
Computational Insight into Intraspecies Distinctions in Pseudoalteromonas distincta: Carotenoid-like Synthesis Traits and Genomic Heterogeneity
 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Results and Discussion
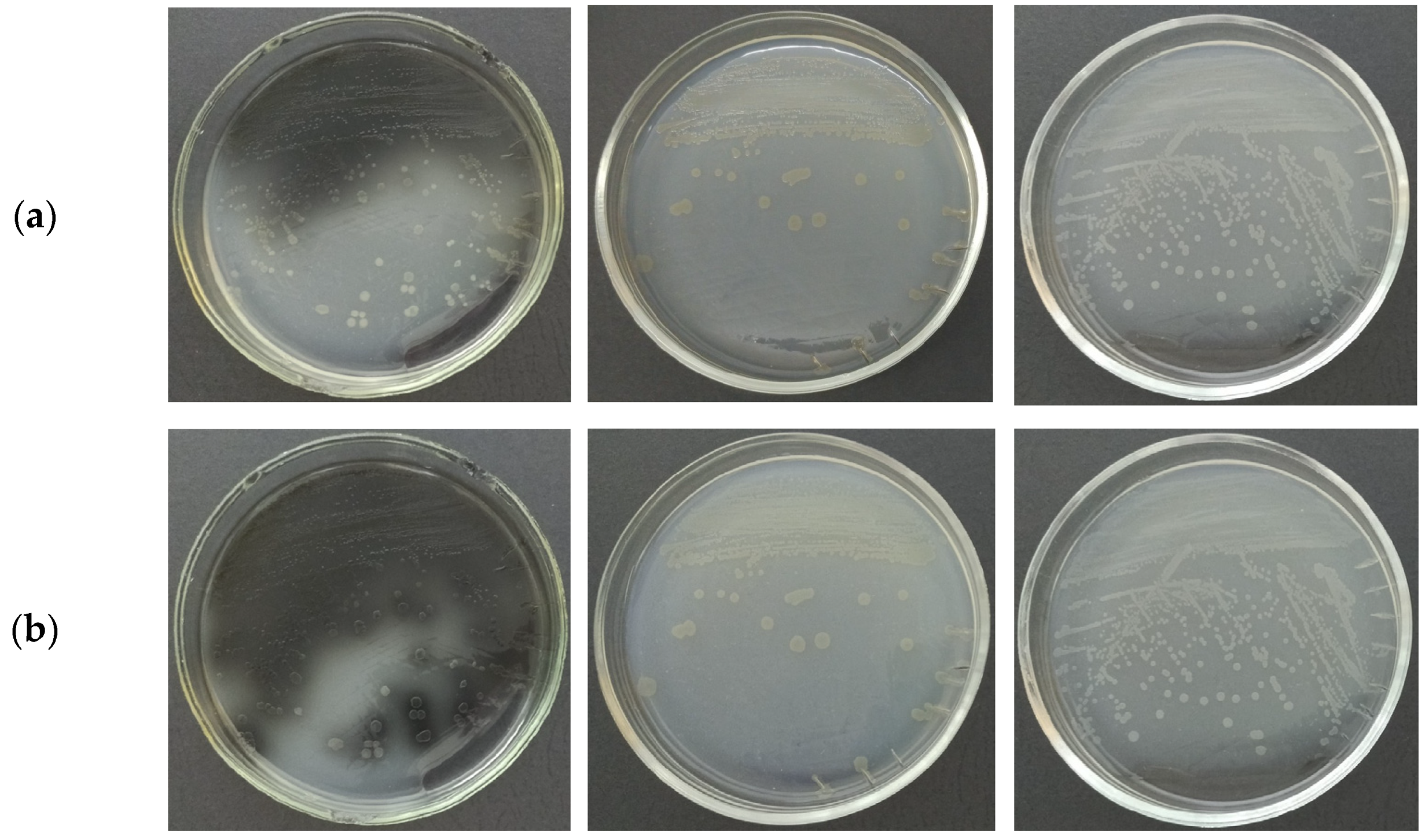
2.1. Phenotypic Characterization of the P. distincta Strains
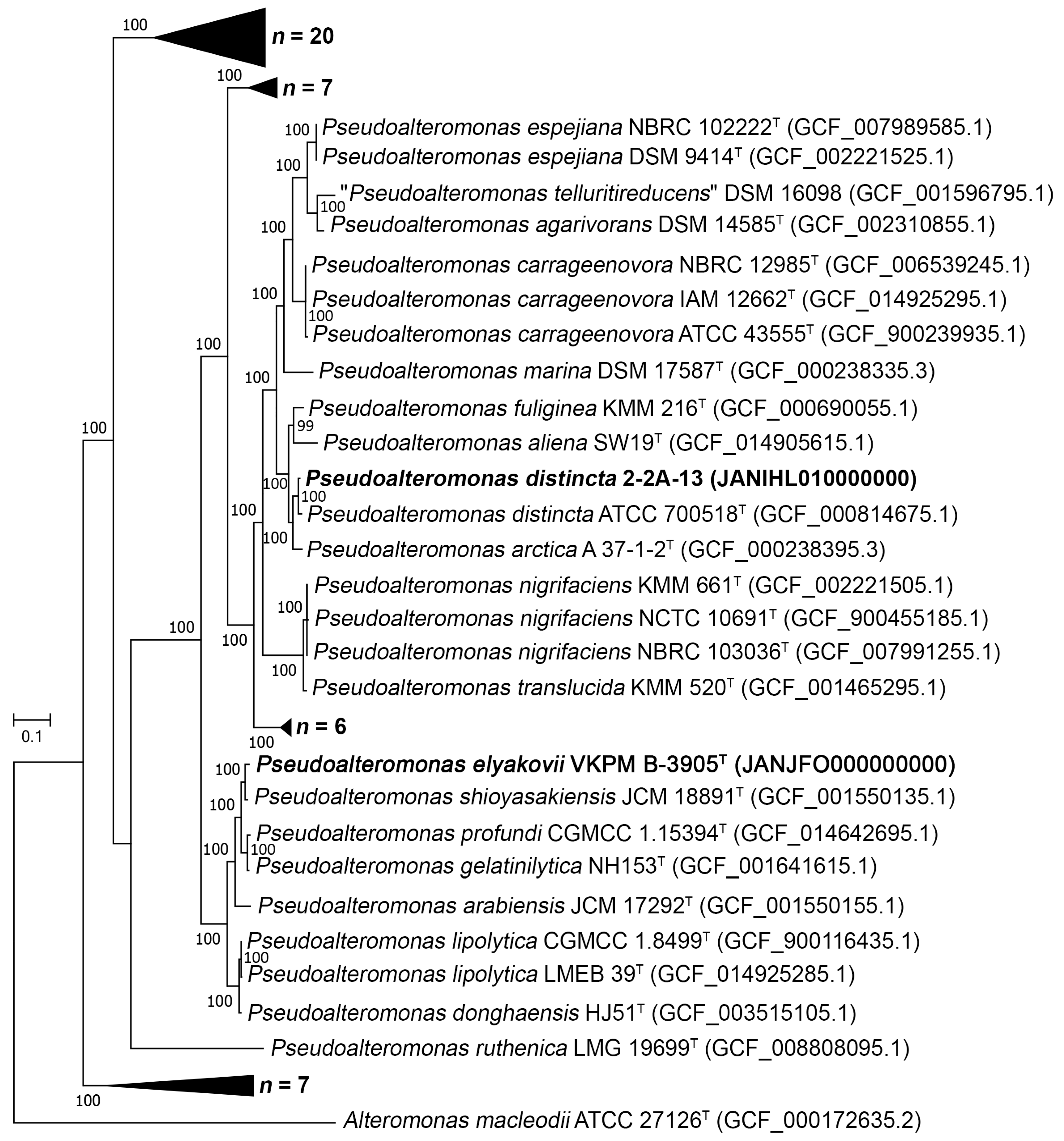
2.2. Phylogenomic Relationships of the P. distincta Strains within the Genus Pseudoalteromonas
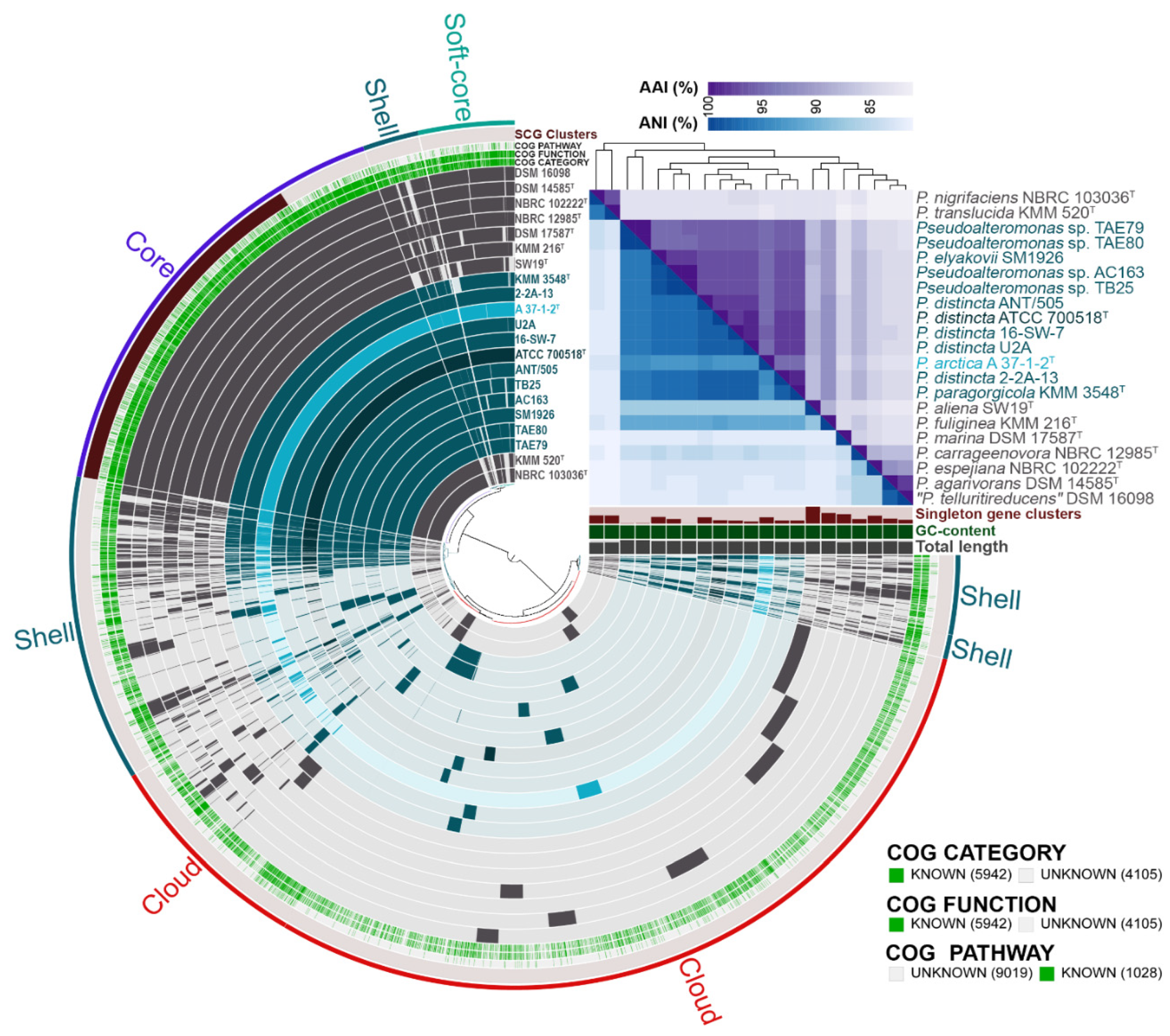
2.3. Pan-Genomic Characterization of the P. distincta Strains
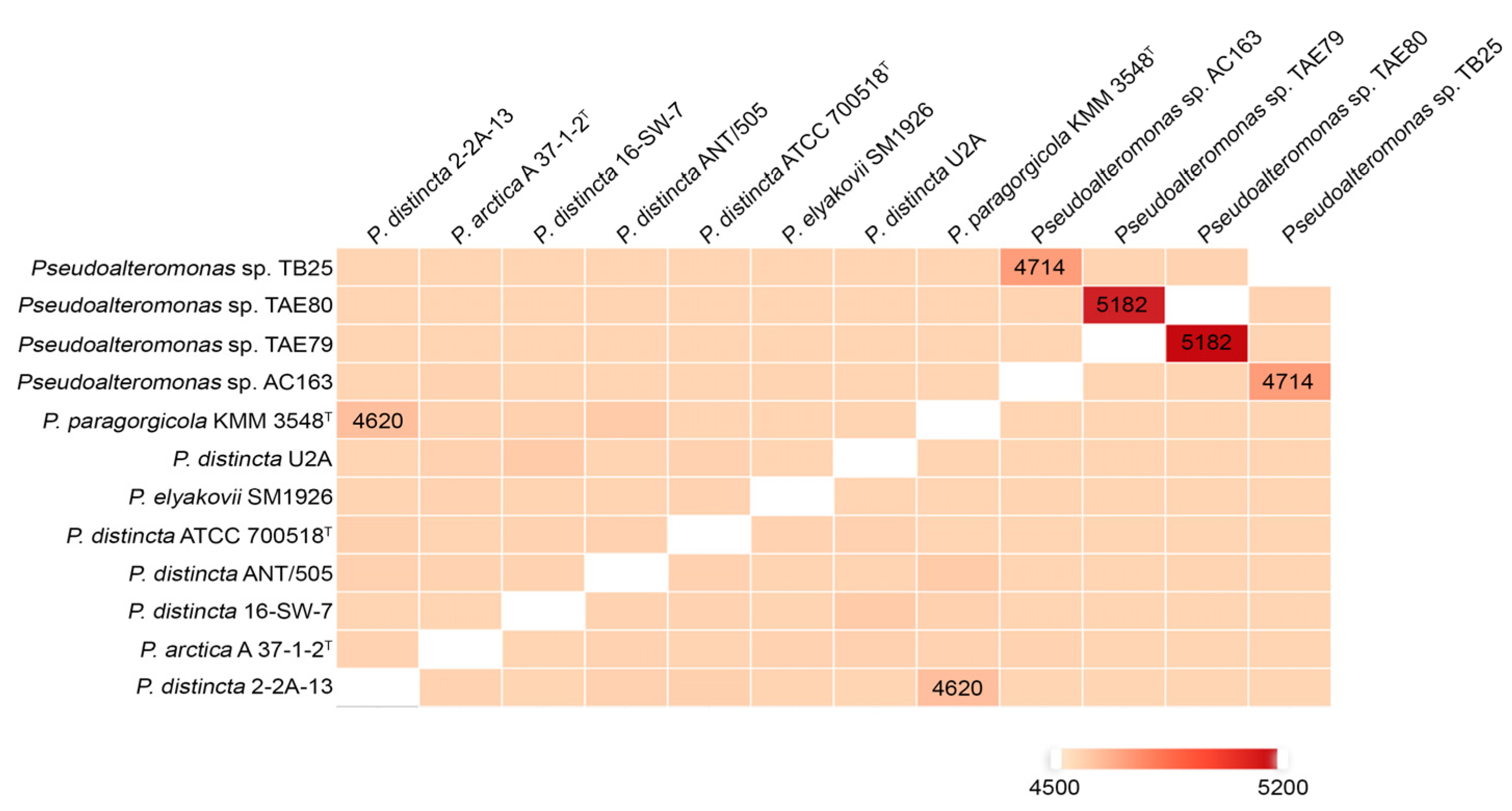
2.4. Phylogenetic Core-Genes Analysis of the P. distincta Strains
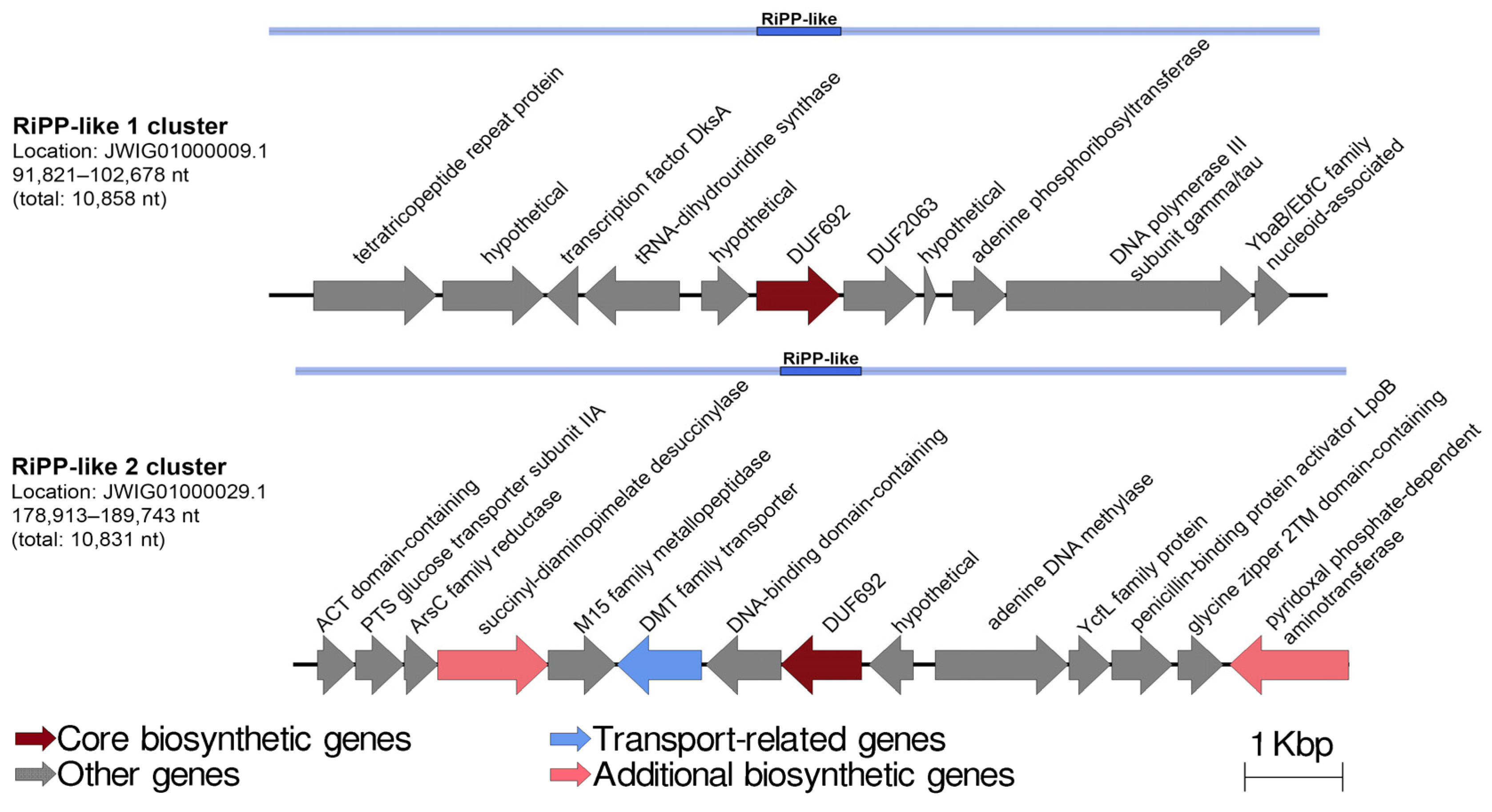
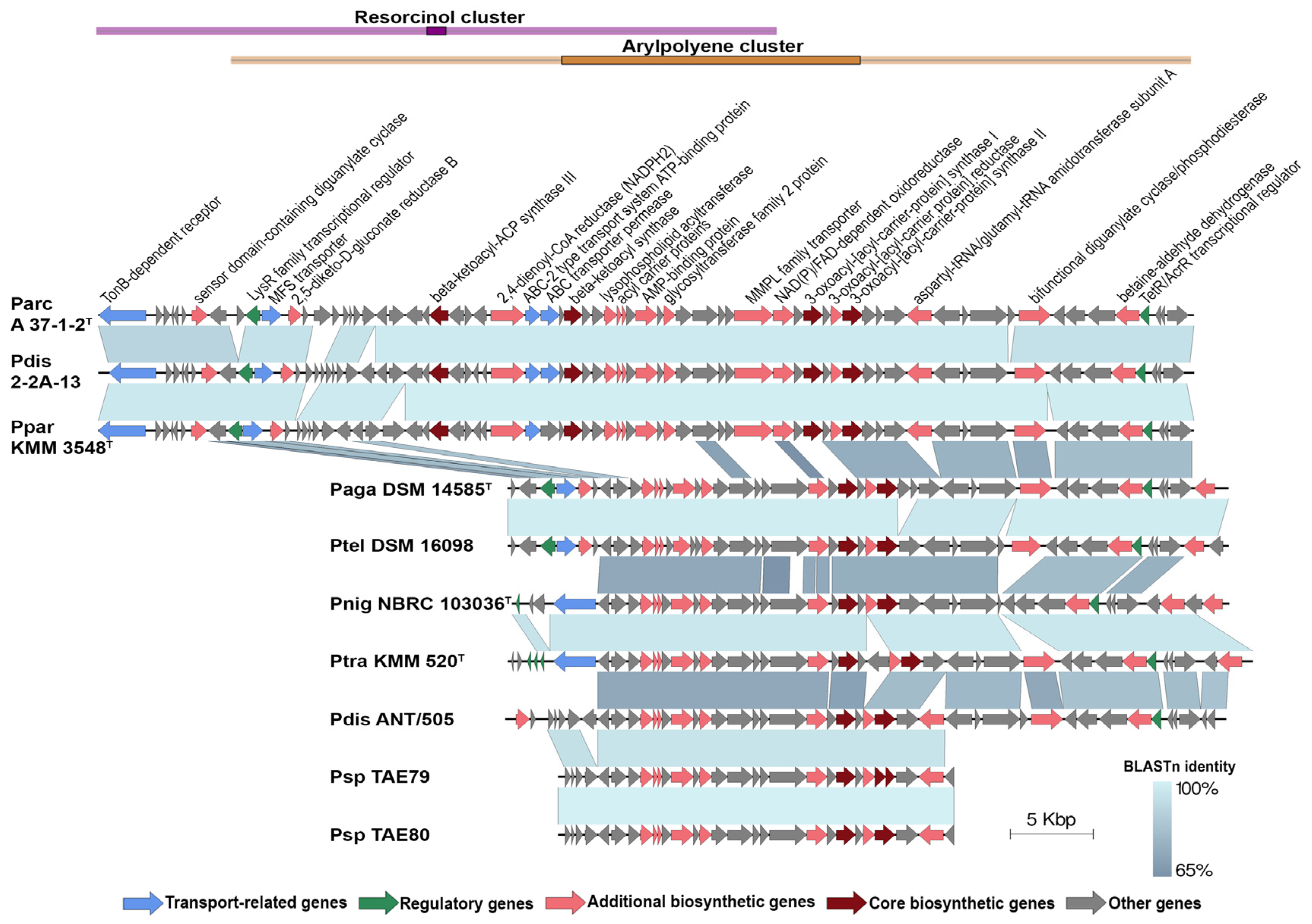
2.5. Secondary Metabolite Biosynthetic Gene Clusters Analysis of the P. distincta Strains
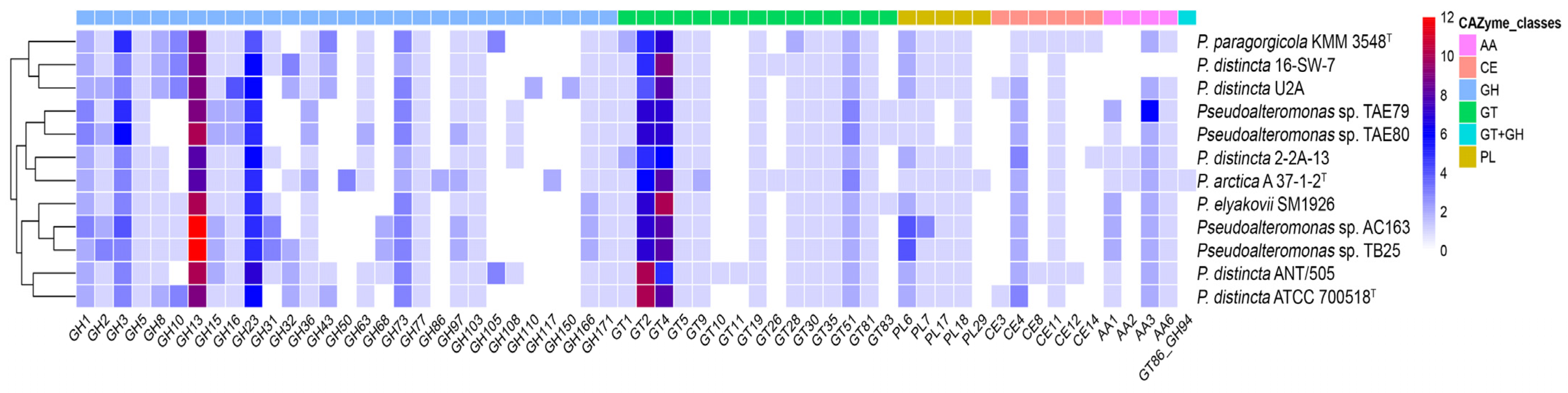
2.6. Carbohydrate-Active Enzymes of the P. distincta Strains
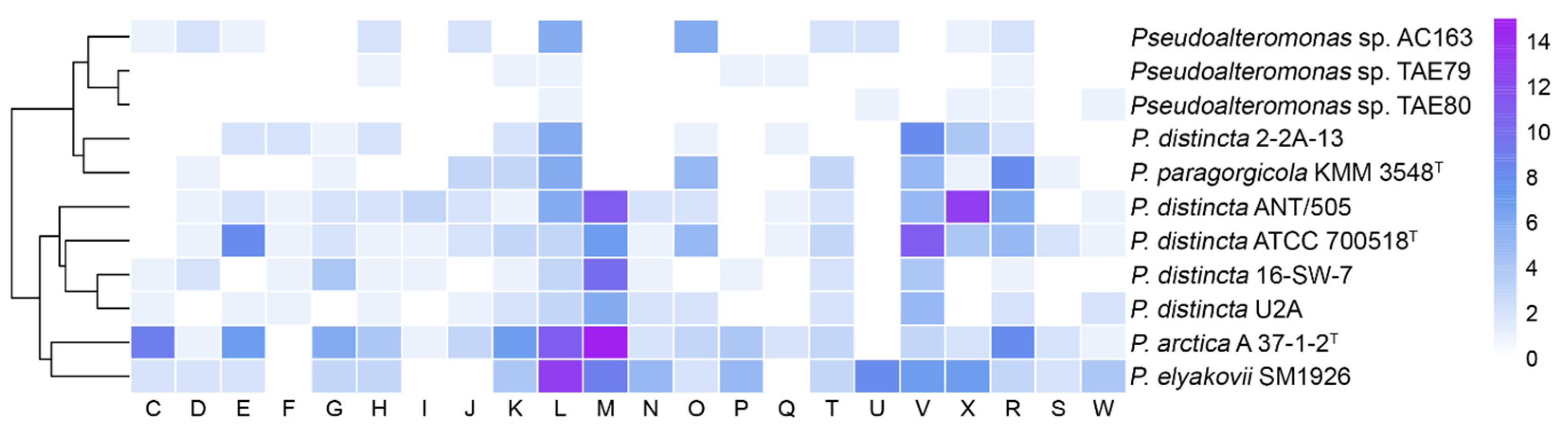
2.7. Unique Genes and Genomic Islands Analysis of the P. disticta Strains
2.8. Metabolic Pathways Analysis of the P. distincta Strains
2.9. Emended Description of the Species Pseudoalteromonas distincta (Romanenko et al., 1995; Ivanova et al., 2000; Nedashkovskaya et al., 2022)
3. Material and Methods
3.1. Bacterial Strains for Phenotype Characterization
3.2. Bacterial Strains for Genome Characterization
3.3. Whole-Genome Sequencing and Assembly
3.4. Whole-Genome, Core-Genome, and Pangenome Phylogeny
3.5. Comparative Analysis of CAZy Families and Biosynthesis Gene Clusters
3.6. Genomic Island Analysis
3.7. Reconstruction of Metabolic Pathways
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nedashkovkaya, O.I.; Kim, S.-G.; Balabanova, L.A.; Zhukova, N.V.; Son, O.M.; Tekutyeva, L.A.; Mikhailov, V.V. Genome-based classification of strain 16-SW-7, a marine bacterium capable of converting B red blood cells, as Pseudoalteromonas distincta and proposal to reclassify Pseudoalteromonas paragorgicola as a later heterotypic synonym of Pseudoalteromonas distincta. Front. Microbiol. 2022, 12, 4325. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. J. Syst. Evol. Microbiol. 2017, 67, 1613. [Google Scholar] [CrossRef]
- Ivanova, E.P.; Chun, J.; Romanenko, L.A.; Matte, M.E.; Mikhailov, V.V.; Frolova, G.M. Reclassification of Alteromonas distincta Romanenko et al. 1995 as Pseudoalteromonas distincta comb. nov. Int. J. Syst. Evol. Microbiol. 2000, 50, 141–144. [Google Scholar] [CrossRef]
- Bosi, E.; Fondi, M.; Orlandini, V.; Perrin, E.; Maida, I.; de Pascale, D.; Fani, R. The pangenome of (Antarctic) Pseudoalteromonas bacteria: Evolutionary and functional insights. BMC Genom. 2017, 18, 93. [Google Scholar] [CrossRef] [Green Version]
- Hettle, A.G.; Hobbs, J.K.; Pluvinage, B.; Vickers, C.; Abe, K.T.; Salama-Alber, O.; McGuire, B.E.; Hehemann, J.-H.; Hui, J.P.M.; Berrue, F.; et al. Insights into the κ/ι-carrageenan metabolism pathway of some marine Pseudoalteromonas species. Commun. Biol. 2019, 2, 474. [Google Scholar] [CrossRef] [Green Version]
- Parrilli, E.; Tedesco, P.; Fondi, M.; Tutino, M.L.; Giudice, A.L.; de Pascale, D.; Fani, R. The art of adapting to extreme environments: The model system Pseudoalteromonas. Phys. Life Rev. 2021, 36, 137–161. [Google Scholar] [CrossRef]
- Chau, R.; Pearson, L.A.; Cain, J.; Kalaitzis, J.A.; Neilan, B.A. A Pseudoalteromonas Clade with remarkable biosynthetic potential. Appl. Environ. Microbiol. 2021, 87, e02604-20. [Google Scholar] [CrossRef]
- El-Naggar, N.E.; El-Ewasy, S.M. Bioproduction, characterization, anticancer and antioxidant activities of extracellular melanin pigment produced by newly isolated microbial cell factories Streptomyces glaucescens NEAE-H. Sci. Rep. 2017, 14, 42129. [Google Scholar] [CrossRef]
- Celedón, R.S.; Díaz, L.B. Natural pigments of bacterial origin and their possible biomedical applications. Microorganisms 2021, 9, 739. [Google Scholar] [CrossRef]
- Takemura, M.; Takagi, C.; Aikawa, M.; Araki, R.; Choi, S.-K.; Itaya, M.; Shindo, K.; Misawa, N. Heterologous production of novel and rare C30-carotenoids using Planococcus carotenoid biosynthesis genes. Microb. Cell Fact. 2021, 20, 194. [Google Scholar] [CrossRef]
- Siziya, I.N.; Hwang, C.Y.; Seo, M.-J. Antioxidant potential and capacity of microorganism-sourced C30 carotenoids—A review. Antioxidants 2022, 11, 1963. [Google Scholar] [CrossRef]
- Metwally, R.A.; El-Sersy, N.A.; El Sikaily, A.; Soraya, A.; Sabry, S.A.; Ghozlan, H.A. Optimization and multiple in vitro activity potentials of carotenoids from marine Kocuria sp. RAM1. Sci. Rep. 2022, 12, 18203. [Google Scholar] [CrossRef]
- Kim, S.H.; Lee, P.C. Functional expression and extension of staphylococcal staphyloxanthin biosynthetic pathway in Escherichia coli. J. Biol. Chem. 2012, 22, 21575–21583. [Google Scholar] [CrossRef] [Green Version]
- de Carvalho, C.C.C.R.; Caramujo, M.J. Carotenoids in aquatic ecosystems and aquaculture: A colorful business with implications for human health. Front. Mar. Sci. 2017, 4, 93. [Google Scholar] [CrossRef] [Green Version]
- Liang, M.H.; Zhu, J.; Jiang, J.G. Carotenoids biosynthesis and cleavage related genes from bacteria to plants. Crit. Rev. Food Sci. Nutr. 2018, 58, 2314–2333. [Google Scholar] [CrossRef]
- Rodriguez-Concepcion, M.; Avalos, J.; Bonet, M.L.; Boronat, A.; Gomez-Gomez, L.; Hornero-Mendez, D.; Limon, M.C.; Meléndez-Martínez, A.J.; Olmedilla-Alonso, B.; Palou, A.; et al. A global perspective on carotenoids: Metabolism, biotechnology, and benefits for nutrition and health. Prog. Lipid Res. 2018, 70, 62–93. [Google Scholar] [CrossRef] [Green Version]
- Maoka, T. Carotenoids as natural functional pigments. J. Nat. Med. 2020, 74, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, K.; Buys, E.M. Insights into the role of bacteria in vitamin A biosynthesis: Future research opportunities. Crit. Rev. Food Sci. Nutr. 2019, 59, 3211–3226. [Google Scholar] [CrossRef]
- Bohn, T. Carotenoids, Chronic Disease Prevention and Dietary Recommendations. Int. J. Vitam. Nutr. Res. 2017, 87, 121–130. [Google Scholar] [CrossRef]
- Zimmermann, J.; Kaleta, C.; Waschina, S. gapseq: Informed prediction of bacterial metabolic pathways and reconstruction of accurate metabolic models. Genome Biol. 2021, 22, 81. [Google Scholar] [CrossRef]
- Schöner, T.A.; Gassel, S.; Osawa, A.; Tobias, N.J.; Okuno, Y.; Sakakibara, Y.; Bode, H.B. Aryl polyenes, a highly abundant class of bacterial natural products, are functionally related to antioxidative carotenoids. Chem. Bio Chem. 2016, 17, 247–253. [Google Scholar] [CrossRef]
- Grammbitter, G.L.C.; Schmalhofer, M.; Karimi, R.; Schöner, T.A.; Tobias, N.J.; Morgner, N.; Groll, M.; Bode, H.B. An uncommon type II PKS catalyzes biosynthesis of aryl polyene pigments. Am. Chem. Soc. 2019, 141, 16615–16623. [Google Scholar] [CrossRef]
- Johnston, I.; Osborn, L.J.; Markley, R.L.; McManus, E.A.; Kadam, A.; Schultz, K.B.; Nagajothi, N.; Ahern, P.P.; Brown, J.M.; Claesen, J. Identification of essential genes for Escherichia coli aryl polyene biosynthesis and function in biofilm formation. NPJ Biofilms Microbiom. 2021, 7, 56. [Google Scholar] [CrossRef]
- He, Y.-W.; Cao, X.-Q.; Poplawsky, A.R. Chemical structure, biological roles, biosynthesis and regulation of the yellow xanthomonadin pigments in the phytopathogenic genus Xanthomonas. Mol. Plant Microbe Interact. 2020, 33, 705–714. [Google Scholar] [CrossRef] [Green Version]
- Grammbitter, G.L.C.; Shi, Y.-M.; Shi, Y.N.; Sahithya, Y.; Vemulapalli, P.B.; Richter, C.; Schwalbe, H.; Alanjary, M.; Schüffler, A.; Witt, M.; et al. The Chemical structure of widespread microbial aryl polyene lipids. bioRxiv 2020. [Google Scholar] [CrossRef]
- Tran-Ly, A.N.; Reyes, C.; Schwarze, F.W.M.R.; Ribera, J. Microbial production of melanin and its various applications. World J. Microbiol. Biotechnol. 2020, 36, 170. [Google Scholar] [CrossRef]
- Ram, S.; Mitra, M.; Shah, F.; Tirkey, S.R.; Mishra, S. Bacteria as an alternate biofactory for carotenoid production: A review of its applications, opportunities and challenges. J. Funct. Foods 2020, 67, 103867. [Google Scholar] [CrossRef]
- Halder, U.; Banerjee, A.; Bandopadhyay, R. Structural and functional properties, biosynthesis, and patenting trends of bacterial succinoglycan: A review. Indian J. Microbiol. 2017, 57, 278–284. [Google Scholar] [CrossRef]
- Bäumgen, M.; Dutschei, T.; Bornscheuer, U.T. Marine polysaccharides: Occurrence, enzymatic degradation and utilization. Chem. Bio Chem. 2021, 22, 2247–2256. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, J.; Wei, F.; Li, W.; Wen, L. Facile synthesis of sugar nucleotides from common sugars by the cascade conversion strategy. Am. Chem. Soc. 2022, 144, 9980–9989. [Google Scholar] [CrossRef]
- Romanenko, L.A.; Mikhailov, V.V.; Lysenko, A.M.; Stepanenko, V.I. A new species of melanin-producing bacteria of the genus Alteromonas. Mikrobiologiya 1995, 64, 74–77. [Google Scholar]
- Al Khudary, R.; Stößer, N.I.; Qoura, F.; Antranikian, G. Pseudoalteromonas arctica sp. nov., an aerobic, psychrotolerant, marine bacterium isolated from Spitzbergen. Int. J. Syst. Evol. Microbiol. 2008, 58, 2018–2024. [Google Scholar] [CrossRef]
- Sawabe, T.; Tanaka, R.; Iqbal, M.M.; Tajima, K.; Ezura, Y.; Ivanova, E.P.; Christen, R. Assignment of Alteromonas elyakovii KMM 162T and five strains isolated from spot-wounded fronds of Laminaria japonica to Pseudoalteromonas elyakovii comb. nov. and the extended description of the species. Int. J. Syst. Evol. Microbiol. 2000, 50, 265–271. [Google Scholar] [CrossRef]
- Asnicar, F.; Thomas, A.M.; Beghini, F.; Mengoni, C.; Manara, S.; Manghi, P.; Zhu, Q.; Bolzan, M.; Cumbo, F.; May, U.; et al. Precise phylogenetic analysis of microbial isolates and genomes from meta-genomes using PhyloPhlAn 3.0. Nat. Commun. 2020, 11, 2500. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [Green Version]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [Green Version]
- Matsuyama, H.; Sawazaki, K.; Minami, H.; Kasahara, H.; Horikawa, K.; Yumoto, I. Pseudoalteromonas shioyasakiensis sp. nov., a marine polysaccharide-producing bacterium. Int. J. Syst. Evol. Microbiol. 2014, 64, 101–106. [Google Scholar] [CrossRef] [Green Version]
- Okie, J.G.; Poret-Peterson, A.T.; Lee, Z.M.P.; Richter, A.; Alcaraz, L.D.; Eguiarte, L.E.; Iefert, J.L.; Souza, V.; Dupont, C.L.; Elser, J.J. Genomic adaptations in information processing underpin trophic strategy in a whole-ecosystem nutrient enrichment experiment. eLife 2020, 9, e49816. [Google Scholar] [CrossRef]
- Wang, X.; Uematsu, K.; O’Rourke, A.; Akita, T.; Kimura, K.; Tomaru, Y.; Voolstra, C.R.; Innan, H.H. A novel bacterial speciation process observed in a symbiotic marine population. bioRxiv 2022. [Google Scholar] [CrossRef]
- Vázquez-Rosas-Landa, M.; Ponce-Soto, G.Y.; Aguirre-Liguori, J.A.; Thakur, S.; Scheinvar, E.; Barrera-Redondo, J.; Ibarra-Laclette, E.; Guttman, D.S.; Luis, E.; Eguiarte, L.E.; et al. Population genomics of Vibrionaceae isolated from an endangered oasis reveals local adaptation after an environmental perturbation. BMC Genom. 2020, 21, 418. [Google Scholar] [CrossRef]
- Feng, X.; Bello, D.; O’Hagan, D. Isolation of 5’-O-sulfamyladenosine and related 3’-O-β-glucosylated adenosines from the nucleocidin producer Streptomyces calvus. RSC Adv. 2021, 28, 5291–5294. [Google Scholar] [CrossRef]
- Botas, A.; Eitel, M.; Schwarz, P.N.; Buchmann, A.; Costales, P.; Núñez, L.E.; Cortés, J.; Morís, F.; Krawiec, M.; Wolański, M.; et al. Genetic engineering in combination with semi-synthesis leads to a new route for gram-scale production of the immunosuppressive natural product Brasilicardin A. Angew. Chem. Int. Ed. 2021, 60, 13536. [Google Scholar] [CrossRef]
- Chattopadhyay, M.K. Mechanism of bacterial adaptation to low temperature. J. Biosci. 2006, 31, 157–165. [Google Scholar] [CrossRef]
- Schöner, T.A.; Fuchs, S.W.; Reinhold-Hurek, B.; Bode, H.B. Identification and biosynthesis of a novel Xanthomonadin-Dialkylresorcinol-Hybrid from Azoarcus sp. BH72. PLoS ONE 2014, 9, e90922. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.M.; Hirschmann, M.; Shi, Y.N.; Ahmed, S.; Abebew, D.; Tobias, N.J.; Grün, P.; Crames, J.J.; Pöschel, L.; Kuttenlochner, W.; et al. Global analysis of biosynthetic gene clusters reveals conserved and unique natural products in entomopathogenic nematode-symbiotic bacteria. Nat. Chem. 2022, 14, 701–712. [Google Scholar] [CrossRef]
- The CAZypedia Consortium, Ten years of CAZypedia: A living encyclopedia of carbohydrate-active enzymes. Glycobiology 2018, 28, 3–8. [CrossRef] [Green Version]
- Li, Q.; Jiang, Y.; Tong, X.; Zhao, L.; Pei, J. Co-production of xylooligosaccharides and xylose from poplar sawdust by recombinant endo-1,4-β-xylanase and β-xylosidase mixture hydrolysis. Front. Bioeng. Biotechnol. 2021, 8, 637397. [Google Scholar] [CrossRef]
- Khorshidian, N.; Khanniri, E.; Koushki, M.R.; Sohrabvandi, S.; Yousefi, M. An overview of antimicrobial activity of lysozyme and its functionality in cheese. Front. Nutr. 2022, 9, 833618. [Google Scholar] [CrossRef]
- Munoz-Munoz, J.; Cartmell, A.; Terrapon, N.; Baslé, A.; Henrissat, B.; Gilbert, H.J. An evolutionarily distinct family of polysaccharide lyases removes rhamnose capping of complex arabinogalactan proteins. J. Biol. Chem. 2017, 11, 13271–13283. [Google Scholar] [CrossRef] [Green Version]
- Koch, H.; Germscheid, N.; Freese, H.M.; Noriega-Ortega, B.; Lücking, D.; Berger, M.; Qiu, G.M.; Marzinelli, E.M.; Campbell, A.H.; Steinberg, P.D.; et al. Genomic, metabolic and phenotypic variability shapes ecological differentiation and intraspecies interactions of Alteromonas macleodii. Sci. Rep. 2020, 10, 809. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Song, Y.; Feng, X.; Tang, K.; Jiao, N.; Tian, J.; Zhang, Y. Genomic characteristics and potential metabolic adaptations of hadal trench roseobacter and alteromonas bacteria based on single-cell genomics analyses. Front. Microbiol. 2020, 11, 1739. [Google Scholar] [CrossRef]
- Bertelli, C.; Laird, M.R.; Williams, K.P.; Simon Fraser University Research Computing Group; Lau, B.Y.; Hoad, G.; Winsor, G.L.; Brinkman, F.S.L. IslandViewer 4: Expanded prediction of genomic islands for larger-scale datasets. Nucleic Acids Res. 2017, 45, W30–W35. [Google Scholar] [CrossRef]
- da Silva Filho, A.C.; Raittz, R.T.; Guizelini, D.; De Pierri, C.R.; Augusto, D.W.; dos Santos-Weiss, I.C.R.; Marchaukoski, J.N. Comparative analysis of genomic island prediction tools. Front. Genet. 2018, 9, 619. [Google Scholar] [CrossRef] [Green Version]
- Norsigiana, C.J.; Danhofb, H.A.; Brandb, C.K.; Midanib, F.S.; Broddrickd, J.T.; Savidgee, T.C.; Brittonb, R.A.; Palssona, B.O.; Spinlerd, J.K.; Jonathan, M.; et al. Systems biology approach to functionally assess the Clostridioides difficile pangenome reveals genetic diversity with discriminatory power. Proc. Natl. Acad. Sci. USA 2022, 119, e2119396119. [Google Scholar] [CrossRef]
- Veaudor, T.; Cassier-Chauvat, C.; Chauvat, F. Genomics of urea transport and catabolism in cyanobacteria: Biotechnological implications. Front. Microbiol. 2019, 10, 2052. [Google Scholar] [CrossRef] [Green Version]
- Chidwick, H.S.; Flack, E.K.P.; Keenan, T.; Walton, J.; Thomas, G.T.; Fascione, M.A. Reconstitution and optimisation of the biosynthesis of bacterial sugar pseudaminic acid (Pse5Ac7Ac) enables preparative enzymatic synthesis of CMP-Pse5Ac7Ac. Sci. Rep. 2021, 11, 4756. [Google Scholar] [CrossRef]
- Wang, J.; Greenway, H.; Li, S.; Wei, M.; Polizzi, S.J.; Wang, P.G. Facile and stereo-selective synthesis of UDP-α-D-xylose and UDP-β-L-arabinose using UDP-sugar pyrophosphorylase. Front. Chem. 2018, 6, 163. [Google Scholar] [CrossRef]
- Casillo, A.; Di Guida, R.; Carillo, S.; Chen, C.; Kamasaka, K.; Kawamoto, J.; Kurihara, T.; Corsaro, M.M. Structural elucidation of a novel lipooligosaccharide from the antarctic bacterium OMVs producer Shewanella sp. HM13. Mar. Drugs 2019, 17, 34. [Google Scholar] [CrossRef] [Green Version]
- Perret, A.; Lechaplais, C.; Tricot, S.; Perchat, N.; Vergne, C.; Pellé, C.; Bastard, K.; Kreimeyer, A.; Vallenet, D.; Zaparucha, A.; et al. A novel acyl-CoA beta-transaminase characterized from a metagenome. PLoS ONE 2011, 6, e22918. [Google Scholar] [CrossRef] [Green Version]
- Koedooder, C.; Guéneuguès, A.; Van Geersdaële, R.; Vergé, V.; Bouget, F.-Y.; Labreuche, Y.; Obernosterer, I.; Blain, S. The role of the glyoxylate shunt in the acclimation to iron limitation in marine heterotrophic bacteria. Front. Mar. Sci. 2018, 5, 435. [Google Scholar] [CrossRef] [Green Version]
- Pavoncello, V.; Barras, F.; Bouveret, E. Degradation of exogenous fatty acids in Escherichia coli. Biomolecules 2022, 12, 1019. [Google Scholar] [CrossRef]
- Kaval, K.G.; Garsin, D.A. Ethanolamine utilization in bacteria. mBio 2018, 20, e00066-18. [Google Scholar] [CrossRef] [Green Version]
- Chai Hao Chiu, C.H.; Paszkowski, U. Mechanisms and impact of symbiotic phosphate acquisition. Cold Spring Harb. Perspect. Biol. 2019, 11, a034603. [Google Scholar] [CrossRef]
- Tenconi, E.; Traxler, M.F.; Hoebreck, C.; van Wezel, G.P.; Rigali, S. Production of prodiginines is part of a programmed cell death process in Streptomyces coelicolor. Front. Microbiol. 2018, 9, 1742. [Google Scholar] [CrossRef]
- van der Beek, S.L.; Zorzoli, A.; Çanak, E.; Chapman, R.N.; Lucas, K.; Meyer, B.H.; Evangelopoulos, D.; de Carvalho, L.P.S.; Boons, G.J.; Dorfmueller, H.C.; et al. Streptococcal dTDP-L-rhamnose biosynthesis enzymes: Functional characterization and lead compound identification. Mol. Microbiol. 2019, 111, 951–964. [Google Scholar] [CrossRef] [Green Version]
- Mikkola, S. Nucleotide sugars in chemistry and biology. Molecules 2020, 25, 5755. [Google Scholar] [CrossRef]
- Balabanova, L.; Averianova, L.; Marchenok, M.; Son, O.; Tekutyeva, L. Microbial and genetic resources for cobalamin (vitamin B12) biosynthesis: From ecosystems to industrial biotechnology. Int. J. Mol. Sci. 2021, 22, 4522. [Google Scholar] [CrossRef]
- Wang, L.; Yan, J.; Wise, M.J.; Liu, Q.; Asenso, J.; Huang, Y.; Dai, S.; Liu, Z.; Du, Y.; Tang, D. Distribution patterns of polyphosphate metabolism pathway and its relationships with bacterial durability and virulence. Front. Microbiol. 2018, 9, 782. [Google Scholar] [CrossRef] [Green Version]
- Wood, J.M.; Chavan, B.; Hafeez, I.; Schallreuter, K.U. Regulation of tyrosinase by tetrahydropteridines and H2O2. Biochem. Biophys. Res. Commun. 2004, 325, 1412–1417. [Google Scholar] [CrossRef]
- Buglak, A.A.; Telegina, T.A.; Vechtomova, Y.L.; Kritsky, M.S. Autoxidation and photooxidation of tetrahydrobiopterin: A theoretical study. Free. Radic. Res. 2021, 55, 499–509. [Google Scholar] [CrossRef]
- Fast, Q.C. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 28 December 2018).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [Green Version]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-R, L.M.; Konstantinidis, K.T. The enveomics collection: A toolbox for specialized analyses of microbial genomes andmetagenomes. Peer J. Prepr. 2016, 4, e1900v1. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Göker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [Green Version]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Yohe, T.; Huang, L.; Entwistle, S.; Wu, P.; Yang, Z.; Busk, P.K.; Xu, Y.; Yin, Y. dbCAN2: A meta server for automatedcarbohydrate-active enzyme annotation. Nucleic Acids Res. 2018, 46, W95–W101. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimatingmaximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. Modelfinder: Fast model selection for accuratephylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Dong, Z.; Fang, L.; Luo, Y.; Wei, Z.; Guo, H.; Zhang, G.; Gu, Y.Q.; Coleman-Derr, D.; Xia, Q.; et al. OrthoVenn2: A web server for whole-genome comparison and annotation of orthologous clusters acrossmultiple species. Nucleic Acids Res. 2019, 47, W52–W58. [Google Scholar] [CrossRef] [Green Version]
- Bauer, E.; Zimmermann, J.; Baldini, F.; Thiele, I.; Kaleta, C. BacArena: Individual-based metabolic modeling of heterogeneous microbes in complex communities. PLoS Comput. Biol. 2017, 13, e1005544. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef] [Green Version]
- Dowle, M.; Srinivasan, A. Data.table: Extension of ‘data.frame’. 2021. R Package Version 1.14.0. Available online: https://CRAN.R-project.org/package=data.table (accessed on 10 November 2022).
- Wickham, H. Stringr: Simple, Consistent Wrappers for Common String Operations. R Package Version 1.4.0. 2019. Available online: https://CRAN.R-project.org/package=stringr (accessed on 10 November 2022).
- Gelius-Dietrich, G.; Desouki, A.A.; Fritzemeier, C.J.; Lercher, M.J. Sybil–efficient constraint-based modelling in R. BMC Syst. Biol. 2013, 7, 125. [Google Scholar] [CrossRef] [Green Version]
- Davis, T.L.; Day, A. Getopt: C-like ‘Getopt’ Behavior. R Package Version 1.20.3. 2019. Available online: https://CRAN.R-project.org/package=getopt (accessed on 10 November 2022).
- Wickham, H. Reshaping data with the reshape package. J. Stat. Softw. 2007, 21, 1–20. [Google Scholar] [CrossRef]
- Corporation, M.; Weston, S. doParallel: Foreach Parallel Adaptor for the ‘Parallel’ Package. R Package Version 1.0.16. 2020. Available online: https://CRAN.R-project.org/package=doParallel (accessed on 10 November 2022).
- Weston, S. Foreach: Provides Foreach Looping Construct. R Package Version 1.5.1. 2019. Available online: https://CRAN.R-project.org/package=foreach (accessed on 15 November 2022).
- Bengtsson, H.R. utils: Various Programming Utilities. R Package Version 2.10.1. 2019. Available online: https://CRAN.R-project.org/package=R.utils (accessed on 15 November 2022).
- Gagolewski, M. R Package Stringi: Character String Processing Facilities. 2020. Available online: http://www.gagolewski.com/software/stringi/ (accessed on 10 November 2022).
- Gelius-Dietrich, G. glpkAPI: R Interface to C API of GLPK. R Package Version 1.3.2. 2020. Available online: https://CRAN.R-project.org/package=glpkAPI (accessed on 10 November 2022).
- Pagès, H.; Aboyoun, P.; Gentleman, R.; DebRoy, S. Biostrings: Efficient Manipulation of Biological Strings, R package version 2.54.0; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Bornstein, B.J.; Keating, S.M.; Jouraku, A.; Hucka, M. LibSBML: An API library for SBML. Bioinformatics 2008, 24, 880–881. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem in 2021: New data content and improved web interfaces. Nucleic Acids Res. 2021, 49, D1388–D1395. [Google Scholar] [CrossRef]
- Karp, P.D.; Billington, R.; Caspi, R.; Fulcher, C.A.; Latendresse, M.; Kothari, A.; Keseler, I.M.; Krummenacker, M.; Midford, P.E.; Ong, Q.; et al. The BioCyc collection of microbial genomes and metabolic pathways. Brief. Bioinform. 2019, 20, 1085–1093. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic * | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|
| Source of isolation | Sponge | Seawater | Gorgonian | Starfish | Seawater |
| Colony color | whitish | whitish | Slightly orange | Slightly orange | Slightly orange |
| Temperature range for growth (°C) | 4–30 | 4–34 | 4–30 | 4–37 | 4–25 |
| Salinity range for growth (% NaCl) | 1–6 | 1–10 | 0.5–8 | 0.5–10 | 0–9 |
| Required seawater or artificial seawater for growth | + | − | − | − | − |
| Production of melanin-like pigments | + | − | − | − | − |
| Acetoin production | − | − | − | w | − |
| H2S production | + | − | + | + | − |
| Hydrolysis of: | |||||
| Casein | − | w | w | + | + |
| Gelatin | + | + | + | + | − |
| Starch | − | − | + | + | − |
| Acid production from: | |||||
| Cellobiose, maltose | − | + | + | + | + |
| Galactose | − | + | + | − | + |
| Glucose, lactose, xylose | − | + | + | − | − |
| Raffinose | − | + | − | − | − |
| Mannitol | + | + | − | + | + |
| Utilization of citrate | − | + | w | w | + |
| Assimilation of: | |||||
| D-Glucose, sucrose, sodium acetate, L-alanine, L-serine, capric acid | + | + | + | − | + |
| Glycogen | + | + | + | − | − |
| Sodium citrate, L-proline, propionic acid, valeric acid | + | + | + | − | ND |
| Potashium-2-keto-gluconate, salicin | − | + | − | − | − |
| Itaconic acid, L-histidine | − | + | − | − | ND |
| D-Melibiose | − | + | + | − | + |
| D-Sorbitol | − | + | + | − | − |
| Inositol, lactic acid, ribose | + | − | − | − | − |
| Sodium malonate, 3-hydroxybutiric acid | + | − | − | − | ND |
| Enzyme activity (API ZYM tests): | |||||
| Esterase (C4) | + | + | − | + | ND |
| Cysteine arylamidase | − | + | − | w | ND |
| Trypsin, α-chymotrypsin | − | + | − | − | ND |
| β-Glucosidase, β-galactosidase | − | − | − | − | + |
| Susceptibility to: | |||||
| Ampicillin, vancomycin | + | − | + | + | ND |
| Benzylpenicillin, cephalexin, cefazolin | − | − | − | + | ND |
| Oleandomycin | − | + | + | + | ND |
| Streptomycin | + | + | + | − | ND |
| Tetracycline | − | − | + | + | ND |
| DNA G + C content (mol%) | 39.2 | 39.3 | 39.2 | 39.1 | 39.1 |
| Fatty Acids * | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| Saturated | ||||
| C12:0 | tr | 2.8 | tr | 2.4 |
| C14:0 | tr | 3.9 | 1.3 | 1.1 |
| C15:0 | 4.7 | 2.5 | 3.0 | 2.2 |
| C16:0 | 15.4 | 18.2 | 16.0 | 12.7 |
| C17:0 | 10.3 | 6.1 | 5.8 | 1.5 |
| C18:0 | 1.8 | 1.9 | 1.4 | - |
| Unsaturated | ||||
| C15:1 ω8c | 4.3 | 2.7 | 4.9 | 1.8 |
| C16:1 ω7c | 29.0 | 30.6 | 32.1 | 40.2 |
| C17:1 ω8c | 17.9 | 11.7 | 15.3 | 5.5 |
| C18:1 ω7c | 5.2 | 11.0 | 7.5 | 7.8 |
| Branched | ||||
| iso-C16:0 | tr | 1.0 | tr | - |
| Hydroxy | ||||
| C10:0 3-OH | tr | tr | tr | 1.2 |
| C11:0 3-OH | tr | tr | tr | 1.1 |
| C12:0 3-OH | 6.3 | 4.8 | 7.5 | 6.4 |
| C13:0 3-OH | 1.2 | tr | 1.0 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balabanova, L.; Nedashkovskaya, O.; Otstavnykh, N.; Isaeva, M.; Kolpakova, O.; Pentehina, I.; Seitkalieva, A.; Noskova, Y.; Stepochkina, V.; Son, O.; et al. Computational Insight into Intraspecies Distinctions in Pseudoalteromonas distincta: Carotenoid-like Synthesis Traits and Genomic Heterogeneity. Int. J. Mol. Sci. 2023, 24, 4158. https://doi.org/10.3390/ijms24044158
Balabanova L, Nedashkovskaya O, Otstavnykh N, Isaeva M, Kolpakova O, Pentehina I, Seitkalieva A, Noskova Y, Stepochkina V, Son O, et al. Computational Insight into Intraspecies Distinctions in Pseudoalteromonas distincta: Carotenoid-like Synthesis Traits and Genomic Heterogeneity. International Journal of Molecular Sciences. 2023; 24(4):4158. https://doi.org/10.3390/ijms24044158
Chicago/Turabian StyleBalabanova, Larissa, Olga Nedashkovskaya, Nadezhda Otstavnykh, Marina Isaeva, Oksana Kolpakova, Iuliia Pentehina, Aleksandra Seitkalieva, Yulia Noskova, Varvara Stepochkina, Oksana Son, and et al. 2023. "Computational Insight into Intraspecies Distinctions in Pseudoalteromonas distincta: Carotenoid-like Synthesis Traits and Genomic Heterogeneity" International Journal of Molecular Sciences 24, no. 4: 4158. https://doi.org/10.3390/ijms24044158