
View from the Biological Property: Insight into the Functional Diversity and Complexity of the Gut Mucus
 ,
,
Abstract
:Highlights
- Mucus, as the most widely distributed biofilm and vital protective barrier on the surface of mucous membranes throughout the body, fulfills a number of critical activities in the maintenance of cellular and organismal homeostasis.
- Morphologic and biochemical evidence corroborate that the mucin family is composed of the transmembrane and gel-forming mucins, which have diverse functional modules.
- Goblet cells are responsible for synthesis, storage, and secreting, which are implicated in either beneficial or detrimental factors.
- Excessive or insufficient mucus secretion as well as phenotypic alternation may be associated with gastrointestinal disorders, offering potential therapeutic targets for prevention in clinical practice.
Abstract
1. Introduction
2. Mucus Biological Properties
2.1. General Biological Characteristics of Mucus
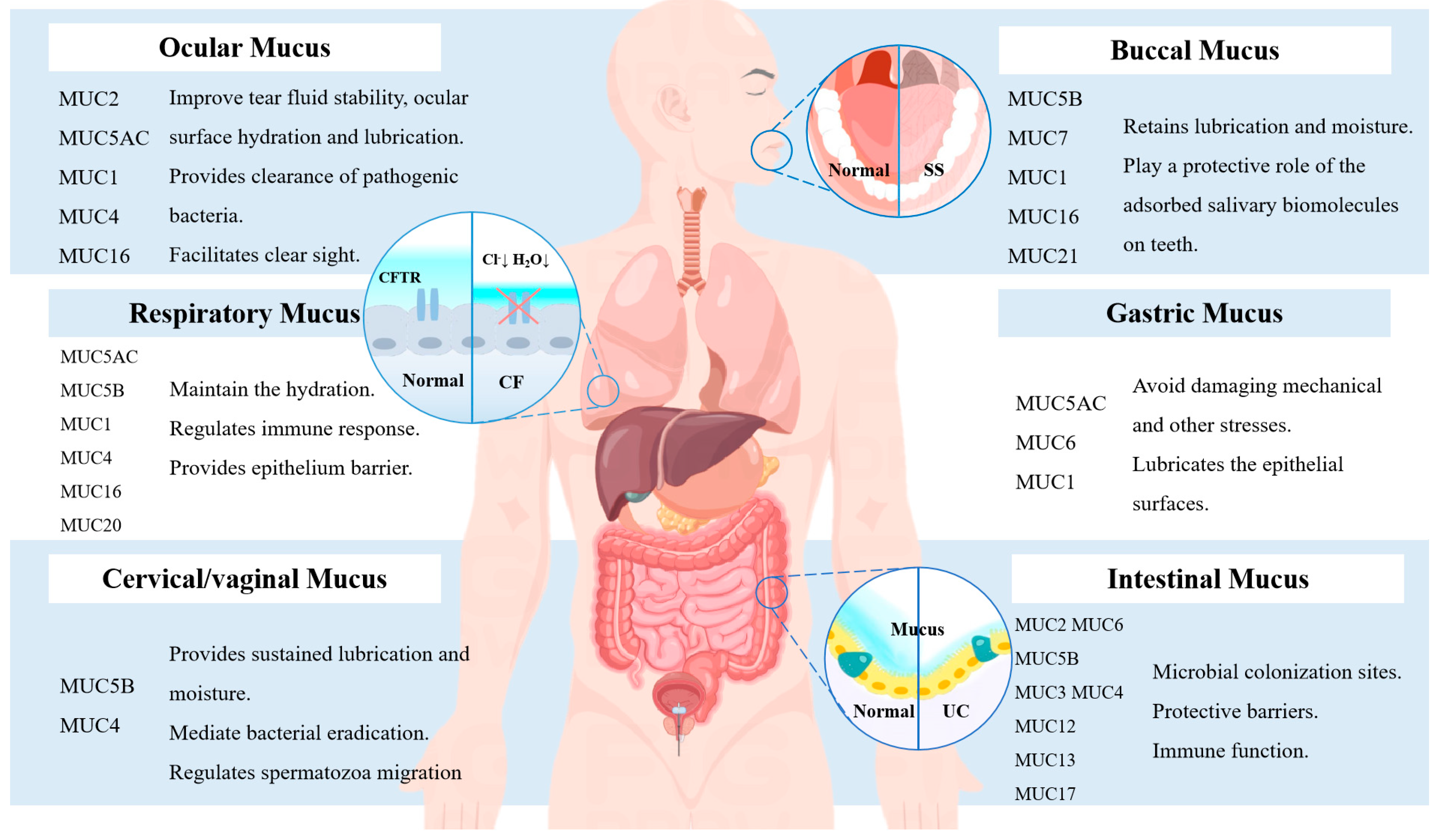
2.2. Mucus Serves as a Crucial Entryway to the Body’s Barrier
2.2.1. Physical Barrier Function of Mucus
2.2.2. Immune Barrier Function of Mucus
3. Mucin Glycoproteins as the Main Component of Mucus
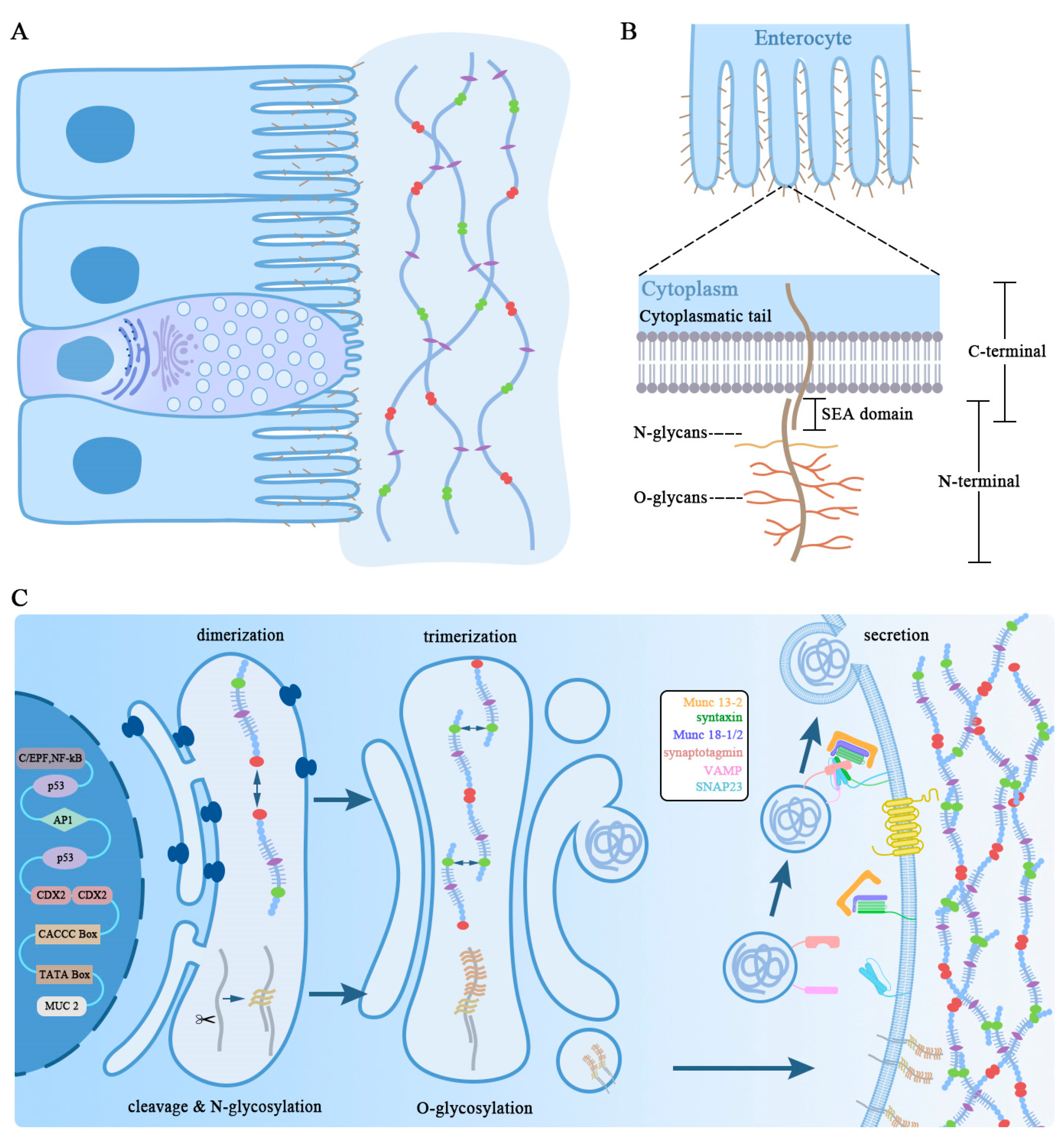
3.1. Synthesis and Secretion of Mucin in Goblet Cell
3.2. Transmembrane Mucins
3.3. Secreted Mucins

{kind=link}
{kind=link}
{kind=link}
| Mucin Type | Mucin/Cytogenetic Band | Cell Type Expression | Protein Domains | Number of Amino Acids | Number (Estimated Length) of Mucin Domains | Section of Gastrointestinal Tract | Functions | Refs. | |
|---|---|---|---|---|---|---|---|---|---|
| Secreted mucin | Gel mucin | MUC2/11p15.5 | Goblet cells Paneth cells | 4 VWD, 2 CysD, 1 CK | ~5200 | 2 (~550 nm) | Small intestine, large intestine | Protection, lubrication, entrapment | [43,44,45,46] |
| MUC5AC/11p15.5 | Mucous cells | 4 VWD, 11 CysD, 1 CK | >5050 | 11 (>350 nm) | Stomach | Protection, lubrication, entrapment | [47,48] | ||
| MUC5B/11p15.5 | Mucous cells Goblet cells | 4 VWD, 7 CysD, 1 CK | ~5700 | 7 (~550 nm) | Mouth, large intestine | Protection, lubrication, entrapment | [49,50] | ||
| MUC6/11p15.5 | Mucous cells | 1 VWD, 1 CK | ~2400 | 1 (~250 nm) | Stomach, small intestine | Protection, lubrication, entrapment | [51,52,53] | ||
| MUC19/12q12 | Mucous cells | 1 VWC | >7000 | 1 | Salivary gland, testis | Protection, lubrication | [54,55] | ||
| Small soluble mucin | MUC7/4q13.3 | Mucous cells | None | 377 | 1 (~230 nm) | Mouth | Protection | [56,57,58] | |
| MUC8/12q24.33 | Epithelial cells | None | 2699 | None | Airway | Protection | [59] | ||
| MUC9/1p13.2 | Epithelial cells | None | 654 | 1 | Oviduct | Fertilization related | [60,61,62] | ||
| Transmembrane mucin | MUC1/1q22 | Epithelial cells | 1 SEA | ~1250 | 1 (~200 nm) | Mouth, stomach, small intestine, large intestine | Signaling, protection | [63,64,65] | |
| MUC3II/7q22 | Enterocytes | 1 SEA | >2550 | 1 (>350 nm) | Small intestine, large intestine | Apical surface protection | [66,67,68] | ||
| MUC4/3q29 | Epithelial cells goblet cells | 1 NIDO, 1 AMOP, 1 VWD | ~5300 | 1 (~800 nm) | Small intestine, large intestine | Signaling, protection | [43,69,70] | ||
| MUC12/7q22.1 | Enterocytes | 1 SEA | ~5500 | 1 (~1000 nm) | Small intestine, large intestine | Apical surface protection | [71] | ||
| MUC13/3q21.2 | Enterocytes | 1 SEA | 512 | 1 (~30 nm) | Small intestine, large intestine | Apical surface protection | [72] | ||
| MUC15/11p14.3 | Gland cells | None | ~334 | 1 | Epididymis, thyroid | Antimicrobial activity | [73] | ||
| MUC16/19p13.2 | Epithelial cells | 33 SEA | ~22,000 | 1 (~2400 nm) | Mouth | Apical surface protection | [74] | ||
| MUC177q22.1 | Enterocytes | 1 SEA | ~4500 | 1 (~800 nm) | Small intestine, large intestine | Apical surface protection | [75,76] | ||
| MUC20/3q29 | Epithelial cells | None | ~709 | 2 | Esophagus, lung, stomach, kidney | Signaling, protection | [77,78] | ||
| MUC21/6p21.33 | Epithelial cells | None | 566 | Mouth, stomach, eyes | Signaling, protection | [79,80] | |||
| MUC22/6p21.33 | Epithelial cells | None | ~1773 | 1 | Esophagus, vagina, lung | Protection | [81,82] | ||
| Acidic Mucin | Neutral Mucin | Refs. | |
|---|---|---|---|
| Main amino acids | Proline, threonine, glycine | Serine, aspartate, alanine | [83] |
| Main glycosylation | Sialic acid, N-acetylgalactosamine | Fucose, galactose, N-acetylglucosamine | [83] |
| Location | Stomach: | Surface epithelium, foveolar cells, most of the mucous neck cells | [84,85] |
| Small intestine: | Glycocalyx of the brush border, goblet cells of both villi and crypts, especially in distal ileum | ||
| Large intestine: | All brush border and goblet cells | ||
| Secernent | Bacterial colonization: Bifidobacterium dentium, Helicobacter pylori Cancer: gastric cancer Drugs, cytokines and chemicals: Moringa oleifera leaf powder, keratinocyte growth factor, trefoil peptide, dietary resistant starch type 3 | Bacterial colonization: Salmonella typhimurium Drugs: butyrate, methotrexate Others: Nippostrongylus brasiliensis infection | [86,87,88,89,90,91,92,93,94,95,96,97,98,99] |
| Antisecretory | Drugs, and chemicals: aspirin, sesame oil | Bacterial colonization: Helicobacter pylori Cancer: gastric cancer Others: food restriction, microgravity, aging, vagotomy | [98,99,100,101,102,103,104,105] |
4. Regulation of Mucin Function
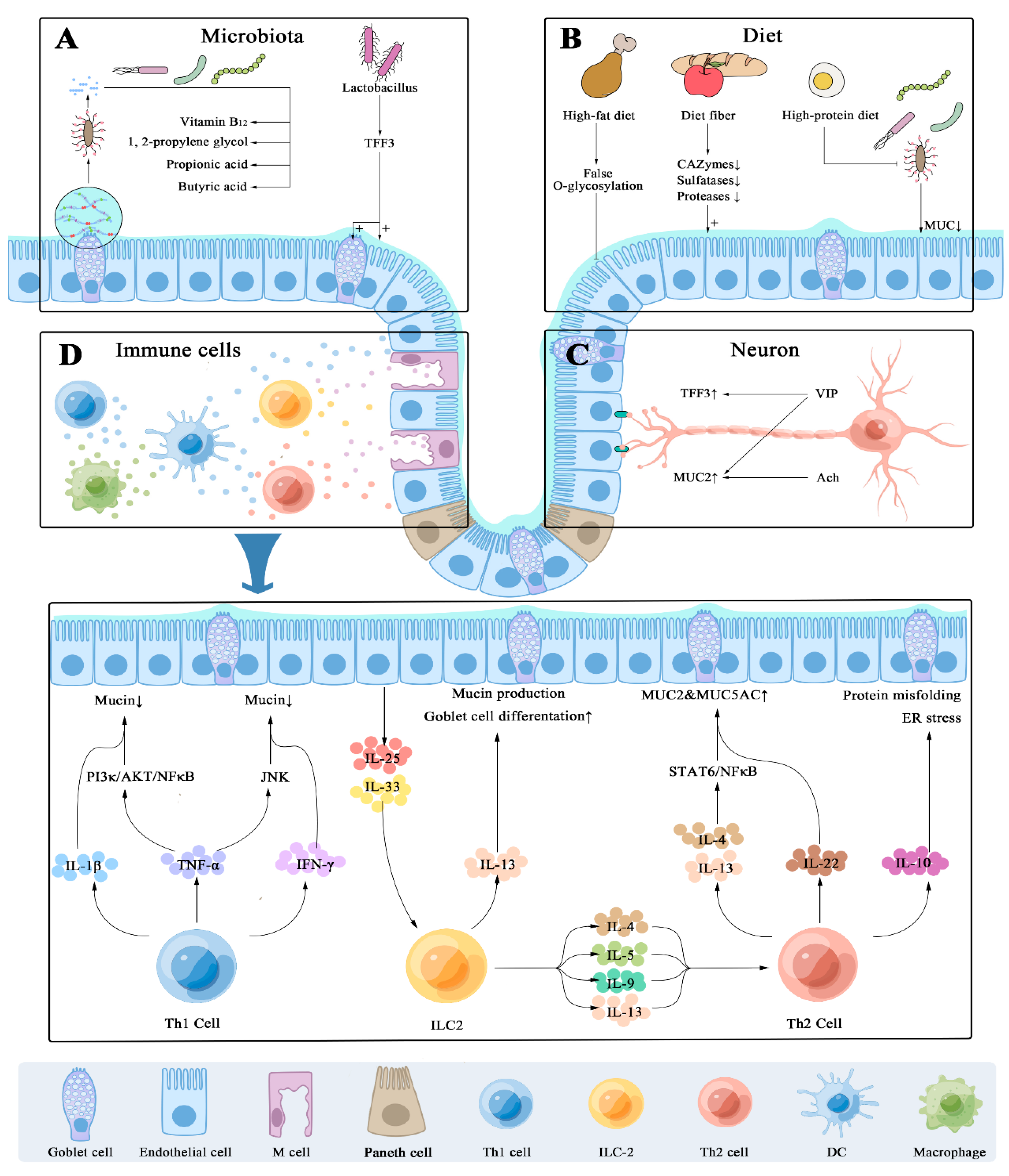
4.1. Host’s Immune-Dependent Regulation
4.2. Enteric Nervous System
4.3. Diet Ingredients Impact on Mucin Secretion
4.4. Microbial Colonization
5. Mucus Abnormalities and Disease
5.1. Sjogren’s Syndrome and Mucin Deficiency
5.2. Bidirectional Relation between Mucin and Cystic Fibrosis
5.3. Helicobacter Pylori and Mucin
5.4. IBS and IBD
5.5. Mucin in Cancer
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| GI | gastrointestinal |
| KO | knock out |
| CAII | carbonic anhydrase |
| CMC | carboxymethylcellulose |
| Cdx2 | caudal-related homeobox-2 |
| CR | Citrobacter rodentium |
| CPs | cysteine protease |
| CF | cystic fibrosis |
| CFTR | cystic fibrosis transmembrane conductance regulator |
| DAG | diacylglycerol |
| ER | endoplasmic reticulum |
| EPCR | endothelial protein C receptor |
| ENS | enteric nervous system |
| EGF | epidermal growth factor |
| EMT | epithelial-mesenchymal transition |
| GF | germ-free |
| GC | goblet cell |
| Gfi1 | growth factor independence 1 |
| HLA | human leukocyte antigen |
| PC-Ac | IgA+-secreting plasma cells |
| IgA | immunoglobulin A |
| ILCs | innate lymphoid cells |
| IP3 | inositol triphosphate |
| ICAM1 | intercellular cell adhesion molecule-1 |
| IFN | interferon |
| IL-1β | interleukin-1β |
| ILD | interstitial lung disease |
| IBS | irritable bowel syndrome |
| JAK-STAT | Janus kinase-signal transducer and activator of transcription |
| Klf4 | Krüppel-like transcription factor 4 |
| MUNC | mammalian ortholog of Caenorhabditis elegans uncoordinated |
| MAPK | mitogen-activated protein kinase |
| Muc2−/− | MUC2-deficient |
| MUC2 | Mucin 2 |
| MALT | mucosa-associated lymphoid tissue |
| MARCKS | myristoylated alanine-rich C-kinase substrate |
| NHE1 | Na+/H+ exchanger |
| NeuNAc | N-acetyl neuraminic |
| GalNAc | N-acetylgalactosamine |
| GlcNAc | N-acetylglucosamine |
| NLRP6 | NOD-like receptor family pyrin domain containing 6 |
| NF-κB | nuclear factor κB |
| OVA | ovalbumin |
| PI3K | phosphatidylinositol 3-kinase |
| PigR | polymeric Ig receptor |
| PAR2 | protease-activated receptor 2 |
| STAT | signal transducer and activator of transcription |
| SS | Sjogren’s syndrome |
| SNARE | soluble N-ethylmaliemide sensitive factor attachment receptor |
| SEA | sperm protein, enterokinase, and agrin |
| TR | tandem-repeat |
| TLR4 | Toll-like receptor 4 |
| TM | transmembrane mucins |
| Tff3 | trefoil factor 3 |
| TAAs | tumor-associated antigens |
| TNF | tumor necrosis factor |
| VEC | vascular endothelial cell |
| VIP | vasoactive intestinal peptide |
| VAMP | vesicle-associated membrane protein |
| vWF | von Willebrand factor |
| vWD | vWF-like D domain |
| Wnt | wingless-type MMTV integration site family member |
References
- Cheng, L.K.; O’Grady, G.; Du, P.; Egbuji, J.U.; Windsor, J.A.; Pullan, A.J. Gastrointestinal system. Wiley Interdiscip. Rev. Syst. Biol. Med. 2010, 2, 65–79. [Google Scholar] [CrossRef] [Green Version]
- McCracken, V.J.; Lorenz, R.G. The gastrointestinal ecosystem: A precarious alliance among epithelium, immunity and microbiota. Cell. Microbiol. 2001, 3, 1–11. [Google Scholar] [CrossRef]
- Okumura, R.; Takeda, K. Maintenance of intestinal homeostasis by mucosal barriers. Inflamm. Regen. 2018, 38, 5. [Google Scholar] [CrossRef]
- Michaudel, C.; Sokol, H. The Gut Microbiota at the Service of Immunometabolism. Cell Metab. 2020, 32, 514–523. [Google Scholar] [CrossRef]
- de Vos, W.M.; Tilg, H.; Van Hul, M.; Cani, P.D. Gut microbiome and health: Mechanistic insights. Gut 2022, 71, 1020–1032. [Google Scholar] [CrossRef]
- Meddings, J. The significance of the gut barrier in disease. Gut 2008, 57, 438–440. [Google Scholar] [CrossRef] [PubMed]
- Ashida, H.; Ogawa, M.; Kim, M.; Mimuro, H.; Sasakawa, C. Bacteria and host interactions in the gut epithelial barrier. Nat. Chem. Biol. 2011, 8, 36–45. [Google Scholar] [CrossRef] [PubMed]
- McGuckin, M.A.; Linden, S.K.; Sutton, P.; Florin, T.H. Mucin dynamics and enteric pathogens. Nat. Rev. Microbiol. 2011, 9, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Paone, P.; Cani, P.D. Mucus barrier, mucins and gut microbiota: The expected slimy partners? Gut 2020, 69, 2232–2243. [Google Scholar] [CrossRef]
- Bansil, R.; Turner, B.S. The biology of mucus: Composition, synthesis and organization. Adv. Drug Deliv. Rev. 2018, 124, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Atuma, C.; Strugala, V.; Allen, A.; Holm, L. The adherent gastrointestinal mucus gel layer: Thickness and physical state in vivo. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 280, G922–G929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, M.E.; Hansson, G.C. Immunological aspects of intestinal mucus and mucins. Nat. Rev. Immunol. 2016, 16, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Capaldo, C.T.; Powell, D.N.; Kalman, D. Layered defense: How mucus and tight junctions seal the intestinal barrier. J. Mol. Med. 2017, 95, 927–934. [Google Scholar] [CrossRef] [Green Version]
- Cornick, S.; Tawiah, A.; Chadee, K. Roles and regulation of the mucus barrier in the gut. Tissue Barriers 2015, 3, e982426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Wu, Y.; Li, J.; Bao, Y.; Guo, Y.; Yang, W. The Dynamic Changes of Gut Microbiota in Muc2 Deficient Mice. Int. J. Mol. Sci. 2018, 19, 2809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor Nordgard, C.; Draget, K.I. Dynamic responses in small intestinal mucus: Relevance for the maintenance of an intact barrier. Eur. J. Pharm. Biopharm. 2015, 95, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Hansson, G.C. Mucus and mucins in diseases of the intestinal and respiratory tracts. J. Intern. Med. 2019, 285, 479–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roos, A.; Bouwman, L.H.; van Gijlswijk-Janssen, D.J.; Faber-Krol, M.C.; Stahl, G.L.; Daha, M.R. Human IgA activates the complement system via the mannan-binding lectin pathway. J. Immunol 2001, 167, 2861–2868. [Google Scholar] [CrossRef] [Green Version]
- Daele, J.; Zicot, A.F. Humoral immunodeficiency in recurrent upper respiratory tract infections. Some basic, clinical and therapeutic features. Acta Oto-Rhino-Laryngol. Belg. 2000, 54, 373–390. [Google Scholar]
- Ludvigsson, J.F.; Neovius, M.; Hammarstrom, L. Risk of Infections Among 2100 Individuals with IgA Deficiency: A Nationwide Cohort Study. J. Clin. Immunol. 2016, 36, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Natvig, I.B.; Johansen, F.E.; Nordeng, T.W.; Haraldsen, G.; Brandtzaeg, P. Mechanism for enhanced external transfer of dimeric IgA over pentameric IgM: Studies of diffusion, binding to the human polymeric Ig receptor, and epithelial transcytosis. J. Immunol. 1997, 159, 4330–4340. [Google Scholar] [CrossRef] [PubMed]
- Ammann, A.J.; Hong, R. Selective IgA deficiency: Presentation of 30 cases and a review of the literature. Medicine 1971, 50, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Faderl, M.; Noti, M.; Corazza, N.; Mueller, C. Keeping bugs in check: The mucus layer as a critical component in maintaining intestinal homeostasis. IUBMB Life 2015, 67, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Shan, M.; Gentile, M.; Yeiser, J.R.; Walland, A.C.; Bornstein, V.U.; Chen, K.; He, B.; Cassis, L.; Bigas, A.; Cols, M.; et al. Mucus enhances gut homeostasis and oral tolerance by delivering immunoregulatory signals. Science 2013, 342, 447–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Yu, M. Role of Goblet Cells in Intestinal Barrier and Mucosal Immunity. J. Inflamm. Res. 2021, 14, 3171–3183. [Google Scholar] [CrossRef]
- Kwon, O.J.; Valdez, J.M.; Zhang, L.; Zhang, B.; Wei, X.; Su, Q.; Ittmann, M.M.; Creighton, C.J.; Xin, L. Increased Notch signalling inhibits anoikis and stimulates proliferation of prostate luminal epithelial cells. Nat. Commun. 2014, 5, 4416. [Google Scholar] [CrossRef] [Green Version]
- van der Flier, L.G.; Clevers, H. Stem cells, self-renewal, and differentiation in the intestinal epithelium. Annu. Rev. Physiol. 2009, 71, 241–260. [Google Scholar] [CrossRef]
- Yang, Q.; Bermingham, N.A.; Finegold, M.J.; Zoghbi, H.Y. Requirement of Math1 for secretory cell lineage commitment in the mouse intestine. Science 2001, 294, 2155–2158. [Google Scholar] [CrossRef]
- Shroyer, N.F.; Wallis, D.; Venken, K.J.; Bellen, H.J.; Zoghbi, H.Y. Gfi1 functions downstream of Math1 to control intestinal secretory cell subtype allocation and differentiation. Genes Dev. 2005, 19, 2412–2417. [Google Scholar] [CrossRef] [Green Version]
- Katz, J.P.; Perreault, N.; Goldstein, B.G.; Lee, C.S.; Labosky, P.A.; Yang, V.W.; Kaestner, K.H. The zinc-finger transcription factor Klf4 is required for terminal differentiation of goblet cells in the colon. Development 2002, 129, 2619–2628. [Google Scholar] [CrossRef]
- Akiba, Y.; Guth, P.H.; Engel, E.; Nastaskin, I.; Kaunitz, J.D. Dynamic regulation of mucus gel thickness in rat duodenum. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, G437–G447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McShane, A.; Bath, J.; Jaramillo, A.M.; Ridley, C.; Walsh, A.A.; Evans, C.M.; Thornton, D.J.; Ribbeck, K. Mucus. Curr. Biol. 2021, 31, R938–R945. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.M.; Koo, J.S. Airway mucus: The good, the bad, the sticky. Pharmacol. Ther. 2009, 121, 332–348. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo, A.M.; Azzegagh, Z.; Tuvim, M.J.; Dickey, B.F. Airway Mucin Secretion. Ann. Am. Thorac Soc. 2018, 15, S164–S170. [Google Scholar] [CrossRef]
- Corfield, A.P. Mucins: A biologically relevant glycan barrier in mucosal protection. Biochim. Biophys. Acta 2015, 1850, 236–252. [Google Scholar] [CrossRef]
- Johansson, M.E.; Sjovall, H.; Hansson, G.C. The gastrointestinal mucus system in health and disease. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 352–361. [Google Scholar] [CrossRef] [Green Version]
- Pelaseyed, T.; Hansson, G.C. Membrane mucins of the intestine at a glance. J. Cell Sci. 2020, 133, jcs240929. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, A.; Gu, Y.; Lin, X.; Peng, J.; Major, P.; Tang, D. MUCIN 1 in Prostate Cancer. In Prostate Cancer; Bott, S.R.J., Ng, K.L., Eds.; Exon Publications: Brisbane, Australia, 2021. [Google Scholar] [CrossRef]
- Striefler, J.K.; Riess, H.; Lohneis, P.; Bischoff, S.; Kurreck, A.; Modest, D.P.; Bahra, M.; Oettle, H.; Sinn, M.; Blaker, H.; et al. Mucin-1 Protein Is a Prognostic Marker for Pancreatic Ductal Adenocarcinoma: Results From the CONKO-001 Study. Front. Oncol. 2021, 11, 670396. [Google Scholar] [CrossRef]
- Utispan, K.; Koontongkaew, S. Mucin 1 regulates the hypoxia response in head and neck cancer cells. J. Pharm. Sci. 2021, 147, 331–339. [Google Scholar] [CrossRef]
- Johansson, M.E.; Gustafsson, J.K.; Holmen-Larsson, J.; Jabbar, K.S.; Xia, L.; Xu, H.; Ghishan, F.K.; Carvalho, F.A.; Gewirtz, A.T.; Sjovall, H.; et al. Bacteria penetrate the normally impenetrable inner colon mucus layer in both murine colitis models and patients with ulcerative colitis. Gut 2014, 63, 281–291. [Google Scholar] [CrossRef]
- Konig, J.; Wells, J.; Cani, P.D.; Garcia-Rodenas, C.L.; MacDonald, T.; Mercenier, A.; Whyte, J.; Troost, F.; Brummer, R.J. Human Intestinal Barrier Function in Health and Disease. Clin. Transl. Gastroenterol. 2016, 7, e196. [Google Scholar] [CrossRef] [PubMed]
- Audie, J.P.; Janin, A.; Porchet, N.; Copin, M.C.; Gosselin, B.; Aubert, J.P. Expression of Human Mucin Genes in Respiratory, Digestive, and Reproductive Tracts Ascertained by in-Situ Hybridization. J. Histochem. Cytochem. 1993, 41, 1479–1485. [Google Scholar] [CrossRef] [PubMed]
- Weiss, A.A.; Babyatsky, M.W.; Ogata, S.; Chen, A.; Itzkowitz, S.H. Expression of MUC2 and MUC3 mRNA in human normal, malignant, and inflammatory intestinal tissues. J. Histochem. Cytochem. 1996, 44, 1161–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, M.E.; Phillipson, M.; Petersson, J.; Velcich, A.; Holm, L.; Hansson, G.C. The inner of the two Muc2 mucin-dependent mucus layers in colon is devoid of bacteria. Proc. Natl. Acad. Sci. USA 2008, 105, 15064–15069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gum, J.R., Jr.; Hicks, J.W.; Toribara, N.W.; Rothe, E.M.; Lagace, R.E.; Kim, Y.S. The human MUC2 intestinal mucin has cysteine-rich subdomains located both upstream and downstream of its central repetitive region. J. Biol. Chem. 1992, 267, 21375–21383. [Google Scholar] [CrossRef]
- Allen, A.; Flemstrom, G. Gastroduodenal mucus bicarbonate barrier: Protection against acid and pepsin. Am. J. Physiol. Cell Physiol. 2005, 288, C1–C19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meezaman, D.; Charles, P.; Daskal, E.; Polymeropoulos, M.H.; Martin, B.M.; Rose, M.C. Cloning and analysis of cDNA encoding a major airway glycoprotein, human tracheobronchial mucin (MUC5). J. Biol. Chem. 1994, 269, 12932–12939. [Google Scholar] [CrossRef]
- Troxler, R.F.; Offner, G.D.; Zhang, F.; Iontcheva, I.; Oppenheim, F.G. Molecular cloning of a novel high molecular weight mucin (MG1) from human sublingual gland. Biochem. Biophys. Res. Commun. 1995, 217, 1112–1119. [Google Scholar] [CrossRef]
- Keates, A.C.; Nunes, D.P.; Afdhal, N.H.; Troxler, R.F.; Offner, G.D. Molecular cloning of a major human gall bladder mucin: Complete C-terminal sequence and genomic organization of MUC5B. Biochem. J. 1997, 324, 295–303. [Google Scholar] [CrossRef] [Green Version]
- Ho, S.B.; Shekels, L.L.; Toribara, N.W.; Kim, Y.S.; Lyftogt, C.; Cherwitz, D.L.; Niehans, G.A. Mucin gene expression in normal, preneoplastic, and neoplastic human gastric epithelium. Cancer Res. 1995, 55, 2681–2690. [Google Scholar]
- Toribara, N.W.; Roberton, A.M.; Ho, S.B.; Kuo, W.L.; Gum, E.; Hicks, J.W.; Gum, J.R., Jr.; Byrd, J.C.; Siddiki, B.; Kim, Y.S. Human gastric mucin. Identification of a unique species by expression cloning. J. Biol. Chem. 1993, 268, 5879–5885. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, K.; Byrne, C.; Kim, Y.S.; Gum, J.R.; Swallow, D.M.; Toribara, N.W. The complete genomic organization of the human MUC6 and MUC2 mucin genes. Genomics 2004, 83, 936–939. [Google Scholar] [CrossRef] [PubMed]
- Culp, D.J.; Robinson, B.; Cash, M.N.; Bhattacharyya, I.; Stewart, C.; Cuadra-Saenz, G. Salivary mucin 19 glycoproteins: Innate immune functions in Streptococcus mutans-induced caries in mice and evidence for expression in human saliva. J. Biol. Chem. 2015, 290, 2993–3008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Zhao, Y.H.; Kalaslavadi, T.B.; Hamati, E.; Nehrke, K.; Le, A.D.; Ann, D.K.; Wu, R. Genome-wide search and identification of a novel gel-forming mucin MUC19/Muc19 in glandular tissues. Am. J. Respir. Cell Mol. Biol. 2004, 30, 155–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langbein, L.; Heid, H.W.; Moll, I.; Franke, W.W. Molecular characterization of the body site-specific human epidermal cytokeratin 9: cDNA cloning, amino acid sequence, and tissue specificity of gene expression. Differentiation 1994, 55, 164. [Google Scholar] [CrossRef] [PubMed]
- Situ, H.; Wei, G.; Smith, C.J.; Mashhoon, S.; Bobek, L.A. Human salivary MUC7 mucin peptides: Effect of size, charge and cysteine residues on antifungal activity. Biochem. J. 2003, 375, 175–182. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, P.A.; Mandel, U.; Therkildsen, M.H.; Clausen, H. Differential expression of human high-molecular-weight salivary mucin (MG1) and low-molecular-weight salivary mucin (MG2). J. Dent. Res. 1996, 75, 1820–1826. [Google Scholar] [CrossRef]
- Song, S.Y.; Jung, E.C.; Bae, C.H.; Choi, Y.S.; Kim, Y.D. Visfatin induces MUC8 and MUC5B expression via p38 MAPK/ROS/NF-kappaB in human airway epithelial cells. J. Biomed. Sci. 2014, 21, 49. [Google Scholar] [CrossRef] [Green Version]
- Slayden, O.D.; Friason, F.K.E.; Bond, K.R.; Mishler, E.C. Hormonal regulation of oviductal glycoprotein 1 (OVGP1; MUC9) in the rhesus macaque cervix. J. Med. Primatol. 2018, 47, 362–370. [Google Scholar] [CrossRef]
- Choudhary, S.; Janjanam, J.; Kumar, S.; Kaushik, J.K.; Mohanty, A.K. Structural and functional characterization of buffalo oviduct-specific glycoprotein (OVGP1) expressed during estrous cycle. Biosci. Rep. 2019, 39, 12. [Google Scholar] [CrossRef] [Green Version]
- Lagow, E.; DeSouza, M.M.; Carson, D.D. Mammalian reproductive tract mucins. Hum. Reprod. Update 1999, 5, 280–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linden, S.K.; Florin, T.H.; McGuckin, M.A. Mucin dynamics in intestinal bacterial infection. PLoS ONE 2008, 3, e3952. [Google Scholar] [CrossRef] [PubMed]
- Gendler, S.J. MUC1, the renaissance molecule. J. Mammary Gland Biol. Neoplasia 2001, 6, 339–353. [Google Scholar] [CrossRef] [PubMed]
- Lan, M.S.; Batra, S.K.; Qi, W.N.; Metzgar, R.S.; Hollingsworth, M.A. Cloning and sequencing of a human pancreatic tumor mucin cDNA. J. Biol. Chem. 1990, 265, 15294–15299. [Google Scholar] [CrossRef] [PubMed]
- Gum, J.R.; Hicks, J.W.; Swallow, D.M.; Lagace, R.L.; Byrd, J.C.; Lamport, D.T.; Siddiki, B.; Kim, Y.S. Molecular cloning of cDNAs derived from a novel human intestinal mucin gene. Biochem. Biophys. Res. Commun. 1990, 171, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.K.; Dohrman, A.F.; Basbaum, C.B.; Ho, S.B.; Tsuda, T.; Toribara, N.W.; Gum, J.R.; Kim, Y.S. Localization of mucin (MUC2 and MUC3) messenger RNA and peptide expression in human normal intestine and colon cancer. Gastroenterology 1994, 107, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Crawley, S.C.; Gum, J.R., Jr.; Hicks, J.W.; Pratt, W.S.; Aubert, J.P.; Swallow, D.M.; Kim, Y.S. Genomic organization and structure of the 3’ region of human MUC3: Alternative splicing predicts membrane-bound and soluble forms of the mucin. Biochem. Biophys. Res. Commun. 1999, 263, 728–736. [Google Scholar] [CrossRef]
- Rong, M.; Rossi, E.A.; Zhang, J.; McNeer, R.R.; van den Brande, J.M.; Yasin, M.; Weed, D.T.; Carothers Carraway, C.A.; Thompson, J.F.; Carraway, K.L. Expression and localization of Muc4/sialomucin complex (SMC) in the adult and developing rat intestine: Implications for Muc4/SMC function. J. Cell. Physiol. 2005, 202, 275–284. [Google Scholar] [CrossRef]
- Moniaux, N.; Nollet, S.; Porchet, N.; Degand, P.; Laine, A.; Aubert, J.P. Complete sequence of the human mucin MUC4: A putative cell membrane-associated mucin. Biochem. J. 1999, 338, 325–333. [Google Scholar] [CrossRef]
- Williams, S.J.; McGuckin, M.A.; Gotley, D.C.; Eyre, H.J.; Sutherland, G.R.; Antalis, T.M. Two novel mucin genes down-regulated in colorectal cancer identified by differential display. Cancer Res. 1999, 59, 4083–4089. [Google Scholar]
- Williams, S.J.; Wreschner, D.H.; Tran, M.; Eyre, H.J.; Sutherland, G.R.; McGuckin, M.A. Muc13, a novel human cell surface mucin expressed by epithelial and hemopoietic cells. J. Biol. Chem. 2001, 276, 18327–18336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, C.; Thi Thao Tran, N.; Van Ngu, T.; Park, S.W.; Song, M.S.; Kim, S.H.; Bae, Y.U.; Ayudthaya, P.D.N.; Munir, J.; Kim, E.; et al. Promotion of tumor progression and cancer stemness by MUC15 in thyroid cancer via the GPCR/ERK and integrin-FAK signaling pathways. Oncogenesis 2018, 7, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, B.W.; Lloyd, K.O. Molecular cloning of the CA125 ovarian cancer antigen: Identification as a new mucin, MUC16. J. Biol. Chem. 2001, 276, 27371–27375. [Google Scholar] [CrossRef] [Green Version]
- Gum, J.R., Jr.; Crawley, S.C.; Hicks, J.W.; Szymkowski, D.E.; Kim, Y.S. MUC17, a novel membrane-tethered mucin. Biochem. Biophys. Res. Commun. 2002, 291, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Senapati, S.; Ho, S.B.; Sharma, P.; Das, S.; Chakraborty, S.; Kaur, S.; Niehans, G.; Batra, S.K. Expression of intestinal MUC17 membrane-bound mucin in inflammatory and neoplastic diseases of the colon. J. Clin. Pathol. 2010, 63, 702–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, L.; Yonemura, A.; Yasuda-Yoshihara, N.; Umemoto, T.; Zhang, J.; Yasuda, T.; Uchihara, T.; Akiyama, T.; Kitamura, F.; Yamashita, K.; et al. Intracellular MUC20 variant 2 maintains mitochondrial calcium homeostasis and enhances drug resistance in gastric cancer. Gastric Cancer 2022, 25, 542–557. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Panjwani, N.; Wang, C.; Sun, L.; Strug, L.J. A flexible summary statistics-based colocalization method with application to the mucin cystic fibrosis lung disease modifier locus. Am. J. Hum. Genet. 2022, 109, 253–269. [Google Scholar] [CrossRef]
- Dai, W.; Liu, J.; Liu, B.; Li, Q.; Sang, Q.; Li, Y.Y. Systematical Analysis of the Cancer Genome Atlas Database Reveals EMCN/MUC15 Combination as a Prognostic Signature for Gastric Cancer. Front. Mol. Biosci. 2020, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Bao, X.; Wiehe, R.; Dommisch, H.; Schaefer, A.S. Entamoeba gingivalis Causes Oral Inflammation and Tissue Destruction. J. Dent. Res. 2020, 99, 561–567. [Google Scholar] [CrossRef]
- Norman, P.J.; Norberg, S.J.; Guethlein, L.A.; Nemat-Gorgani, N.; Royce, T.; Wroblewski, E.E.; Dunn, T.; Mann, T.; Alicata, C.; Hollenbach, J.A.; et al. Sequences of 95 human MHC haplotypes reveal extreme coding variation in genes other than highly polymorphic HLA class I and II. Genome Res. 2017, 27, 813–823. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Carrasco, R.; Argueso, P.; Fini, M.E. Membrane-associated mucins of the human ocular surface in health and disease. Ocul. Surf. 2021, 21, 313–330. [Google Scholar] [CrossRef] [PubMed]
- Wesley, A.; Mantle, M.; Man, D.; Qureshi, R.; Forstner, G.; Forstner, J. Neutral and acidic species of human intestinal mucin. Evidence for different core peptides. J. Biol. Chem. 1985, 260, 7955–7959. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, L.C. Analysis of the Distribution of Mucins in Adult Human Gastric Mucosa and Its Functional Significance. J. Clin. Diagn. Res. 2016, 10, AC01–AC04. [Google Scholar] [CrossRef]
- Sheahan, D.G.; Jervis, H.R. Comparative histochemistry of gastrointestinal mucosubstances. Am. J. Anat. 1976, 146, 103–131. [Google Scholar] [CrossRef] [PubMed]
- Enss, M.L.; Grosse-Siestrup, H.; Riedesel, H. Acidification of the colonic mucins following polyvalent colonization of the germ-free rat. Zent. Vet. B 1992, 39, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Ulaganathan, M.; Familari, M.; Yeomans, N.D.; Giraud, A.S.; Cook, G.A. Spatio-temporal expression of trefoil peptide following severe gastric ulceration in the rat implicates it in late-stage repair processes. J. Gastroenterol. Hepatol. 2001, 16, 506–512. [Google Scholar] [CrossRef]
- Bauer-Marinovic, M.; Florian, S.; Müller-Schmehl, K.; Glatt, H.; Jacobasch, G. Dietary resistant starch type 3 prevents tumor induction by 1,2-dimethylhydrazine and alters proliferation, apoptosis and dedifferentiation in rat colon. Carcinogenesis 2006, 27, 1849–1859. [Google Scholar] [CrossRef] [Green Version]
- Iijima, K.; Ichikawa, T.; Okada, S.; Ogawa, M.; Koike, T.; Ohara, S.; Shimosegawa, T. Rebamipide, a cytoprotective drug, increases gastric mucus secretion in human: Evaluations with endoscopic gastrin test. Dig. Dis. Sci. 2009, 54, 1500–1507. [Google Scholar] [CrossRef] [Green Version]
- Egger, B.; Inglin, R.; Zeeh, J.; Dirsch, O.; Huang, Y.; Büchler, M.W. Insulin-like growth factor I and truncated keratinocyte growth factor accelerate healing of left-sided colonic anastomoses. Br. J. Surg. 2001, 88, 90–98. [Google Scholar] [CrossRef]
- Khan, I.; Zaneb, H.; Masood, S.; Yousaf, M.S.; Rehman, H.F.; Rehman, H. Effect of Moringa oleifera leaf powder supplementation on growth performance and intestinal morphology in broiler chickens. J. Anim. Physiol. Anim. Nutr. 2017, 101 (Suppl. 1), 114–121. [Google Scholar] [CrossRef] [Green Version]
- Tse, S.K.; Chadee, K. Biochemical characterization of rat colonic mucins secreted in response to Entamoeba histolytica. Infect. Immun. 1992, 60, 1603–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engevik, M.A.; Luk, B.; Chang-Graham, A.L.; Hall, A.; Herrmann, B.; Ruan, W.; Endres, B.T.; Shi, Z.; Garey, K.W.; Hyser, J.M.; et al. Bifidobacterium dentium Fortifies the Intestinal Mucus Layer via Autophagy and Calcium Signaling Pathways. MBio 2019, 10, e01087-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koninkx, J.F.; Mirck, M.H.; Hendriks, H.G.; Mouwen, J.M.; van Dijk, J.E. Nippostrongylus brasiliensis: Histochemical changes in the composition of mucins in goblet cells during infection in rats. Exp. Parasitol. 1988, 65, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Koninkx, J.F.; Stemerdink, A.F.; Mirck, M.H.; Egberts, H.J.; van Dijk, J.E.; Mouwen, J.M. Histochemical changes in the composition of mucins in goblet cells during methotrexate-induced mucosal atrophy in rats. Exp. Pathol. 1988, 34, 125–132. [Google Scholar] [CrossRef]
- Meslin, J.C.; Bensaada, M.; Popot, F.; Andrieux, C. Differential influence of butyrate concentration on proximal and distal colonic mucosa in rats born germ-free and associated with a strain of Clostridium paraputrificum. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2001, 128, 379–384. [Google Scholar] [CrossRef]
- Koninkx, J.F.; Mirck, M.H.; Hendriks, H.G.; Mouwen, J.M.; van Dijk, J.E. Changes in the histochemical composition of mucins in goblet cells in the course of a Nippostrongylus brasiliensis infection in the rat. Tijdschr. Voor Diergeneeskd. 1990, 115, 1051–1057. [Google Scholar]
- Kim, D.H.; Kim, S.W.; Song, Y.J.; Oh, T.Y.; Han, S.U.; Kim, Y.B.; Joo, H.J.; Cho, Y.K.; Kim, D.Y.; Cho, S.W.; et al. Long-term evaluation of mice model infected with Helicobacter pylori: Focus on gastric pathology including gastric cancer. Aliment. Pharm. 2003, 18 (Suppl. 1), 14–23. [Google Scholar] [CrossRef]
- Turani, H.; Lurie, B.; Chaimoff, C.; Kessler, E. The diagnostic significance of sulfated acid mucin content in gastric intestinal metaplasia with early gastric cancer. Am. J. Gastroenterol. 1986, 81, 343–345. [Google Scholar]
- Periasamy, S.; Hsu, D.Z.; Chandrasekaran, V.R.; Liu, M.Y. Sesame oil accelerates healing of 2,4,6-trinitrobenzenesulfonic acid-induced acute colitis by attenuating inflammation and fibrosis. JPEN J. Parenter. Enter. Nutr. 2013, 37, 674–682. [Google Scholar] [CrossRef]
- Ishihara, K.; Ohara, S.; Azuumi, Y.; Goso, K.; Hotta, K. Changes of gastric mucus glycoproteins with aspirin administration in rats. Digestion 1984, 29, 98–102. [Google Scholar] [CrossRef]
- Schoffen, J.P.; Vicentini, F.A.; Marcelino, C.G.; Araujo, E.J.; Pedrosa, M.M.; Natali, M.R. Food restriction beginning at lactation interferes with the cellular dynamics of the mucosa and colonic myenteric innervation in adult rats. Acad. Bras. Cienc. 2014, 86, 1833–1848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabot, S.; Szylit, O.; Nugon-Baudon, L.; Meslin, J.C.; Vaissade, P.; Popot, F.; Viso, M. Variations in digestive physiology of rats after short duration flights aboard the US space shuttle. Dig. Dis. Sci. 2000, 45, 1687–1695. [Google Scholar] [CrossRef] [PubMed]
- Rubio, C.; Lizárraga, E.; Álvarez-Cilleros, D.; Pérez-Pardo, P.; Sanmartín-Salinas, P.; Toledo-Lobo, M.V.; Alvarez, C.; Escrivá, F.; Fernández-Lobato, M.; Guijarro, L.G.; et al. Aging in Male Wistar Rats Associates with Changes in Intestinal Microbiota, Gut Structure, and Cholecystokinin-Mediated Gut-Brain Axis Function. J. Gerontol. A Biol. Sci. Med. Sci. 2021, 76, 1915–1921. [Google Scholar] [CrossRef] [PubMed]
- Kaminishi, M.; Shimoyama, S.; Yamaguchi, H.; Ogawa, T.; Kawahara, M.; Jojima, Y.; Ohara, T. Effect of gastric mucosal denervation on gastric carcinogenesis. Nihon Geka Gakkai Zasshi 1992, 93, 927–931. [Google Scholar]
- Liu, J.; Walker, N.M.; Ootani, A.; Strubberg, A.M.; Clarke, L.L. Defective goblet cell exocytosis contributes to murine cystic fibrosis-associated intestinal disease. J. Clin. Investig. 2015, 125, 1056–1068. [Google Scholar] [CrossRef] [Green Version]
- Delvecchio, K.; Seman, S. Successful surgical excision of a massive symptomatic partially obstructing Brunner’s gland hamartoma: A case report. J. Surg. Case Rep. 2016, 2016, rjw206. [Google Scholar] [CrossRef] [Green Version]
- Camacho, M.; Machado, J.D.; Montesinos, M.S.; Criado, M.; Borges, R. Intragranular pH rapidly modulates exocytosis in adrenal chromaffin cells. J. Neurochem. 2006, 96, 324–334. [Google Scholar] [CrossRef]
- Ahn, D.H.; Crawley, S.C.; Hokari, R.; Kato, S.; Yang, S.C.; Li, J.D.; Kim, Y.S. TNF-alpha activates MUC2 transcription via NF-kappaB but inhibits via JNK activation. Cell Physiol. Biochem. 2005, 15, 29–40. [Google Scholar] [CrossRef]
- Enss, M.L.; Cornberg, M.; Wagner, S.; Gebert, A.; Henrichs, M.; Eisenblatter, R.; Beil, W.; Kownatzki, R.; Hedrich, H.J. Proinflammatory cytokines trigger MUC gene expression and mucin release in the intestinal cancer cell line LS180. Inflamm. Res. 2000, 49, 162–169. [Google Scholar] [CrossRef]
- Jarry, A.; Merlin, D.; Velcich, A.; Hopfer, U.; Augenlicht, L.H.; Laboisse, C.L. Interferon-Gamma Modulates Camp-Induced Mucin Exocytosis without Affecting Mucin Gene-Expression in a Human Colonic Goblet Cell-Line. Eur. J. Pharmacol. Mol. Pharmacol. 1994, 267, 95–103. [Google Scholar] [CrossRef]
- Dharmani, P.; Srivastava, V.; Kissoon-Singh, V.; Chadee, K. Role of intestinal mucins in innate host defense mechanisms against pathogens. J. Innate Immun. 2009, 1, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.D.; Kwon, E.J.; Park, D.W.; Song, S.Y.; Yoon, S.K.; Baek, S.H. Interleukin-1beta induces MUC2 and MUC5AC synthesis through cyclooxygenase-2 in NCI-H292 cells. Mol. Pharm. 2002, 62, 1112–1118. [Google Scholar] [CrossRef] [Green Version]
- Sharba, S.; Navabi, N.; Padra, M.; Persson, J.A.; Quintana-Hayashi, M.P.; Gustafsson, J.K.; Szeponik, L.; Venkatakrishnan, V.; Sjoling, A.; Nilsson, S.; et al. Interleukin 4 induces rapid mucin transport, increases mucus thickness and quality and decreases colitis and Citrobacter rodentium in contact with epithelial cells. Virulence 2019, 10, 97–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wynn, T.A. Type 2 cytokines: Mechanisms and therapeutic strategies. Nat. Rev. Immunol. 2015, 15, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Iwashita, J.; Sato, Y.; Sugaya, H.; Takahashi, N.; Sasaki, H.; Abe, T. mRNA of MUC2 is stimulated by IL-4, IL-13 or TNF-alpha through a mitogen-activated protein kinase pathway in human colon cancer cells. Immunol. Cell Biol. 2003, 81, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yao, N.; Fu, X.; Wei, L.; Ding, M.M.; Pang, Y.; Liu, D.; Ren, Y.; Guo, M. Butylphthalide ameliorates airway inflammation and mucus hypersecretion via NF-kappaB in a murine asthma model. Int. Immunopharmacol. 2019, 76, 105873. [Google Scholar] [CrossRef]
- Guan, Y.; Zhu, J.P.; Shen, J.; Jia, Y.L.; Jin, Y.C.; Dong, X.W.; Xie, Q.M. Salvianolic acid B improves airway hyperresponsiveness by inhibiting MUC5AC overproduction associated with Erk1/2/P38 signaling. Eur. J. Pharm. 2018, 824, 30–39. [Google Scholar] [CrossRef]
- Fuell, C.; Kober, O.I.; Hautefort, I.; Juge, N. Mice deficient in intestinal gammadelta intraepithelial lymphocytes display an altered intestinal O-glycan profile compared with wild-type littermates. Glycobiology 2015, 25, 42–54. [Google Scholar] [CrossRef] [Green Version]
- Giron, L.B.; Tanes, C.E.; Schleimann, M.H.; Engen, P.A.; Mattei, L.M.; Anzurez, A.; Damra, M.; Zhang, H.; Bittinger, K.; Bushman, F.; et al. Sialylation and fucosylation modulate inflammasome-activating eIF2 Signaling and microbial translocation during HIV infection. Mucosal Immunol. 2020, 13, 753–766. [Google Scholar] [CrossRef] [Green Version]
- Beum, P.V.; Basma, H.; Bastola, D.R.; Cheng, P.W. Mucin biosynthesis: Upregulation of core 2 beta 1,6 N-acetylglucosaminyltransferase by retinoic acid and Th2 cytokines in a human airway epithelial cell line. Am. J. Physiol. Lung Cell Mol. Physiol. 2005, 288, L116–124. [Google Scholar] [CrossRef] [Green Version]
- Melhem, H.; Regan-Komito, D.; Niess, J.H. Mucins Dynamics in Physiological and Pathological Conditions. Int. J. Mol. Sci. 2021, 22, 13642. [Google Scholar] [CrossRef] [PubMed]
- Fekete, E.; Allain, T.; Amat, C.B.; Mihara, K.; Saifeddine, M.; Hollenberg, M.D.; Chadee, K.; Buret, A.G. Giardia duodenalis cysteine proteases cleave proteinase-activated receptor-2 to regulate intestinal goblet cell mucin gene expression. Int. J. Parasitol. 2022, 52, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Birchenough, G.M.; Johansson, M.E.; Gustafsson, J.K.; Bergstrom, J.H.; Hansson, G.C. New developments in goblet cell mucus secretion and function. Mucosal Immunol. 2015, 8, 712–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grondin, J.A.; Kwon, Y.H.; Far, P.M.; Haq, S.; Khan, W.I. Mucins in Intestinal Mucosal Defense and Inflammation: Learning From Clinical and Experimental Studies. Front. Immunol. 2020, 11, 2054. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, J.K.; Davis, J.E.; Rappai, T.; McDonald, K.G.; Kulkarni, D.H.; Knoop, K.A.; Hogan, S.P.; Fitzpatrick, J.A.; Lencer, W.I.; Newberry, R.D. Intestinal goblet cells sample and deliver lumenal antigens by regulated endocytic uptake and transcytosis. Elife 2021, 10, e67292. [Google Scholar] [CrossRef]
- Wu, X.; Conlin, V.S.; Morampudi, V.; Ryz, N.R.; Nasser, Y.; Bhinder, G.; Bergstrom, K.S.; Yu, H.B.; Waterhouse, C.C.; Buchan, A.M.; et al. Vasoactive intestinal polypeptide promotes intestinal barrier homeostasis and protection against colitis in mice. PLoS ONE 2015, 10, e0125225. [Google Scholar] [CrossRef]
- Schwerdtfeger, L.A.; Tobet, S.A. Vasoactive intestinal peptide regulates ileal goblet cell production in mice. Physiol. Rep. 2020, 8, e14363. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Yoshizaki, K.; Kishimoto, T.; Ito, H. IL-6 is required for the development of Th1 cell-mediated murine colitis. J. Immunol. 2000, 164, 4878–4882. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Walia, B.; Evans, J.; Gewirtz, A.T.; Merlin, D.; Sitaraman, S.V. IL-6 induces NF-kappa B activation in the intestinal epithelia. J. Immunol. 2003, 171, 3194–3201. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.H.; Guo, J.L.; Wen, Y.Q.; Li, Y.X.; Wei, D.D.; Yang, R.L.; Mu, X.Y.; Wang, H.C. Role of IgG plasma cells in the change of protein C system in ulcerative colitis. Acta Physiol. Sin. 2017, 69, 172–182. [Google Scholar]
- Collins, J.W.; Keeney, K.M.; Crepin, V.F.; Rathinam, V.A.; Fitzgerald, K.A.; Finlay, B.B.; Frankel, G. Citrobacter rodentium: Infection, inflammation and the microbiota. Nat. Rev. Microbiol. 2014, 12, 612–623. [Google Scholar] [CrossRef] [PubMed]
- Desai, M.S.; Seekatz, A.M.; Koropatkin, N.M.; Kamada, N.; Hickey, C.A.; Wolter, M.; Pudlo, N.A.; Kitamoto, S.; Terrapon, N.; Muller, A.; et al. A Dietary Fiber-Deprived Gut Microbiota Degrades the Colonic Mucus Barrier and Enhances Pathogen Susceptibility. Cell 2016, 167, 1339–1353.E21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauvaitre, T.; Etienne-Mesmin, L.; Sivignon, A.; Mosoni, P.; Courtin, C.M.; Van de Wiele, T.; Blanquet-Diot, S. Tripartite relationship between gut microbiota, intestinal mucus and dietary fibers: Towards preventive strategies against enteric infections. FEMS Microbiol. Rev. 2021, 45, fuaa052. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, J.; Yi, J.; Liu, Y.; Yu, Z.; Chen, S.; Liu, X. Increased mucin-degrading bacteria by high protein diet leads to thinner mucus layer and aggravates experimental colitis. J. Gastroenterol. Hepatol. 2021, 36, 2864–2874. [Google Scholar] [CrossRef]
- Rohr, M.W.; Narasimhulu, C.A.; Rudeski-Rohr, T.A.; Parthasarathy, S. Negative Effects of a High-Fat Diet on Intestinal Permeability: A Review. Adv. Nutr. 2020, 11, 77–91. [Google Scholar] [CrossRef] [Green Version]
- Mastrodonato, M.; Calamita, G.; Mentino, D.; Scillitani, G. High-fat Diet Alters the Glycosylation Patterns of Duodenal Mucins in a Murine Model. J. Histochem. Cytochem. 2020, 68, 279–294. [Google Scholar] [CrossRef]
- Corfield, A.P.; Myerscough, N.; Longman, R.; Sylvester, P.; Arul, S.; Pignatelli, M. Mucins and mucosal protection in the gastrointestinal tract: New prospects for mucins in the pathology of gastrointestinal disease. Gut 2000, 47, 589–594. [Google Scholar] [CrossRef] [Green Version]
- Chassaing, B.; Koren, O.; Goodrich, J.K.; Poole, A.C.; Srinivasan, S.; Ley, R.E.; Gewirtz, A.T. Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature 2015, 519, 92–96. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, I.; Yusuf, K.; Roy, B.C.; Stubbs, J.; Anant, S.; Attard, T.M.; Sampath, V.; Umar, S. Dietary Interventions Ameliorate Infectious Colitis by Restoring the Microbiome and Promoting Stem Cell Proliferation in Mice. Int. J. Mol. Sci. 2021, 23, 339. [Google Scholar] [CrossRef]
- Rinninella, E.; Cintoni, M.; Raoul, P.; Gasbarrini, A.; Mele, M.C. Food Additives, Gut Microbiota, and Irritable Bowel Syndrome: A Hidden Track. Int. J. Env. Res. Public Health 2020, 17, 8816. [Google Scholar] [CrossRef]
- Desvaux, M.; Dumas, E.; Chafsey, I.; Hebraud, M. Protein cell surface display in Gram-positive bacteria: From single protein to macromolecular protein structure. FEMS Microbiol. Lett. 2006, 256, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Mukai, T.; Arihara, K. Presence of Intestinal Lectin-Binding Glycoproteins on the Cell-Surface of Lactobacillus-Acidophilus. Biosci. Biotechnol. Biochem. 1994, 58, 1851–1854. [Google Scholar] [CrossRef] [Green Version]
- Mukai, T.; Kaneko, S.; Ohori, H. Haemagglutination and glycolipid-binding activities of Lactobacillus reuteri. Lett. Appl. Microbiol. 1998, 27, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Belzer, C.; Chia, L.W.; Aalvink, S.; Chamlagain, B.; Piironen, V.; Knol, J.; de Vos, W.M. Microbial Metabolic Networks at the Mucus Layer Lead to Diet-Independent Butyrate and Vitamin B12 Production by Intestinal Symbionts. MBio 2017, 8, e00770-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jara, D.; Carvajal, P.; Castro, I.; Barrera, M.J.; Aguilera, S.; Gonzalez, S.; Molina, C.; Hermoso, M.; Gonzalez, M.J. Type I Interferon Dependent hsa-miR-145-5p Downregulation Modulates MUC1 and TLR4 Overexpression in Salivary Glands From Sjogren’s Syndrome Patients. Front. Immunol. 2021, 12, 685837. [Google Scholar] [CrossRef] [PubMed]
- Das, N.; Menon, N.G.; de Almeida, L.G.N.; Woods, P.S.; Heynen, M.L.; Jay, G.D.; Caffery, B.; Jones, L.; Krawetz, R.; Schmidt, T.A.; et al. Proteomics Analysis of Tears and Saliva From Sjogren’s Syndrome Patients. Front. Pharm. 2021, 12, 787193. [Google Scholar] [CrossRef]
- Culp, D.J.; Stewart, C.; Wallet, S.M. Oral epithelial membrane-associated mucins and transcriptional changes with Sjogren’s syndrome. Oral. Dis. 2019, 25, 1325–1334. [Google Scholar] [CrossRef]
- Chaudhury, N.M.; Proctor, G.B.; Karlsson, N.G.; Carpenter, G.H.; Flowers, S.A. Reduced Mucin-7 (Muc7) Sialylation and Altered Saliva Rheology in Sjogren’s Syndrome Associated Oral Dryness. Mol. Cell. Proteom. 2016, 15, 1048–1059. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.F.; Chen, Y.; Han, J.M.; Zhang, H.; Chen, X.P.; Zou, W.J.; Liang, L.Y.; Xu, C.C.; Liu, Z.G. MUC19 expression in human ocular surface and lacrimal gland and its alteration in Sjogren syndrome patients. Exp. Eye Res. 2008, 86, 403–411. [Google Scholar] [CrossRef]
- Boucher, R.C. Muco-Obstructive Lung Diseases. N. Engl. J. Med. 2019, 380, 1941–1953. [Google Scholar] [CrossRef]
- Bafna, S.; Kaur, S.; Batra, S.K. Membrane-bound mucins: The mechanistic basis for alterations in the growth and survival of cancer cells. Oncogene 2010, 29, 2893–2904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khatri, I.A.; Ho, C.; Specian, R.D.; Forstner, J.F. Characteristics of rodent intestinal mucin Muc3 and alterations in a mouse model of human cystic fibrosis. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 280, G1321–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finkbeiner, W.E.; Zlock, L.T.; Morikawa, M.; Lao, A.Y.; Dasari, V.; Widdicombe, J.H. Cystic fibrosis and the relationship between mucin and chloride secretion by cultures of human airway gland mucous cells. Am. J. Physiol. Lung Cell Mol. Physiol. 2011, 301, L402–L414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, C.J.; Hyde, K.; Ho, S.B.; Harris, A. Cystic fibrosis of the pancreas: Involvement of MUC6 mucin in obstruction of pancreatic ducts. Mol. Med. 1997, 3, 403–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batson, B.; Zorn, B.; Radicioni, G.; Livengood, S.; Kumagai, T.; Dang, H.; Ceppe, A.; Clapp, P.; Tunney, M.; Elborn, S.; et al. Cystic Fibrosis Airway Mucus Hyperconcentration Produces a Vicious Cycle of Mucin, Pathogen, and Inflammatory Interactions that Promote Disease Persistence. Am. J. Respir. Cell Mol. Biol. 2022, 67, 253–265. [Google Scholar] [CrossRef]
- Niv, Y. Helicobacter pylori and gastric mucin expression: A systematic review and meta-analysis. World J. Gastroenterol. 2015, 21, 9430–9436. [Google Scholar] [CrossRef]
- Duarte, H.O.; Freitas, D.; Gomes, C.; Gomes, J.; Magalhaes, A.; Reis, C.A. Mucin-Type O-Glycosylation in Gastric Carcinogenesis. Biomolecules 2016, 6, 33. [Google Scholar] [CrossRef] [Green Version]
- Jin, C.; Kenny, D.T.; Skoog, E.C.; Padra, M.; Adamczyk, B.; Vitizeva, V.; Thorell, A.; Venkatakrishnan, V.; Linden, S.K.; Karlsson, N.G. Structural diversity of human gastric mucin glycans. Mol. Cell. Proteom. 2017, 16, 743–758. [Google Scholar] [CrossRef]
- Buisine, M.P.; Desreumaux, P.; Leteurtre, E.; Copin, M.C.; Colombel, J.F.; Porchet, N.; Aubert, J.P. Mucin gene expression in intestinal epithelial cells in Crohn’s disease. Gut 2001, 49, 544–551. [Google Scholar] [CrossRef] [Green Version]
- Moehle, C.; Ackermann, N.; Langmann, T.; Aslanidis, C.; Kel, A.; Kel-Margoulis, O.; Schmitz-Madry, A.; Zahn, A.; Stremmel, W.; Schmitz, G. Aberrant intestinal expression and allelic variants of mucin genes associated with inflammatory bowel disease. J. Mol. Med. 2006, 84, 1055–1066. [Google Scholar] [CrossRef]
- Breugelmans, T.; Van Spaendonk, H.; De Man, J.G.; De Schepper, H.U.; Jauregui-Amezaga, A.; Macken, E.; Linden, S.K.; Pintelon, I.; Timmermans, J.P.; De Winter, B.Y.; et al. In-Depth Study of Transmembrane Mucins in Association with Intestinal Barrier Dysfunction During the Course of T Cell Transfer and DSS-Induced Colitis. J. Crohns Colitis 2020, 14, 974–994. [Google Scholar] [CrossRef] [PubMed]
- Bardin, N.; Reumaux, D.; Geboes, K.; Colombel, J.F.; Blot-Chabaud, M.; Sampol, J.; Duthilleul, P.; Dignat-George, F. Increased expression of CD146, a new marker of the endothelial junction in active inflammatory bowel disease. Inflamm. Bowel Dis. 2006, 12, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Whitcomb, E.; Liu, X.; Xiao, S.Y. Crohn enteritis-associated small bowel adenocarcinomas exhibit gastric differentiation. Hum. Pathol. 2014, 45, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Borralho, P.; Vieira, A.; Freitas, J.; Chaves, P.; Soares, J. Aberrant gastric apomucin expression in ulcerative colitis and associated neoplasia. J. Crohns Colitis 2007, 1, 35–40. [Google Scholar] [CrossRef] [Green Version]
- Pothuraju, R.; Rachagani, S.; Krishn, S.R.; Chaudhary, S.; Nimmakayala, R.K.; Siddiqui, J.A.; Ganguly, K.; Lakshmanan, I.; Cox, J.L.; Mallya, K.; et al. Molecular implications of MUC5AC-CD44 axis in colorectal cancer progression and chemoresistance. Mol. Cancer 2020, 19, 37. [Google Scholar] [CrossRef] [Green Version]
- Byrd, J.C.; Bresalier, R.S. Mucins and mucin binding proteins in colorectal cancer. Cancer Metastasis Rev. 2004, 23, 77–99. [Google Scholar] [CrossRef]
- Coleman, O.I.; Haller, D. Microbe-Mucus Interface in the Pathogenesis of Colorectal Cancer. Cancers 2021, 13, 616. [Google Scholar] [CrossRef]
- Sheng, Y.H.; Wong, K.Y.; Seim, I.; Wang, R.; He, Y.; Wu, A.; Patrick, M.; Lourie, R.; Schreiber, V.; Giri, R.; et al. MUC13 promotes the development of colitis-associated colorectal tumors via beta-catenin activity. Oncogene 2019, 38, 7294–7310. [Google Scholar] [CrossRef]
- Lu, S.; Catalano, C.; Huhn, S.; Pardini, B.; Partu, L.; Vymetalkova, V.; Vodickova, L.; Levy, M.; Buchler, T.; Hemminki, K.; et al. Single nucleotide polymorphisms within MUC4 are associated with colorectal cancer survival. PLoS ONE 2019, 14, e0216666. [Google Scholar] [CrossRef]
- Sylvester, P.A.; Myerscough, N.; Warren, B.F.; Carlstedt, I.; Corfield, A.P.; Durdey, P.; Thomas, M.G. Differential expression of the chromosome 11 mucin genes in colorectal cancer. J. Pathol. 2001, 195, 327–335. [Google Scholar] [CrossRef]
- Oh, H.R.; An, C.H.; Yoo, N.J.; Lee, S.H. Frameshift mutations of MUC15 gene in gastric and its regional heterogeneity in gastric and colorectal cancers. Pathol. Oncol. Res. 2015, 21, 713–718. [Google Scholar] [CrossRef]
- Liu, Z.; Gu, Y.; Li, X.; Zhou, L.; Cheng, X.; Jiang, H.; Huang, Y.; Zhang, Y.; Xu, T.; Yang, W.; et al. Mucin 16 Promotes Colorectal Cancer Development and Progression Through Activation of Janus Kinase 2. Dig. Dis. Sci. 2022, 67, 2195–2208. [Google Scholar] [CrossRef] [PubMed]
- Iranmanesh, H.; Entezari, M.; Rejali, L.; Nazemalhosseini-Mojarad, E.; Maghsoudloo, M.; Aghdaei, H.A.; Zali, M.R.; Hushmandi, K.; Rabiee, N.; Makvandi, P.; et al. The association of clinicopathological characterizations of colorectal cancer with membrane-bound mucins genes and LncRNAs. Pathol.-Res. Pract. 2022, 233, 153883. [Google Scholar] [CrossRef] [PubMed]
- Sumida, T.; Azuma, N.; Moriyama, M.; Takahashi, H.; Asashima, H.; Honda, F.; Abe, S.; Ono, Y.; Hirota, T.; Hirata, S.; et al. Clinical practice guideline for Sjogren’s syndrome 2017. Mod. Rheumatol. 2018, 28, 383–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, Y.; Takeuchi, T.; Tsubota, K. Autoimmune Epithelitis and Chronic Inflammation in Sjogren’s Syndrome-Related Dry Eye Disease. Int. J. Mol. Sci. 2021, 22, 11820. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Carrasco, M.; Fuentes-Alexandro, S.; Escarcega, R.O.; Salgado, G.; Riebeling, C.; Cervera, R. Pathophysiology of Sjogren’s syndrome. Arch. Med. Res. 2006, 37, 921–932. [Google Scholar] [CrossRef] [PubMed]
- Decker, P.; Moulinet, T.; Pontille, F.; Cravat, M.; De Carvalho Bittencourt, M.; Jaussaud, R. An updated review of anti-Ro52 (TRIM21) antibodies impact in connective tissue diseases clinical management. Autoimmun. Rev. 2022, 21, 103013. [Google Scholar] [CrossRef] [PubMed]
- The definition and classification of dry eye disease: Report of the Definition and Classification Subcommittee of the International Dry Eye WorkShop (2007). Ocul. Surf. 2007, 5, 75–92. [CrossRef]
- Dartt, D.A.; Masli, S. Conjunctival epithelial and goblet cell function in chronic inflammation and ocular allergic inflammation. Curr. Opin. Allergy Clin. Immunol. 2014, 14, 464–470. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Bair, J.A.; Hodges, R.R.; Serhan, C.N.; Dartt, D.A. Resolvin E1 Reduces Leukotriene B4-Induced Intracellular Calcium Increase and Mucin Secretion in Rat Conjunctival Goblet Cells. Am. J. Pathol. 2020, 190, 1823–1832. [Google Scholar] [CrossRef]
- Puro, D.G. Impact of P2X7 Purinoceptors on Goblet Cell Function: Implications for Dry Eye. Int. J. Mol. Sci. 2021, 22, 6935. [Google Scholar] [CrossRef] [PubMed]
- Keith, J.D.; Henderson, A.G.; Fernandez-Petty, C.M.; Davis, J.M.; Oden, A.M.; Birket, S.E. Muc5b Contributes to Mucus Abnormality in Rat Models of Cystic Fibrosis. Front. Physiol. 2022, 13, 884166. [Google Scholar] [CrossRef] [PubMed]
- Ota, H.; Nakayama, J.; Momose, M.; Hayama, M.; Akamatsu, T.; Katsuyama, T.; Graham, D.Y.; Genta, R.M. Helicobacter pylori infection produces reversible glycosylation changes to gastric mucins. Virchows Arch. 1998, 433, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Sidebotham, R.L.; Baron, J.H. Hypothesis: Helicobacter pylori, urease, mucus, and gastric ulcer. Lancet 1990, 335, 193–195. [Google Scholar] [CrossRef]
- Linden, S.K.; Wickstrom, C.; Lindell, G.; Gilshenan, K.; Carlstedt, I. Four modes of adhesion are used during Helicobacter pylori binding to human mucins in the oral and gastric niches. Helicobacter 2008, 13, 81–93. [Google Scholar] [CrossRef]
- Xavier, R.J.; Podolsky, D.K. Unravelling the pathogenesis of inflammatory bowel disease. Nature 2007, 448, 427–434. [Google Scholar] [CrossRef]
- Ng, Q.X.; Soh, A.Y.S.; Loke, W.; Lim, D.Y.; Yeo, W.S. The role of inflammation in irritable bowel syndrome (IBS). J. Inflamm. Res. 2018, 11, 345–349. [Google Scholar] [CrossRef] [Green Version]
- Niv, Y. Mucin gene expression in the intestine of ulcerative colitis patients: A systematic review and meta-analysis. Eur. J. Gastroenterol. Hepatol. 2016, 28, 1241–1245. [Google Scholar] [CrossRef]
- Buisine, M.P.; Desreumaux, P.; Debailleul, V.; Gambiez, L.; Geboes, K.; Ectors, N.; Delescaut, M.P.; Degand, P.; Aubert, J.P.; Colombel, J.F.; et al. Abnormalities in mucin gene expression in Crohn’s disease. Inflamm. Bowel Dis. 1999, 5, 24–32. [Google Scholar] [CrossRef]
- Da Silva, S.; Robbe-Masselot, C.; Ait-Belgnaoui, A.; Mancuso, A.; Mercade-Loubiere, M.; Salvador-Cartier, C.; Gillet, M.; Ferrier, L.; Loubiere, P.; Dague, E.; et al. Stress disrupts intestinal mucus barrier in rats via mucin O-glycosylation shift: Prevention by a probiotic treatment. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G420–429. [Google Scholar] [CrossRef]
- Kandy, S.K.; Radhakrishnan, R. Crowding induced membrane remodeling: Interplay of membrane tension, polymer density, architecture. Biophys. J. 2022, 121, 3674–3683. [Google Scholar] [CrossRef] [PubMed]
- Breugelmans, T.; Oosterlinck, B.; Arras, W.; Ceuleers, H.; De Man, J.; Hold, G.L.; De Winter, B.Y.; Smet, A. The role of mucins in gastrointestinal barrier function during health and disease. Lancet Gastroenterol. Hepatol. 2022, 7, 455–471. [Google Scholar] [CrossRef] [PubMed]
- Mockl, L. The Emerging Role of the Mammalian Glycocalyx in Functional Membrane Organization and Immune System Regulation. Front. Cell Dev. Biol. 2020, 8, 253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantero-Recasens, G.; Alonso-Maranon, J.; Lobo-Jarne, T.; Garrido, M.; Iglesias, M.; Espinosa, L.; Malhotra, V. Reversing chemorefraction in colorectal cancer cells by controlling mucin secretion. Elife 2022, 11, e73926. [Google Scholar] [CrossRef] [PubMed]



| Disease | Sjögren’s Syndrome | Cystic Fibrosis | H. pylori Infection | CD | UC | CRC |
|---|---|---|---|---|---|---|
| Main organs | Mouth, eye | Lungs, pancreas | Stomach | Small intestine | Small intestine | Colon, rectum |
| Cardinal symptoms | Dry accompanies other immune system disorders | Cough, infection, nutritional deficiencies | Pain, bloating, ulcers | Pain, diarrhea, rectal bleeding | Bloody purulent stool, abdominal pain or cramping | Bloody purulent stool, change in bowel habits, abdominal pain or cramping |
| Mucus thickness | ↓ | Sticky and thick | ↓ | ↑ | ↓ | ↓ |
| Glycosylation | ↓ | ↑ | ↓ | Unknown | ↓ | ↓ |
| Sialylation | ↓ | ↑ | ↑ | ↑ | ↑ | ↑ |
| MUC1 | ↑ | ↑ | ↑ | ↓ | ↑ | ↑ |
| MUC2 | ? | ↑ | ↑ | ↓ | ↓ | ↓ |
| MUC3 | ? | ↑ | Undetectable | ↓ | Unchanged | Unchanged |
| MUC4 | Undetectable/unchanged | ↑ | Undetectable | ↓ | ↑ | ↓ |
| MUC5AC | ↑ | ↑ | ↓ | ↓ | ↑ | ↑ |
| MUC5B | Unchanged | ↑ | ↑ | ↓ | ↓ | Unchanged |
| MUC6 | ? | ↑ | ↑ | ↑ | ↑ | Undetectable |
| MUC7 | ↑ | ? | Undetectable | ? | ? | Unchanged |
| MUC8 | ? | Unchanged | Undetectable | Undetectable | Undetectable | ? |
| MUC10 | ? | ? | ? | Undetectable | Undetectable | ? |
| MUC12 | Undetectable | ? | ? | ↓ | ↓ | ↓ |
| MUC13 | Undetectable | ? | ? | ↑ | ↑ | ↑ |
| MUC15 | Unchanged | ? | ? | ? | ? | ↑ |
| MUC16 | Unchanged | ↑ | Undetectable | ? | ? | ↑ |
| MUC17 | Undetectable | ? | ? | ↓ | ↓ | ? |
| MUC18 | ? | ? | ? | ↑ | ↑ | ? |
| MUC19 | ↓ | ? | ? | ? | ? | ? |
| MUC20 | ↓ | ? | ? | ↓ | ↓ | ↑ |
| MUC21 | ↓ | ? | ? | ? | ? | ? |
| Refs. | [146,147,148,149,150] | [151,152,153,154,155,156] | [157,158,159] | [160,161,162,163,164] | [160,161,162,163,165] | [71,166,167,168,169,170,171,172,173,174] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, C.; Gao, H.; Xin, S.; Hua, R.; Guo, X.; Han, Y.; Shang, H.; Xu, J. View from the Biological Property: Insight into the Functional Diversity and Complexity of the Gut Mucus. Int. J. Mol. Sci. 2023, 24, 4227. https://doi.org/10.3390/ijms24044227
He C, Gao H, Xin S, Hua R, Guo X, Han Y, Shang H, Xu J. View from the Biological Property: Insight into the Functional Diversity and Complexity of the Gut Mucus. International Journal of Molecular Sciences. 2023; 24(4):4227. https://doi.org/10.3390/ijms24044227
Chicago/Turabian StyleHe, Chengwei, Han Gao, Shuzi Xin, Rongxuan Hua, Xueran Guo, Yimin Han, Hongwei Shang, and Jingdong Xu. 2023. "View from the Biological Property: Insight into the Functional Diversity and Complexity of the Gut Mucus" International Journal of Molecular Sciences 24, no. 4: 4227. https://doi.org/10.3390/ijms24044227