
Untargeted Metabolomics Based Prediction of Therapeutic Potential for Apigenin and Chrysin
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Apigenin and Chrysin Treatment Altered the Whole Cell Metabolome in MEF Cells
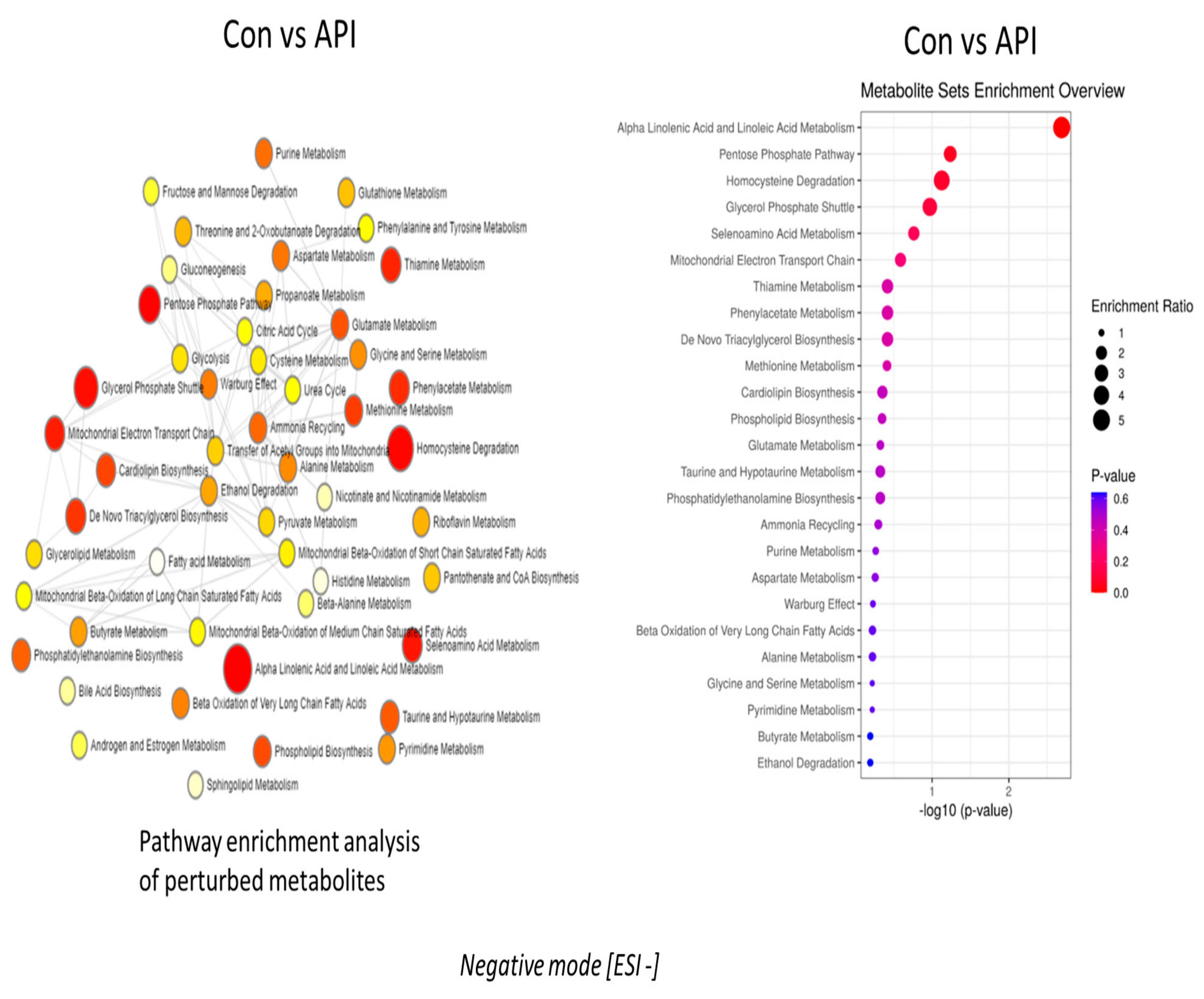
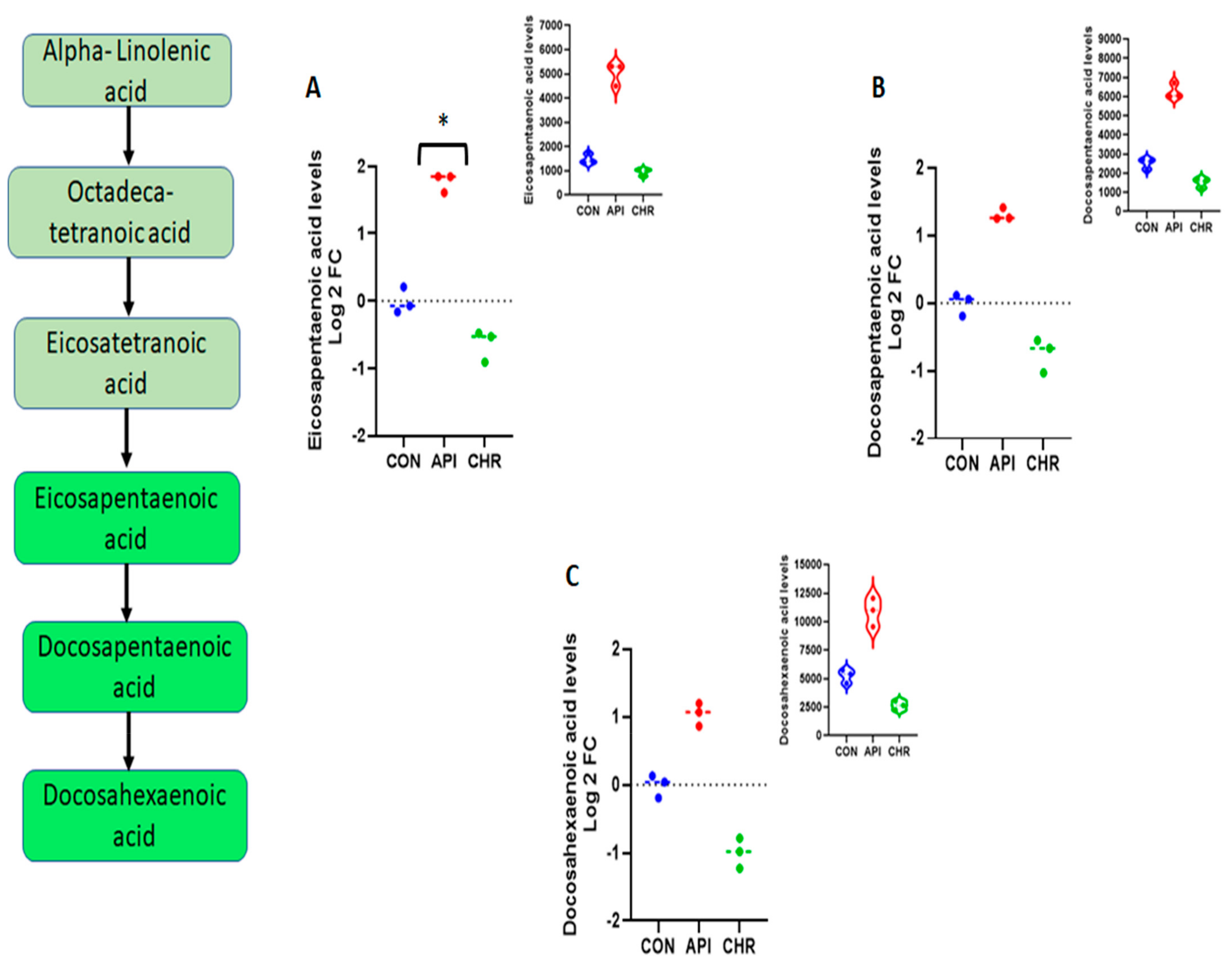
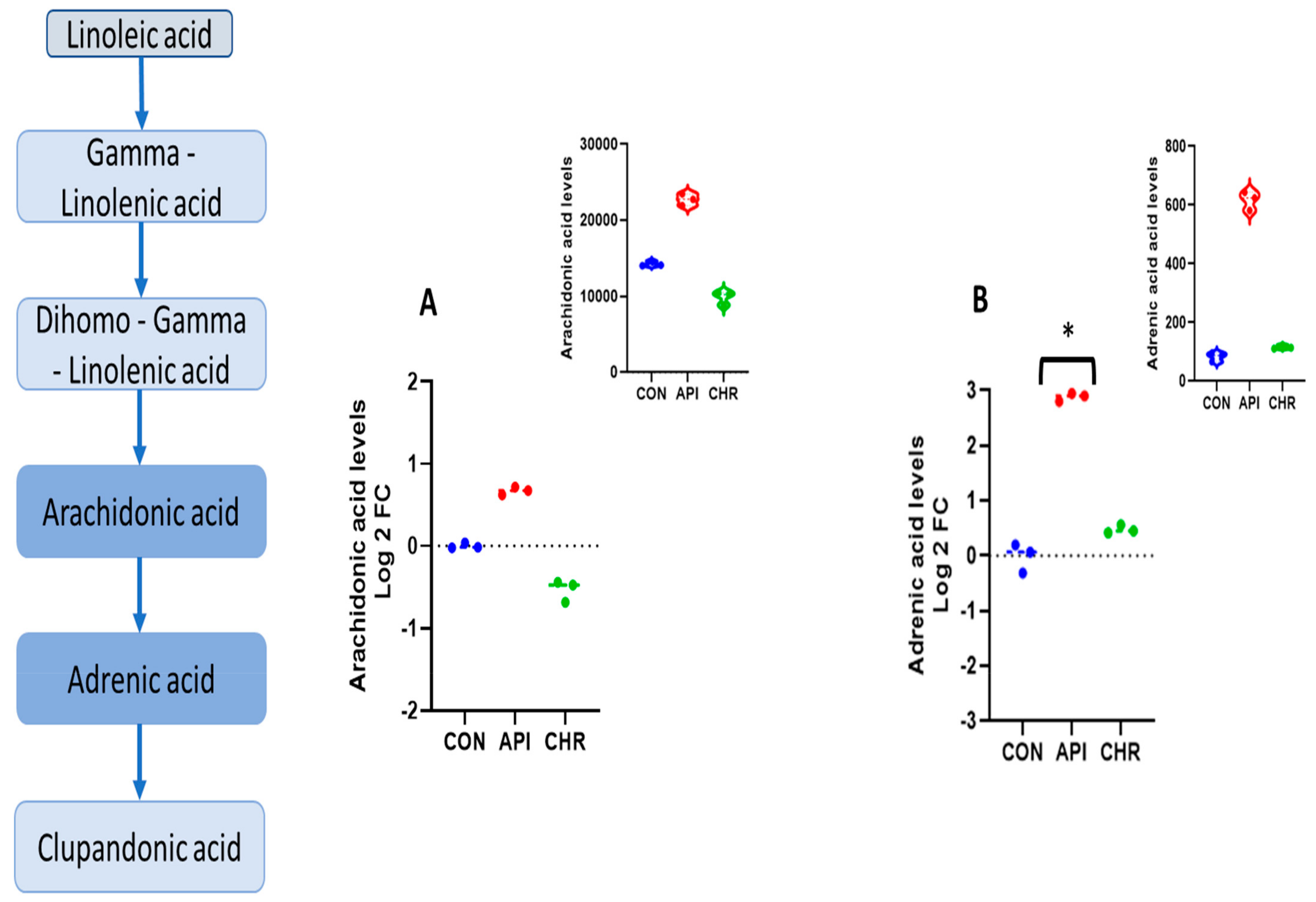
2.2. Alpha-Linolenic Acid and Linoleic Acid Metabolism Emerged as the Major Metabolic Pathways Specifically Regulated by Apigenin
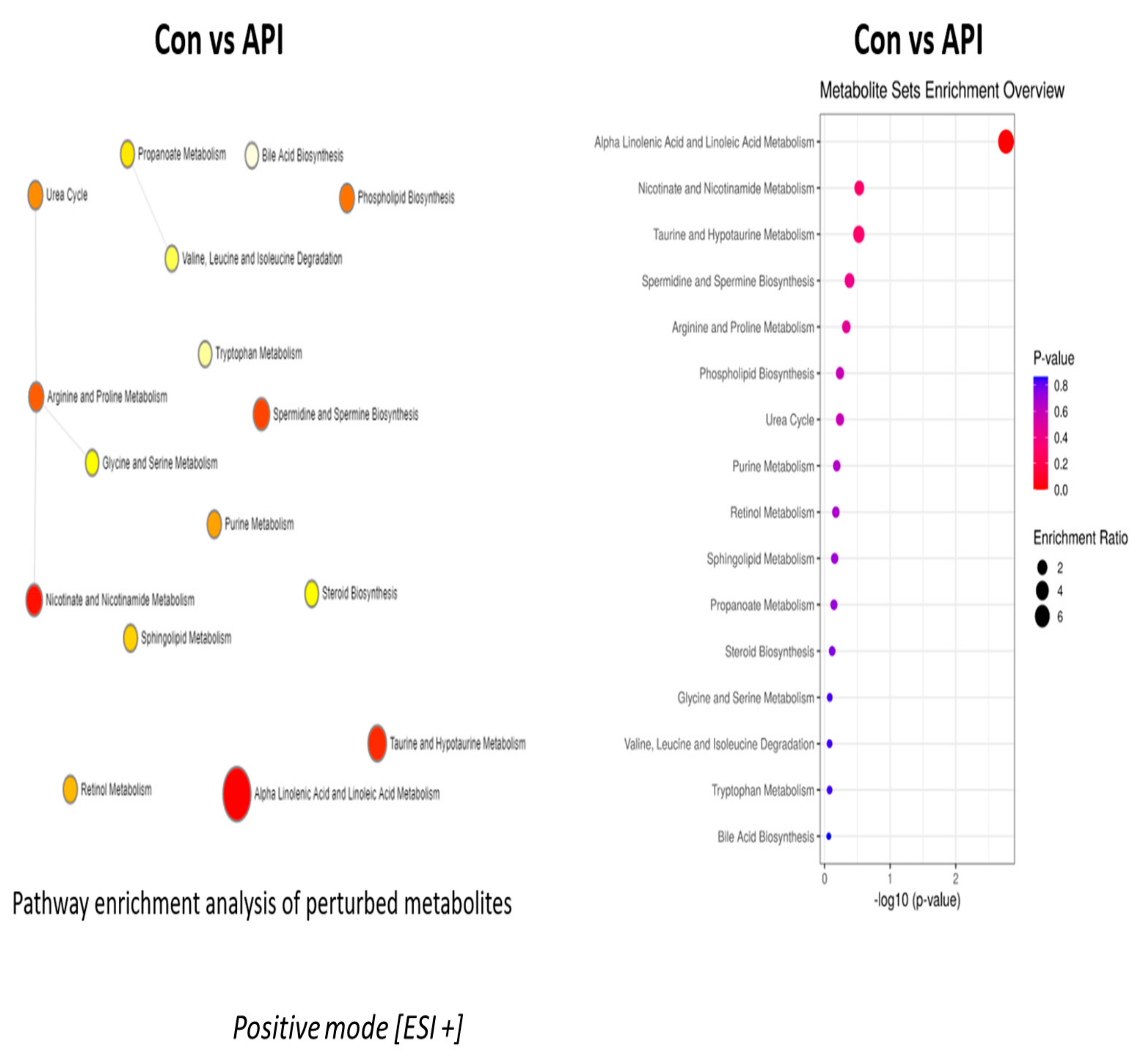
2.3. Alanine Metabolism and Urea Cycle Are the Major Metabolic Pathways Specifically Controlled by Chrysin
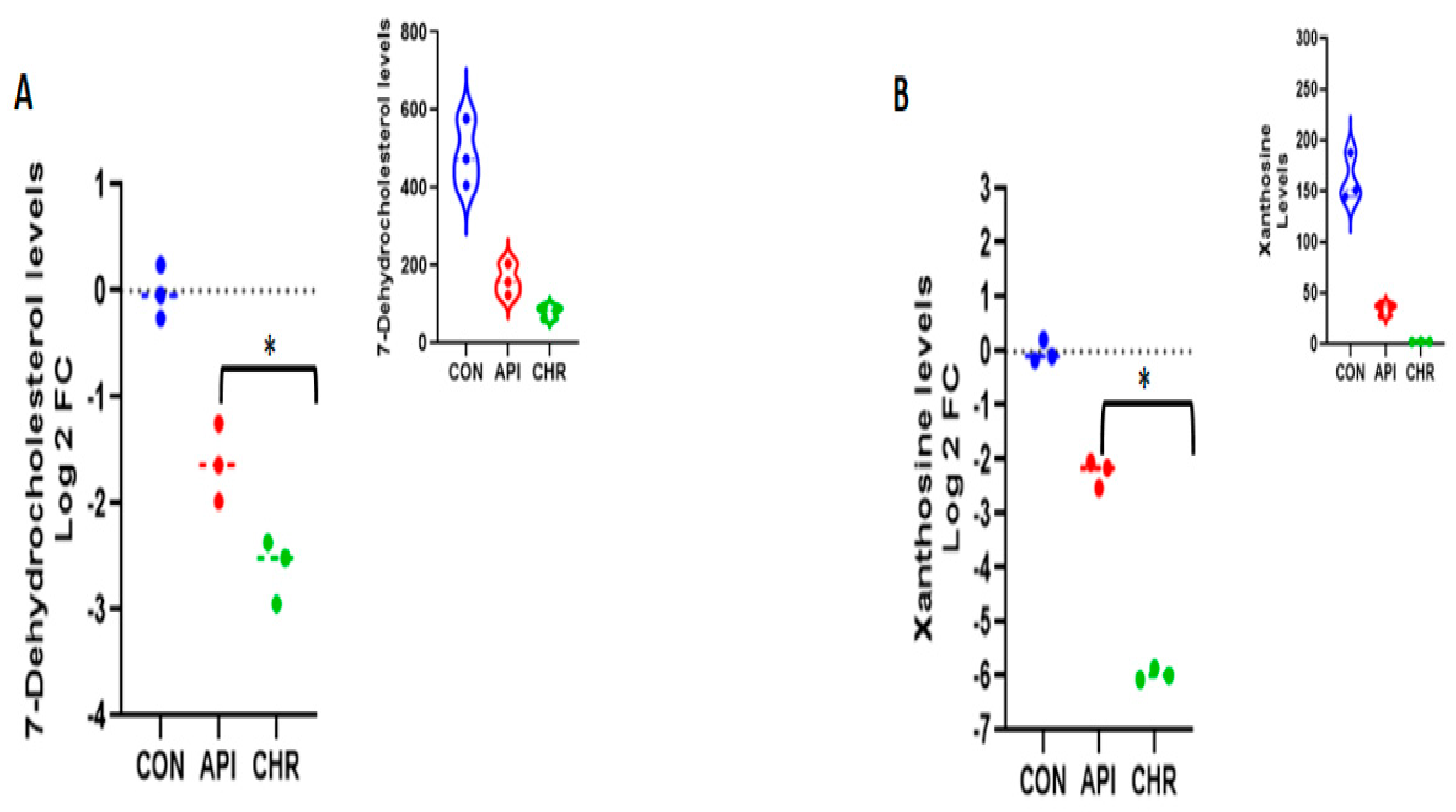
2.4. Apigenin and Chrysin Demonstrated Similarity in Downregulating Metabolites Involved in Cholesterol and Uric Acid Biosynthesis Pathways
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatments
4.2. Sample Preparation
4.3. Chemical Structures
4.4. UPLC-TOF-MS Technology
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, D.R.; Brewster, L.G. The food environment is a complex social network. Soc. Sci. Med. 2015, 133, 202–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, B.V.; Oliveira, C.D.; Lopes, A.C. Food environment of fruits and vegetables in the territory of the Health Academy Program. Cad. Saude Publica 2015, 31 (Suppl. S1), 159–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nesheim, M.; Stover, P.J.; Oria, M. Food systems: Healthy diet sustains the environment too. Nature 2015, 522, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-Barquero, S.; Tresserra-Rimbau, A.; Vitelli-Storelli, F.; Domenech, M.; Salas-Salvado, J.; Martin-Sanchez, V.; Rubin-Garcia, M.; Buil-Cosiales, P.; Corella, D.; Fito, M.; et al. Dietary Polyphenol Intake is Associated with HDL-Cholesterol and A Better Profile of other Components of the Metabolic Syndrome: A PREDIMED-Plus Sub-Study. Nutrients 2020, 12, 689. [Google Scholar] [CrossRef] [Green Version]
- Jantan, I.; Haque, M.A.; Arshad, L.; Harikrishnan, H.; Septama, A.W.; Mohamed-Hussein, Z.A. Dietary polyphenols suppress chronic inflammation by modulation of multiple inflammation-associated cell signaling pathways. J. Nutr. Biochem. 2021, 93, 108634. [Google Scholar] [CrossRef]
- Gentile, D.; Fornai, M.; Pellegrini, C.; Colucci, R.; Blandizzi, C.; Antonioli, L. Dietary flavonoids as a potential intervention to improve redox balance in obesity and related co-morbidities: A review. Nutr. Res. Rev. 2018, 31, 239–247. [Google Scholar] [CrossRef]
- Puthanveetil, P.; Kong, X.; Brase, S.; Voros, G.; Peer, W.A. Transcriptome analysis of two structurally related flavonoids; Apigenin and Chrysin revealed hypocholesterolemic and ketogenic effects in mouse embryonic fibroblasts. Eur. J. Pharmacol. 2021, 893, 173804. [Google Scholar] [CrossRef]
- Granda, H.; de Pascual-Teresa, S. Interaction of Polyphenols with Other Food Components as a Means for Their Neurological Health Benefits. J. Agric. Food Chem. 2018, 66, 8224–8230. [Google Scholar] [CrossRef]
- Wijendran, V.; Hayes, K.C. Dietary n-6 and n-3 fatty acid balance and cardiovascular health. Annu. Rev. Nutr. 2004, 24, 597–615. [Google Scholar] [CrossRef]
- Sadi, A.M.; Toda, T.; Oku, H.; Hokama, S. Dietary effects of corn oil, oleic acid, perilla oil, and evening [corrected] primrose oil on plasma and hepatic lipid level and atherosclerosis in Japanese quail. Exp. Anim. 1996, 45, 55–62. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Hu, Q.; Wu, H.; Xue, Y.; Cai, L.; Fang, M.; Liu, Z.; Yao, P.; Wu, Y.; Gong, Z. Protective role of n6/n3 PUFA supplementation with varying DHA/EPA ratios against atherosclerosis in mice. J. Nutr. Biochem. 2016, 32, 171–180. [Google Scholar] [CrossRef]
- Hussein, J.; El-Naggar, M.; Badawy, E.; El-Laithy, N.; El-Waseef, M.; Hassan, H.; Abdel-Latif, Y. Homocysteine and Asymmetrical Dimethylarginine in Diabetic Rats Treated with Docosahexaenoic Acid-Loaded Zinc Oxide Nanoparticles. Appl. Biochem. Biotechnol. 2020, 191, 1127–1139. [Google Scholar] [CrossRef]
- Hennessy, A.A.; Ross, R.P.; Devery, R.; Stanton, C. The health promoting properties of the conjugated isomers of alpha-linolenic acid. Lipids 2011, 46, 105–119. [Google Scholar] [CrossRef]
- Vaklavas, C.; Blume, S.W.; Grizzle, W.E. Translational Dysregulation in Cancer: Molecular Insights and Potential Clinical Applications in Biomarker Development. Front. Oncol. 2017, 7, 158. [Google Scholar] [CrossRef] [Green Version]
- Laham-Karam, N.; Pinto, G.P.; Poso, A.; Kokkonen, P. Transcription and Translation Inhibitors in Cancer Treatment. Front. Chem. 2020, 8, 276. [Google Scholar] [CrossRef] [Green Version]
- Young, A. Inhibition of glucagon secretion. Adv. Pharmacol. 2005, 52, 151–171. [Google Scholar] [CrossRef]
- Van Poelje, P.D.; Potter, S.C.; Chandramouli, V.C.; Landau, B.R.; Dang, Q.; Erion, M.D. Inhibition of fructose 1,6-bisphosphatase reduces excessive endogenous glucose production and attenuates hyperglycemia in Zucker diabetic fatty rats. Diabetes 2006, 55, 1747–1754. [Google Scholar] [CrossRef] [Green Version]
- Shao, J.; Qiao, L.; Janssen, R.C.; Pagliassotti, M.; Friedman, J.E. Chronic hyperglycemia enhances PEPCK gene expression and hepatocellular glucose production via elevated liver activating protein/liver inhibitory protein ratio. Diabetes 2005, 54, 976–984. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Perdomo, G.; Zhang, T.; Slusher, S.; Lee, S.; Phillips, B.E.; Fan, Y.; Giannoukakis, N.; Gramignoli, R.; Strom, S.; et al. FoxO6 integrates insulin signaling with gluconeogenesis in the liver. Diabetes 2011, 60, 2763–2774. [Google Scholar] [CrossRef] [Green Version]
- Fanelli, C.G.; Porcellati, F.; Rossetti, P.; Bolli, G.B. Glucagon: The effects of its excess and deficiency on insulin action. Nutr. Metab. Cardiovasc. Dis. 2006, 16 (Suppl. S1), S28–S34. [Google Scholar] [CrossRef]
- Dang, Q.; Kasibhatla, S.R.; Reddy, K.R.; Jiang, T.; Reddy, M.R.; Potter, S.C.; Fujitaki, J.M.; van Poelje, P.D.; Huang, J.; Lipscomb, W.N.; et al. Discovery of potent and specific fructose-1,6-bisphosphatase inhibitors and a series of orally-bioavailable phosphoramidase-sensitive prodrugs for the treatment of type 2 diabetes. J. Am. Chem. Soc. 2007, 129, 15491–15502. [Google Scholar] [CrossRef] [PubMed]
- Dunbar, B.S.; Bosire, R.V.; Deckelbaum, R.J. Omega 3 and omega 6 fatty acids in human and animal health: An African perspective. Mol. Cell. Endocrinol. 2014, 398, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Fretts, A.M.; Mozaffarian, D.; Siscovick, D.S.; Sitlani, C.; Psaty, B.M.; Rimm, E.B.; Song, X.; McKnight, B.; Spiegelman, D.; King, I.B.; et al. Plasma phospholipid and dietary alpha-linolenic acid, mortality, CHD and stroke: The Cardiovascular Health Study. Br. J. Nutr. 2014, 112, 1206–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.B.; Nam, Y.A.; Kim, H.S.; Hayes, A.W.; Lee, B.M. alpha-Linolenic acid: Nutraceutical, pharmacological and toxicological evaluation. Food Chem. Toxicol. 2014, 70, 163–178. [Google Scholar] [CrossRef] [PubMed]
- O'Neill, C.M.; Minihane, A.M. The impact of fatty acid desaturase genotype on fatty acid status and cardiovascular health in adults. Proc. Nutr. Soc. 2017, 76, 64–75. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance—A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef]
- Wysoczanski, T.; Sokola-Wysoczanska, E.; Pekala, J.; Lochynski, S.; Czyz, K.; Bodkowski, R.; Herbinger, G.; Patkowska-Sokola, B.; Librowski, T. Omega-3 Fatty Acids and their Role in Central Nervous System—A Review. Curr. Med. Chem. 2016, 23, 816–831. [Google Scholar] [CrossRef]
- DeGiorgio, C.M.; Miller, P.R.; Harper, R.; Gornbein, J.; Schrader, L.; Soss, J.; Meymandi, S. Fish oil (n-3 fatty acids) in drug resistant epilepsy: A randomised placebo-controlled crossover study. J. Neurol. Neurosurg. Psychiatry 2015, 86, 65–70. [Google Scholar] [CrossRef] [Green Version]
- Janssen, C.I.; Kiliaan, A.J. Long-chain polyunsaturated fatty acids (LCPUFA) from genesis to senescence: The influence of LCPUFA on neural development, aging, and neurodegeneration. Prog. Lipid Res. 2014, 53, 1–17. [Google Scholar] [CrossRef]
- Lecomte, M.; Paget, C.; Ruggiero, D.; Wiernsperger, N.; Lagarde, M. Docosahexaenoic acid is a major n-3 polyunsaturated fatty acid in bovine retinal microvessels. J. Neurochem. 1996, 66, 2160–2167. [Google Scholar] [CrossRef]
- Allaire, J.; Vors, C.; Tremblay, A.J.; Marin, J.; Charest, A.; Tchernof, A.; Couture, P.; Lamarche, B. High-Dose DHA Has More Profound Effects on LDL-Related Features Than High-Dose EPA: The ComparED Study. J. Clin. Endocrinol. Metab. 2018, 103, 2909–2917. [Google Scholar] [CrossRef] [Green Version]
- Innes, J.K.; Calder, P.C. The Differential Effects of Eicosapentaenoic Acid and Docosahexaenoic Acid on Cardiometabolic Risk Factors: A Systematic Review. Int. J. Mol. Sci. 2018, 19, 532. [Google Scholar] [CrossRef] [Green Version]
- Bhatt, D.L.; Steg, P.G.; Brinton, E.A.; Jacobson, T.A.; Miller, M.; Tardif, J.C.; Ketchum, S.B.; Doyle, R.T., Jr.; Murphy, S.A.; Soni, P.N.; et al. Rationale and design of REDUCE-IT: Reduction of Cardiovascular Events with Icosapent Ethyl-Intervention Trial. Clin. Cardiol. 2017, 40, 138–148. [Google Scholar] [CrossRef] [Green Version]
- Sherratt, S.C.R.; Juliano, R.A.; Copland, C.; Bhatt, D.L.; Libby, P.; Mason, R.P. EPA and DHA containing phospholipids have contrasting effects on membrane structure. J. Lipid Res. 2021, 62, 100106. [Google Scholar] [CrossRef]
- Yi, X.Y.; Gauthier, K.M.; Cui, L.; Nithipatikom, K.; Falck, J.R.; Campbell, W.B. Metabolism of adrenic acid to vasodilatory 1alpha,1beta-dihomo-epoxyeicosatrienoic acids by bovine coronary arteries. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H2265–H2274. [Google Scholar] [CrossRef]
- Kopf, P.G.; Zhang, D.X.; Gauthier, K.M.; Nithipatikom, K.; Yi, X.Y.; Falck, J.R.; Campbell, W.B. Adrenic acid metabolites as endogenous endothelium-derived and zona glomerulosa-derived hyperpolarizing factors. Hypertension 2010, 55, 547–554. [Google Scholar] [CrossRef] [Green Version]
- Brouwers, H.; Jonasdottir, H.S.; Kuipers, M.E.; Kwekkeboom, J.C.; Auger, J.L.; Gonzalez-Torres, M.; Lopez-Vicario, C.; Claria, J.; Freysdottir, J.; Hardardottir, I.; et al. Anti-Inflammatory and Proresolving Effects of the Omega-6 Polyunsaturated Fatty Acid Adrenic Acid. J. Immunol. 2020, 205, 2840–2849. [Google Scholar] [CrossRef]
- Burelle, Y.; Fillipi, C.; Peronnet, F.; Leverve, X. Mechanisms of increased gluconeogenesis from alanine in rat isolated hepatocytes after endurance training. Am. J. Physiol. Endocrinol. Metab. 2000, 278, E35–E42. [Google Scholar] [CrossRef] [Green Version]
- Leij-Halfwerk, S.; van den Berg, J.W.; Sijens, P.E.; Wilson, J.H.; Oudkerk, M.; Dagnelie, P.C. Altered hepatic gluconeogenesis during L-alanine infusion in weight-losing lung cancer patients as observed by phosphorus magnetic resonance spectroscopy and turnover measurements. Cancer Res. 2000, 60, 618–623. [Google Scholar]
- Perez-Sala, D.; Parrilla, R.; Ayuso, M.S. Key role of L-alanine in the control of hepatic protein synthesis. Biochem. J. 1987, 241, 491–498. [Google Scholar] [CrossRef]
- Leij-Halfwerk, S.; Dagnelie, P.C.; van Den Berg, J.W.; Wattimena, J.D.; Hordijk-Luijk, C.H.; Wilson, J.P. Weight loss and elevated gluconeogenesis from alanine in lung cancer patients. Am. J. Clin. Nutr. 2000, 71, 583–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connell, T.M. The complex role of branched chain amino acids in diabetes and cancer. Metabolites 2013, 3, 931–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martino, M.R.; Gutierrez-Aguilar, M.; Yiew, N.K.H.; Lutkewitte, A.J.; Singer, J.M.; McCommis, K.S.; Ferguson, D.; Liss, K.H.H.; Yoshino, J.; Renkemeyer, M.K.; et al. Silencing alanine transaminase 2 in diabetic liver attenuates hyperglycemia by reducing gluconeogenesis from amino acids. Cell Rep. 2022, 39, 110733. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, S.; Fanburg, B.L. Pyrimidine nucleotide synthesis in the normal and hypertrophying rat heart. Relative importance of the de novo and "salvage" pathways. Circ. Res. 1970, 27, 415–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.H.; Larson, B.L. Pyrimidine synthesis pathway enzymes and orotic acid in bovine mammary tissue. J. Dairy Sci. 1971, 54, 842–846. [Google Scholar] [CrossRef]
- Iida, S. Overview: A New Era of Cancer Genomics in Lymphoid Malignancies. Oncology 2015, 89 (Suppl. S1), 4–6. [Google Scholar] [CrossRef]
- Wang, W.; Cui, J.; Ma, H.; Lu, W.; Huang, J. Targeting Pyrimidine Metabolism in the Era of Precision Cancer Medicine. Front. Oncol. 2021, 11, 684961. [Google Scholar] [CrossRef]
- Mollick, T.; Lain, S. Modulating pyrimidine ribonucleotide levels for the treatment of cancer. Cancer Metab. 2020, 8, 12. [Google Scholar] [CrossRef]
- Koene, R.J.; Prizment, A.E.; Blaes, A.; Konety, S.H. Shared Risk Factors in Cardiovascular Disease and Cancer. Circulation 2016, 133, 1104–1114. [Google Scholar] [CrossRef] [Green Version]
- Johnson, C.B.; Davis, M.K.; Law, A.; Sulpher, J. Shared Risk Factors for Cardiovascular Disease and Cancer: Implications for Preventive Health and Clinical Care in Oncology Patients. Can. J. Cardiol. 2016, 32, 900–907. [Google Scholar] [CrossRef]
- Penson, P.; Long, D.L.; Howard, G.; Howard, V.J.; Jones, S.R.; Martin, S.S.; Mikhailidis, D.P.; Muntner, P.; Rizzo, M.; Rader, D.J.; et al. Associations between cardiovascular disease, cancer, and very low high-density lipoprotein cholesterol in the REasons for Geographical and Racial Differences in Stroke (REGARDS) study. Cardiovasc. Res. 2019, 115, 204–212. [Google Scholar] [CrossRef]
- Feig, D.I.; Kang, D.H.; Johnson, R.J. Uric acid and cardiovascular risk. N. Engl. J. Med. 2008, 359, 1811–1821. [Google Scholar] [CrossRef]
- Taghizadeh, N.; Vonk, J.M.; Boezen, H.M. Serum uric acid levels and cancer mortality risk among males in a large general population-based cohort study. Cancer Causes Control. 2014, 25, 1075–1080. [Google Scholar] [CrossRef] [Green Version]
- Yourman, L.C.; Cenzer, I.S.; Boscardin, W.J.; Nguyen, B.T.; Smith, A.K.; Schonberg, M.A.; Schoenborn, N.L.; Widera, E.W.; Orkaby, A.; Rodriguez, A.; et al. Evaluation of Time to Benefit of Statins for the Primary Prevention of Cardiovascular Events in Adults Aged 50 to 75 Years: A Meta-analysis. JAMA Intern. Med. 2021, 181, 179–185. [Google Scholar] [CrossRef]
- Cheung, B.M.; Lauder, I.J.; Lau, C.P.; Kumana, C.R. Meta-analysis of large randomized controlled trials to evaluate the impact of statins on cardiovascular outcomes. Br. J. Clin. Pharmacol. 2004, 57, 640–651. [Google Scholar] [CrossRef] [Green Version]
- Samuel, M.; Tardif, J.C.; Khairy, P.; Roubille, F.; Waters, D.D.; Gregoire, J.C.; Pinto, F.J.; Maggioni, A.P.; Diaz, R.; Berry, C.; et al. Cost-effectiveness of low-dose colchicine after myocardial infarction in the Colchicine Cardiovascular Outcomes Trial (COLCOT). Eur. Heart J. Qual. Care Clin. Outcomes 2021, 7, 486–495. [Google Scholar] [CrossRef]
- Hennessy, T.; Soh, L.; Bowman, M.; Kurup, R.; Schultz, C.; Patel, S.; Hillis, G.S. The Low Dose Colchicine after Myocardial Infarction (LoDoCo-MI) study: A pilot randomized placebo controlled trial of colchicine following acute myocardial infarction. Am. Heart J. 2019, 215, 62–69. [Google Scholar] [CrossRef]
- Deng, S.; West, B.J.; Jensen, C.J. UPLC-TOF-MS Characterization and Identification of Bioactive Iridoids in Cornus mas Fruit. J. Ana.l Methods Chem. 2013, 2013, 710972. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.X.; Cong, W.; Chen, X.Q.; He, S.Y.; Elsheikha, H.M.; Zhu, X.Q. Serum Metabolic Profiling of Oocyst-Induced Toxoplasma gondii Acute and Chronic Infections in Mice Using Mass-Spectrometry. Front. Microbiol. 2017, 8, 2612. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Mei, B.; Chen, D.; Cai, L. GC-MS metabolomics identifies novel biomarkers to distinguish tuberculosis pleural effusion from malignant pleural effusion. J. Clin. Lab. Anal. 2021, 35, e23706. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cochran, C.; Martin, K.; Rafferty, D.; Choi, J.; Leontyev, A.; Shetty, A.; Kurup, S.; Puthanveetil, P. Untargeted Metabolomics Based Prediction of Therapeutic Potential for Apigenin and Chrysin. Int. J. Mol. Sci. 2023, 24, 4066. https://doi.org/10.3390/ijms24044066
Cochran C, Martin K, Rafferty D, Choi J, Leontyev A, Shetty A, Kurup S, Puthanveetil P. Untargeted Metabolomics Based Prediction of Therapeutic Potential for Apigenin and Chrysin. International Journal of Molecular Sciences. 2023; 24(4):4066. https://doi.org/10.3390/ijms24044066
Chicago/Turabian StyleCochran, Cole, Katelyn Martin, Daniel Rafferty, Jennifer Choi, Angela Leontyev, Akanksh Shetty, Sonali Kurup, and Prasanth Puthanveetil. 2023. "Untargeted Metabolomics Based Prediction of Therapeutic Potential for Apigenin and Chrysin" International Journal of Molecular Sciences 24, no. 4: 4066. https://doi.org/10.3390/ijms24044066