519. https://doi.org/10.3390/ijms24054519